

Abstract
Infection and dissemination of HIV-1 through the female body after vaginal intercourse depends on the activation/differentiation status of mucosal CD4 T cells. Here, we investigated this status and the susceptibility to HIV-1 infection of human cervico-vaginal tissue ex vivo. We found that virtually all T cells are of the effector memory phenotype with broad CCR5 expression. As it does in vivo, human cervico-vaginal tissue ex vivo preferentially supports the productive infection of R5 HIV-1 rather than that of X4 HIV-1 in spite of broad expression of CXCR4. X4 HIV-1 replicated only in the few tissues that were enriched in CD27+CD28+ effector memory CD4 T cells. Productive infection of R5 HIV-1 occurred preferentially in activated CD38+CD4 T cells and was followed by a similar activation of HIV-1-uninfected (bystander) CD4 T cells that may amplify viral infection. These results provide new insights into the dependence of HIV-1 infection and dissemination on the activation/differentiation of cervico-vaginal lymphocytes.
INTRODUCTION
Heterosexual transmission of HIV-1 occurs predominantly through vaginal intercourse1, rendering the female lower genital tract mucosa an important gateway for virus entry. At this mucosal site, HIV-1 encounters its target cells, and after an initial local amplification eventually disseminates to the draining lymph nodes, establishing a systemic infection2. However, not all HIV-1 variants are capable of establishing such infection. For example, although both CCR5-tropic (R5) and CXCR4-tropic (X4) HIV-1 variants are present in semen, only virus utilizing CCR5 but not CXCR4 is apparently transmitted and dominant in the early stages of HIV-1 disease3,4. Moreover, data have been recently published on the genetic and phenotypic characteristics of “founder” R5 HIV-1 variants that are capable of being efficiently transmitted and of establishing systemic infection5. Since cervico-vaginal tissue is the first gateway for HIV-1 during heterosexual transmission, the lower genital tract may serve as the first “gatekeeper”, conferring protection from at least some HIV-1 variants. Understanding of the “gatekeeping” mechanisms of HIV-1 transmission is critically important for the development of effective HIV-1-preventive measures (microbicides, virustats, and vaccines). Such an understanding requires a robust knowledge of the phenotypes and functional features of the cells which in the lower genital mucosa are implicated in the early events of HIV-1 transmission6. It seems that CD4 T cells are the predominant target for HIV-1 infection at the mucosal invasion sites2,7. Although macrophages and various dendritic cells (DC) are important for the transmission of HIV-1, in particular by presenting infectious virions to CD4 T cells, the latter appear to be the cells that predominantly support productive HIV infection 5,8,9,10,11. Thus, investigating the pattern of HIV-1 infection in CD4 T cells and characterizing cells that are major viral targets is crucial for our understanding of the mechanisms of HIV-1 transmission in the female genital tract.
In other anatomical sites that are critical for HIV-1 pathogenesis and transmission (lymph nodes and rectal mucosa), cell activation, cell differentiation, and expression of HIV-1 coreceptors on CD4 T-cell subsets have been identified as the most important factors that determine the efficiency of HIV-1 production. In spite of the critical importance of cervico-vaginal tissue to HIV-1 infection, these key characteristics remain largely unknown for cervico-vaginal T cells. Here, we report data on these characteristics.
We found that in cervico-vaginal tissue CD4 and CD8 T cells are predominantly of the highly differentiated effector memory phenotype with a variable pattern of expression of activation markers. Using an ex vivo system of cervical tissue12 that we have now further developed, we identified the phenotype and the activation and differentiation status of CD4 T cells that are preferentially infected by HIV-1. We also found that R5 HIV-1 replication in cervico-vaginal tissue increases the fraction of activated cells that are preferentially infected, creating a vicious circle between R5 HIV-1 replication and the number of cell targets that support this infection. Finally, investigating the gatekeeping mechanism that selects between R5 and X4 HIV-1 infection we found that, in ex vivo cervico-vaginal tissue, susceptibility to productive R5 HIV-1 infection is much higher than that to X4 HIV-1. We linked this difference to the particular phenotypic and activation patterns of the CD4 T cells in this tissue.
RESULTS
We analyzed various cell populations in human cervico-vaginal tissue. In particular, we focused on T cells which are located in the epithelial layer, lamina propria or submucosa and therefore are among the first cells that encounter HIV-1 in vaginal intercourse13. Towards this goal, in fresh surgical samples of this tissue we identified lymphocytic populations from their expression of membrane antigens CD3, CD4, CD8, CCR7, CD45RA, CD27, and CD28, their activation status from their expression of CD25, CD38, CD57, CD69, CD95, and HLA-DR, and their potential susceptibility to HIV-1 infection from the expression of HIV-1 coreceptors CCR5 and CXCR4. To study HIV-1 infection in this tissue, we developed an original protocol for its long-term culture. We infected this tissue with R5 and X4 HIV-1, monitored viral replication, identified HIV-1-infected cells and their activation status as well as the activation status of uninfected (bystander) cells, and evaluated cellular depletion.
T-cell subsets in human cervico-vaginal tissue
First, using flow cytometry we evaluated the distribution of cells of various subtypes in cervico-vaginal tissue (Fig. 1A–C). On average, T (CD3+) cells constitute 82±5.4% of total lymphocytes, whereas B cells, as identified from the expression of CD19 or CD20, constitute 31.5±1.5% of CD3− cells. On average, cervico-vaginal tissue contains 684±100 T (CD3+) cells per milligram of wet tissue (n=4). Also, we identified plasmacytoid DCs (PDCs) and myeloid DCs (MDCs) and calculated that per milligram of wet tissue there are on average 96.1±22 and 80.7±39 (n=5) of these, respectively. Also, we found that there are on average 235±92 (n=5) macrophages per milligram of wet tissue (Fig. 1B, C).
Figure 1. Identification of T lymphocytes, dendritic cells and macrophages in human cervico-vaginal tissue.
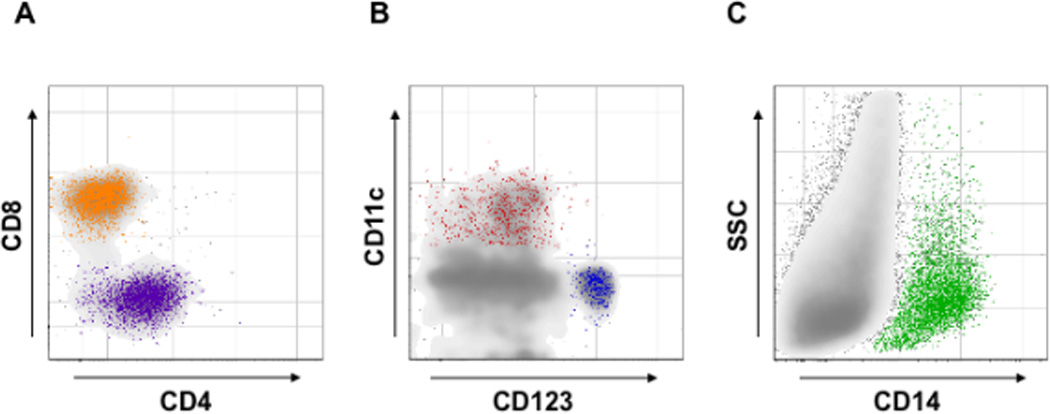
Cells suspension from digested fresh cervico-vaginal tissue blocks was stained for markers of T lymphocytes, dendritic cells [plasmacytoid (PDCs) and myeloid (MDCs)] and macrophages and analyzed with flow cytometry. Presented are bivariate dot plots of the distribution of CD4 and CD8 T cells (A), PDCs and MDCs (B), and macrophages (C). CD4- and CD8-positive T cells were identified gating on CD3+ lymphocytes. PDCs (CD123+) and MDCs (CD11c+) were identified gating on cells negative for CD3, CD14, CD16, CD56, CD19, CD20 and positive for HLA-DR. Macrophages (CD14+) were identified gating on CD3, CD56, CD19, CD20 negative cells. Presented plots are representative of experiments with cervico-vaginal tissues from five to twelve donors.
Note that T lymphocytes, PDCs, MDCs, and macrophages that are involved in HIV-1 infection are present in human cervico-vaginal tissue.
We compared the distribution of T cells in cervico-vaginal tissue with that in lymphoid (tonsillar) tissue. CD4 and CD8 T cells accounted for almost the entire CD3+ cell subset in cervico-vaginal tissue (46±4.5% and 52±3.8%, respectively, n=12), with an average CD4:CD8 ratio of 1.1±0.1 (Fig. 1A, Fig. 2A). In contrast, in tonsillar tissue there were 133,747±32,457 T (CD3+) cells per milligram of wet tissue. B cells constituted 97.7±0.3% of CD3− cells. CD4 T cells represented the predominant T-cell subpopulation, with a CD4:CD8 ratio of 4.4±0.5% (n=5).
Figure 2. T-lymphocyte subpopulations in human cervico-vaginal and tonsillar tissue.
