

Abstract
Across plant species, leaves vary enormously in their size and their venation architecture, of which one major function is to replace water lost to transpiration. The leaf hydraulic conductance (Kleaf) represents the capacity of the transport system to deliver water, allowing stomata to remain open for photosynthesis. Previous studies showed that Kleaf relates to vein density (vein length per area). Additionally, venation architecture determines the sensitivity of Kleaf to damage; severing the midrib caused Kleaf and gas exchange to decline, with lesser impacts in leaves with higher major vein density that provided more numerous water flow pathways around the damaged vein. Because xylem embolism during dehydration also reduces Kleaf, we hypothesized that higher major vein density would also reduce hydraulic vulnerability. Smaller leaves, which generally have higher major vein density, would thus have lower hydraulic vulnerability. Tests using simulations with a spatially explicit model confirmed that smaller leaves with higher major vein density were more tolerant of major vein embolism. Additionally, for 10 species ranging strongly in drought tolerance, hydraulic vulnerability, determined as the leaf water potential at 50% and 80% loss of Kleaf, was lower with greater major vein density and smaller leaf size (|r| = 0.85–0.90; P < 0.01). These relationships were independent of other aspects of physiological and morphological drought tolerance. These findings point to a new functional role of venation architecture and small leaf size in drought tolerance, potentially contributing to well-known biogeographic trends in leaf size.
Leaf venation architecture has common functions across plant species, serving for mechanical support (Niklas, 1999), sugar and hormone transport (Kehr and Buhtz, 2008), and the replacement of water lost to transpiration during photosynthesis (Sack and Holbrook, 2006). However, venation architecture is highly diverse across species (Uhl and Mosbrugger, 1999; Roth-Nebelsick et al., 2001; Sack and Frole, 2006; Ellis et al., 2009; Brodribb et al., 2010). In dicotyledons, the leaf venation system typically consists of three orders of major veins and up to five higher orders of minor veins embedded in the mesophyll, with the vein orders arranged in a hierarchy; lower order veins are larger in diameter, with greater xylem conduit numbers and sizes, whereas higher order veins have greater length per area (vein density; Sack and Holbrook, 2006; McKown et al., 2010). Species vary strongly in the density of given vein orders and their conductivities (Cochard et al., 2004b; Sack and Frole, 2006). The aim of this study was to test for novel functional consequences of variation in leaf venation architecture and leaf size, and particularly a role in drought tolerance.
The leaf is a critical component in the plant water transport system, accounting for 30% or more of whole-plant hydraulic resistance (Sack and Holbrook, 2006). The leaf hydraulic conductance (Kleaf; flow rate/water potential driving force, i.e. the inverse of hydraulic resistance) quantifies a complex microhydrological system, including the conductances in series of the vein xylem (Kx) and the mesophyll pathways outside the xylem (Kox). The venation architecture is thus an important determinant of Kleaf and its dynamics. Total vein density is a determinant of both Kx and Kox, because, all else being equal, higher densities represent more numerous xylem flow pathways in parallel per leaf area and shorter pathways for water movement outside the xylem (Sack and Frole, 2006; Brodribb et al., 2007; McKown et al., 2010). Additionally, venation structure may contribute to the ability of Kleaf to withstand vein damage (Sack et al., 2008). Minor veins had been classically hypothesized to provide “conductive overload,” consisting of many parallel pathways for water flow such that a leaf could tolerate hydraulic disruption of major veins (Wylie, 1938). However, detailed studies found that Kleaf, stomatal conductance, and photosynthesis were very sensitive to damage of the large major veins, which supply water to the downstream vein hierarchy (Nardini et al., 2001; Huve et al., 2002; Nardini and Salleo, 2003; Sack et al., 2003a; Salleo et al., 2003; Delaney and Higley, 2006). Additionally, the impact of severing the midrib near its base differed among species. The decline of Kleaf was lower in palmately than in pinnately veined species, with greater major vein density providing flow pathways around the disrupted vein (Sack et al., 2008). The impact of midrib damage on Kleaf also varied among pinnately veined species. Smaller leaves, with their major veins spaced more closely and thus greater major vein densities, had greater tolerance of midrib damage (Sack et al., 2008).
Just as for leaves with damaged veins, Kleaf declines in dehydrating leaves, resulting in reductions of leaf gas exchange and whole-plant growth (Salleo et al., 2000; Brodribb and Holbrook, 2003; Sack and Holbrook, 2006; Johnson et al., 2009b). The Kleaf decline with dehydration arises at least in part from embolism in the vein xylem (Kikuta et al., 1997; Salleo et al., 2000, 2001; Nardini et al., 2003; Trifilò et al., 2003a, 2003b; Cochard et al., 2004a; Brodribb and Holbrook, 2005; Choat et al., 2005; Brodribb et al., 2009; Johnson et al., 2009a). Cavitation in turn will depend on the intrinsic vulnerability of each vein, with major veins likely to be more vulnerable because of their long and wide conduits (Choat et al., 2005). We hypothesized that higher major vein density, by providing transport pathways around embolized major veins, would confer tolerance of Kleaf to dehydration, i.e. more negative leaf water potential values (Ψleaf) at 50% and 80% loss of Kleaf (P50 and P80, respectively). Such a role for leaf venation could be important in the optimization of leaf size. Leaf size is highly variable across environments, with smaller leaves more frequent in dry habitats, both within and among species (Givnish, 1987; Sultan and Bazzaz, 1993; Gibson, 1998; Cunningham et al., 1999; Ackerly, 2003, 2004), as well as among community assemblages (Dolph and Dilcher, 1980; Fonseca et al., 2000). One advantage for small leaves is their thinner boundary layer, enabling more rapid convective cooling (Vogel, 1968, 1970; Parkhurst and Loucks, 1972; Gibson, 1998; Vogel, 2009; Nobel, 2010). Additionally, there may be a direct hydraulic benefit of small leaves, if their greater major vein redundancy protects Kleaf from decline and thus contributes to drought tolerance. To test these hypotheses, we conducted computer simulations of the impact of vein cavitation on Kleaf. We compared theoretical results with experimentally measured relationships among leaf hydraulic vulnerability, leaf size, venation architecture, and other aspects of leaf drought tolerance for species diverse in leaf form and drought sensitivity.
