

Abstract
Phosphofructokinase (PFK), a regulatory enzyme in glycolytic pathway, has been purified to electrophoretic homogeneity from adult female Setaria cervi and partially characterized. For this enzyme, the Lineweaver-Burk's double reciprocal plots of initial rates and D-fructose-6-phosphate (F-6-P) or Mg-ATP concentrations for varying values of cosubstrate concentration gave intersecting lines indicating that K m values for F-6-P (1.05 mM) and ATP (3 μM) were independent of each other. S. cervi PFK, when assayed at inhibitory concentration of ATP (>0.1 mM), exhibited sigmoidal behavior towards binding with F-6-P with a Hill coefficient (n) value equal to 1.8 and 1.7 at 1.0 and 0.33 mM ATP, respectively. D-fructose-1,6-diphosphate (FDP) competitively inhibited the filarial enzyme: K i and Hill coefficient values being 0.18 μM and 2.0, respectively. Phosphoenolpyruvate (PEP) also inhibited the enzyme competitively with the K i value equal to 0.8 mM. The Hill coefficient values (>1.5) for F-6-P (at inhibitory concentration of ATP) and FDP suggested its positive cooperative kinetics towards F-6-P and FDP, showing presence of more than one binding sites for these molecules in enzyme protein and allosteric nature of the filarial enzyme. The product inhibition studies gave us the only compatible mechanism of random addition process with a probable orientation of substrates and products on the enzyme surface.
1. Introduction
Phosphofructokinase (ATP: D-fructose-6-phosphotrans-ferase, EC 2.7.1.11, PFK) is an enzyme of prime importance in the regulation of glycolytic flux in both the mammalian [1–5] and parasites [6–8], for example nematodes such as filarial worms Setaria cervi [9–13] and Dirofilaria immitis [14], Ascaris suum [15], malaria parasite Plasmodium berghei [16], intestinal parasite Entamoeba histolytica [17], liver fluke Fasciola hepatica [18], and a few others including Toxoplasma gondii [19], Trypanosoma cruzi [20], and Trypanosoma brucei [21]. The filarial worms studied so far employ predominantly anaerobic metabolism of carbohydrate (glycogen and/glucose) as a major source of energy [22]. However, they do not catalyze the complete oxidation of the substrate to CO2 and reduced organic acids as end product of the metabolism. The filarial nematodes utilize limited quantity of oxygen, when available and possess rudimentary and unusual electron transport chains that catalyze limited terminal oxidation with generation of little energy [23].
The kinetic and regulatory properties of the purified enzyme have been extensively studied in several mammalian systems, plants, and bacteria [24, 25]. In most of the cases, it is shown that PFK regulates glucose metabolism. Because of the multiplicity of modifiers, PFK has served as a model in studies of allosteric regulation of enzymes. The enzyme activity appears to be modulated to meet the metabolic needs of the cell, with the metabolites serving as intracellular indicators [26]. However, very little attention has been paid towards the systematic studies on the nature and properties of this enzyme from the filarial parasites.
Setaria cervi, a bovine filarial parasite, dwelling in the intraperitoneal cavity and lymphatics, possesses similarities with the human filarial worms in many ways such as nocturnal periodicity, antigenic composition, and the metabolic pathways. This parasite relies mainly on glycolysis for deriving energy [27]. It contains comparatively low levels of PFK and is thus highly vulnerable to the inhibitors of the glycolytic enzymes. PFK from S. cervi has been purified to electrophoretic homogeneity and some of its molecular properties have been studied [22]. It has been shown that this enzyme possess two different pH optima depending on ATP concentrations, the values being 8.0 at low (0.1 mM) concentration which decreases to pH 7.4 at high ATP (>0.1 mM) concentration. These results indicated that the activity of filarial PFK was possibly under regulation of ATP levels [28].
The present paper describes certain kinetic properties of PFK purified from adult female filarial worm, S. cervi, at low and high concentrations of ATP. It is shown that the molecular characteristics and reaction mechanisms of filarial PFK as deduced using steady state kinetic method differ radically from those of the complex vertebrates, thereby suggesting the adaptation of the filarial parasite in the anaerobic habitat.
2. Materials and Methods
2.1. Materials
2.1.1. Parasite
Motile adult female worms were collected from the peritoneal folds of freshly slaughtered buffaloes at a local abattoir and brought to the laboratory in glucose saline. Worms were thoroughly washed three to four times with lukewarm isotonic saline. The fresh or the frozen worms at −20°C for a week were utilized for this study.
2.1.2. Chemicals/Biochemicals
D-fructose-6-phosphate (F-6-P), adenosine-3′, 5′-triphosphate (ATP), α-glycerophosphate dehydrogenase (GDH), D-fructose-1,6-diphosphate (FDP), triosephosphate isomerase (TPI), aldolase, and phosphoenolpyruvate (PEP) were purchased from Sigma Chemical Co., USA. Nicotinamide adenine dinucleotide reduced (NADH) was obtained from CSIR Centre of Biochemical Technology, New Delhi. Other reagents used were analytical grade.
2.2. Methods
2.2.1. Purification of S. cervi PFK
PFK from adult female S. cervi has been purified to electrophoretic homogeneity using very simple procedures and the activity has been stabilized using suitable reagents [22]. This purified enzyme preparation was used in the present study.
2.2.2. Enzyme Assay
S. cervi PFK was assayed using an enzyme coupled reaction method described by Racker (1947) [29] with slight modification as described in [22]. In this method, we measured the formation of the D-Fructose-1,6-diphosphate (FDP) using aldolase, TPI, GDH, and NADH. The reaction mixture (3 mL) contained Tris HCl buffer (50 mM, pH 8.0), F-6-P (3.3 mM), ATP (0.1 mM), MgCl2 (3.3 mM), NADH (0.04 mM), GDH (0.66 Units/mL), TPI (5.6 Units/mL), aldolase (0.21 Units/mL), and suitable amount of enzyme protein (10–20 μg). The reaction was always started by adding substrate to the reaction mixture and the change in absorbance (oxidation of NADH to NAD+) after every 30 sec interval was measured spectrophotometrically at 340 nm. The three auxiliary enzymes such as aldolase, GDH, and TPI were added in excess so that the overall reaction was governed by PFK activity present in the assay mixture. The concentration of Mg2+ was kept higher than that of ATP (unless stated otherwise) for generating Mg-ATP complex (the substrate for the enzyme) and avoiding presence of free ATP molecules, which are known to be inhibitory in nature to PFK from other sources [30].
2.2.3. Determination of K m and K cat Values for Substrates of S. cervi PFK
The mechanism of enzyme action was investigated using steady state kinetic methods [31] and it was measured in the direction of formation of FDP (from F-6-P, ATP, Mg2+ ion) in the presence of NADH and nonratelimiting concentrations of auxiliary enzymes (aldolase, TPI and GDH). The K m and K cat values of F-6-P and ATP were determined by studying the rate of PFK catalyzed reaction at different concentrations of one substrate keeping the concentration of the second substrate constant. The K cat values were based on a subunit mass of 90 kDa [22].
2.2.4. Determination of K m Value of MgCl2 for S. cervi PFK
The S. cervi PFK was assayed at varying concentrations of MgCl2 (0.05–7mM) keeping concentrations of ATP (0.1 mM) and F-6-P (3.3 mM) constant. The K m value of MgCl2 was computed from equation deduced from Lineweaver-Burk's double reciprocal plot.