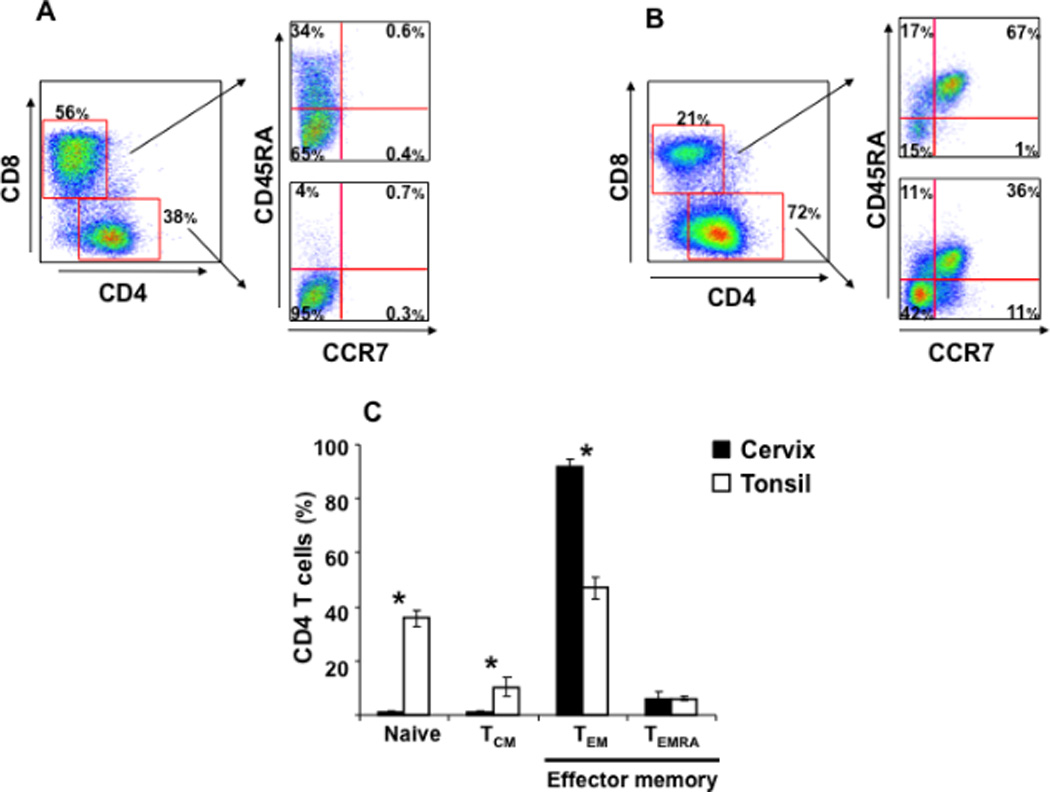
Cells suspension was stained for lymphocytic markers and analyzed with flow cytometry. Presented are bivariate dot plots of the distribution of different T-lymphocyte subpopulations for cervico-vaginal (A) and tonsillar (B) tissues. For each tissue, the distribution of CD4 and CD8 T cells is displayed in the left panel. The distribution of naïve, TCM, TEM, and TEMRA subpopulations among CD4 and CD8 T cells is displayed in the upper right panel and in the lower right panel, respectively. Presented plots are representative of experiments with cervico-vaginal tissues from eleven donors and tonsillar tissues from five donors. In C are presented the means ± SEM of the fractions of naïve, TCM, TEM, and TEMRA among CD4 T cells from these tissues. *Indicates a statistically significant difference with p<0.05.
Note that the distribution of naïve, TCM, and TEM CD4 T cells in cervico-vaginal tissues is different from that in tonsillar tissues.
Second, to characterize the phenotypes of CD4 and CD8 T-cell subpopulations in cervico-vaginal tissue, we investigated the distributions of naïve (CCR7+/CD45RA+), central memory (TCM; CCR7+/CD45RA−), effector memory (TEM; CCR7−/CD45RA−), and terminal effector memory (TEMRA; CCR7−/CD45RA+) cells14 (Fig. 2A). We found that in cervico-vaginal tissue 92±3% of total CD4 T cells were TEM and 6±2.4% were TEMRA, while 0.8±0.5% and 1±0.3% were respectively naïve and TCM (n=11) (Fig. 2C). Similar fractions of naïve and TCM cells were found among CD8 T cells (1.3±0.6% and 0.5±0.2%, respectively; p>0.05), while the fractions of TEM and TEMRA among CD8 T cells (respectively 69±5.5% and 29±5%) were significantly different from those in CD4 T cells (n=11, p<0.0003). In tonsillar tissue, naive, TCM, TEM, and TEMRA cells constituted 35.8±3.1%, 10.3±3.4%, 47±5.8%, and 6±0.7% of CD4 and 62.8±3.2%, 5±0.4%, 20.2±4.6%, and 13.8±3.6% of CD8 T cells, respectively (n=5) (Fig. 2B). Thus, in the CD4 T cell subset the fractions of naïve, TCM, and TEM cells were significantly different between cervico-vaginal and tonsillar tissue (p<0.03) (Fig. 2C).
Since we found that the majority of T cells in cervico-vaginal tissue were of the effector memory phenotype (TEM, TEMRA), we analyzed their expression of the costimulatory molecules CD27 and CD28, which define cell differentiation towards specialized effector functions15,16. Among CD4 T cells, 21.2±5.3% were of early-differentiated effector phenotype (ED; CD27+CD28+), 47.7±5% were of intermediate effector phenotype (ID; CD27+CD28− or CD27−CD28+), and 31.2±8.4% were of late-differentiated effector phenotype (LD; CD27−CD28−) (n=10) (Fig. 3). Similar results were obtained for CD8 T cells in cervico-vaginal tissue, in which phenotypes ED, ID, and LD constituted 16.4±4%, 52.6±4%, and 30.9±2.9% of CD8 T cells, respectively (n=10).
Figure 3. Distribution of early-, intermediate-. and late-differentiated effector memory CD4 T cells in human cervico-vaginal and tonsillar tissues.
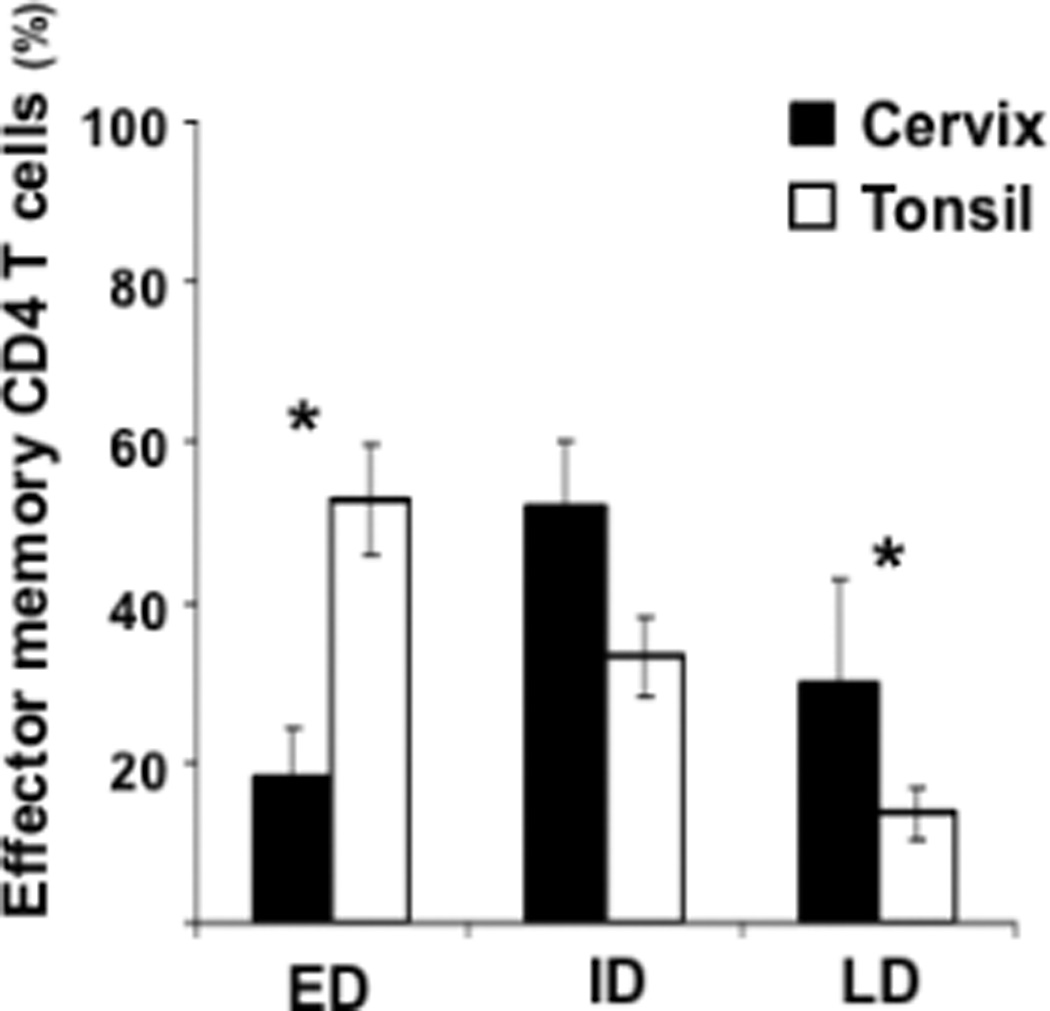
Early- (ED), intermediate- (ID) and late-differentiated (LD) effector memory CD4 T cells (TEM, TEMRA) were identified on the basis of the expression of CD27 and CD28 costimulatory molecules. Presented are means ± SEM of the fractions of ED, ID, and LD effector memory CD4 T cells in cervico-vaginal (n=10) and tonsillar (n=5) tissues. *Indicates a statistically significant difference with p<0.05.
Note that the distribution of ED and LD effector memory CD4 T cells in cervico-vaginal tissues is different from that in tonsillar tissues.
The distribution of these cell subpopulations among effector memory cells (TEM, TEMRA) in lymphoid tissue (tonsils) was significantly different from that in cervico-vaginal tissue (p<0.05). Among CD4 T cells, the ED, ID, and LD subpopulations constituted 53±6.9%, 33.2±5.1%, and 13.7±3.3%, respectively (n=5), while these subsets constituted respectively 38.6±6%, 49±1.6%, and 12.2±4.5% among CD8 T cells (n=5) (Fig. 3).