RESULTS
Impacts of Vein Cavitation Depend on Venation Architecture: Computer Simulations
Simulations implemented in the program K_leaf showed that the impacts of vein cavitation depended on vein density and leaf size (Fig. 1; Table I). Leaves were simulated of different sizes but with the same number of second-order (2°) veins; thus, larger leaves had their 2° veins spaced farther apart, and major vein density declined geometrically with increasing leaf size (major vein density = 1.194 × leaf size−0.5; rp = 0.999, P < 0.001). By contrast, across the simulated leaves, minor vein density was varied independently of leaf size (rp = 0.02, P = 0.89). Kx for uncavitated leaves correlated positively with both major vein density and minor vein density (rp = 0.75, P < 0.001 and rp = 0.15, P = 0.01 respectively).
Figure 1.

A and B, Results of computer model simulations of the percentage loss of whole-leaf xylem hydraulic conductance (PLC of Kx) after reducing by 90% major (A) and minor (B) vein conductivities to simulate cavitation for realistic leaves varying in major or minor vein density (n = 42 simulated leaves; see “Materials and Methods”). C, The dependence of PLC of Kx due to cavitation of major (gray circles) and minor (black circles) veins on the ratio of major to minor vein density.
Table I. Results of computer model simulations of the PLC of Kx, after reducing major and minor vein conductivities to simulate cavitation, in realistic leaves varying in major or minor vein density.
Contrasting impacts were found for the impacts of higher major and minor vein densities on the PLC of Kx due to vein cavitation, and contrasting impacts were found when cavitating major or minor veins. These effects can be understood in terms of the relative leverage of major or minor veins on the overall vein system. When the major vein density is increased, its greater redundancy gives the minor vein system a greater leverage; thus, the leaf is less sensitive to cavitation in the major veins and more sensitive to cavitation in the minor veins. By contrast, when the minor vein density is increased, its greater redundancy gives the major vein system a greater leverage; thus, the leaf is more sensitive to cavitation in the major veins and less sensitive to cavitation in the minor veins.
| Leaf Simulation | Cavitation in Major Veins | Cavitation in Minor Veins |
| Higher major vein density | Lower Kx decline | Greater Kx decline |
| Higher minor vein density | Greater Kx decline | Lower Kx decline |
| Higher major:minor vein density | Lower Kx decline | Greater Kx decline |
When major veins were reduced by 90% in cross-sectional conductivity to simulate the dysfunction of conduits due to embolism, the smaller leaves with greater major vein density showed a lesser impact on total xylem and whole-leaf hydraulic conductance per leaf area (Kx and Kleaf; i.e. a lower percentage loss of conductance [PLC]; Fig. 1A). Thus, across simulated leaves, the PLC of Kx resulting from major vein cavitation correlated negatively with major vein density (rp = −0.85, P < 0.001). For the simulated leaves with cavitated major veins, the Kx itself correlated strongly with major vein density, due both to the increase of maximum Kx by higher major vein density and the protective role of higher major vein density (rp = 0.97, P < 0.001). The importance of minor vein density was opposite from that of major vein density. A higher minor vein density increased the impact of cavitation of the major veins on the Kx (Fig. 1A). Thus, across the simulated leaves, the PLC of Kx resulting from major vein cavitation correlated positively with minor vein density (rp = 0.42, P = 0.001). For these simulated leaves with cavitated major veins, the Kx was not related to minor vein density (rp = 0.08, P = 0.57), because the greater Kx conferred by higher minor vein density was counteracted by a greater sensitivity to major vein cavitation. Overall, because of the contrasting effect of major and minor vein density, the PLC of Kx due to the cavitation of major veins was least for leaves with highest major vein density and lowest minor vein density and correlated negatively with the ratio of major to minor vein density (Fig. 1C; rp = −0.97, P < 0.001).
When the minor veins rather than the major veins were cavitated, Kx and Kleaf had a different dependency on venation architecture. When minor veins were reduced by 90% in cross-sectional conductivity to simulate cavitation, the smaller leaves with greater major vein density showed higher PLC of Kx (Fig. 1B). Thus, across simulated leaves, the PLC of Kx resulting from minor vein cavitation was positively related to major vein density (rp = 0.77, P < 0.001). By contrast, a higher minor vein density reduced the impact of cavitation of the minor veins on the Kx (Fig. 1B). Thus, across the simulated leaves, the PLC of Kx resulting from minor vein cavitation was negatively correlated with minor vein density (rp = −0.48, P = 0.001). For the simulated leaves with cavitated minor veins, the Kx itself correlated positively with minor vein density, due both to the increase of maximum Kx by higher minor vein density and the protective role of high minor vein density (rp = 0.66, P < 0.001). For these simulated leaves with cavitated minor veins, the Kx also positively correlated with major vein density, but more weakly than for uncavitated leaves, because the increase of maximum Kx due to higher major vein density was counteracted by the greater PLC driven by minor vein cavitation in leaves with higher major vein density (rp = 0.35 rather than 0.75, P < 0.001). Overall, the PLC of Kx resulting from cavitation of the minor veins was strongly positively correlated with the ratio of major to minor vein density (Fig. 1C; rp = 0.92, P < 0.001).