2.2.5. Effect of Divalent Cations
To determine the effect of divalent metal ions such as Mg2+, Ca2+, Mn2+, Co2+, Cd2+, and Ba2+ on S. cervi PFK catalyzed reaction, assays were carried out under standard conditions (0.1 mM ATP, 3.3 mM F-6-P). The metal ion concentration in each case was 3.3 mM. The rate observed at 3.3 mM MgCl2 has been taken to be 100%.
2.2.6. Product Inhibition Studies
The inhibition of S. cervi PFK by fructose-1,6-diphosphate (FDP) and phosphoenol pyruvate (PEP) was determined by assaying the enzyme at varying concentrations of F-6-P (0.5–5.0 mM) in presence and absence of fixed concentrations of FDP or PEP keeping the concentrations of ATP (0.1 mM) and Mg2+ (3.3 mM) constant. While studying the effect of FDP on activity of filarial PFK, suitable control was used (employing the reaction mixture containing FDP in the presence and absence of enzyme protein), which was deduced from experimental one to record the actual effect of FDP on activity of filarial PFK. Inhibition constants were calculated using the equation deduced from Lineweaver-Burk's double reciprocal plot.
2.2.7. Other Analytical Methods
The mechanism of enzyme action was investigated using steady state kinetic methods and it was measured in the direction of formation of FDP (from F-6-P, ATP, Mg2+ ion) in the presence of NADH and nonratelimiting concentrations of auxiliary enzymes (aldolase, TPI and GDH). K m values of F-6-P and ATP were determined by studying the rate of PFK catalyzed reaction at different concentrations of one substrate keeping the concentration of the second substrate constant.
3. Results
3.1. Effect of D-Fructose-6-Phosphate and ATP on the Phosphofructokinase Activity
In order to study the effect of F-6-P, the enzyme was assayed using varying concentrations of F-6-P at two different concentrations of ATP (50 and 100 μM). Similarly, the effect of ATP was studied by assaying the enzyme activity using varying concentrations of ATP and two fixed concentrations of F-6-P (1.7 and 3.3 mM). The K m and Vmax values were computed from the Lineweaver-Burk's double reciprocal plots. The results are shown in Figures 1 and 2. In each case the enzyme exhibited hyperbolic saturation curves. The double reciprocal plots as shown in Figures 1(b) and 2(b) gave sets of straight lines intersecting at common points on the negative abscissa from which the K m values of 1.05 mm and 3 μM were obtained for F-6-P and Mg-ATP2−, respectively. These observations further suggested that the K m values of Mg-ATP2− and F-6-P were not affected by varying the concentrations of other reactant (noncompetitive behavior). This result is consistent with the random order of attachment of the substrates to the enzyme. The low K m value of for ATP (3 μM) is noteworthy. Further the Mg2+ was always quite high (3.3 mM), so that all ATP must have existed as Mg-ATP complex. Thus the results suggested that S. cervi PFK had high affinity for Mg-ATP complex. However, the S. cervi enzyme was found to be highly specific for F-6-P: at 3.3 mM the relative activities with D-glucose-6-phosphate and D-glucose-1-phosphate were 10.7 and 3.2%, respectively, of the rate with F-6-P. The K cat value was found to be 320 ± 15 sec−1.
Figure 1.
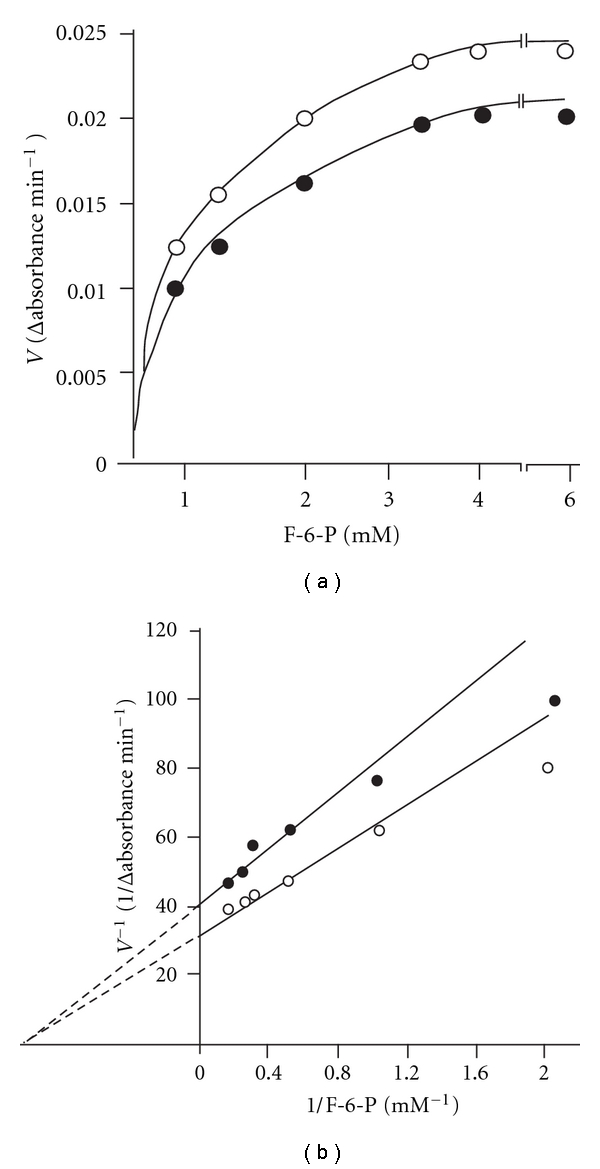
Effect of F-6-P concentration on the rate of S. cervi PFK catalyzed reaction. ATP concentration was 50 (○) and 100 (•) μM and Mg2+ concentration was 3.3 mM. Enzyme concentration was 6.6 μg/mL. Other conditions were same as in standard enzyme assay. (b) Double reciprocal plot of the data of (a). ATP concentrations were 50 (○) and 100 (•) μM.
Figure 2.