Expression of activation markers and HIV-1 coreceptors on T cells in human cervico-vaginal tissue
Since for several types of non-genital tissue it has been shown that productive infection by HIV-1 occurs mainly in activated T cells17, we evaluated the expression of activation markers on T cells in fresh cervico-vaginal tissue. Both in CD4 and in CD8 T cells, CD69 and CD95 were expressed on more than 90% of cells (Table 1). In contrast, CD57, CD38, and HLA-DR were expressed on a smaller fraction of T cells, and differentially in CD4 and CD8 T cells: 10.6±2.7%, 19.8±4%, and 3.3±1% of CD4 T cells, respectively, expressed these markers, while among CD8 T cells these fractions constituted 36.5±5.2%, 30.6±6.2%, and 7.4±2.4%, respectively (p<0.05, n=5, n=12, n=9). CD25 expression was similar on CD4 and CD8 T cells (7.6±2% and 6.7±4.6%, respectively; p=0.2, n=10) (Table 1).
Table 1.
Expression of activation markers on CD4 and CD8 T cells from cervico-vaginal and tonsillar tissue.
| Cervix (%) | Tonsil (%) | ||
|---|---|---|---|
| CD4 | CD69 | 91.7 ± 7.7 | 73.3 ± 4.3 * |
| CD25 | 7.6 ± 2 | 4.5 ± 0.9 | |
| HLA-DR | 3.3 ± 1 | 6.6 ± 1.2 | |
| CD38 | 19.8 ± 4 | 45.8 ± 3.8 * | |
| CD57 | 10.6 ± 2.7 | 12 ± 1.2 | |
| CD95 | 92.1 ± 3 | 66.3 ± 4.8 * | |
| CD8 | CD69 | 97.1 ± 1.5 | 47 ± 5.2 * |
| CD25 | 6.7 ± 4.4 | 7.5 ± 2.1 | |
| HLA-DR | 7.4 ± 2.4 | 5.2 ± 1 | |
| CD38 | 30.6 ± 6.2 | 33.1 ± 2 | |
| CD57 | 36.5 ± 5.2 | 11.1 ± 2.6 * | |
| CD95 | 96 ± 1.4 | 31.5 ± 7.4 * |
indicate a statistical significant difference with p< 0.05
The expression of activation markers in cervico-vaginal tissue was different from that in lymphoid tissue: in particular, in CD4 and CD8 T cells from cervico-vaginal tissue there was a higher expression of CD95 and CD69 than in lymphoid tissue (p<0.05, n=6). The expression of CD38 was higher on CD4 T cells in lymphoid than in cervico-vaginal tissue (p=0.0003, n=12). There was no significant difference in the expression of CD25 and HLA-DR on CD4 and CD8 T cells between lymphoid and cervico-vaginal tissue (p>0.05, n=9), while the expression of CD57 on CD8 T cells was lower in lymphoid than in cervico-vaginal tissue (p=0.005, n=5) (Table 1).
Furthermore, we evaluated the expression of the HIV-1 coreceptors CCR5 and CXCR4. We found that 44±1.6% of CD4 T lymphocytes expressed both CXCR4 and CCR5, and 44±1.5% of CD4 T cells expressed CXCR4 exclusively, whereas 7±0.8% expressed CCR5 exclusively (n=3). The distribution of the expression of these HIV-1 coreceptors was different from that in lymphoid tissue (Fig. 4). In this tissue, 15±4% of CD4 T cells coexpressed CCR5 and CXCR4, 4±1% expressed CCR5 exclusively, and 71±4.5% expressed only CXCR43.
Figure 4. Expression of HIV-1 coreceptors on CD4 T cells from human cervico-vaginal and tonsillar tissues.
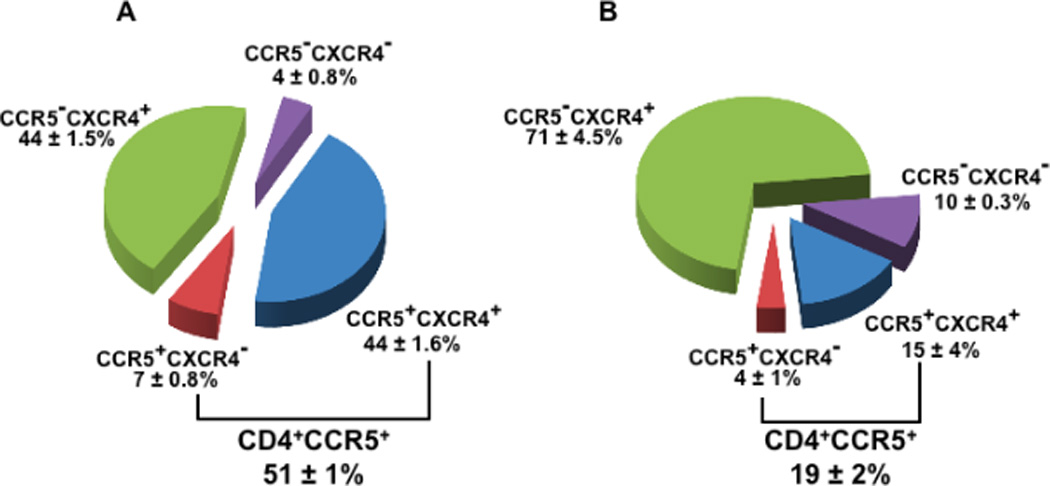
Cells isolated from fresh tissue blocks were stained for lymphocytic markers and analyzed with flow cytometry. Presented are means ± SEM of the fractions of total CD4 T cells expressing CCR5 and CXCR4 in cervico-vaginal (A, n=4) and tonsillar (B, n=3) tissues.
Note that the fraction of CD4 T cells expressing CCR5 is higher in cervico-vaginal than in tonsillar tissue, while the fraction of CD4 T cells expressing CXCR4 is similar.
Development of a raft-based culture system of human cervico-vaginal tissue that efficiently supports HIV-1 infection ex vivo
Culture on collagen sponge gel rafts at the liquid-air interface has been described for several tissues as a system that preserves tissue viability, cytoarchitecture, and functions for at least 2 to 3 weeks12,18,19,20. Here, we used such a system to culture cervico-vaginal tissue and infect it with HIV-1 under controlled laboratory conditions.
Cervical tissue obtained from surgery was dissected into tissue blocks and cultured on collagen sponge gel rafts for 12 days. Phenotypic analysis of T cells in these tissue blocks showed that the distributions of all the T-cell populations described above as well as the fractions of cells expressing activation markers and HIV-1 coreceptors did not change between day 0 and day 12 of culture (n=8, p>0.5). Cervico-vaginal tissue blocks were infected with HIV-1 (16 blocks per experimental condition), and viral replication was evaluated from the release of p24gag into the culture medium. In tissue blocks inoculated with R5BaL HIV-1, viral replication became evident on day 6 post-inoculation and increased during the entire course of the experiment up to day 12. At day 12 post-infection, the release of p24gag into the supernatant was 1,619±635 pg/mL (Fig. 5A). On average, the total cumulative release of p24gag into the culture medium between day 6 and day 12 post-inoculation was 3,191±1,158 pg/mL of p24gag (n=16). Over this period, each block of tissue produced, on average, 199±72 pg.
Figure 5. R5BaL HIV-1 infection of human cervico-vaginal tissues.
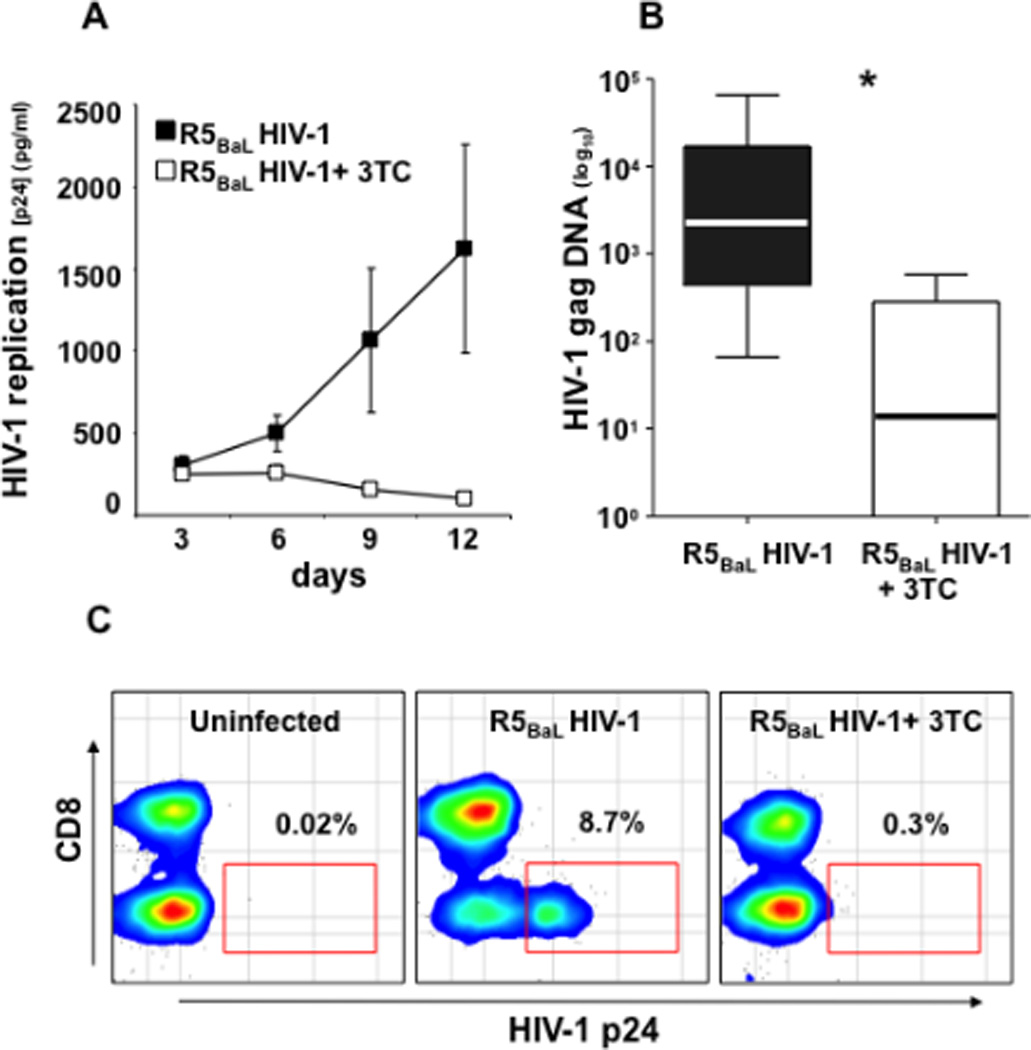
Donor-matched blocks of human cervico-vaginal tissue were inoculated ex vivo with R5BaL HIV-1 and treated or not with 3TC (5 µM).