Notably, in the model simulations, the PLC of Kx resulting from major vein cavitation varied widely across the entire range of tested leaves with different major and minor vein densities (22%–87%; Fig. 1, A and C). By contrast, the PLC of Kx resulting from minor vein cavitation was less variable and very strong across the entire range of tested leaves (62%–90% and more than 80% for most simulated leaves; Fig. 1, B and C).
Diversity in Leaf Venation and Drought Tolerance Traits across Species of Moist and Dry Habitat
Across the 10 species diverse in drought tolerance, there were strong differences in leaf hydraulic conductance at full hydration (Kmax) and in their vulnerability to dehydration. The species varied in Kmax, P50, and P80 by 12- to 32-fold. The Kmax varied from 2.96 to 34.1 mmol m−2 s−1 MPa−1 for Comarostaphylos diversifolia and Platanus racemosa, in P50 from −0.09 to −2.85 MPa for P. racemosa and C. diversifolia; and in P80 from −0.35 to −5.25 MPa for P. racemosa and Cercocarpus betuloides (Supplemental Table S1).
The species differed strongly in leaf venation architecture and gross morphology, with substantial variation between moist and dry habitat species (Table II; Supplemental Table S1). Species varied 4-fold in major vein density, 3-fold in minor vein density, and 7-fold in the ratio of major to minor vein density. Species of moist and dry habitats did not differ significantly in minor and total vein densities (P = 0.11–0.74), but dry habitat species had 18% higher major vein density (with 14%–18% higher midrib, 2°, and third-order [3°] vein densities) and 50% higher ratio of major to minor vein density. Moist habitat species had 24% more free-ending veinlets per area, 13% higher minor vein diameters, and 14% higher number of 2° veins than dry habitat species (P < 0.001). Moist and dry habitat species did not differ in the diameters of their major veins (Supplemental Table S1). Species varied 18-fold in leaf area, with dry habitat species having on average 30% smaller leaves than moist habitat species. Leaf shape indices (length:width and perimeter squared:area) did not differ between habitats (P = 0.35–0.46).
Table II. Species, family, native range, and mean values ± se for morphological and physiological traits, and results of ANOVAs for the differences between moist and dry habitat species and among species nested within those categories.
*** P < 0.001, NS P > 0.05.
| Species | Family | Native Range | Leaf Area | Leaf Mass per Area | Major Vein Density | Minor Vein Density | Ratio of Major to Minor Vein Density | Cuticular Conductance |
| cm2 | g m−2 | mm−1 | mmol m−2 s−1 | |||||
| Dry habitat species | ||||||||
| Cercocarpus betuloides | Rosaceae | California, Mexico | 7.04 ± 1.73 | 156 ± 19.9 | 1.40 ± 0.14 | 7.74 ± 0.76 | 0.19 ± 0.03 | 3.99 ± 0.41 |
| Comarostaphylis diversifolia | Ericaceae | California, Mexico | 7.93 ± 1.89 | 254 ± 7.73 | 1.57 ± 0.18 | 4.17 ± 0.18 | 0.38 ± 0.04 | 2.87 ± 0.35 |
| Hedera canariensis | Araliacaeae | Canary Islands | 53.2 ± 14.4 | 78.1 ± 6.32 | 0.53 ± 0.06 | 3.00 ± 0.10 | 0.18 ± 0.01 | 0.44 ± 0.03 |
| Heteromeles arbutifolia | Rosaceae | California, Mexico | 14.6 ± 2.89 | 146 ± 13.2 | 0.88 ± 0.04 | 4.63 ± 0.11 | 0.19 ± 0.005 | 4.21 ± 1.22 |
| Quercus agrifolia | Fagaceae | California, Mexico | 13.5 ± 1.32 | 166 ± 7.64 | 1.07 ± 0.07 | 7.30 ± 0.23 | 0.15 ± 0.02 | 1.72 ± 0.23 |
| Moist habitat species | ||||||||
| Camellia sasanqua | Theaceae | Japan | 11.1 ± 0.45 | 144 ± 13.4 | 0.78 ± 0.04 | 3.31 ± 0.26 | 0.24 ± 0.03 | 1.77 ± 0.13 |
| Helianthus annuus | Asteraceae | Across North America | 44.3 ± 1.64 | 56.2 ± 6.98 | 0.48 ± 0.03 | 9.32 ± 0.44 | 0.05 ± 0.002 | 18.3 ± 1.92 |
| Lantana camara | Verbenaceae | Pantropical | 12.8 ± 3.09 | 79.0 ± 4.48 | 0.97 ± 0.12 | 9.75 ± 0.40 | 0.11 ± 0.02 | 12.0 ± 0.85 |
| Magnolia grandiflora | Magnoliaceae | Southern United States | 69.5 ± 5.51 | 180 ± 17.3 | 0.48 ± 0.02 | 5.16 ± 0.29 | 0.09 ± 0.003 | 3.88 ± 0.41 |
| Platanus racemosa | Platanaceae | California, Mexico | 80.9 ± 2.79 | 109 ± 6.54 | 0.40 ± 0.06 | 4.97 ± 0.14 | 0.08 ± 0.01 | 6.61 ± 0.41 |
| Average trait values | Dry area species | 19.2 ± 4.44 | 162 ± 11 | 1.09 ± 0.09 | 5.18 ± 0.35 | 0.22 ± 0.04 | 2.65 ± 0.45 | |
| Moist area species | 43.7 ± 14.3 | 113 ± 9.75 | 0.62 ± 0.05 | 6.23 ± 0.33 | 0.12 ± 0.03 | 8.44 ± 0.82 | ||
| ANOVA | Dry/moist Species | *** *** | *** *** | *** *** | NS *** | *** *** | *** *** | |
Several venation traits correlated with leaf size. The densities of first-order (1°), 2°, and 3° veins declined with increasing leaf size (rp and rs = −0.70 to −0.92, P < 0.05), as did the total major vein density (rp = −0.95, rs = −0.89, P < 0.001; Figs. 2A and 3) and the ratio of major to minor vein density (rp = −0.67, rs = −0.70, P = 0.03). Major vein density declined geometrically with increasing leaf size (i.e. with an exponent of −0.5; Fig. 2A). By contrast, minor vein density was independent of leaf size (rp and rs = −0.10 to −0.20, P > 0.05; Figs. 2B and 3), as were total vein density (as minor vein density accounted for 73%–95% of total vein density), vein diameters, and the number of free-ending veinlets per area (|rp| and |rs| = 0.02–0.43, P > 0.05; Fig. 3; Supplemental Table S2).