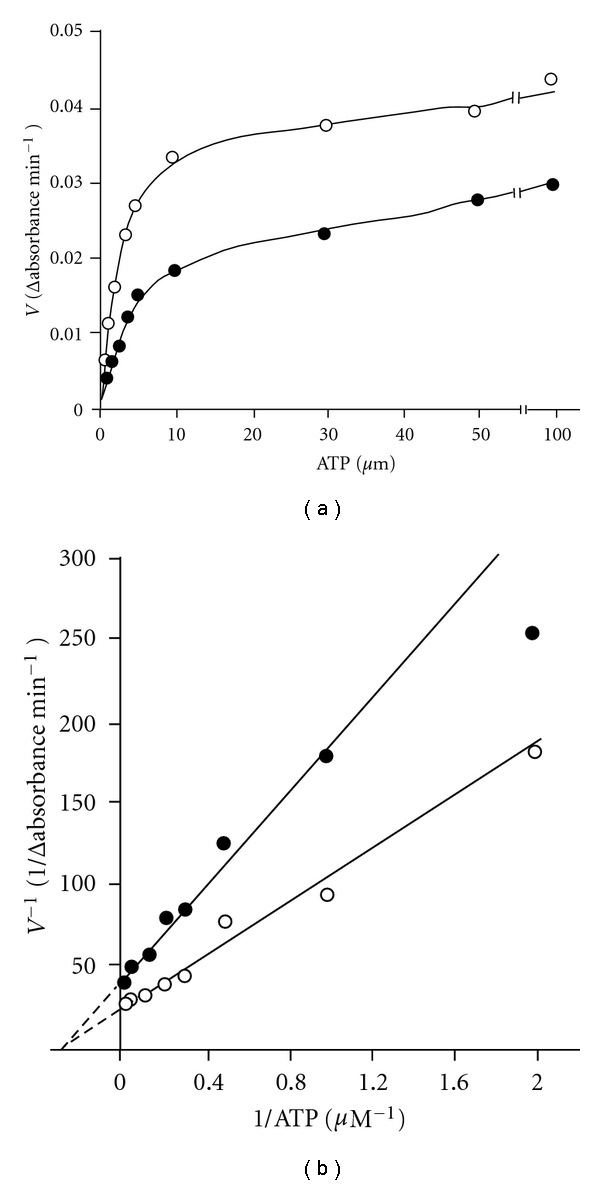
Effect of ATP concentration on the rate of PFK catalyzed reaction. F-6-P concentration was 1.6 (○) and 3.3 (•) mM and Mg2+ concentration was 3.3 mM. Enzyme concentration was 10 μg/mL. Other conditions were same as in standard enzyme assay. (b) Double reciprocal plot of the data of (a). F-6-P concentrations were 1.6 (Δ) and 3.3 (▲) mM.
3.2. Effect of Higher Concentrations of ATP on Filarial PFK Catalyzed Reaction
At higher concentration of ATP (330 and 1000 μM) and 3.3 mM Mg2+ concentration, the reaction rate versus F-6-P concentration curve was found to be sigmoidal in shape (Figure 3). The sigmoidicity in these data appears to be function of ATP concentration. Correspondingly, slope of the Hill plot (n) of the data (Figure 3(b)) was found to depend on ATP concentration. Its values were found to be 1.8, 1.7, and 1.1 at 1.0, 0.33 and 0.1 mM ATP concentrations, respectively.
Figure 3.
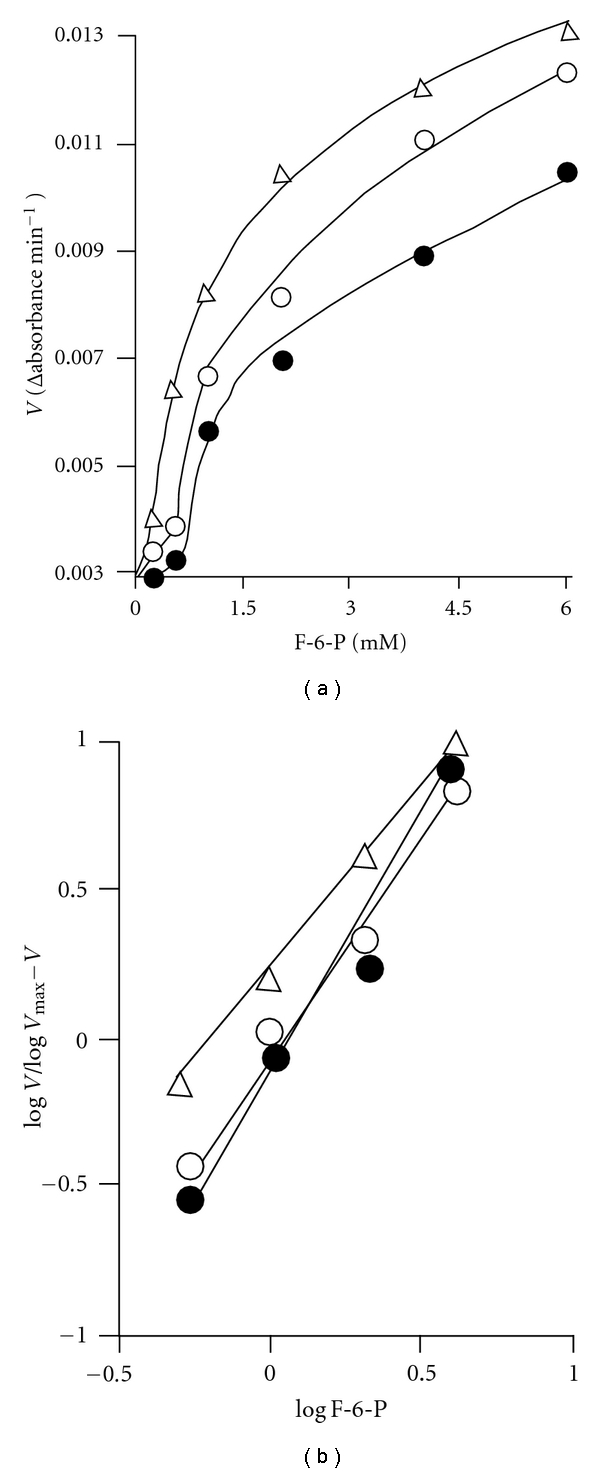
Effect of F-6-P concentration on the rate of S. cervi PFK catalyzed reaction at different fixed and high concentration of ATP. ATP concentration was 0.1 (Δ), 0.33 (○), and 1.0 (•) mM and Mg2+ concentration was 3.3 mM. Enzyme concentration was 3.3 μg/mL. Other conditions were same as in standard enzyme assay. Inset. Hill plot of the data of (a) at 0.2–6.0 mM F-6-P concentrations. ATP concentrations were 0.1 (Δ), 0.33 (○), and 1.0 (•).
It may be noted that concentrations of ATP, which gave rise to sigmoidicity in F-6-P saturation curve in Figure 3 were higher than those used in Figures 1 and 2, where simple hyperbolic saturation curves were obtained for both the substrates. It became necessary, therefore, to extend the experiments for the effect of ATP to higher concentration ranges. Results of such an experiment carried out at a single fixed and high F-6-P concentration (3.3 mM) are shown in Figure 4. The concentration of Mg2+ was held high and constant (3.3 mM). Rate of reaction increased progressively up to 0.10 mM ATP concentration, beyond which a sharp decline was observed suggesting inhibition of the enzyme at higher ATP concentrations. The plots of the data obtained at high and inhibitory ATP concentrations (1/rate of reaction versus ATP) and at two fixed F-6-P concentrations, namely, 1.7 and 3.33 mM (Figure 4; inset) showed that the enzyme had an allosteric site for ATP with low affinity. The substrate inhibition constant (K i s) value was 1.2 ± 0.1 mM.
Figure 4.
