

Abstract
Objective
Generalized social phobia (GSP) is defined by a persistent fear of social disapproval. However, the neural underpinnings of this increased fear and its mediating factors are unclear. Using event-related fMRI, we examined whether the intent of an event, which mediates the neural response to social disapproval in healthy individuals, differentially affects response in GSP. Specifically, would patients with GSP show particularly increased response to embarrassing, unintentional transgressions?
Method
Sixteen patients with GSP and sixteen age, IQ, and gender matched healthy individuals read stories during fMRI scans that either involved neutral social events, unintentional social transgressions (e.g., choking on food at party, and coughing it up), or intentional social transgressions (e.g., disliking food at party, and spitting it out).
Results
Significant group-by-transgression interactions were observed within ventral regions of medial prefrontal cortex (MPFC). Healthy individuals tended to show increased BOLD responses to intentional, relative to unintentional transgressions. The patients with GSP, however, showed significantly increased responses to the unintentional transgressions. In addition, they rated the unintentional transgressions as significantly more embarrassing than the comparison individuals. We also observed significant group main effects within the amygdala and bilateral insula, reflecting elevated GSP responses within these regions to all event types.
Conclusions
These results further implicate the MPFC in the pathophysiology of GSP, specifically through its involvement in distorted self-referential processing. In addition, the current results further underscore the extended role of the amygdala and insula in the processing of social stimuli more generally in GSP.
INTRODUCTION
Generalized Social Phobia (GSP) typically manifests in early adolescence and predicts risk for depression, substance abuse, and suicide (1-3). At the neural-network level, fMRI studies of GSP find atypically increased responses to social stimuli within the amygdala (4-7). In addition, recent studies also find atypical activity within medial prefrontal cortex (MPFC) regions previously implicated in self-relevant processing (8-11).
Self-referential processing plays a critical role in regulating interpersonal behavior. Self-referential processing occurs following social transgressions which result in harmless norm violations (e.g., spitting out food at a dinner party). An individual’s response to such transgressions reflects their social calculations related to self-reference (i.e., how will observers react to my behavior?), and GSP involves perturbations in such social calculations. These perturbations manifest as heightened fear of social disapproval. Indeed, patients with GSP show anomalous neural (8, 11) and autonomic (12) responses to the receipt of verbal criticism and during anticipation of negative peer evaluations (4). Moreover, social anxiety is related to increased levels of self-criticism (13, 14).
No prior fMRI study of GSP examines the response to actions that bring social disapproval. A variable modulating social disapproval is the transgressor’s presumed intent (15). Unintentional conventional transgressions (e.g., accidentally choking and consequently spitting out the host’s food) lead to displays of embarrassment that, it is argued, serve to indicate the absence of intent and a desire for appeasement (16). In contrast, intentional transgressions (e.g., spitting out the host’s unpalatable food) are not associated with embarrassment or other signs of appeasement. Rather, behavior in this situation reflects the transgressor’s intent to challenge the social hierarchy (15).
Previous fMRI work with healthy adults has implicated ventral MPFC and temporo-parietal regions in the processing of conventional transgressions (15, 17, 18). These are regions implicated in self-referential processing and representations of the intentional states of others (19-21). For healthy individuals, this region responds to the occurrence of intentional, relative to unintentional, conventional social transgressions (15). This is thought to reflect heightened representation of the protagonists’ intent to challenge the social order (15, 18).
GSP involves marked and persistent fear of embarrassment (22), which might lead GSP patients to frequently seek appeasement. A tendency to fear embarrassment and seek appeasement in GSP might be reflected in enhanced neural responses specifically to unintentional, potentially embarrassing, social transgressions (cf. 23). The current study tests this prediction using an adaptation of Berthoz et al. (2006), whereby intentional transgressions, unintentional transgressions, and normative vignettes are presented to GSP patients and comparison individuals. Given GSP-related heightened propensity for embarrassment, coupled with prior imaging work in GSP, we expect unintentional social transgressions, specifically, when presented to patients, to elicit increased activity in MPFC regions implicated in social cognition (8, 10, 11).
Of note, the amygdala and insula also appear hyper-responsive in GSP to a range of negative social stimuli. However, unlike for the MPFC, this hyper-sensitivity manifests in many contexts (e.g., 6-8). Thus, we also expected patients with GSP to show increased responding in these areas. However, unlike the MPFC, which might precisely code context for social transgressions, we expect the amygdala and insula to show hyper-responsivity to social stimuli, regardless of context. This would be reflected in a main effect of “group,” in the absence of “group-by-condition” interactions. This study tests these predictions.