A: Kinetics of release of p24gag by inoculated tissues. Presented are means ± SEM for tissues from 16 donors. For each donor, each datum point represents pooled viral release from 16 tissue blocks over 3-day periods. B: The amount of HIV-1 gag complete reverse transcripts at day 12 post-infection. Presented are medians and the interquartile ranges; the whiskers of the graph show the largest and smallest values. C: The distribution of HIV-1-infected cells in donor-matched tissue blocks: uninfected, inoculated with R5BaL HIV-1, inoculated with R5BAL, and treated with 3TC (5 µM). The amounts of HIV-1 p24gag-positive cells are expressed as percentage of total CD3+ T cells. Presented are bivariate density plots of a representative experiment with cervico-vaginal tissues from one out of 13 donors. *Indicates a statistically significant difference with p<0.05.
Note that p24gag accumulation in culture medium measured by cytometric bead assay, production of HIV-1 complete reverse transcripts measured by real-time PCR, and identification of HIV-1-infected cells by flow cytometry confirm productive infection of human cervico-vaginal tissues by R5BaL HIV-1.
Potentially, some of the p24gag measured in the medium can reflect the slow release of viral inoculum adsorbed on the tissue rather than de novo HIV-1 production. To discriminate between these two processes, we performed control experiments in which in donor-matched tissues we prevented de novo viral production with an inhibitor of HIV-1 reverse transcriptase [lamivudine (3TC), 5 µM]. In tissues treated with 3TC there was an 89.7±5.3% reduction of p24gag release on day 12 of culture compared with matched tissues not treated with 3TC (p=0.02, n=16) (Fig. 5A). These data were confirmed in experiments with another HIV-1 inhibitor, PSC-RANTES (which downregulates R5 HIV-1 coreceptor CCR5)21. In these tissues, there was an 80.5±10.2% reduction in p24gag release into the medium (n=2). Hence, the amount of p24gag in the medium bathing inoculated tissue largely reflects productive HIV-1 infection.
To confirm productive infection by R5BaL HIV-1 in cervico-vaginal tissue, we quantified the complete reverse transcripts using real-time PCR22. In R5BaL HIV-1-inoculated tissues, the median amount of complete reverse transcripts on day 12 post-infection was 2,288 copies/106 cells (IQR 432−716,869), while in 3TC-treated cultures it was reduced to 13.9 copies/106 cells (IQR 0−286) (p=0.0001, n=14) (Fig. 5B).
Finally, flow cytometry of tissue cells stained for p24gag confirmed productive R5BaL HIV-1 infection. Among CD3+ cells on day 12 post-inoculation, the fraction of p24gag-expressing cells was 6.6±1.4% (n=13). Treatment with 3TC reduced this fraction to 0.8±0.3% (n=13, p=0.002). Gating on CD3+CD8− cells (to include T lymphocytes that downregulated CD4 because of HIV-1 infection23) revealed that on average, in R5BaL HIV-1-inoculated tissue on day 12 post-inoculation 14.1±3.3% (n=13) of CD8− T cells expressed p24gag. Treatment with 3TC reduced the fraction of HIV-1-infected cells on average to 1.5±0.8% (p=0.002, n=13) (Fig. 5C). Among CD3+CD8+ cells, the fraction of p24gag-positive cells was 0.3±0.07, and this fraction was not significantly reduced by 3TC treatment (0.2±0.07; n=13, p=0.3). Also, we evaluated the fraction of HIV-infected cells on day 12 post-HIV-1BAL-infection in the CD3− cell subset. In tissues infected with HIV-1BAL, 0.45±0.1% of CD3− cells expressed p24gag, while in tissues infected and treated with 3TC 0.36±0.06% (n=13) of CD3− cells expressed p24gag. The difference between these values was not statistically significant (p=0.31).
Thus, cervico-vaginal tissue ex vivo preserves different lymphocyte populations and supports productive HIV-1 infection in CD3+CD8− T cells but not in CD3+CD8+ T cells or in CD3− cells.
T-cell activation and depletion in R5 HIV-1-infected human cervico-vaginal tissue
To evaluate depletion in various T-cell subpopulations in human cervico-vaginal tissue infected ex vivo with R5BaL HIV-1, we enumerated, using flow cytometry, the numbers of total and CCR5-expressing CD8− T cells. On day 12 post-infection, 50.9±8.1% of CD8− T cells were depleted (n=16). Among CD8−CCR5+ T-cells, this depletion was 63.6±7.9%. Productive infection by R5BaL HIV-1 was the main cause for CD4 T-cell depletion, since the suppression of R5BaL HIV-1 infection by 3TC also decreased the number of depleted cells by 80.4±7.6% (p<0.05, n=16). Similar results were obtained when R5BaL HIV-1 infection was prevented by PSC-RANTES: in tissues treated with this inhibitor, the depletion of CD4 T-cells by R5BaL HIV-1 was reduced by approximately 80% compared with untreated tissues. To evaluate the role of apoptosis in cell depletion, we measured the expression of the early apoptotic marker APO2.7 in HIV-1-infected and uninfected T cells from cervico-vaginal tissue. On day 12 post-infection with R5BaL HIV-1, 34±8.8% of the apoptotic CD4 (CD8−) T cells were p24gag+, whereas among non-apoptotic CD4 (CD8−) T cells 19.3±5.7% expressed p24gag (p=0.01; n=6).
To study the relation between T-cell activation status and HIV-1 infection in cervico-vaginal tissue, we evaluated the fractions of activated and non-activated CD8− T cells in control and HIV-1-infected tissues. Also, we evaluated the fractions of activated cells among productively infected and bystander cells. HIV-1-infected cells were identified by means of intracellular staining for p24gag; activated cells were identified from the expression of the activation markers CD25, CD38, CD57, CD69, CD95 and HLA-DR. We found that on day 12 after R5BaL HIV-1 inoculation into cervico-vaginal tissue blocks, the expression of CD38 was higher among HIV-1-infected T cells (21.9±3.3%) than among the bystander (uninfected) T cells (15.8±2.4%; p=0.01, n=12). Also, the fraction of activated cells expressing CD38 among bystander T cells in HIV-1-inoculated tissue was higher than the fraction of activated CD8− T cells in donor-matched uninfected controls (15.8±2.4% vs. 12.7±1.3; p=0.02, n=12).
Except for CD38, there was no difference in the expression of other tested activation markers (HLA-DR, CD25, CD57, CD69, and CD95) between infected and bystander CD8− T cells in HIV-1-infected tissue. Similarly, no difference was observed between the expression of these activation markers on bystander CD8− T cells from HIV-1-infected and from donor-matched uninfected tissues. No increased expression of activation markers, including CD38, was observed on CD8 T cells in HIV-1-infected compared with donor-matched uninfected tissues.
Thus, R5BaL HIV-1 preferentially infects CD4 T cells of activated (CD38+) phenotype, and the number of such cells among uninfected bystander CD4 T cells is increased upon R5BaL HIV-1 infection of human cervico-vaginal tissue.
Infection of human cervico-vaginal tissue ex vivo with the X4 HIV-1 variant
To compare the ability of human cervico-vaginal tissue to support productive infection by HIV-1 variants of different coreceptor tropisms, we inoculated tissue blocks with X4LAI HIV-1 and R5BaL HIV-1. On average, X4LAI HIV-1 replicated less efficiently than R5BaL HIV-1. For X4LAI HIV-1, the average cumulative p24gag release between day 6 and day 12 post-inoculation was 536±86 pg/mL: 64.6±5.3% less than that for R5BaL HIV-1 (p=0.03, n=27). Also, in contrast to the production of R5BaL HIV-1, which constantly increased over the 12 days of infection, there was a steady drop of the release of X4LAI HIV-1 through the entire culture period (Fig. 6A).
Figure 6. X4LAI HIV-1 infection of human cervico-vaginal tissues.