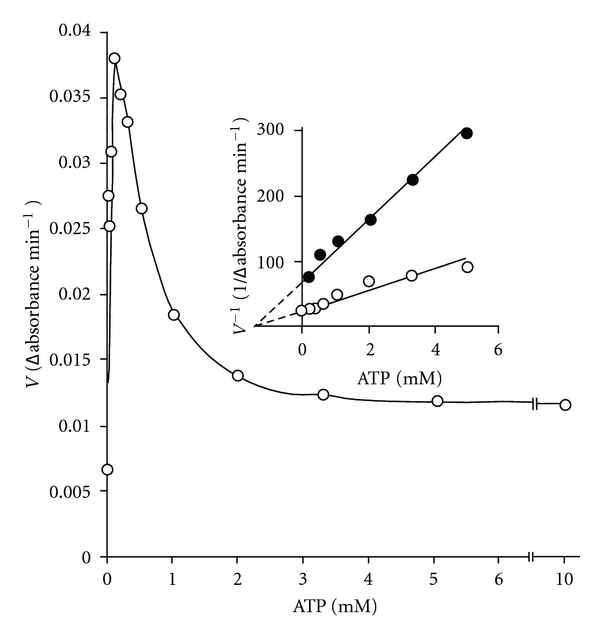
Effect of ATP concentration on the rate of PFK catalyzed reaction at fixed concentration of F-6-P and Mg2+ (3.3 mM each). Inset shows the plot obtained on assaying the enzyme at variable inhibitory (>0.1 mM) concentration of ATP at two fixed concentrations of F-6-P such as 1.6 (•) and 3.3 (○) mM. Mg2+ concentration was constant (3.3 mM). Enzyme concentration was 10 μg/mL. Other conditions were same as in standard enzyme assay.
3.3. Concentration of Mg2+ Ion Is Limiting for Filarial PFK Activity
In a separate set of experiments, concentration of F-6-P was maintained constant at 3.3 mM and concentrations of Mg2+ and ATP were varied together and kept equal to each other. The data showed an apparent sigmoidicity when rate of reaction was plotted against [ATP]. The results indicate that sigmoidicity is observed at low values of Mg2+ and ATP concentrations (Figure 5). Under these conditions, the dissociation of Mg-ATP complex will be significant
Figure 5.
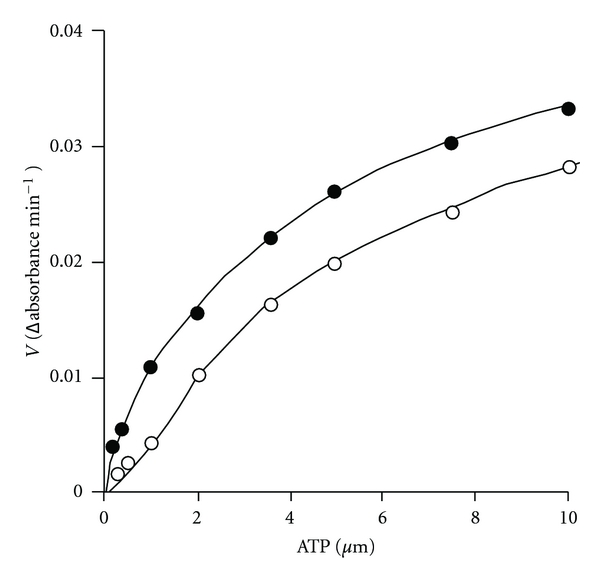
Effect of Mg2+ and ATP concentrations on the rate of PFK-catalyzed reaction. For (○), Mg2+ ion concentration was equal to and varied together with that of ATP. For (•), Mg2+ ion concentration was equal to 3.33 mM. F-6-P concentration in each case was 3.33 mM. Enzyme concentration was 10 μg/mL. Other conditions were same as in standard enzyme assay.
| (1) |
At lower concentrations, the concentration of complex will be less than that of Mg2+and ATP added. If Mg-ATP complex (rather than free ATP and Mg2+) is the real substrate, the observed rate of reaction will be lower than expected. This will make the curve sigmoidal. Correspondingly, when concentration of Mg2+ is kept high and constant (3.3 mM), all available ATP will exist as Mg-ATP complex. Under these conditions rate versus ATP concentration plot is no longer sigmoidal but gives usual hyperbolic curve (Figure 5).
3.4. Presence of Excess Concentration of Cations, Specifically Mg2+ Ions, Is Required by Filarial PFK
The effect of divalent metal ions (Mg2+, Ca2+, Mn2+, Co2+, Cd2+, and Ba2+) on S. cervi PFK catalyzed reaction was studied in order to find out the specificity and preferential utilization of metals for optimal activity. The S. cervi PFK exhibited very low (2%) activity when Mg2+ was omitted from the assay solution (Table 1). The filarial PFK activity exhibits Mg2+ concentration dependence (Figure 6). The K m of Mg2+ was found to be 0.23 ± 0.02 mM. Mn2+, Ca2+ and Co2+ (each at 3.3 mM concentration) showed 36, 11 and 5% of the activity observed with Mg2+. The other metal ions tested, Cd2+ and Ba2+ did not show any activity.
Table 1.
Effect of some divalent metal ions on the activity of PFK from S. cervi.
| Salt | Relative activity |
|---|---|
| MgCl2 | 100 |
| CaCl2 | 36 |
| MnCl2 | 11 |
| CoCl2 | 5 |
| CdCl2 | 0 |
| BaCl2 | 0 |
| No salt | 2 |
Assays were carried out at optimal concentration of F-6-P (3.3 mM) and ATP (0.1 mM) following the procedure as described in Section 2. The metal ion concentration in each case was 3.3 mM. The rate of reaction determined at 3.3 mM MgCl2 is considered to be 100%.
Figure 6.
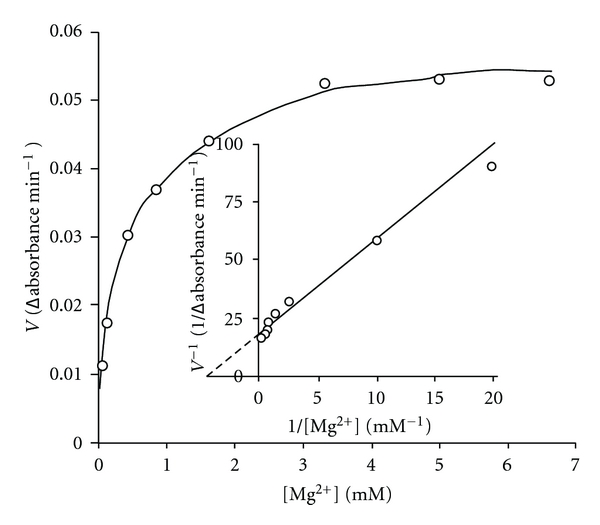
Effect of Mg2+ concentration on the rate of PFK-catalyzed reaction. Concentrations of F-6-P and ATP were 3.33 and 0.1 mM, respectively. Enzyme concentration was 10 μg/mL. Other conditions were same as in standard enzyme assay. Inset shows the double reciprocal plot of the data.
3.5. Effect of Products (D-Fructose-1,6-Diphosphate (FDP)) and Phosphoenolpyruvate (PEP) on the Activity of S. cervi Phosphofructokinase
The product inhibition studies were carried out using F-6-P as a variable substrate and near saturating (0.1 mM) concentration of cosubstrate (Mg-ATP). The results showed that the inhibition of PFK activity by FDP was competitive in nature (Figure 7(a)) with respect to F-6-P; the K i value as computed from the Lineweaver-Burk's double reciprocal plot was found to be 0.18 μM. A Hill plot of the data at several FDP concentrations and a fixed F-6-P concentration (Figure 7(b)) showed a slope equal to 2.0, suggesting cooperativity in the binding of FDP. PEP, another metabolite in glycolysis, also showed a cooperative type of inhibition with respective F-6-P (Figure 8) and its K i value was found to be 0.8 mM.