METHODS AND MATERIALS
Subjects
This study included 16 patients with GSP and 16 healthy comparison (HC) individuals, group-matched on age, gender, and IQ (see Table 1). Subjects were recruited from NIMH Institutional-Review-Board (IRB) approved advertisements.
Table 1.
Subject Characteristics S.D. in Brackets ().
| Patients with GSP (N = 16) | Healthy subjects (N = 16) | P < | |
|---|---|---|---|
| Age | 35.1 (9.60) | 30.0 (8.37) | ns |
| Gender | 9 M/ 7 F | 7 M/ 9 F | ns |
| Race | |||
| Caucasian | 13 | 14 | ns |
| African-American | 3 | 2 | |
| IQ | 118.6 (8.54) | 117.2 (9.74) | ns |
| LSAS-SR | 67.1 (23.43) | 16.2 (11.65) | 0.001 |
| IDS | 11.0 (7.20) | 5.1 (3.50) | 0.05 |
| GAF | 61.3 (5.34) | - | - |
M = Male; F = Female; LSAS = Liebowitz Social Anxiety Scale; IDS = Inventory of Depressive Symptomatology; GAF = Global Assessment of Functioning.
Subjects with GSP had to meet criteria for generalized social phobia according to the DSM-IV (1994) criteria based on the Structural Clinical interview for DSM-IV Axis I disorders (SCID) (24) and a confirmatory clinical interview by a board-certified psychiatrist (DSP). No GSP patient had another Axis-1 diagnosis; all were currently medication-free 6+ months. HCs were excluded if they had a history of any psychiatric illness. All subjects were in good physical health, as confirmed by a complete physical exam, and provided written informed consent.
Further, as part of the assessment, all subjects completed the Liebowitz Social Anxiety Scale - Self Report (LSAS-SR), and the Inventory of Depressive Symptomatology – Self Report (IDS-SR). In addition, for the patients with GSP, the level of overall social, occupational and psychological functioning was assessed by the Global Assessment of Functioning (GAF). Scores on the these measures characterized the GSP group as having moderate levels of social anxiety with mild/ some associated impairment in functioning; Table 1.
Behavioral Task
Subjects read stories (e.g., Joanna is invited for dinner at a friend’s house, she has a bite of the first course, chews…) that could either involve a neutral social event (… and swallows the food), an unintentional transgression (…chokes and coughs up the food), or an intentional transgression (…dislikes it and spits out the food). Twenty-six different stem stories were used, each presented with the three different type endings. Thus, a total of 78 endings were used. The three different type endings were matched on number of letters and words. Moreover, care was taken to ensure that the framing of the comments was consistent across the three type endings. Prior to scanning, subjects were told that they would read different stories, and to imagine what they would feel like if they were in the situation described. For each story, subjects were simply required to press a button with their left hand when they had read the story. Each stem comment was presented on its own for 3000 ms. The ending of the story would then appear underneath the stem, and the stem + ending (full story) would be shown on the screen together for 6000 ms. In addition, for each experimental run, twenty-two 3000 ms fixation points were presented between the stimuli (five at beginning of run, five at end of run and twelve randomized throughout the run), serving as an implicit baseline. Subjects completed three randomly-presented runs.
Following EPI acquisition, subjects rated each of the 78 individual stories, presented in a randomized order across subjects, on a 5-point Likert scale according to how embarrassing they thought the behaviors were, where 1 = not at all embarrassing; 3 = somewhat embarrassing; and 5 = extremely embarrassing. In addition, they rated the stories according to how inappropriate they thought the behaviors were, where 1 = not at all inappropriate; 3 = somewhat inappropriate; 5 = extremely inappropriate.
fMRI Parameters
Whole-brain blood oxygen level dependent (BOLD) fMRI data were acquired using a 1.5 Tesla GE MRI scanner. Following sagital localization, functional T2* weighted images were acquired using an echo-planar single-shot gradient echo pulse sequence (matrix = 64 × 64 mm, repetition time (TR) = 3000 ms, echo time (TE) = 30 ms, field-of-view (FOV) = 240 mm (3.75 × 3.75 × 4 mm voxels). Images were acquired in 31 contiguous 4 mm axial slices per brain volume, with each run lasting 4 minutes 54 seconds. In the same session, a high-resolution T1-weighed anatomical image was acquired to aid with spatial normalization (three-dimensional Spoiled GRASS; TR = 8.1 ms; TE = 3.2 ms, flip angle = 20°; FOV = 240 mm, 124 axial slices, thickness = 1.0 mm; 256 × 256 acquisition matrix).