Figure 2.

The scaling of vein density with leaf size for 10 species varying strongly in drought tolerance. A, Major vein density versus leaf area. B, The independence of minor vein density from leaf size. Gray circles represent dry habitat species, and white circles represent moist habitat species. Fitted regression in A: major vein density = 0.32 × leaf area−0.53. *** P < 0.001, NS P > 0.05.
Figure 3.

Relation of the vulnerability of Kleaf to major and minor vein densities for 10 species varying strongly in drought tolerance. Vulnerability curves are plotted in the left column; each point represents a different measured leaf (n = 26–74 per curve). For each vulnerability curve, the fitted line is the maximum likelihood function for a given species (linear for C. sasanqua, C. diversifolia, Q. agrifolia, and H. arbutifolia; logistic for M. grandiflora, P. racemosa, H. annuus, H. canariensis, and L. camara; and sigmoidal for C. betuloides; r2 = 0.39–0.89; P < 0.001; see “Materials and Methods”) and the vertical line represents the water potential at P80. Leaf schematics are drawn to scale with major veins (1° and 2° veins). Micrographs of the minor vein architecture are represented on the right, indicating the independence of minor vein density from leaf size; in each image, the largest vein at the top is a 2° vein, with 3° veins branching off, and the minor veins make up the rest of the network.
Species of dry habitats also had greater expression of leaf drought tolerance traits than species from moist habitats, with 14% higher leaf mass per area (LMA), 11% thicker leaves, 18% higher modulus of elasticity, 15% to 16% more negative values for osmotic potential at full turgor and at turgor loss point, and 30% lower cuticular conductance. By contrast, species of moist habitats had on average 2- to 3-fold higher saturated water content and capacitances before and after turgor loss point (Supplemental Table S1; P < 0.001).
Relationships among Hydraulic Vulnerability, Venation, and Other Drought Tolerance and Morphological Traits
Across species, P50 and P80 were strongly correlated, and more negative values occurred in leaves with higher major vein density and smaller leaf size (|rp| and |rs| = 0.78–0.90, P < 0.01; Figs. 3 and 4; Supplemental Table S2). These relationships for the major vein system also held for component vein orders; the densities of 1°, 2°, and 3° veins, all intercorrelated, were greater in leaves with more negative P50 and P80 (|rp| and |rs| = 0.64–0.90, P < 0.01). Because leaf size and major vein densities were themselves negatively correlated (Fig. 2A), no correlation could be observed of leaf hydraulic vulnerability with leaf size or with major vein density independently of the other. Thus, in a partial correlation analysis, the relationships of P50 and P80 with leaf area were not significant after partialing out major vein density, and their relationships with major vein density were not significant after partialing out leaf area (|rpartial| = 0.08–0.29, P > 0.05). By contrast with major vein densities, other vein traits did not correlate with P50 or P80, including the minor vein density, total vein density, number of secondary veins, vein diameters, and number of free-ending veinlets per area (|rp| and |rs| = 0.01–0.57, P > 0.05). The ratio of major to minor vein density was positively correlated with P50 and P80 (rp and rs = 0.67–0.77, P < 0.05; Supplemental Table S2).
Figure 4.

Dependence of leaf hydraulic vulnerability, quantified as the water potential at 80% loss of conductivity (P80) on major vein density (A) and leaf area (B). Gray circles represent dry habitat species, and white circles represent moist habitat species. Fitted standardized major axis in A: P80 = ‒4.2 × major vein density + 0.92; fitted standardized major axis in B: P80 = ‒33.6 × leaf area−0.91.
Leaf hydraulic vulnerability also correlated with several other drought tolerance traits. The osmotic potentials at full and zero turgor, which were intercorrelated (rs and rp = 0.95–0.98, P < 0.001), both correlated with P50 and P80 (rp and rs = 0.71–0.85, P < 0.05; Supplemental Table S2). Leaves with higher LMA values tended to have more negative P80 (rp and rs = 0.71–0.73, P < 0.05; Supplemental Table S2), but LMA was not correlated with P50 (P > 0.05). However, both P50 and P80 were independent of other aspects of venation architecture and leaf morphology and physiology, including leaf shape indices (length:width and perimeter squared:area), the modulus of elasticity, capacitances at full and zero turgor, saturated water content, leaf thickness and density, and cuticular conductance (P > 0.05; Supplemental Table S2).