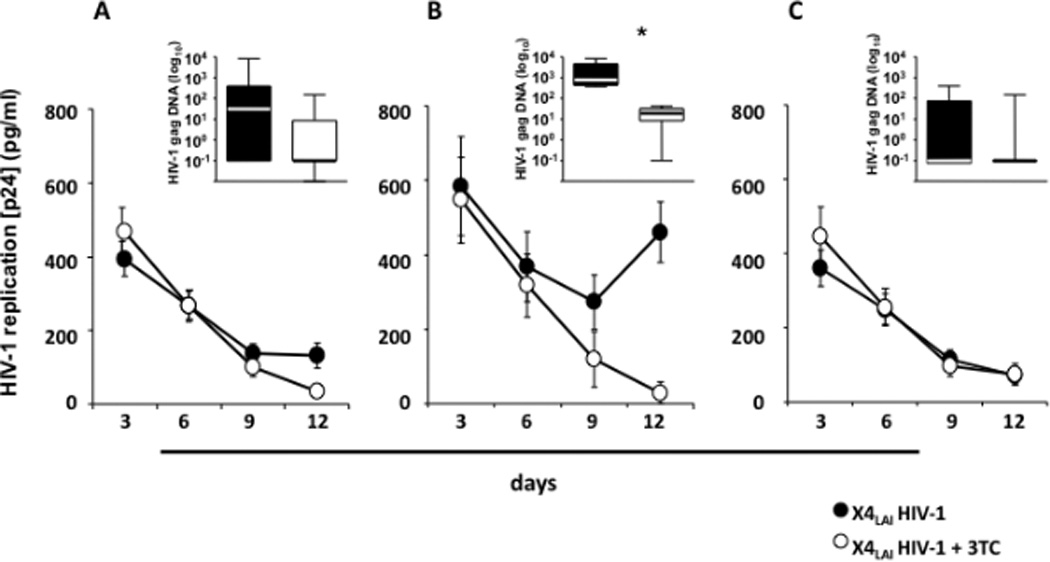
Donor-matched blocks of human cervico-vaginal tissue were inoculated ex vivo with X4LAI HIV-1 and treated or not with 3TC (5 µM).
A: Kinetics of release of p24gag by tissues from 27 donors. B: Kinetics of release of p24gag by tissues from 4 donors. These tissues were selected from the 27 presented in A based on the increase of p24gag release during culture. C: Kinetics of release of p24gag by tissues from the remaining 23 donor.
Each datum point for each tissue reflects p24gag produced by 16 tissue blocks over 3- day periods. Presented are means ± SEM.
Insets: The amount of HIV-1 gag complete reverse transcripts measured in tissue blocks at day 12 post-infection. Presented are the medians and interquartile ranges. The whiskers of the graphs show the largest and smallest values. *Indicates a statistically significant difference with p<0.05.An arbitrary value of 0.1 log10 copies/ml was assigned to samples negative for HIV-1 gag complete reverse transcripts to allow graphical representation of the data.
Note that in human cervico-vaginal tissues inoculation with X4LAI HIV-1 resulted in a productive infection in 4 out of 27 tested tissues as demonstrated by comparison of p24gag release and complete reverse transcripts in tissues treated or not with 3TC.
To evaluate the contribution of de novo production to the amount of the X4LAI HIV-1 released, we suppressed viral replication with 3TC. In contrast to R5BaL HIV-1-infected tissues, in tissues infected with X4LAI HIV-1, 3TC did not decrease the release of p24gag compared with the donor-matched untreated tissues (the average cumulative p24gag release between day 6 and day 12 of culture was 403±81pg/mL vs. 536±86pg/mL; n=27, p>0.05) (Fig. 6A). Also, there was no significant difference in the median numbers of complete reverse transcripts between X4LAI HIV-1–infected tissues untreated and treated with 3TC (respectively, 31.1 copies/106 cells (IQR 0−384) and 0 copies /106 cells (IQR 0−8.6); p=0.1, n=14). Thus, apparently on average no replication of X4LAI HIV-1 occurred in inoculated tissues, but rather the release of viral inoculum.
However, out of the 27 tissues we reported above, in 4 of them inoculation with X4LAI HIV-1 resulted in a productive infection, although with slower kinetics than R5BaL HIV-1 (Fig. 6B).
In these four cultures, the average cumulative p24gag release into the medium between day 6 and day 12 post-inoculation was 1,102±238 pg/mL (n=4), while for the rest of the tissues it was 438±77 pg/mL (n=23). Moreover, in tissue from these four donors there was a 61.8±9% reduction of p24gag release after treatment with 3TC (465±183 pg/mL) (p=0.005) in contrast to the twenty-three tissues in which 3TC did not significantly reduce p24gag release (23.3±5.7%; p=0.1) (Fig. 6C).
The analysis of complete reverse transcripts with real-time PCR confirmed the productive infection of these four tissues by X4LAI HIV-1: on day 12 post-infection the median amount of complete reverse transcripts was 437.5 copies/106 cells (IQR 239.4−4428) while in the other twenty-three tissues there were 0 copies/106 cells (IQR 0−172.1). In these four tissues, 3TC decreased the number of complete reverse transcripts by 97.6±1.3% (to the level of 19.1 copies/106 cells, IQR 8.5−31.7; p=0.01), confirming productive infection by X4LAI HIV-1.
We investigated whether the ability to support productive X4LAI HIV-1 infection in these four tissues was related to a particular distribution of various T-cell subsets or to their activation status. There was no significant difference in the fractions of naïve, TCM, TEM, TEMRA, and activated cells (CD25, CD38, CD57, CD69, CD95, and HLA-DR) as evaluated on day 12 post-inoculation between tissues that support and those that do not support X4LAI HIV-1 infection (p>0.05). However, we found that in tissues that supported productive X4LAI HIV-1 infection there were significantly more ED (CD27+CD28+) effector memory CD4 T cells than in tissues that did not support productive X4LAI HIV-1 infection (40.2±3.4%, n=4, vs. 19.9±3.7%, n=14; p=0.007) (Fig. 7). Accordingly, in tissues that supported productive X4LAI HIV-1 infection there were significantly fewer LD (CD27−CD28−) effector memory CD4 T cells than in tissues that did not support productive X4LAI HIV-1 infection (3.6±1.1%, n=4, vs. 21.2±5.6%, n=14; p=0.01). There was no statistical difference in the fraction of ID CD4 T cells between tissues that supported and those that did not support X4LAI HIV-1 infection (p=0.7) (Fig. 7).
Figure 7. X4LAI HIV-1 infection of human cervico-vaginal tissue and distribution of early-, intermediate-, and late-differentiated effector memory CD4 T cells.
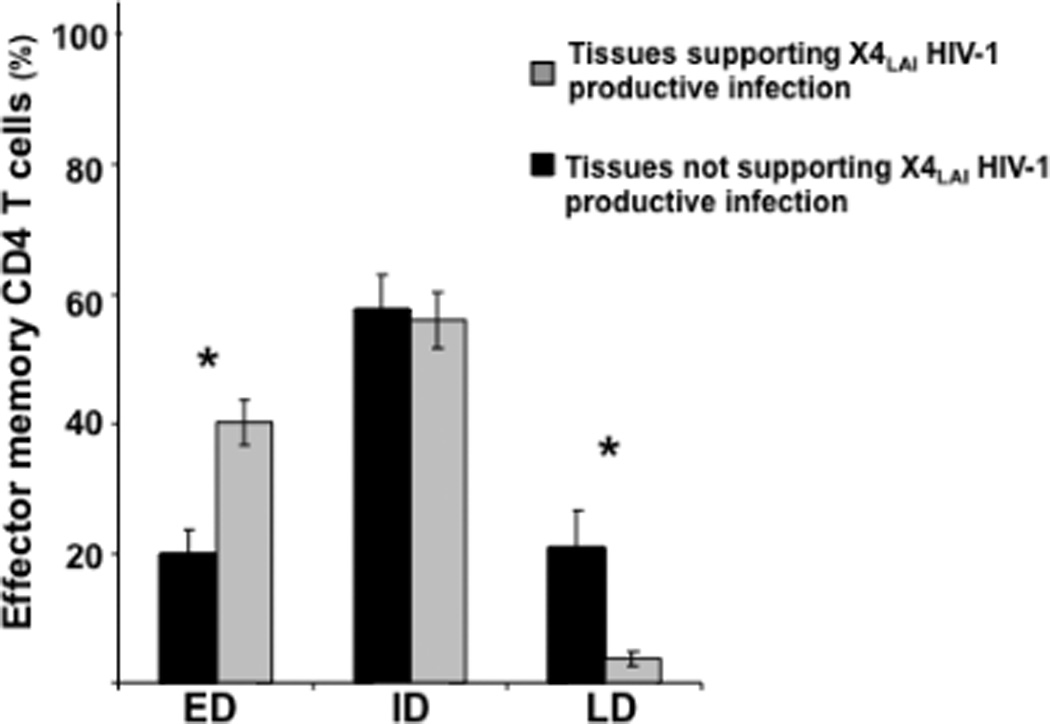
Presented are means ± SEM of the fractions of ED, ID, and LD effector memory CD4 T cells in cervico-vaginal tissues that supported (n=4) or did not support (n=23) productive infection by X4LAI HIV-1. *Indicates a statistically significant difference with p<0.05.
Note that these tissues differ in the distribution of ED and LD effector memory CD4 T cells.
Thus, in contrast to R5 HIV-1, which efficiently replicated in all tested cervico-vaginal tissues, X4 HIV-1 typically did not replicate. However, in 4 out of 27 tested tissues, X4 replicated, although with slower kinetics than R5 HIV-1. These tissues differed from the rest by a higher fraction of effector memory CD4 T cells expressing CD27 and CD28, but not by the distribution of other tested T-cell phenotypes or by the activation status of these cells.
DISCUSSION
In principle, the vaginal and cervical mucosa constitute a reliable natural barrier for HIV-1 transmission4,12,24. However, recent epidemiological data demonstrate a disproportionate increase in female infections, predominantly through vaginal intercourse1. It has been suggested that this route of HIV-1 transmission is facilitated by micro-abrasions occurring during the intercourse, to lesions due to sexually transmitted diseases, and to environmental factors that breach the integrity of the cervico-vaginal epithelial barrier4. Upon penetration of this barrier, HIV-1 interacts with mucosal cells, CD4 T cells, macrophages, and various dendritic cells. Although all these cells may play an important role in the early stages of HIV-1 infection, it seems that CD4 T cells are crucial to the establishment and dissemination of the infection25,26. Macrophages are productively infected ex vivo but do not seem to be the main target for the initial SIV/HIV infection in vivo11,27–29. On the other hand, even though intra- or sub-epithelial dendritic cells efficiently incorporate virions by endocytosis, it remains controversial whether they are productively infected26,27. Similarly, although in cervico-vaginal tissue various dendritic cells may contribute to the establishment of HIV infection (e.g., by presenting infectious virus to T lymphocytes as well as by the dissemination of newly produced virus), our ex vivo results show that CD4+T lymphocytes are the prevalent HIV-1-infected cells. Therefore, in the present work we focused on cells of this subtype.