Data were analyzed within the framework of the general linear model using Analysis of Functional Neuroimages (AFNI) (25). Both individual and group-level analyses were conducted. The first four volumes in each scan series, collected before equilibrium magnetization was reached, were discarded. Motion correction was performed by registering all volumes in the EPI dataset to a volume collected close to acquisition of the high resolution anatomical dataset.
The EPI datasets for each subject were spatially smoothed (isotropic 6mm kernel) to reduce variability among individuals and generate group maps. Next, the time series data were normalized by dividing the signal intensity of a voxel at each time point by the mean signal intensity of that voxel for each run and multiplying the result by 100, producing regression coefficients representing percent-signal change. Regressors for the three comment categories (Transgression: Intentional, Unintentional, None) were created by convolving the train of stimulus events with a gamma-variate haemodynamic response function. Linear regression modeling was performed using these regressors plus regressors for a first-order baseline drift function. This produced for each voxel and each regressor, a beta coefficient and its associated t-statistic.
Voxel-wise group analyses involved transforming single subject beta coefficients into the standard coordinate space of Talairach and Tournoux (26). Subsequently, a 2 (Group: GSP, HC) by 2 (Transgression: Intentional, Unintentional, None) ANOVA was performed to produce statistical maps of the main effect of group and transgression and group-by-transgression interaction (p < 0.005). To correct for multiple comparisons for the whole-brain analysis at p < 0.005, we performed a spatial clustering operation using AlphaSim (http://afni.nimh.nih.gov/pub/dist/doc/manual/AlphaSim.pdf) with 1,000 Monte Carlo simulations taking into account the entire EPI matrix. This procedure yielded a minimum cluster size with a map-wise false-positive probability of p < 0.05, corrected for multiple comparisons. We report results primarily for brain regions that survived this whole-brain correction procedure. Nevertheless, prior results for specific brain regions generate regionally-specific a priori hypotheses. Accordingly, in secondary analyses, we also report results using a lower threshold for the insula and MPFC. Precise statistical results, with correction for multiple comparisons, are clearly indicated in Table 3.
Table 3.
Significant areas of activation for the group-by-transgression interaction †
| REGION | BA | Mm3 | X | Y | Z | F-value |
|---|---|---|---|---|---|---|
| Group-by-transgression interaction | ||||||
| L ventromedial prefrontal gyrus | 10 | 1008 | -7 | 59 | 13 | 8.74 |
| R medial prefrontal gyrus* | 9 | 364 | 5 | 58 | 38 | 7.74 |
| Group main effect (GSP > HC for all) | ||||||
| L ventromedial prefrontal gyrus | 10 | 12603 | -13 | 60 | 16 | 33.90 |
| L dorsomedial prefrontal gyrus* | 9 | 644 | -23 | 32 | 34 | 19.32 |
| L insula* | 13 | 623 | -37 | -14 | 0 | 18.23 |
| R insula* | 13 | 305 | 26 | 16 | -6 | 13.67 |
| R amygdala/ parahippocampal gyrus | 1088 | 41 | -8 | -18 | 14.07 |
All activations are effects observed in whole brain analyses significant at p < 0.005 corrected for multiple comparisons (significant at p < 0.05) except
significant at p < 0.005 uncorrected.
After observing hypothesized group differences, post-hoc analyses were performed to facilitate interpretations. For these analyses, average percent signal change was measured across all voxels within each ROI generated from the functional mask, and data for main effects and interactions were unpacked and analyzed using appropriate follow-up tests, principally one-way repeated measures ANOVAs, within SPSS.
RESULTS
Behavioral Ratings Data
The behavioral data pertaining to the ratings of embarrassment and inappropriateness were analyzed using two separate 2 (Group: GSP, HC) by 2 (Transgression: Intentional, Unintentional, None) ANOVAs. Subsequent one-way repeated measures ANOVAs were used to unpack significant interactions. Here we first consider the embarrassment ratings. There was a significant group-by-transgression interaction (F = 3.15; p < 0.05); whereas the patients with GSP rated the unintentional transgressions as being significantly more embarrassing compared to the healthy comparison individuals (F = 11.32; p < 0.005), the two groups did not differ significantly in their ratings of neutral or intentional behaviors (F = 1.62 & 0.90 respectively; ns); see Table 2). There was also a significant main effect of group; overall the patients with GSP rated the behaviors as significantly more embarrassing compared to the healthy comparison individuals (F = 4.26; p < 0.05).