The linkage of hydraulic vulnerability (P50 and P80) with major vein density was independent from the relationships of hydraulic vulnerability to other leaf drought tolerance traits. The linkage of P50 or P80 with major vein density was apparently more fundamental. Thus, partialing out LMA or osmotic potentials at full and zero turgor did not remove the correlation of P50 or P80 with major vein density (rpartial = 0.68–0.78, P < 0.05). However, when partialing out the effect of major vein density on P50 or P80, their correlations with LMA and with osmotic potentials at full and zero turgor were lost (|rpartial| = 0.04–0.29, P > 0.05). Notably, the Kmax did not correlate with any venation architecture or morphological trait, including leaf size or leaf drought tolerance, for this set of diverse leaves (P > 0.05).
DISCUSSION
The Importance of Major Vein Density and Leaf Size in Resistance to Drought
We found novel, strong correlations of P50 and P80 with major vein density and leaf size across 10 species with diverse leaves, consistent with findings from the computer model simulations. All else being equal, leaf and whole-plant drought resistance would be conferred by a higher major vein density, which is generally associated with small leaf size (Dunbar-Co et al., 2009; McKown et al., 2010).
Such a role for venation and leaf size in determining hydraulic vulnerability has important potential ecological and biogeographic implications. A link between leaf size and P50 and P80 provides a new additional mechanism for the ecological distribution of leaf sizes. Leaf size evolves relatively quickly via several independent genetic pathways (Ackerly, 2009; Gonzalez et al., 2010). Small leaves are more common in dry and exposed habitats and larger leaves in moister and/or shaded habitats (Dolph and Dilcher, 1980; Givnish, 1987; Fonseca et al., 2000), and fossil leaf size is thus used as an indicator of past climate (Wilf, 1997). A direct hydraulic benefit of small leaves in drought tolerance, and the greater risk associated with large leaves under dry conditions, should thus be considered in addition to other demonstrated roles for leaf size in determining drought tolerance. A very well-established benefit of smaller leaves in warmer environments is their thinner boundary layer and more rapid convective cooling (Nobel, 1976; Nicotra et al., 2008; Vogel, 2009; Yates et al., 2010). Another advantage of small leaves in exposed conditions is that more leaves can be packed into a smaller space to capture irradiance, although this benefit carries a greater cost in support mass; more branch allocation is needed to support many small leaves than for fewer larger leaves, and this outweighs the lower requirement for petiole and midrib support of smaller leaves (Bragg and Westoby, 2002; Niinemets et al., 2006, 2007). Future work needs to tease apart the importance of the direct hydraulic mechanism in providing an advantage in drought tolerance for smaller leaves from these other benefits across different species sets.
Is the relationship of hydraulic tolerance of dehydration to leaf size and major vein density necessarily causal? We considered the possibility that these relationships could be coincidental, i.e. that small leaf size and high major vein density on the one hand, and more negative P50 and P80 on the other hand, might be independently selected in drought-tolerant species. In our study, two lines of evidence supported a causal relationship. First, the relationship was established by the computer simulations of leaves with altered sizes and venation architecture, all else being held fixed. In these simulations, cavitation of the major veins, as often observed during dehydration (see below), was better tolerated by smaller leaves with higher major vein density. Second, the partial correlation analyses of our experimental data showed that the relationship of venation architecture to leaf hydraulic vulnerability was independent of other drought tolerance traits, including turgor loss point (πTLP). Indeed, πTLP is the most reliable single predictor of a species’ drought tolerance to our knowledge (Auge et al., 1998; Sack et al., 2003b; Baltzer et al., 2008); thus, if selection for drought tolerance were to result in a coincidental correlation of vulnerability and venation architecture, both should show underlying correlations with πTLP. For our 10 species, P50 and P80 were strongly correlated with πTLP (Crombie et al., 1985; Blackman et al., 2010), but venation architecture was unrelated to πTLP. Furthermore, P50 and P80 lost their relationship with πTLP after partialing out major vein density, whereas they remained correlated with major vein density even after partialing out πTLP, indicating that the relationships of P50 and P80 to major vein density are more directly causal than any relationships with πTLP (Shipley, 2000).
We propose that the relationship between leaf size and hydraulic vulnerability in both simulated and real leaves supports a general mechanism, to be tested in other species sets, including closely related species within lineages in a phylogenetic context, because smaller leaves have evolved reliably in drier habitats (Ackerly et al., 2002; McDonald et al., 2003; Dunbar-Co et al., 2009; Santiago and Kim, 2009). In our modeled leaves and experimental species set, as across species in general, leaf size and major vein density were linked (Dunbar-Co et al., 2009; McKown et al., 2010). Future studies of species similar in leaf size but different in major vein densities are necessary to establish the role of venation independently of size in determining P50 and P80. A similar test could be conducted using Arabidopsis (Arabidopsis thaliana) vein mutants of similar leaf size with variation in major vein densities.