Figure 7.
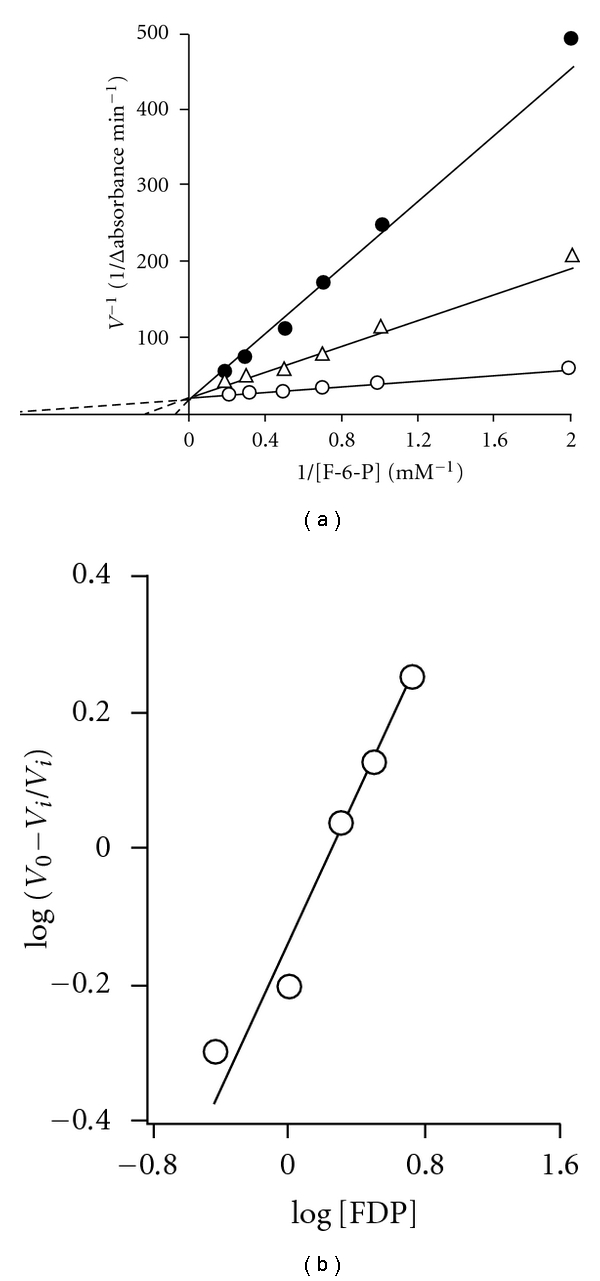
(a) Effect of FDP on the rate of PFK catalyzed reaction. FDP concentrations were nil (○), 1.0 (Δ), and 2.0 (▲) μM. Mg2+ concentration was constant (3.3 mM). The concentration of ATP was 0.1 mM. Enzyme concentration was 10 μg/mL. Other conditions were same as in standard enzyme assay. (b) Hill plot of the data of the effect of FDP on the rate of PFK catalyzed reaction. The concentrations of F-6-P, ATP, and Mg2+ were 3.3, 0.1, and 3.3 mM, respectively. Enzyme concentration was 6.6 μg/mL. V i and V 0 are the rates of reaction in the presence and absence of FDP. Other conditions were same as in standard enzyme assay.
Figure 8.
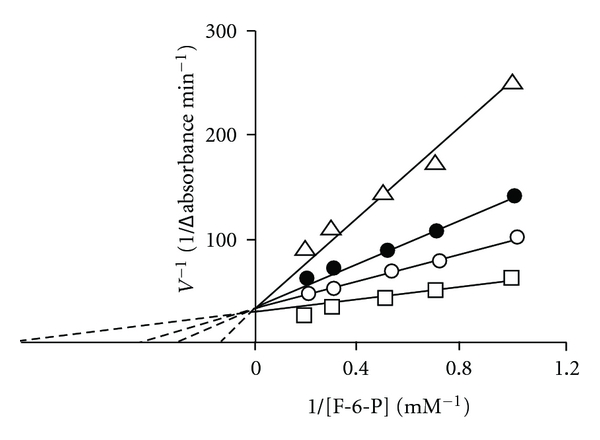
Effect of PEP on the rate of PFK-catalyzed reaction. PEP concentrations were nil (□), 1.0 (○), 2.0 (•), and 5.0 (Δ) mM. Mg2+ and ATP concentrations were 3.3 and 0.1 mM, respectively. Enzyme concentration was 10 μg/mL. Other conditions were same as in standard enzyme assay.
4. Discussion
4.1. The Crude and Purified Preparations of S. cervi PFK Have Same K m for F-6-P
Earlier works from this laboratory have demonstrated the existence of PFK in S. cervi comparatively in lesser amount. This enzyme has been purified and some of its molecular characteristics were studied [22]. The K m of F-6-P for S. cervi PFK and its inhibition by high concentration of ATP with crude preparation [28] have been further confirmed with the purified enzyme [22]. This agreement suggests that no kinetically relevant proteolytic modifications occurred during purification.
4.2. S. cervi PFK Exhibits Specificity in Having More Affinity to Mg-ATP Complex Than F-6-P
The much lower K m value of Mg-ATP (3 μM) as compared to that of F-6-P (1.05 mM) for S. cervi enzyme at pH 8.0, suggests higher affinity of the enzyme to ATP than F-6-P. Similar wide differences in K m values of Mg-ATP and F-6-P were also observed with the enzyme isolated from other parasites such as M. expansa [32], A. suum [15] T. brucei [21], and T. cruzi [20] whereas the enzyme from vertebrate tissues exhibited very less difference between the K m values of Mg-ATP and F-6-P [33]. The binding of the substrate Mg-ATP to PFKs from different origins shows a hyperbolic dependence with half saturation values in the range of 10–100 μM in most of the cases. However, it was quite low (3 μM) for PFK from S. cervi.
4.3. The Affinity of S. cervi PFK for One Substrate Is Independent of the Concentration of the Cosubstrate
The complex allosteric interactions of substrate and effectors with PFK hampered the study of the reaction mechanism of this enzyme from most of the sources. The study of the reaction mechanism of S. cervi PFK was carried out under optimal conditions using steady state kinetic methods [34, 35]. The intersecting lines of Lineweaver-Burk's double reciprocal plots (Figures 1 and 2) clearly indicated that the affinity of the enzyme for one substrate was independent of the concentration of the cosubstrate. Similar results were reported for the enzyme of T. cruzi [20] However, for the enzyme in M. expansa and T. brucei [36], the K m value of Mg-ATP2- has been shown to increase at higher F-6-P concentration. The K cat value (320 ± 15 sec−1) for F-6-P as recorded for S. cervi PFK in present investigation was found to be similar to that from E. histolytica (344 sec−1) [37] and about five times higher than that of T. brucei (61 sec−1) [38].