Table 2.
Embarrassment and inappropriateness ratings for the three transgression types. S.E. in Brackets ().
| Transgression | Embarrassment | Inappropriateness | ||
|---|---|---|---|---|
| GSP | HC | GSP | HC | |
| Unintentional | 3.48 (0.14) | 2.81 (0.14) | 2.71 (0.19) | 2.02 (0.19) |
| Intentional | 3.20 (0.26) | 2.90 (0.20) | 4.40 (0.13) | 3.92 (0.15) |
| None (Neutral) | 1.13 (0.03) | 1.15 (0.03) | 1.15 (0.03) | 1.09 (0.03) |
There was also a significant group-by-transgression interaction for the inappropriateness ratings (F = 3.50; p < 0.05); the patients with GSP rated the unintentional and intentional transgressions as being significantly more inappropriate compared to the healthy comparison individuals (F = 6.57 & 5.85 respectively; p < 0.05), however the two groups did not differ significantly in their ratings of neutral behaviors (F = 1.42; ns); Table 2. As with the embarrassment ratings, there once again was a significant main effect of group; overall the patients with GSP rated the behaviors as significantly more inappropriate compared to the healthy comparison individuals (F = 9.32; p < 0.05).
EPI Data
The BOLD response data were analyzed by a 2 (Group: GSP, HC) by 3 (Transgression: Intentional, Unintentional, None) ANOVA. Our goal was to determine whether patients with GSP show a particularly increased activation to unintentional (embarrassing) transgressions relative to comparison individuals. Two statistical maps were critical to our predictions (group-by-transgression and group) and will be discussed in turn; see Table 3.
The analysis for group-by-transgression interactions identified two regions: the left ventral MPFC and a small region of more dorsal right MPFC (this second region did not survive correction for multiple comparisons); see Table 3. In line with predictions, patients with GSP showed significantly greater BOLD responses in both regions to unintentional relative to intentional transgressions (F = 12.80 & 6.43; p < 0.005 & 0.05 respectively). In contrast, the healthy comparison individuals showed a trend in both regions for significantly greater activation to intentional relative to unintentional transgressions (F = 3.29 & 3.00; p = 0.09 & 0.10 respectively), as well as an overall reduction in response to stories. Importantly, group differences in activation were particularly marked for unintentional transgressions (F = 20.90; p < 0.001, compared to F = 14.36 & 5.67; p < 0.005 and 0.05 for intentional and neutral transgressions respectively); see Figure 1.
Figure 1.
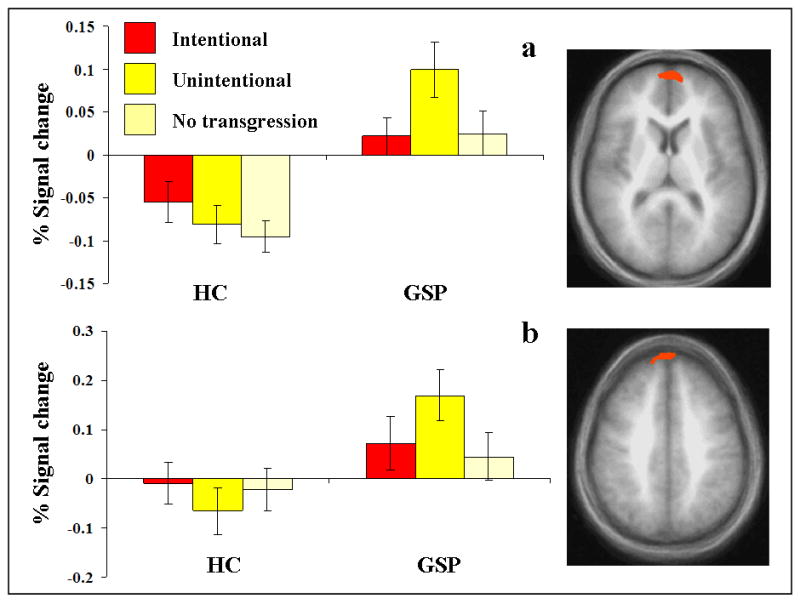
Interactions of group-by-transgression. BOLD responses within (a) left MPFC (-7, 59, 13), and (b) right MPFC (5, 58, 38) to behaviors involving intentional, unintentional and no social transgressions for the two groups.