The Potential Roles of Venation in Determining Kleaf Decline
The linkage of hydraulic vulnerability with venation architecture shown here would be expected because of the reduction of xylem conductivity, due to cavitation or collapse of conduits in the vein xylem, as shown previously by dye experiments, cryo-scanning electron microscopy, and acoustic methods (Kikuta et al., 1997; Salleo et al., 2000; Nardini and Salleo, 2003; Cochard et al., 2004a; Brodribb and Holbrook, 2005; Johnson et al., 2009a). Notably, other factors besides vein density can determine leaf hydraulic vulnerability, which is a higher-level trait influenced by multiple lower-level traits (Marks and Lechowicz, 2006; McKown et al., 2010). Thus, species may additionally differ in the air-seeding thresholds of xylem conduits and in the responses of extravascular tissues to dehydration. Some small-leaved species in moist habitats might not have the low vulnerability suggested by their leaf size, if other factors were to override the benefit of high major vein density. However, our findings from simulated leaves and from our 10 species diverse in drought tolerance indicated a strong potential role of major vein density and leaf size in determining P50 and P80 across diverse species.
Model simulations showed that the vulnerability of Kx due to cavitation of major and minor veins was associated differently with venation architecture. Thus, loss of Kx resulting from cavitation in the major veins correlated with lower major vein density, the same relationship found in our experiments. However, loss of Kx resulting from cavitation in the major veins correlated with higher minor vein density. Furthermore, the opposite patterns were found for loss of Kx when cavitation was simulated in the minor veins. These various model results can be understood according to the simple principle of the relative leverage of the major or minor veins in the overall vein system (McKown et al., 2010). The hydraulic leverage of one vein system depends inversely on its density (i.e. its redundancy) relative to the other vein system, and cavitation has a stronger impact on the overall system when the leverage of that vein system is strongest. Thus, when the major vein density is higher, it has less leverage relative to the minor vein system, and cavitation of the major veins will cause a lower decrease in Kx and Kleaf (Table I). By contrast, when the minor vein density is higher, the major vein system gains leverage relative to the minor vein system; thus, cavitation of the major veins would cause a greater decrease in Kx and Kleaf (Table I). The model indicated that a higher major vein density is more effective for reducing hydraulic vulnerability due to major vein cavitation than a lower minor vein density. The model showed that reducing minor vein density only led to a strong reduction in PLC (i.e. to a strong gain in tolerance) at high major vein density. Furthermore, a higher major vein density also increases Kmax for well-hydrated leaves, both in absolute terms and relative to vein construction cost, all else being equal, whereas a lower minor vein density leads to losses of Kmax (McKown et al., 2010). Thus, for leaves with high major vein density, a low minor vein density might be a mechanism to achieve additional drought tolerance, all else being equal, but at the cost of maximum hydraulic capacity and providing no gain in absolute conductance when the major veins are cavitated. The mechanism of achieving higher major vein density with small leaf sizes was found in the model simulations and experimental study to provide a strong benefit in reducing vulnerability and thus for drought tolerance.
Notably, the model findings indicated that leaves with higher major vein density, although less sensitive to cavitation of the major veins, were more sensitive to cavitation of the minor vein system. Thus, our empirical findings, of reduced vulnerability in leaves with higher major vein density, suggested that major vein cavitation was more important than minor vein cavitation in driving loss of Kx and Kleaf. There are four lines of evidence that support the greater probability of cavitation in the major than minor veins. First, the major veins have wide and long xylem conduits especially vulnerable to air seeding (Choat et al., 2005). Second, in naturally dehydrated leaves, embolism is readily observed by cryo-scanning electron microscopy of major veins (Ball et al., 2004, 2006; Marenco et al., 2006; Johnson et al., 2009a), and acoustic studies have indicated cavitation in the major veins at Ψleaf values as high as −0.3 MPa (Crombie et al., 1985; Kikuta et al., 1997; Salleo et al., 2000; Johnson et al., 2009a). By contrast, the cryo-scanning electron microscopy studies published thus far have not shown cavitation of minor vein conduits (Canny, 2001). Studies of dye uptake into transpiring leaves did show less staining of minor veins in dehydrated leaves (Salleo et al., 2001; Nardini et al., 2003; Trifilo et al., 2003b); these findings are consistent with cavitation occurring principally in major veins, blocking uptake to the minor veins. Third, leaves that did not render their minor vein system resistant to cavitation would tend to be extremely sensitive to decline during drought, regardless of their venation architecture. The model simulations showed that cavitation of minor veins led to drastic declines in Kx across leaves of all venation architectures. As previously argued by Brodribb and Holbrook (2006), it seems improbable that leaves would invest in a fine vein network that becomes embolized at high water potentials and thus decline strongly in function. Fourth, a study of diverse angiosperms found that the leaf P50 was more negative in species with thicker-walled conduits in their minor veins (Blackman et al., 2010). That finding suggested that conduits are built to resist collapse at the tensions experienced during strong dehydration. In wood, such investment to avoid collapse signifies that the xylem can withstand cavitation to close to that degree of tension, as cavitation precedes collapse (Hacke and Sperry, 2001; Hacke et al., 2001; Blackman et al., 2010). Thus, minor vein conduits too should resist both cavitation and collapse at high levels of dehydration.