Analysis of CD4 T cells isolated from other tissues relevant to HIV-1 pathogenesis and transmission has revealed that the CD4 T-cell population is not homogeneous in its ability to be infected and to support viral replication: this ability greatly depends on whether the cell is a memory or a naïve one, activated or non-activated, and on the pattern of expression of HIV-1 coreceptors30,31. These features may also contribute to the tissue’s ability to differentially support productive infection of different HIV-1 variants (“gatekeeping”). Surprisingly, with regard to the cervico-vaginal tissue which serves as an important gateway for HIV-1, there are few data on the status of T cells and on its relation to HIV-1 infection and production.
Here, we report data on these phenomena. We investigated the status of CD4 T cells in cervico-vaginal tissue and their ability to selectively support productive infection with R5 but not with X4 HIV-1. For these experiments, we further developed a previously described technique for the culture of cervical explants32, 33. We maintained cervico-vaginal tissue blocks on top of gel rafts at the liquid-air interface, instead of submerging them in the culture medium. In a raft-based culture, the collagen support and the oxygenation are sufficient to preserve the cytoarchitecture and thus to delay tissue deterioration18. Under these ex vivo conditions, cervico-vaginal tissue blocks readily support HIV-1 replication, as evidenced by the presence of complete reverse transcripts and of p24-positive T cells, and by the release of p24gag in the culture medium. Tissue treatment with 3TC almost completely prevented formation of the complete reverse transcripts, as well as the emergence of p24-positive T cells and the release of p24gag, thus confirming de novo viral production in the inoculated tissues. The residual viral infection in 3TC-treated tissue blocks is negligible and may reflect the extent of the first round of infection before the drug fully penetrates the tissue.
As with in vivo tissue, HIV-1 replication resulted in the depletion of CD4 T cells. We found that the apoptotic CD4+ T-cell population was enriched with HIV-infected cells, suggesting that induction of apoptosis contributes to the depletion of CD4 T cells in ex vivo cervico-vaginal tissue. The quantitative relation between CD4 T-cell apoptosis and their depletion is difficult to establish, since the number of apoptotic cells represents a “snapshot” while cell depletion is a cumulative parameter.
Also, since a raft-based culture system has been used to maintain lymphoid (lymph nodes or tonsils) and rectal tissues20, we were able to compare the abilities of cervico-vaginal, rectal, and lymphoid tissues to support replication of HIV-1 and to investigate the relation of these abilities to the distribution of T cells of various phenotypes in these tissues.
An important factor that may control HIV-1 replication is the functional status of T cells34. It has been shown that HIV-1 replication predominantly occurs in effector memory CD4 T cells31. Therefore, we analyzed the subpopulation of such T cells in cervico-vaginal tissue using flow cytometry and found that virtually all T cells are of the effector memory phenotype (TEM and TEMRA), with an almost complete absence of naïve and central memory T cells. In rectal mucosa, naïve, central memory, and effector memory T cells are differently distributed in Peyer`s patches and in the lamina propria: in the former the great majority of T lymphocytes are of naïve and central memory phenotype, while in the latter it is almost exclusively effector memory T cells that are present35. In contrast, in lymphoid tissue naïve T cells constitute more than 60% of the T-cell population.
The prevalence of effector memory cells among cervico-vaginal T cells is critical for guarding against various incoming pathogens. The ability of the cervico-vaginal tissue to defend against pathogens is complemented by the high prevalence of CD8 T cells. We found that in cervico-vaginal tissue the fraction of CD8 T cells was 3–4 times larger than in rectal, tonsillar, or lymph node tissues3,34. Besides their direct cytolytic function, CD8 T cells are the major source of soluble factors36 that suppress HIV-1 infection of CD4 T cells; thus, the higher prevalence of CD8 T cells in cervico-vaginal tissue may contribute to the higher resistance of this tissue to HIV-1 transmission compared with the highly efficient transmission through the rectal mucosa. Moreover, in cervico-vaginal tissue, CD8 T cells had a higher proportion of TEMRA than did CD4 T cells. This difference was accompanied by a higher expression of the activation markers CD38, HLA-DR, and CD57 in CD8 than in CD4 T cells. However, these defense mechanisms are not sufficient to prevent HIV-1 infection, and HIV-1 circumvents some of them for its benefit. Indeed, the prevalence of effector memory CD4 T cells of activated phenotype at the site of infection provides HIV-1 with more cell targets, thus creating a favorable environment for its efficient replication31.
Since, unlike isolated cells, cells in ex vivo tissues support HIV-1 replication without exogenous stimulation and activation, this experimental system is particular useful for the study of the effect of HIV-1 on cell activation. We found that in cervico-vaginal tissue R5 HIV-1 replication occurred preferentially in CD4 T cells that express the activation marker CD38. Surprisingly, this increase in activation of HIV-1-infected cells was associated with a concomitant increase in the expression of CD38 in HIV-1-uninfected (bystander) CD4 T cells, but not in CD8 T cells. No other tested activation markers were upregulated in cervico-vaginal tissue upon HIV-1 infection. Thus, it appears that HIV-1 replication in cervico-vaginal tissue is associated with specific activation of CD4 bystander T cells. Although HIV-1 viral proteins like Tat, Nef, and gp12037 may be directly involved in this process, it is also possible that host soluble mediators (e.g., type I interferon or other cytokines) 34,38 produced by HIV-1-infected and uninfected cells may mediate the upregulation of CD38 on bystander T cells. A similar phenomenon, albeit with different activation markers (CD25+HLA-DR+), has been reported for lymphoid tissue17. Thus, it appears that in both lymphoid and cervico-vaginal tissues, HIV-1 specifically upregulates surface molecules that are characteristic of the cells that are preferentially infected by this virus. This phenomenon may expand the local pool of cells that HIV-1 preferentially infects. Such an interpretation is consistent with a recent report, based on work on SIV-infected rhesus macaques, that demonstrated that viral infection in the genital tract spreads from a few isolated CD4 T cells. These cells trigger a local inflammatory response that induces the recruitment of other CD4 T cells. Thus, the infection of a few isolated CD4 T cells rapidly expands into the infection of clusters of numerous CD4 T cells, eventually favoring dissemination of the virus to the draining lymph node25. The bystander activation of local CD4 T cells that we observed in HIV-1-infected human cervico-vaginal tissue may represent a similar mechanism that facilitates the expansion of infection. Such bystander activation may in turn further amplify HIV-1 replication, and this vicious circle of infection–activation–infection would favor the systemic dissemination of the virus.
Although such a vicious circle seems to be general both for lymphoid and cervical tissue, in the former it can be generated by both R5 and X4 HIV-1, while cervico-vaginal tissue predominantly supports productive infection by R5 rather than X4 HIV-1. One of the factors that may contribute to this phenomenon is the high prevalence, demonstrated here, of CCR5 expression among cervico-vaginal CD4 T lymphocytes. However, the ability of this tissue to support R5 rather than X4 HIV-1 infection is not explained solely by the pattern of HIV-1 coreceptor expression, since although the expression of CCR5 on CD4 T cells is high almost all CD4 T cells express CXCR4 as well.
Other restriction factors that “gatekeep” against X4 HIV-1 infection should be investigated39,40. We identified the differentiation status of effector memory CD4 T cells as one such factor. On average, inoculation with X4LAI HIV-1 did not result in significant infection of cervico-vaginal explants: the amount of p24gag released into the medium from X4LAI HIV-1-inoculated tissue was similar to that released from 3TC-treated cultures inoculated with X4LAI HIV-1 or R5BaL HIV-1. However, in tissues from 4 out of 27 donors we documented a productive infection by X4LAI HIV-1 as evaluated from the release of p24gag in the culture supernatant, from detection of p24gag-positive cells with flow cytometry, and from production of complete reverse transcripts measured with real-time PCR.
What makes these four tissues different from the rest? It is not the presence of the CXCR4 coreceptor, since in tissues that did not support X4 HIV-1 replication CXCR4 was expressed on almost 90% of CD4+ T cells. A phenotypic analysis of the lymphocytic population in the four tissues that supported X4 HIV-1 replication revealed their enrichment in effector memory CD4 T cells of an early-differentiated phenotype (CD27+CD28+) associated with a smaller fraction of effector memory CD4 T cells of a late-differentiated phenotype (CD27−CD28−). Possibly, the cellular machinery of early-differentiated memory T cells is better equipped to support X4 HIV-1 replication than is that of more differentiated cells. Alternatively, a smaller fraction of late-differentiated CD4 T cells may be associated with a reduced production by these or other cells of X4 HIV-1-inhibitory soluble factors41. Although further studies are required to investigate the exact mechanisms by which the equilibrium between early- and late-differentiated effector memory CD4 T cells controls X4 HIV-1 productive infection, our results demonstrate that the susceptibility to X4 or R5 HIV-1 infection is not defined by the mere expression of specific coreceptors and can be restricted by innate immunological mechanisms operating in the human genital mucosa.
In summary, we analyzed the phenotypes of the lymphocytic populations in cervico-vaginal tissue and found that CD4 T cells, the major HIV-1 cell targets, are in this tissue mostly in an activated and differentiated state; also, the majority of CD4 T cells express CCR5, thus making this tissue particularly susceptible to R5 HIV-1 infection. We confirmed this in actual experiments on human cervico-vaginal tissue ex vivo, a system we further developed. We found that R5 HIV-1 replication causes activation of uninfected bystander CD4 T cells, resulting in expansion of the pool of preferentially infected cells. This mechanism can promote the local expansion of infection that precedes the establishment of a systemic infection.