It is notable that there were also significant main effects of group within both a large region of ventral MPFC and a slightly lateral region of dorsal MPFC as well as bilateral insula and the right amygdala; see Table 3. Within all these regions, patients with GSP showed significantly greater BOLD responses across transgressions relative to the healthy comparison individuals (F range 12.50 to 32.69; p < 0.001 for all).
MDD was an exclusion criteria for participation in this study. However, patients with GSP did report a higher level of depressive symptoms on the IDS-SR relative to the healthy comparison individuals (F = 8.70; p < 0.05; Table 1). To exclude the possibility that the higher level of depression scores in the patients with GSP influenced results, we used depression scores as a covariate in the follow-up analyses on the extracted average percent signal change from the two main regions of interests, the MPFC regions identified by the group-by-transgression interaction. Importantly, this inclusion of depression scores as a covariate did not change the significant results (F = 5.28 & 5.77; p = 0.008 & 0.005 respectively), suggesting that level of depression did not significantly contribute towards the current results.
Correlational Analysis
Using correlational analysis, we examined whether the increased responses to unintentional transgressions within MPFC in GSP related to severity of symptomatology. Specifically, we tested whether there was a significant relationship between level of social anxiety symptoms, as indexed by the LSAS-SR and the activation to unintentional transgression within the two MPFC regions identified by the group-by-transgressions interaction. There was a trend towards a significant positive correlation for one of the two MPFC regions. However, neither test reached two-tailed significance (Pearson’s r= 0.48 & 0.19; p = 0.059 & 0.487 respectively).
DISCUSSION
In this study we examined the neural responses in GSP to intentional and unintentional, embarrassing social transgressions. In line with predictions, we observed a significant group-by-transgression interaction in both the behavioural and MPFC neural response patterns. Patients with GSP rated unintentional, but not intentional, transgressions as significantly more embarrassing than the healthy comparison individuals. Moreover, healthy individuals showed greater activation to intentional relative to unintentional transgressions within ventral MPFC, consistent with previous research (15). Patients with GSP, in contrast, showed the opposite: significantly greater activation to unintentional, relative to intentional transgressions. In addition, and again in line with predictions, patients with GSP showed increased responses, relative to the comparison individuals, within an extensive region of ventral MPFC as well as the amygdala and bilateral insula to the social transgressions regardless whether they were intentional or unintentional.
Work on MPFC with respect to social cognition suggests a functional distinction between ventral and more dorsal MPFC. It has been argued that ventral MPFC is particularly implicated in self-referential processing; i.e., the processing of stimuli that are experienced as strongly related to one’s own person (27). In contrast, dorsal MPFC has been implicated in the representation of the mental states of others (20, 28-34). It is notable that in the current study patients with GSP showed an atypically increased response to unintentional, embarrassing social transgressions within ventral MPFC, the region particularly implicated in self-referential processing (it should be noted though that there was also a significant activation within a more dorsal region of MPFC, although this region did not survive correction for multiple comparisons). We would suggest that this is because they judge such transgressions as significantly more self-relevant than healthy individuals. Indeed, healthy individuals show a very different pattern of activity within this region, showing in this and previous work (15) greater activity to intentional rather than unintentional transgressions. It is argued that intentional transgressions have greater significance for healthy individuals and lead to generally increased activity in regions engaged in social cognition. This is because a social transgression, unmarked by embarrassment and instead demonstrating intent, marks a challenge to the social hierarchy. As such, we speculate that these results together with those of Blair et al. (11), indicate a fundamental reorganization in self-relevant processing in patients with GSP. Evaluations of the self in GSP primarily focus on potentially embarrassing events. In healthy individuals, in contrast, evaluations of the self focus more closely on potential status challenges. Importantly, these data indicate that the pathophysiology of GSP has, perhaps developmentally, extended beyond a heightened amygdala response to social threat.