4.4. ATP Regulates Activity of S. cervi PFK
The purified enzyme from S. cervi displayed regulatory properties which confirmed previous reports from this laboratory with a cytosolic preparation particularly its inhibition by higher concentration of ATP (>0.1 mM), which is the basic regulatory property of PFK in vertebrates, bacteria, and a few plants responsible for Pasteur effect [12]. The steady state kinetic results, as stated above, were obtained when ATP concentration was equal to or less than 0.1 mM. The rate of reaction increased to maximum with ATP concentration up to 0.1 mM (in presence of 3.3 mM F-6-P and 3.3 mM Mg2+) (Figure 4). At higher ATP concentration, a lowering of catalyzed rate was observed (inhibition by excess ATP). The enzyme from other mammalian [39] and parasite sources [18] required more ATP for optimal activity. The corresponding optimum ATP concentrations for the monkey liver [40] and rat small intestine enzymes [41] were found to be 0.4 and 1.0 mM, respectively. The enzymes from A. suum [15] and F. hepatica [42] require 50–200 and 200 μM ATP concentrations, respectively, for showing maximal activity.
The kinetics of ATP and F-6-P saturation for S. cervi enzyme are almost identical to those reported in D. immitis and A. suum [43]. The inhibitory action of ATP was reported for the first time with rabbit muscle PFK and the inhibition was attributed to isosteric competition with the true substrate, that is, Mg-ATP complex. However, with the increase in F-6-P concentration, the inhibitory action of ATP was reduced suggesting that the ability to demonstrate the regulatory effects of either substrate is a function of relative concentration of other substrate.
4.5. Allosteric Regulation of S. cervi PFK
The inhibition of rabbit muscle PFK by ATP has been reported to be pH dependent [44–46]. In contrast to these findings, the enzyme from T. cruzi was not inhibited by ATP even at 1.0 mM concentration at pH 7.4. The inhibition pattern (noncompetitive behavior) of S. cervi PFK by ATP is in close agreement with that of pig spleen enzyme. The filarial PFK showed sigmoidicity in saturation with respect to F-6-P when assayed at inhibitory concentration of ATP with Hill coefficient (n) values found to be 1.8. The complex kinetics of pure S. cervi enzyme towards F-6-P (Figure 3) has also been reported for analogous enzyme isolated from mammalian and parasite sources. It has been explained assuming more than two binding sites per catalytic unit with cooperative interactions, by the presence of two or more isozymes with different kinetic properties or by changes in the aggregation state and the activity of the enzyme induced by the combination of protein and substrate (particularly F-6-P) concentrations.
4.6. Mg-ATP Complex Is the Real Substrate of S. cervi PFK
In all the experiments described above, Mg2+ concentration was maintained high and constant (3.3 mM). If Mg2+ and ATP concentrations are maintained equal and varied together keeping F-6-P concentration high and constant (3.3 mM), some deviation from the hyperbolic saturation is observed. The deviation is observed when both Mg2+ and ATP concentrations are low (2 μM). The rate versus (Mg2+) = (ATP) curve has a sigmoidal shape. At low Mg2+ and ATP concentrations, dissociation of Mg-ATP complex into free Mg2+ and ATP will be considerable and significant. At higher concentrations, on the other hand, Mg2+ and ATP will exist mostly in the form of Mg-ATP complex. If the latter (rather than free ATP) is the real substrate for enzyme, its concentration will decrease sharply (and disproportionately) at lower concentrations. Consequently, lower rates of reaction will be obtained than those calculated if all ATP + Mg2+ existed as Mg-ATP complex. This will generate an apparent sigmoidicity without there being any cooperativity in binding of Mg-ATP to enzyme. Further, free ATP is expected to bind better to the allosteric or inhibitory site of enzyme. This will enhance the sigmoidicity. Thus our data suggest that Mg-ATP is the real substrate of S. cervi PFK.
The complex kinetics of pure S. cervi enzyme towards F-6-P (Figure 3) has also been reported for analogous enzyme isolated from many other sources. It has been explained assuming more than two binding sites per catalytic unit with cooperative interactions, by changes in activity of the enzyme induced by the combination of protein and substrate (particularly F-6-P) concentrations.
In summary, three types of steady state kinetic results have been described. In one set of experiments, noninhibitory concentrations of ATP (≤0.1 mM) and high Mg2+ concentration (3.3 mM) were used and hyperbolic saturation was observed for F-6-P as well as ATP. Their K m values (1.05 mM and 3 μM, respectively) were independent of the concentration of other substrate. In second set, when inhibitory concentrations of ATP (1.0 mM or above) were employed at high Mg2+ concentration (3.3 mM), F-6-P binding shows sigmoidicity with a Hill coefficient (n) equal to 1.8. In third set, non-inhibitory concentrations of ATP were employed, but Mg2+ and ATP concentrations were held equal and varied together. Concentration of F-6-P was held constant at 3.3 mM. An apparent sigmoidicity was observed in the plot of rate versus ATP concentration. This is explained on the basis of assumption that Mg-ATP complex, rather than free ATP is the real substrate for this enzyme.
4.7. S. cervi PFK Preferentially Utilizes Mg2+ for Its Optimal Activity
The results of the effect of some divalent cations on activity of S. cervi PFK suggested that this enzyme possess a wide specificity for metal ions like analogous enzyme from mammalian sources. However, unlike other systems, filarial PFK exhibits preferential utilization of Mg2+ for its optimal activity. The K m of Mg2+ for S. cervi PFK (0.23 ± 0.02 mM) is nearly five times lower than the value reported for rabbit brain (1.2 mM), and very close to value for T. brucei (0.294 mM) [21].
4.8. Feedback Inhibition of S. cervi PFK
The product inhibition studies were carried out using F-6-P as a variable substrate (because of its simple Michaelis-Menten kinetics at all concentrations) and near saturating (0.1 mM) concentration of Mg-ATP. S. cervi PFK was found to be inhibited by FDP and PEP in a competitive manner, the former was inhibitory at micromolar, while the latter was effective at millimolar concentrations. The enzymes from T. brucei and T. cruzi were also inhibited by FDP, while that from A. suum and epimastigotes of T. cruzi remained unaffected on treatment with FDP. The enzyme from mammalian sources, however, was significantly activated by FDP. PEP is well-known inhibitor of PFK from mammalian systems and parasites. It has been reported that PEP acts as an allosteric inhibitor of PFK from T. brucei. In present investigation, the K i value of PEP for S. cervi PFK was half of the value reported for the pig spleen enzyme. PEP, however, has been shown to activate the enzyme from malaria parasite [16].
The product inhibition patterns displayed by S. cervi PFK (Figures 5 and 6) are only compatible with random addition process with the probable orientation of the substrate and product on the enzyme surface in which random entry of substrates produces a central complex which isomerizes and releases the products in random fashion. Again the proposed mechanism for S. cervi PFK is markedly different from those reported for the analogous enzyme from many other sources: the results of the studies on the mechanism of mammalian PFK has been shown to exhibit both random and ordered sequential bi-bi process [39]; bacterial enzyme possesses an ordered bi-bi mechanism [24], while the enzyme from the yeast and slime mold favors an apparent ping-pong bi-bi. On the other hand, the proposed reaction mechanism of random order of the entry of the substrates to the active site of S. cervi enzyme has been reported mainly for allosteric PFK, which is much different from those reported for nonallosteric analogous enzyme from certain other sources [47, 48].