Also, we found that in contrast to R5, replication of X4 HIV-1 was negligible except in a few tissues. In these tissues, a significant fraction of effector memory CD4 T cells coexpressed the markers CD27 and CD28, which characterize early-differentiated cells. Thus, it seems that in cervico-vaginal tissue expression of CXCR4 is not enough to support productive infection of X4 HIV-1. It has been suggested earlier that the “gatekeeping” mechanism is a superimposition of various incomplete restrictive barriers in the course of HIV-1 transmission, rather than a unique complete barrier4. Identification of the factors associated with restriction of X4 HIV-1 infection in cervico-vaginal tissue will reveal mechanisms for these “gatekeeping” barriers.
In conclusion, our results provide new insights into the relationship between HIV-1 infection and the activation/differentiation status of the lymphocytic population in cervico-vaginal tissue, thus contributing to a better understanding of the early events of sexual transmission of HIV-1. Such an understanding may lead to the development of new anti-HIV-1 strategies, in particular the expansion of the “gatekeeping” mechanisms to all HIV-1 variants and the disruption of the vicious circle of HIV-1-triggered T-cell activation that facilitates HIV-1 dissemination.
METHODS
Tissue histoculture and HIV-1 infection
Tonsillar tissues from routine tonsillectomy were obtained from the Children's National Medical Center (Washington, DC). Cervico-vaginal tissues were obtained from routine hysterectomy through the National Disease Research Interchange (Philadelphia, PA). All tissues were obtained according to an IRB-approved protocol. Tonsillar tissues were dissected into 2-mm3 blocks and digested for flow cytometry applications as described below. Culture of cervico-vaginal tissue was set up as previously described, with minor modifications20. Briefly, mucosal epithelium and underlying stroma of both ecto- and endo-cervix were separated from muscular tissue, dissected into approximately 2-mm3 blocks, and infected with the HIV-1 variant BaL (R5BaL) or LAI.04 (X4LAI.).
Both viral preparations were obtained in the Virology Quality Assurance Laboratory at Rush University (Chicago, IL). Viral stocks were obtained from the medium of PBMC cultures inoculated with either BaL (R5BaL) or LAI.04 (X4LAI.), originally received from the NIH AIDS Reagent Program. The p24 stock concentrations were 49±3 and 53±3 ng/ml for BaL (R5BaL) and LAI.04 (X4LAI.), respectively. Viral inocula were adjusted to obtain similar replication levels in lymphoid tissue 42.
For each experiment, 32 cervical blocks were transferred into two 1.5-mL conical tubes (16 blocks per tube), each containing 0.5 mL of viral stock. Simultaneously, lamivudine (3TC) at the dose of 5 µM was added to one of these tubes. After 2 hours of incubation at 37°C, tissue blocks were gently washed three times with phosphate-buffered saline (PBS), placed on top of a collagen sponge gel into a 12 well-plate (8 blocks/well), and cultured for 12 days, with a change of medium every third day. To the tissue blocks that were incubated with 3TC, the drug was added at each medium change.
Flow cytometry
Tonsillar and cervico-vaginal tissue blocks were digested with Collagenase IV (Invitrogen, Carlsbad, CA) at 5 mg/mL in RPMI 5% calf serum for 30 and 90 minutes, respectively. Single-cell suspensions were washed in staining buffer (PBS supplemented with 2% normal mouse serum; Gemini Bioproducts, West Sacramento, CA). To characterize tissue lymphocytes, cell suspensions were stained with different combinations of the monoclonal antibodies anti-CD3, anti-CD19, anti-CD20, anti-CD4, anti-CD8, anti-CD45RA, anti-CCR7, anti-CD27, anti-CD28, anti-CD25, anti-CD38, anti-CD57, anti-CD69, anti-CD95, anti-HLA-DR, anti-CCR5, and anti-CXCR4, coupled to a combination of the fluorochromes Qdot 605, Qdot 655, Pacific Blue, phycoerythrine (Pe), Pe-Cy5, Pe-Cy5.5, Pe-Cy7, APC, APC-Cy5.5, and APC-Cy7. To identify plasmacytoid dendritic cells (PDC) and myeloid dendritic cells (MDC), cell suspensions were stained with combinations of the monoclonal antibodies anti-HLA-DR (Pe-Cy5-labeled), anti- CD123 (Pe), and anti-CD11c (APC), as well as with FITC-labeled anti-CD3, anti-CD14, anti-CD16, anti-CD56, anti-CD19, and anti-CD20. We identified DCs as positive for HLA-DR but negative for CD3, CD14, CD16, CD56, CD19, and CD20; among this population we identified PDCs as CD123+ and MDCs as CD11c+. To identify macrophages, cell suspensions were stained with combinations of anti-CD14 (APC-Cy7-labeled) and FITC-labeled anti-CD3, anti-CD56, anti-CD19, and anti-CD20 monoclonal antibodies. Macrophages were identified as positive for CD14 and negative for CD3, CD56, CD19, and CD20. All the antibodies were purchased from Invitrogen and BD Biosciences (San Jose, CA).
After surface staining, cells were permeabilized with Fix&Perm™ (Invitrogen) and stained with an anti-HIV-1 p24gag monoclonal antibody (KC57) conjugate with fluorescein isothiocyanate (Beckman Coulter, Fullerton, CA). Data were acquired on an LSRII flow cytometer equipped with 355, 407, 488, 532, and 638 nm laser lines using DIVA 6.1.2 software and analyzed with FlowJo version 8.7.3 (Tree Star, Ashland, OR). We identified and excluded dead cells from the analysis using the LIVE/DEAD fixable Blue Dead Cell Stain™ kit (Invitrogen) and identified lymphocytes according to their light-scattering properties. Lymphocyte depletion was quantified by addition of TruCount™ beads (Invitrogen) according to the manufacturers’ instructions.
Dynamic immunofluorescent cytometric bead assay for HIV-1 p24gag quantification
We evaluated productive HIV-1 infection from measurements of HIV-1 antigen p24gag accumulated in tissue culture medium, using a dynamic immunofluorescent cytometric bead assay. The assay was performed as previously described43.
Real-time PCR for HIV-1 complete reverse transcription products
DNA was extracted from tissue blocks with a DNeasy tissue kit from QIAGEN (Valencia, CA). We performed real-time PCR to quantify complete reverse transcripts using the primer sets MH531 and MH532 (Butler SL et al., 2001) as previously described22. We performed amplification and detection on an Applied Biosystems Prism 7500 sequence detection system using a Taqman universal master mix (Invitrogen), 300 nM forward and reverse primer, 100 nM probe, and 100 ng of template DNA in a 25-µL reaction volume. We created a reference standard curve using 8E51 cells serially diluted. The amount of cellular DNA was assessed by use of erv-3 as a reference gene44.
Statistical analysis
Each datum point is the result of analysis of sets of 16 tissue blocks derived from each of n donors, where n is indicated in the text. The statistical significance of differences between data from various experimental groups was evaluated with the paired or unpaired Student's t test. Because the distribution of the numbers of HIV-1 complete reverse transcription products failed the Kolmogorov-Smirnov normality test, we used distribution-free nonparametric methods (Wilcoxon Match-Pairs Signed-Ranks Test and Mann-Whitney U test) to evaluate the significance of the differences between various experimental groups. However, when these data were log10-transformed, normality was achieved and we applied parametric methods (paired or unpaired Student's t test). The statistical significance of differences between data from various experimental groups evaluated after transformation was similar to that evaluated with nonparametric methods applied to non-transformed results. Depending on the type of statistical analysis, the pooled data are presented either as means ± standard error of the mean (SEM) or as medians and interquartile range (IQR). All of the hypothesis tests were two-tailed, and a p value of ≤ 0.05 defined statistical significance.
ACKNOWLEDGEMENTS
We thank the entire staff of the Department of Pathology of Children’s National Medical Center for their generous assistance in obtaining human tonsillar tissues. We thank Dr. Oliver Hartley for a generous gift of PSC-RANTES, Dr. Raymond Schinazi for providing us with lamivudine, and Dr. Barry Alpher for correcting the English style of our paper. This work was supported by the Intramural Research Program of the Eunice Kennedy-Shriver National Institute of Child Health and Human Development, National Institutes of Health.