These findings might meaningfully inform clinicians’ attempts to understand socially anxious patients’ concerns and generate ideas on novel therapies. For example, the current findings delineate an information-processing bias that forces socially-anxious individuals to struggle when evaluating the significance of unintentional, potentially embarrassing acts. This difficulty is associated with perturbations in the prefrontal cortex, a brain region that enables flexible assignment of meaning to events. As such, clinicians might help patients to understand their problems as related to difficulty with flexible thought. In doing so, the clinician might work with the patient to devise alternative information-processing strategies that more accurately evaluate the significance of embarrassing acts. Finally, prior work on other forms of anxiety-related information-processing biases generates ideas on novel treatments, which emerge as targets of the underlying neural substrate supporting particularly information-processing biases. Thus, as knowledge increases on the precise neural-system that support the social-attribution biases found in the current study, this knowledge also might generate novel insights on strategies for changing function in this neural system, leading to novel therapies.
In addition to the significant group-by-transgression interaction within ventral MPFC there was a highly significant main effect of group within a large region of ventral MPFC, as well as a slightly lateral region of dorsal MPFC together with bilateral insula and the amygdala. The amygdala and the insula have been frequently identified in imaging studies involving social phobia (7), suggesting that hyper-responsiveness within those regions in GSP is not specific to the anomalous processes underlying self-referential processing in GSP, but rather extends to the processing of social stimuli more generally.
One aspect of data from the current study should be evaluated particularly carefully and might suggest avenues for future work. Namely, for some brain regions, the presence of a group effect, in the absence of a group-by-condition interaction, reflects increased responsiveness in GSP to all the social stimuli, regardless of their meaning, context, or intent. As such, findings in the current study differentiate regions showing greater responsiveness in patients with GSP to social stimuli more generally (e.g., the amygdala and insula), relative to regions showing a greater responsiveness to social stimuli occurring specifically in self referential contexts (e.g., MPFC). Nevertheless, the current design only allows an initial comparison of findings across these brain regions, to a relatively restricted class of event types. Future work might employ larger collections of stimuli extending across broader classes of events, including non-social control stimuli. By including such stimuli, such future work could evaluate the possibility that GSP involves heightened generalized responsiveness, in some regions, to any form of emotionally salient stimulus, including both non-social and social stimuli.
In summary, we found that within ventral MPFC, and in contrast to the healthy comparison individuals, the patients with GSP showed increased responses to unintentional (embarrassing-mediated), relative to intentional, transgressions. Behaviorally, patients with GSP showed particularly elevated embarrassment ratings for the unintentional transgressions relative to comparison adults. These results underscore the importance of ventral MPFC as part of the pathophysiology of GSP and, we speculate, may relate to distorted self referential processing in this disorder. In addition, we observed increased amygdala and insula activation across both intentional and unintentional transgressions in the patients with GSP, further implicating those two regions in the pathophysiology of GSP.
Acknowledgments
This research was supported by the Intramural Research Program of the NIH: NIMH.
Footnotes
The authors have no conflicts of interest or financial disclosures to report.
REFERENCE LIST
- 1.Kaufman J, Charney D. Comorbidity of mood and anxiety disorders. Depress Anxiety. 2000;12(Suppl 1):69–76. doi: 10.1002/1520-6394(2000)12:1+<69::AID-DA9>3.0.CO;2-K. [DOI] [PubMed] [Google Scholar]