5. Conclusion
In conclusion, the filarial enzyme shows allosteric properties similar to PFK from various sources; it is inhibited by ATP and exhibits two different pH optima depending on the ATP concentrations but it differs with allosteric regulation. The S. cervi follows a random mechanism of substrate binding. The filarial enzyme appears to be parasite specific which differs in molecular and kinetic properties from that of the mammalian sources. These differences may be attributed with an adaptation of this parasite to an anaerobic utilization of glucose. Further, the unique properties of S. cervi enzyme could be exploited for an effective and parasite-specific antifilarial drug design.
Acknowledgments
The author is grateful to Professor O. P. Malhotra, BHU-Varanasi and Dr. S. N. Ghatak, Lucknow for their valuable inputs. The financial assistance to B. Sarma in the form of a fellowship from CSIR-New Delhi is acknowledged. B. Sarma thanks Mr. Ratnesh K.Sharma for drawing the figures.
References
- 1.Jenkins CM, Yang J, Sims HF, Gross RW. Reversible high affinity inhibition of phosphofructokinase-1 by Acyl-CoA: a mechanism integrating glycolytic flux with lipid metabolism. Journal of Biological Chemistry. 2011;286(14):11937–11950. doi: 10.1074/jbc.M110.203661. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Li BW, Rush AC, Jiang DJ, Mitreva M, Abubucker S, Weil GJ. Gender-associated genes in filarial nematodes are important for reproduction and potential intervention targets. PLoS Neglected Tropical Diseases. 2011;5(1) doi: 10.1371/journal.pntd.0000947. Article ID e947. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Langer S, Kaminski MT, Lenzen S, Baltrusch S. Endogenous activation of glucokinase by 6-phosphofructo-2-kinase/fructose- 2,6-bisphosphatase is glucose dependent. Molecular Endocrinology. 2010;24(10):1988–1997. doi: 10.1210/me.2010-0115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.MacDonald JA, Storey KB. Reassessment of the cold-labile nature of phosphofructokinase from a hibernating ground squirrel. Molecular and Cellular Biochemistry. 2001;225(1-2):51–57. doi: 10.1023/a:1012264723657. [DOI] [PubMed] [Google Scholar]
- 5.Usenik A, Legiša M. Evolution of allosteric citrate binding sites on 6- phosphofructo-1-kinase. PLoS One. 2010;5(11) doi: 10.1371/journal.pone.0015447. Article ID e15447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Ahmad R, Srivastava AK. Biochemical composition and metabolic pathways of filarial worms, Setaria cervi: search for new antifilarial agents. Journal of Helminthology. 2007;81(3):261–280. doi: 10.1017/S0022149X07799133. [DOI] [PubMed] [Google Scholar]
- 7.Barrett J. Forty years of helminth biochemistry. Parasitology. 2009;136(12):1633–1642. doi: 10.1017/S003118200900568X. [DOI] [PubMed] [Google Scholar]
- 8.Bryant C, Behm CA, Michael J. Howell Biochemical Adaptation in Parasites. London, UK: Chapman and Hall; 1989. [Google Scholar]
- 9.Barrett J. Biochemistry of filarial worms. Helminthology Abstracts Series A. 1983;52:1–18. [Google Scholar]
- 10.Gupta S, Srivastava AK. Biochemical targets in filarial worms for selective antifilarial drug design. Acta Parasitologica. 2005;50(1):1–18. [Google Scholar]
- 11.Ogushi S, Lawson JW, Dobson GP, Veech RL, Uyeda K. A new transient activator of phosphofructokinase during initiation of rapid glycolysis in brain. Journal of Biological Chemistry. 1990;265(19):10943–10949. [PubMed] [Google Scholar]
- 12.Rafi MM, Raj RK. Phosphoenolpyruvate-succinate-glyoxylate pathway in the filarial parasite Setaria digitata. Journal of Biosciences. 1991;16(3):121–126. [Google Scholar]
- 13.Uchida Y, Koyama T, Hachimori A. Stability and conformation of porcine phosphofructokinase M and L. Comparative Biochemistry and Physiology. 1990;96(2):399–404. doi: 10.1016/0305-0491(90)90395-a. [DOI] [PubMed] [Google Scholar]
- 14.Srinivasan NG, Rao GSJ, Harris BG. Phosphofructokinase from Dirofilaria immitis: effect of fructose 2,6-bisphosphate and AMP on the non-phosphorylated and phosphorylated forms of the enzyme. Molecular and Biochemical Parasitology. 1990;38(1):151–158. doi: 10.1016/0166-6851(90)90215-8. [DOI] [PubMed] [Google Scholar]
- 15.Hofer HW, Allen BL, Kaeini MR, Pette D, Harris BG. Phosphofructokinase from Ascaris suum: regulatory kinetic studies and activity near physiological conditions. Journal of Biological Chemistry. 1982;257(7):3801–3806. [PubMed] [Google Scholar]
- 16.Buckwitz D, Jacobasch G, Gerth C. Phosphofructokinase from Plasmodium berghei: influence of Mg2+, ATP and MG2+-complexed ATP. Biochemical Journal. 1990;267(2):353–357. doi: 10.1042/bj2670353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Saavedra E, Encalada R, Pineda E, Jasso-Chavez R, Moreno-Sanchez R. Glycolysis in Entamoeba histolytica: biochemical characterization of recombinant glycolytic enzymes and flux control analysis. FEBS Journal. 2005;272(7):1767–1783. doi: 10.1111/j.1742-4658.2005.04610.x. [DOI] [PubMed] [Google Scholar]
- 18.Lloyd GM. Kinetic properties of phosphofructokinase (and fructose bisphosphatase) of the liver fluke, Fasciola hepatica . International Journal for Parasitology. 1983;13(5):475–481. doi: 10.1016/s0020-7519(83)80011-3. [DOI] [PubMed] [Google Scholar]
- 19.Fleige T, Fischer K, Ferguson DJP, Gross U, Bohne W. Carbohydrate metabolism in the Toxoplasma gondii apicoplast: localization of three glycolytic isoenzymes, the single pyruvate dehydrogenase complex, and a plastid phosphate translocator. Eukaryotic Cell. 2007;6(6):984–996. doi: 10.1128/EC.00061-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Aguilar Z, Urbina JA. The phosphofructokinase of Trypanosoma (Schizotrypanum) cruzi: purification and kinetic mechanism. Molecular and Biochemical Parasitology. 1986;21(2):103–111. doi: 10.1016/0166-6851(86)90013-7. [DOI] [PubMed] [Google Scholar]
- 21.Cronin CN, Tipton KF. The roles of magnesium ions in the reaction catalysed by phosphofructokinase from Trypanosoma brucei . Biochemical Journal. 1987;247(1):41–46. doi: 10.1042/bj2470041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Sharma B, Ghatak S, Malhotra OP, Kaushal NA. Stabilisation and characterisation of phosphofructokinase purified from Setaria cervi, a bovine filarial parasite. Helminthologia. 1995;32:15–23. [Google Scholar]
- 23.Saz HJ. Biochemical aspects of filarial parasites. Trends in Biochemical Sciences. 1981;6:117–119. [Google Scholar]