REFERENCES
- 1.Haase AT. Perils at mucosal front lines for HIV and SIV and their hosts. Nat Rev Immunol. 2005;5:783–792. doi: 10.1038/nri1706. [DOI] [PubMed] [Google Scholar]
- 2.Pope M, Haase AT. Transmission, acute HIV-1 infection and the quest for strategies to prevent infection. Nat Med. 2003;9:847–852. doi: 10.1038/nm0703-847. [DOI] [PubMed] [Google Scholar]
- 3.Grivel JC, et al. HIV-1 pathogenesis differs in rectosigmoid and tonsillar tissues infected ex vivo with CCR5- and CXCR4-tropic HIV-1. Aids. 2007;21:1263–1272. doi: 10.1097/QAD.0b013e3281864667. [DOI] [PubMed] [Google Scholar]
- 4.Margolis L, Shattock R. Selective transmission of CCR5-utilizing HIV-1: the 'gatekeeper' problem resolved? Nat Rev Microbiol. 2006;4:312–317. doi: 10.1038/nrmicro1387. [DOI] [PubMed] [Google Scholar]
- 5.Salazar-Gonzalez JF, et al. Genetic identity, biological phenotype, and evolutionary pathways of transmitted/founder viruses in acute and early HIV-1 infection. J Exp Med. 2009;206:1273–1289. doi: 10.1084/jem.20090378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Stevceva L, et al. Cervicovaginal lamina propria lymphocytes: phenotypic characterization and their importance in cytotoxic T-lymphocyte responses to simian immunodeficiency virus SIVmac251. J Virol. 2002;76:9–18. doi: 10.1128/JVI.76.1.9-18.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Broliden K, Haase AT, Ahuja SK, Shearer GM, Andersson J. Introduction: Back to basics: mucosal immunity and novel HIV vaccine concepts. J Intern Med. 2009;265:5–17. doi: 10.1111/j.1365-2796.2008.02053.x. [DOI] [PubMed] [Google Scholar]
- 8.Cavrois M, Neidleman J, Kreisberg JF, Greene WC. In vitro derived dendritic cells trans-infect CD4 T cells primarily with surface-bound HIV-1 virions. PLoS Pathog. 2007;3:e4. doi: 10.1371/journal.ppat.0030004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Bouschbacher M, et al. Early events in HIV transmission through a human reconstructed vaginal mucosa. Aids. 2008;22:1257–1266. doi: 10.1097/QAD.0b013e3282f736f4. [DOI] [PubMed] [Google Scholar]
- 10.Hladik F, Hope TJ. HIV infection of the genital mucosa in women. Curr HIV/AIDS Rep. 2009;6:20–28. doi: 10.1007/s11904-009-0004-1. [DOI] [PubMed] [Google Scholar]
- 11.Turville SG, Peretti S, Pope M. Lymphocyte-dendritic cell interactions and mucosal acquisition of SIV/HIV infection. Curr Opin HIV AIDS. 2006;1:3–9. doi: 10.1097/01.COH.0000194109.14601.20. [DOI] [PubMed] [Google Scholar]
- 12.Fletcher PS, et al. Ex vivo culture of human colorectal tissue for the evaluation of candidate microbicides. AIDS. 2006;20:1237–1245. doi: 10.1097/01.aids.0000232230.96134.80. [DOI] [PubMed] [Google Scholar]
- 13.Johansson EL, Rudin A, Wassen L, Holmgren J. Distribution of lymphocytes and adhesion molecules in human cervix and vagina. Immunology. 1999;96:272–277. doi: 10.1046/j.1365-2567.1999.00675.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Sallusto F, Geginat J, Lanzavecchia A. Central memory and effector memory T cell subsets: function, generation, and maintenance. Annu Rev Immunol. 2004;22:745–763. doi: 10.1146/annurev.immunol.22.012703.104702. [DOI] [PubMed] [Google Scholar]
- 15.Okada R, Kondo T, Matsuki F, Takata H, Takiguchi M. Phenotypic classification of human CD4+ T cell subsets and their differentiation. Int Immunol. 2008;20:1189–1199. doi: 10.1093/intimm/dxn075. [DOI] [PubMed] [Google Scholar]
- 16.Appay V, van Lier RA, Sallusto F, Roederer M. Phenotype and function of human T lymphocyte subsets: consensus and issues. Cytometry A. 2008;73:975–983. doi: 10.1002/cyto.a.20643. [DOI] [PubMed] [Google Scholar]
- 17.Biancotto A, et al. HIV-1 induced activation of CD4+ T cells creates new targets for HIV-1 infection in human lymphoid tissue ex vivo. Blood. 2008;111:699–704. doi: 10.1182/blood-2007-05-088435. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Hoffman RM. Three-dimensional histoculture: origins and applications in cancer research. Cancer Cells. 1991;3:86–92. [PubMed] [Google Scholar]
- 19.Glushakova S, Baibakov B, Margolis LB, Zimmerberg J. Infection of human tonsil histocultures: a model for HIV pathogenesis. Nat Med. 1995;1:1320–1322. doi: 10.1038/nm1295-1320. [DOI] [PubMed] [Google Scholar]
- 20.Grivel JC, Margolis L. Use of human tissue explants to study human infectious agents. Nat Protoc. 2009;4:256–269. doi: 10.1038/nprot.2008.245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Kawamura T, et al. PSC-RANTES blocks R5 human immunodeficiency virus infection of Langerhans cells isolated from individuals with a variety of CCR5 diplotypes. J Virol. 2004;78:7602–7609. doi: 10.1128/JVI.78.14.7602-7609.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Biancotto A, et al. Dual role of prostratin in inhibition of infection and reactivation of human immunodeficiency virus from latency in primary blood lymphocytes and lymphoid tissue. J Virol. 2004;78:10507–10515. doi: 10.1128/JVI.78.19.10507-10515.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Karlsson I, et al. Differential pathogenesis of primary CCR5-using human immunodeficiency virus type 1 isolates in ex vivo human lymphoid tissue. J Virol. 2005;79:11151–11160. doi: 10.1128/JVI.79.17.11151-11160.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Miller CJ, Shattock RJ. Target cells in vaginal HIV transmission. Microbes Infect. 2003;5:59–67. doi: 10.1016/s1286-4579(02)00056-4. [DOI] [PubMed] [Google Scholar]
- 25.Li Q, et al. Glycerol monolaurate prevents mucosal SIV transmission. Nature. 2009;458:1034–1038. doi: 10.1038/nature07831. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Hladik F, et al. Initial events in establishing vaginal entry and infection by human immunodeficiency virus type-1. Immunity. 2007;26:257–270. doi: 10.1016/j.immuni.2007.01.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Hu J, Gardner MB, Miller CJ. Simian immunodeficiency virus rapidly penetrates the cervicovaginal mucosa after intravaginal inoculation and infects intraepithelial dendritic cells. J Virol. 2000;74:6087–6095. doi: 10.1128/jvi.74.13.6087-6095.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Zhang Z, et al. Sexual transmission and propagation of SIV and HIV in resting and activated CD4+ T cells. Science. 1999;286:1353–1357. doi: 10.1126/science.286.5443.1353. [DOI] [PubMed] [Google Scholar]
- 29.Spira AI, et al. Cellular targets of infection and route of viral dissemination after an intravaginal inoculation of simian immunodeficiency virus into rhesus macaques. J Exp Med. 1996;183:215–225. doi: 10.1084/jem.183.1.215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Dai J, et al. Human immunodeficiency virus integrates directly into naive resting CD4+ T cells but enters naive cells less efficiently than memory cells. J Virol. 2009;83:4528–4537. doi: 10.1128/JVI.01910-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Picker LJ. Immunopathogenesis of acute AIDS virus infection. Curr Opin Immunol. 2006;18:399–405. doi: 10.1016/j.coi.2006.05.001. [DOI] [PubMed] [Google Scholar]
- 32.Greenhead P, et al. Parameters of human immunodeficiency virus infection of human cervical tissue and inhibition by vaginal virucides. J Virol. 2000;74:5577–5586. doi: 10.1128/jvi.74.12.5577-5586.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Cummins JE, Jr, et al. Preclinical testing of candidate topical microbicides for anti-human immunodeficiency virus type 1 activity and tissue toxicity in a human cervical explant culture. Antimicrob Agents Chemother. 2007;51:1770–1779. doi: 10.1128/AAC.01129-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Biancotto A, et al. Abnormal activation and cytokine spectra in lymph nodes of people chronically infected with HIV-1. Blood. 2007;109:4272–4279. doi: 10.1182/blood-2006-11-055764. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Estes J, et al. Collagen deposition limits immune reconstitution in the gut. J Infect Dis. 2008;198:456–464. doi: 10.1086/590112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Barbour JD, et al. High CD8+ T cell activation marks a less differentiated HIV-1 specific CD8+ T cell response that is not altered by suppression of viral replication. PLoS One. 2009;4:e4408. doi: 10.1371/journal.pone.0004408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Reinhart TA. Chemokine induction by HIV-1: recruitment to the cause. Trends Immunol. 2003;24:351–353. doi: 10.1016/s1471-4906(03)00163-7. [DOI] [PubMed] [Google Scholar]
- 38.Stacey AR, et al. Induction of a striking systemic cytokine cascade prior to peak viremia in acute human immunodeficiency virus type 1 infection, in contrast to more modest and delayed responses in acute hepatitis B and C virus infections. J Virol. 2009;83:3719–3733. doi: 10.1128/JVI.01844-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Gomez C, Hope TJ. The ins and outs of HIV replication. Cell Microbiol. 2005;7:621–626. doi: 10.1111/j.1462-5822.2005.00516.x. [DOI] [PubMed] [Google Scholar]
- 40.Perez O, Hope TJ. Cellular restriction factors affecting the early stages of HIV replication. Curr HIV/AIDS Rep. 2006;3:20–25. doi: 10.1007/s11904-006-0004-3. [DOI] [PubMed] [Google Scholar]
- 41.Mikulak J, et al. Biological and physical characterization of the X4 HIV-1 suppressive factor secreted by LPS-stimulated human macrophages. Virology. 2009;390:37–44. doi: 10.1016/j.virol.2009.04.005. [DOI] [PubMed] [Google Scholar]
- 42.Glushakova S, Baibakov B, Zimmerberg J, Margolis LB. Experimental HIV infection of human lymphoid tissue: correlation of CD4+ T cell depletion and virus syncytium-inducing/non-syncytium-inducing phenotype in histocultures inoculated with laboratory strains and patient isolates of HIV type 1. AIDS Res Hum Retroviruses. 1997;13:461–471. doi: 10.1089/aid.1997.13.461. [DOI] [PubMed] [Google Scholar]
- 43.Biancotto A, et al. A highly sensitive and dynamic immunofluorescent cytometric bead assay for the detection of HIV-1 p24. J Virol Methods. 2009;157:98–101. doi: 10.1016/j.jviromet.2008.11.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Lisco A, et al. Acyclovir is activated into a HIV-1 reverse transcriptase inhibitor in herpesvirus-infected human tissues. Cell Host Microbe. 2008;4:260–270. doi: 10.1016/j.chom.2008.07.008. [DOI] [PMC free article] [PubMed] [Google Scholar]