- 2.Kessler RC. The impairments caused by social phobia in the general population: implications for intervention. Acta Psychiatr Scand Suppl. 2003;(417):19–27. doi: 10.1034/j.1600-0447.108.s417.2.x. [DOI] [PubMed] [Google Scholar]
- 3.Beesdo K, Bittner A, Pine DS, Stein MB, Hofler M, Lieb R, Wittchen HU. Incidence of social anxiety disorder and the consistent risk for secondary depression in the first three decades of life. Arch Gen Psychiatry. 2007;64(8):903–12. doi: 10.1001/archpsyc.64.8.903. [DOI] [PubMed] [Google Scholar]
- 4.Guyer AE, Lau JY, McClure-Tone EB, Parrish J, Shiffrin ND, Reynolds RC, Chen G, Blair RJ, Leibenluft E, Fox NA, Ernst M, Pine DS, Nelson EE. Amygdala and ventrolateral prefrontal cortex function during anticipated peer evaluation in pediatric social anxiety. Arch Gen Psychiatry. 2008;65(11):1303–12. doi: 10.1001/archpsyc.65.11.1303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Blair KS, Shaywitz J, Smith BW, Rhodes R, Geraci M, Jones M, McCaffrey D, Vythilingam M, Finger E, Mondillo K, Jacobs M, Charney D, Blair RJR, Drevets WC, Pine DS. Response to emotional expressions in Generalized Social Phobia (GSP) and Generalized Anxiety Disorder (GAD): Evidence for separate disorders. Am J Psychiatry. 2008;165(9):1193–202. doi: 10.1176/appi.ajp.2008.07071060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Stein MB, Goldin PR, Sareen J, Zorrilla LT, Brown GG. Increased amygdala activation to angry and contemptuous faces in generalized social phobia. Archives of general psychiatry. 2002;59(11):1027–34. doi: 10.1001/archpsyc.59.11.1027. [DOI] [PubMed] [Google Scholar]
- 7.Etkin A, Wager TD. Functional neuroimaging of anxiety: a meta-analysis of emotional processing in PTSD, social anxiety disorder, and specific phobia. Am J Psychiatry. 2007;164(10):1476–88. doi: 10.1176/appi.ajp.2007.07030504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Blair KS, Geraci M, Devido J, McCaffrey D, Chen G, Vythilingam M, Ng P, Hollon N, Jones M, Blair RJ, Pine DS. Neural response to self- and other referential praise and criticism in generalized social phobia. Archives of General Psychiatry. 2008;65(10):1176–84. doi: 10.1001/archpsyc.65.10.1176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Sripada CS, Angstadt M, Banks S, Nathan PJ, Liberzon I, Phan KL. Functional neuroimaging of mentalizing during the trust game in social anxiety disorder. Neuroreport. 2009;20(11):984–9. doi: 10.1097/WNR.0b013e32832d0a67. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Goldin PR, Manber-Ball T, Werner K, Heimberg R, Gross JJ. Neural Mechanisms of Cognitive Reappraisal of Negative Self-Beliefs in Social Anxiety Disorder. Biol Psychiatry. 2009 doi: 10.1016/j.biopsych.2009.07.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Blair KS, Geraci M, Majestic K, Odenheimer S, Jacobs M, Blair KS, Pine DS. Atypical modulation of medial prefrontal cortex to self-referential comments in Generalized Social Phobia. doi: 10.1016/j.pscychresns.2010.12.016. submitted. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Lissek S, Levenson J, Biggs AL, Johnson LL, Ameli R, Pine DS, Grillon C. Elevated fear conditioning to socially relevant unconditioned stimuli in social anxiety disorder. Am J Psychiatry. 2008;165(1):124–32. doi: 10.1176/appi.ajp.2007.06091513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Cox BJ, Walker JR, Enns MW, McWilliams LA. Self-criticism in generalized social phobia and response to cognitive-behaviour treatment. Behav Ther. 2002;33:479–91. [Google Scholar]
- 14.Cox BJ, Fleet C, Stein MB. Self-criticism and social phobia in the US national comorbidity survey. J Affect Disord. 2004;82(2):227–34. doi: 10.1016/j.jad.2003.12.012. [DOI] [PubMed] [Google Scholar]
- 15.Berthoz S, Armony J, Blair RJR, Dolan R. Neural correlates of violation of social norms and embarrassment. Brain. 2002;125(8):1696–1708. doi: 10.1093/brain/awf190. [DOI] [PubMed] [Google Scholar]
- 16.Keltner D, Buswell BN. Embarrassment: its distinct form and appeasement functions. Psychological Bulletin. 1997;122(3):250–270. doi: 10.1037/0033-2909.122.3.250. [DOI] [PubMed] [Google Scholar]