The collapse of xylem conduits during leaf dehydration cannot be entirely excluded. Collapse of conduits in the major or minor veins has never yet been shown for angiosperms but has been found in conifer needles dehydrated to water potentials ranging −1.5 to −3.5 MPa (Cochard et al., 2004a; Brodribb and Holbrook, 2005). Future studies are needed to analyze in detail the progression of cavitation and collapse in veins of different orders during leaf dehydration. Such work will also need to consider other aspects of the structure of the vein system (e.g. vessel widths and lengths and the degree that conduits span across vein orders), as these factors have been found to have great importance in stem vulnerability (Sperry, 2003; Sperry et al., 2005) and vary greatly across species (Sack and Frole, 2006). These traits may contribute to the correlation of vulnerability with low major vein density, because the major veins have especially long and wide vessels that span multiple orders. Other aspects of leaf vein arrangement in addition to vein density, such as looping in the major veins, might also influence resistance to hydraulic decline (Corson, 2010; Katifori et al., 2010).
While this discussion has focused on the decline of Kleaf with dehydration that is driven by declines in the xylem pathway conductivity (i.e. in Kx), there is also a potential role for declines in Kox in the correlation of P80 with major vein density and leaf size. The Kox may decline in dehydrating leaves due to changes in the permeability of membranes (Sack and Holbrook, 2006). Notably, given that a high major vein density would entail a large bundle sheath area by which water exits the major veins, if the lamina near the major veins accounts for a large part of transpiration (Fricke, 2002; Sack et al., 2002; Nardini et al., 2010), then leaves with high major vein density would likely maintain a greater bundle sheath area and a greater Kox when cells lose turgor in dehydrating leaves (Kim and Steudle, 2007). Additionally, we note that as leaves dehydrate, it is possible that Kleaf, the bulk leaf parameter, may not well describe the water transport pathways if sectors or “patches” of lamina become isolated, each with its own hydraulic supply (Barbour and Farquhar, 2004). Leaves with high major vein density may better provide for access of isolated leaf sectors to the lower-order vein distribution system. Future studies are needed of the impact of dehydration on the bundle sheath and mesophyll tissues and on the potential heterogeneity of water supply in dehydrating leaves. Detailed characterization of the causes of Kleaf decline and its dependence on xylem and mesophyll characters will contribute to an ability to predict variation in a species’ drought responses from cell- and tissue-level properties.
Relationship of Leaf Vulnerability to Leaf and Whole-Plant Drought Tolerance
Our results also highlighted the importance of cell properties and leaf morphology in drought tolerance. Species from dry habitats had higher LMA and modulus of elasticity, more negative osmotic potentials at full and zero turgor (πTLP), and lower cuticular conductance than species from moist habitats, which by contrast had higher saturated water content and capacitance. Furthermore, the πTLP correlated with P50 and P80, as shown in two previous studies (Crombie et al., 1985; Blackman et al., 2010). Given that stomata tend to close near the πTLP (Hao et al., 2010), this linkage points to a control of stomatal aperture during drought by hydraulic vulnerability. The linkage might arise mechanistically if a low osmotic potential in leaf tissues, reflected by πTLP, acted to reduce turgor loss and the decline of membrane permeability (Canny and Huang, 2006). Alternatively, the πTLP might be coselected with P50 and P80 in drought-tolerant leaves such that stomatal closure precedes hydraulic dysfunction. Our results were most consistent with this second possibility. In the partial correlation analysis, πTLP had no impact on P50 and P80 when major vein density was partialed out. Thus, as found in the model simulations, leaf size and major vein density are putative causes of high P50 and P80, whereas πTLP is apparently a structurally independent but coordinated trait that modulates the leaf and plant response to drought.
The finding that major vein density and small leaf size reduce leaf hydraulic vulnerability points to potential roles in determining whole-plant drought tolerance. These traits may be especially important because the leaf is a key locus in whole-plant vulnerability, with typically greater hydraulic sensitivity than stems and roots (Hao et al., 2008; Brodribb and Cochard, 2009; Domec et al., 2009). Thus, for three of the four chaparral species in this study, the leaf P50 was 2.7 to 4.7 MPa less negative than that previously reported for stems (Jacobsen et al., 2007), with only Quercus agrifolia having stems apparently similar to leaves in their vulnerability (−2 versus −2.4 MPa, respectively). The importance of leaf hydraulic vulnerability in whole-plant drought tolerance was further supported in this study both by the more negative P50 and P80 values for species of dry than moist habitats and by the general correlation of these traits with others related to leaf drought tolerance. It is important to recognize, however, that there can be other possible routes to leaf drought tolerance than a low hydraulic vulnerability and thus being able to maintain hydraulic and photosynthetic function during drought. Thus, some species can achieve substantial drought tolerance via a low cuticular conductance and water storage, with an extreme development of this mechanism in succulent plants (Ogburn and Edwards, 2009). In this study, Hedera canariensis showed this drought tolerance mechanism (Sack et al., 2003c; Metcalfe, 2005), possibly explaining its relatively high Kleaf vulnerability for a species that can tolerate drought. Other species can achieve drought tolerance via an ability to tolerate low tissue water potentials via dehydrin expression that prevents mechanical failure of the cell walls (e.g. the resurrection fern [Polypodium polypodioides]; Layton et al., 2010). Species with these alternative mechanisms of leaf drought tolerance can achieve large leaf sizes even given dry conditions (Nobel and Jordan, 1983). Furthermore, we note that drought tolerance achieved at the level of the whole plant may not always correspond to leaf-level drought tolerance. Some species with drought-sensitive leaves can tolerate dry soil by shedding leaves or achieving deep roots, as is the case for Lantana camara (Castillo et al., 2007), which had relatively high Kleaf vulnerability. By contrast, some species with drought-tolerant leaves may be sensitive to drought in the field due to relatively shallow roots (e.g. Magnolia grandiflora; Klos et al., 2009). Future work will establish the degree to which, even despite such complexity, key leaf traits such as major vein density, leaf size, and P50 and P80 contribute to drought tolerance. This work has strong potential to explain leaf function during drought from cell-, tissue-, and organ-level physiological properties and to predict the drought tolerance of diverse species and landscapes in current and extinct vegetation from their leaf traits.