- 24.Campos G, Guixe V, Babul G. Kinetic mechanism of phosphofructokinase-2 from Escherichia coli. A mutant enzyme with a different mechanism. Journal of Biological Chemistry. 1984;259(10):6147–6152. [PubMed] [Google Scholar]
- 25.Arechaga I, Martínez-Costa OH, Ferreras C, Carrascosa JL, Aragón JJ. Electron microscopy analysis of mammalian phosphofructokinase reveals an unusual 3-dimensional structure with significant implications for enzyme function. FASEB Journal. 2010;24(12):4960–4968. doi: 10.1096/fj.10-165845. [DOI] [PubMed] [Google Scholar]
- 26.Storey KB. Phosphofructokinase from oyster adductor muscle. In: Wood WA, Colowick SP, Kaplan NO, editors. Methods in Enzymology. Vol. 90. New York, NY, USA: Academic Press; 1982. pp. 39–44. [DOI] [PubMed] [Google Scholar]
- 27.Köhler P. The pathways of energy generation in filarial parasites. Parasitology Today. 1991;7(1):21–25. doi: 10.1016/0169-4758(91)90081-x. [DOI] [PubMed] [Google Scholar]
- 28.Sharma B, Kaushal NA, Ghatak S. Phosphofructokinase of bovine filarial parasite, Setaria cervi . Indian Journal of Parasitology. 1987;11:5–8. [Google Scholar]
- 29.Racker E. Spectrophotometric measurement of hexokinase and phosphohexokinase activity. Journal of Biological Chemistry. 1947;167:843–854. [PubMed] [Google Scholar]
- 30.Leite TC, Da Silva D, Coelho RG, Zancan P, Sola-Penna M. Lactate favours the dissociation of skeletal muscle 6-phosphofructo-1- kinase tetramers down-regulating the enzyme and muscle glycolysis. Biochemical Journal. 2007;408(1):123–130. doi: 10.1042/BJ20070687. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Storey KB. Phosphofructokinase from flight muscle of the cockroach, Periplaneta Americana: control of enzyme activation during flight. Insect Biochemistry. 1985;15(5):663–666. [Google Scholar]
- 32.Mettrick DF. Interactions between parasites and their hosts: metabolic aspects. International Journal for Parasitology. 1987;17(1):111–117. doi: 10.1016/0020-7519(87)90032-4. [DOI] [PubMed] [Google Scholar]
- 33.Ferreras C, Hernández ED, Martínez-Costa OH, Aragón JJ. Subunit interactions and composition of the fructose 6-phosphate catalytic site and the fructose 2,6-bisphosphate allosteric site of mammalian phosphofructokinase. Journal of Biological Chemistry. 2009;284(14):9124–9131. doi: 10.1074/jbc.M807737200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Claustre S, Colette DC, Lakhdar-Ghazal F, et al. Exploring the active site of Trypanosoma brucei phosphofructokinase by inhibition studies: specific irreversible inhibition. Biochemistry. 2002;41(32):10183–10193. doi: 10.1021/bi020082z. [DOI] [PubMed] [Google Scholar]
- 35.Suarez RK, Lighton JR, Joos B, Roberts SP, Harrison JF. Energy metabolism, enzymatic flux capacities, and metabolic flux rates in flying honeybees. Proceedings of the National Academy of Sciences of the United States of America. 1996;93(22):12616–12620. doi: 10.1073/pnas.93.22.12616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Rodríguez E, Lander N, Ramirez JL. Molecular and biochemical characterisation of Trypanosoma cruzi phosphofructokinase. Memorias do Instituto Oswaldo Cruz. 2009;104(5):745–748. doi: 10.1590/s0074-02762009000500014. [DOI] [PubMed] [Google Scholar]
- 37.Deng Z, Huang M, Singh K, et al. Cloning and expression of the gene for the active PPi-dependent phosphofructokinase of Entamoeba histolytica . Biochemical Journal. 1998;329(3):659–664. doi: 10.1042/bj3290659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Navid A, Ortoleva PJ. Simulated complex dynamics of glycolysis in the protozoan parasite Trypanosoma brucei . Journal of Theoretical Biology. 2004;228(4):449–458. doi: 10.1016/j.jtbi.2004.02.007. [DOI] [PubMed] [Google Scholar]
- 39.Taylor M, Gutteridge WE. The regulation of phosphofructokinase in epimastigote Trypanosoma cruzi . FEBS Letters. 1986;201(2):262–266. doi: 10.1016/0014-5793(86)80620-2. [DOI] [PubMed] [Google Scholar]
- 40.Yamasaki T, Nakajima H. Phosphofructokinase (PFK) Nippon Rinsho. 2004;62(supplement 12):835–839. [PubMed] [Google Scholar]
- 41.Khoja SM, Beach NL, Kellett GL. The isolation and characterization of phosphofructokinase from the epithelial cells of rat small intestine. Biochemical Journal. 1983;211(2):373–379. doi: 10.1042/bj2110373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Kamemoto ES, Lan L, Mansour TE. In vivo regulation of phosphorylation level and activity of phosphofructokinase by serotonin in Fasciola hepatica . Archives of Biochemistry and Biophysics. 1989;271(2):553–559. doi: 10.1016/0003-9861(89)90307-x. [DOI] [PubMed] [Google Scholar]
- 43.Srinivasan NG, Wariso BA, Kulkarni G, Rao GSJ, Harris BG. Phosphofructokinase from Dirofilaria immitis: stimulation of activity by phosphorylation with cyclic AMP-dependent protein kinase. Journal of Biological Chemistry. 1988;263(7):3482–3485. [PubMed] [Google Scholar]
- 44.Andrés V, Carreras J, Cussó R. Myofibril-bound muscle phosphofructokinase is less sensitive to inhibition by ATP than the free enzyme, but retains its sensitivity to stimulation by bisphosphorylated hexoses. International Journal of Biochemistry and Cell Biology. 1996;28(10):1179–1184. doi: 10.1016/1357-2725(96)00046-5. [DOI] [PubMed] [Google Scholar]
- 45.Cadefau JA, Parra J, Tauler A, Cussó R. Contractile activity modifies Fru-2,6-P2 metabolism in rabbit fast twitch skeletal muscle. Journal of Biological Chemistry. 1999;274(45):31961–31966. doi: 10.1074/jbc.274.45.31961. [DOI] [PubMed] [Google Scholar]
- 46.Zancan P, Almeida FV, Faber-Barata J, Dellias JM, Sola-Penna M. Fructose-2,6-bisphosphate counteracts guanidinium chloride-, thermal-, and ATP-induced dissociation of skeletal muscle key glycolytic enzyme 6-phosphofructo-1-kinase: a structural mechanism for PFK allosteric regulation. Archives of Biochemistry and Biophysics. 2007;467(2):275–282. doi: 10.1016/j.abb.2007.08.032. [DOI] [PubMed] [Google Scholar]
- 47.Hansen T, Schönheit P. Purification and properties of the first-identified, archaeal, ATP- dependent 6-phosphofructokinase, an extremely thermophilic non-allosteric enzyme, from the hyperthermophile Desulfurococcus amylolyticus . Archives of Microbiology. 2000;173(2):103–109. doi: 10.1007/s002039900114. [DOI] [PubMed] [Google Scholar]
- 48.Estévez AM, Heinisch JJ, Aragón JJ. Functional complementation of yeast phosphofructokinase mutants by the non-allosteric enzyme from Dictyostelium discoideum. FEBS Letters. 1995;374(1):100–104. doi: 10.1016/0014-5793(95)01085-s. [DOI] [PubMed] [Google Scholar]