- 17.Takahashi H, Yahata N, Koeda M, Matsuda T, Asai K, Okubo Y. Brain activation associated with evaluative processes of guilt and embarrassment: an fMRI study. Neuroimage. 2004;23(3):967–74. doi: 10.1016/j.neuroimage.2004.07.054. [DOI] [PubMed] [Google Scholar]
- 18.Finger EC, Marsh AA, Kamel N, Mitchell DG, Blair JR. Caught in the act: The impact of audience on the neural response to morally and socially inappropriate behavior. Neuroimage. 2006;33(1):414–21. doi: 10.1016/j.neuroimage.2006.06.011. [DOI] [PubMed] [Google Scholar]
- 19.Moran JM, Heatherton TF, Kelley WM. Modulation of cortical midline structures by implicit and explicit self-relevance evaluation. Soc Neurosci. 2009;4(3):197–211. doi: 10.1080/17470910802250519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Mitchell JP, Macrae CN, Banaji MR. Dissociable medial prefrontal contributions to judgments of similar and dissimilar others. Neuron. 2006;50(4):655–63. doi: 10.1016/j.neuron.2006.03.040. [DOI] [PubMed] [Google Scholar]
- 21.Frith U, Frith CD. Development and neurophysiology of mentalizing. Philosophical Transactioins of the Royal Society B: Biological Sciences. 2003;358:459–73. doi: 10.1098/rstb.2002.1218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Association. AP. Diagnostic and Statistical Manual of Mental Disorders. Washington, DC: American Psychiatric Association; 1994. [Google Scholar]
- 23.Trower P, Gilbert P. New theoretical conceptions of social anxiety and social phobia. Clin Psychol Rev. 1989;9(19):19–35. [Google Scholar]
- 24.First M, Spitzer R, Gibbon M, Williams J. Structured Clinical Interview for DSM-IV. Washington, DC: American Psychiatric Press; 1997. [Google Scholar]
- 25.Cox RW. AFNI: software for analysis and visualization of functional magnetic resonance neuroimages. Computers and Biomedical Research. 1996;29:162–73. doi: 10.1006/cbmr.1996.0014. [DOI] [PubMed] [Google Scholar]
- 26.Talairach J, Tournoux P. Co-planar stereotaxic atlas of the human brain. Stuttgart, Thieme. 1988 [Google Scholar]
- 27.Northoff G, Heinzel A, de Greck M, Bermpohl F, Dobrowolny H, Panksepp J. Self-referential processing in our brain--a meta-analysis of imaging studies on the self. Neuroimage. 2006;31(1):440–57. doi: 10.1016/j.neuroimage.2005.12.002. [DOI] [PubMed] [Google Scholar]
- 28.Fletcher PC, Happe F, Frith U, Baker SC, Dolan RJ, Frackowiak RS, Frith CD. Other minds in the brain: a functional imaging study of "theory of mind" in story comprehension. Cognition. 1995;57:109–128. doi: 10.1016/0010-0277(95)00692-r. [DOI] [PubMed] [Google Scholar]
- 29.Gallagher HL, Happe F, Brunswick N, Fletcher PC, Frith U, Frith CD. Reading the mind in cartoons and stories: an fMRI study of ’theory of mind’ in verbal and nonverbal tasks. Neuropsychologia. 2000;38(1):11–21. doi: 10.1016/s0028-3932(99)00053-6. [DOI] [PubMed] [Google Scholar]
- 30.Brunet E, Sarfati Y, Hardy-Bayle MC, Decety J. A PET investigation of the attribution of intentions with a nonverbal task. Neuroimage. 2000;11(2):157–66. doi: 10.1006/nimg.1999.0525. [DOI] [PubMed] [Google Scholar]
- 31.Mitchell JP, Banaji MR, Macrae CN. The link between social cognition and self-referential thought in the medial prefrontal cortex. J Cogn Neurosci. 2005;17(8):1306–15. doi: 10.1162/0898929055002418. [DOI] [PubMed] [Google Scholar]
- 32.Moran JM, Macrae CN, Heatherton TF, Wyland CL, Kelley WM. Neuroanatomical evidence for distinct cognitive and affective components of self. J Cogn Neurosci. 2006;18(9):1586–94. doi: 10.1162/jocn.2006.18.9.1586. [DOI] [PubMed] [Google Scholar]
- 33.Phan KL, Taylor SF, Welsh RC, Ho SH, Britton JC, Liberzon I. Neural correlates of individual ratings of emotional salience: a trial-related fMRI study. Neuroimage. 2004;21(2):768–80. doi: 10.1016/j.neuroimage.2003.09.072. [DOI] [PubMed] [Google Scholar]
- 34.Seger CA, Stone M, Keenan JP. Cortical Activations during judgments about the self and an other person. Neuropsychologia. 2004;42(9):1168–77. doi: 10.1016/j.neuropsychologia.2004.02.003. [DOI] [PubMed] [Google Scholar]
- 35.Guyer AE, McClure-Tone EB, Shiffrin ND, Pine DS, Nelson EE. Probing the neural correlates of anticipated peer evaluation in adolescence. Child Dev. 2009;80(4):1000–15. doi: 10.1111/j.1467-8624.2009.01313.x. [DOI] [PMC free article] [PubMed] [Google Scholar]