MATERIALS AND METHODS
Computer Simulations of the Importance of Vein Architecture in Leaf Hydraulic Vulnerability
Simulations of the impact of cavitation in leaves with varying venation architecture were generated using the program K_leaf, version 6 (written by H. Cochard, Institut National de la Recherche Agronomique, Clermont-Ferrand, France [Cochard et al., 2004b; McKown et al., 2010]; available on request at herve.cochard@clermont.inra.fr). Based on specified parameters, K_leaf creates a spatially explicit model of a leaf with up to six vein orders represented as a square grid of xylem resistors and outside-xylem resistors (“mesophyll” resistors) branching orthogonally from each junction of the vein grid. In modeled leaves, water exits through the mesophyll resistor located at each vein junction, and the bulk of the water thus exits from the numerous minor veins. The model determines three parameters, Kx, Kox, and Kleaf, where:
for leaves given specified size, densities, and cross-sectional conductivities for each vein order and mesophyll conductance. Simulations were modeled using an elliptical leaf with 12 pairs of 2° veins and with vein densities and conductivities based on those for a Juglans regia terminal leaflet (Cochard et al., 2004b). Our findings should be applicable to other leaves with hierarchical, reticulate venation (McKown et al., 2010). Individual vein conductivities (kv) were based on estimations from xylem conduit lumen dimensions in Juglansvein cross-sections using the formula:
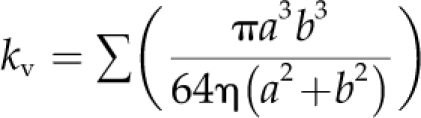 |
where a and b are the major and minor axes of ellipses and η is the viscosity of water at 25°C (units are mmol m s−1 MPa−1; Lewis and Boose, 1995; Cochard et al., 2004b; Sack and Frole, 2006). The Kx calculated by the model depends on the individual vein order conductivities and densities, and Kox depends on the specified mesophyll conductance and the total vein density, which determines the number of junctions and thus of mesophyll resistors in parallel.
Values of Kx, Kox, and Kleaf were determined in typical units normalized by leaf area (mmol m−2 s−1 MPa−1). The relative responses of Kx, Kox, and Kleaf to alteration of venation features in our simulations are expected to accurately indicate relative trends and principles of leaf venation design; however, the empirical values are not to be taken as meaningful, and units are not presented in our simulation results. For instance, the simulations based on the Juglans leaflet anatomical data set produced a Kx of 462 mmol m−2 s−1 MPa−1, which is very high relative to experimentally measured Kx and many times greater than measured Kox (Cochard et al., 2004b). Cochard et al. (2004b) introduced the “xylem hydraulic efficiency” parameter in K_leaf (modeled Kx divided by measured Kx) to calibrate the modeled Kx with measured values and thus to account for other factors than xylem conduit numbers and diameters that cannot currently be modeled, such as pit membrane resistance (Sperry et al., 2005) or conduit blockage by embolism or tyloses (Salleo et al., 2002; Choat et al., 2005). In our simulations, xylem hydraulic efficiency was set to 1. While not significant for this study, future work should better reconcile modeled Kx with experimentally measured values (McKown et al., 2010).
For this study, we focused on the impacts of simulated cavitation on Kx values, which would result in a reduction of Kleaf by a degree that depends on the value of Kx relative to Kox. The ratio of Kox and Kx depends on species and on environmental variables that affect these compartments differently, but the available data suggest that Kox and Kx are of similar magnitude on average (Sack and Holbrook, 2006), and in that case, a given decline of Kx would reduce Kleaf by approximately half that amount. We also note that dehydration could also impact on the extraxylem pathways (e.g. due to cell shrinkage and/or aquaporin deactivation; Kim and Steudle, 2007), which would lead to stronger overall impacts on Kleaf.
The program generates leaves of a specified size and number and arrangement of 2° veins, from which it determines the 2° vein density. Thus, designating leaves of larger sizes leads to the 2° veins being spaced farther apart, just as observed in real leaves (Sack et al., 2008; see “Results”). The density of the minor veins (in this model, 3° and higher) depends on the areole size, which is specified independently, and thus is manipulated independently of major vein density. We altered leaf size (and thus the density of 1° and 2° veins) and also minor vein densities, simulating a total of 42 leaves of six different sizes (4.8–65.4 cm2), associated with a 3.5-fold range of major vein densities (0.15–0.53 mm mm−2) × seven different minor vein densities (spanning a 3.5-fold range; 2.8–8.7 mm mm−2). For each leaf, we additionally applied cavitation “treatments” to determine the impact on Kx: (1) to test the impact of cavitating the major veins, we reduced the cross-sectional conductivities of the 1° and 2° veins by 90%, to simulate the great majority of vessels being cavitated; and (2) to test the impact of cavitating the minor veins, we reduced the cross-sectional conductivities of the 3°, 4°, 5°, and 6° veins by 90%. For the simulated leaves of contrasting venation architecture, we present the PLC of Kx that resulted from these treatments (i.e. the decline relative to a control, uncavitated leaf).
Plant Material and Leaf Hydraulic Vulnerability
Leaf hydraulic vulnerability was determined for 10 species sampled in and around the campus of the University of California, Los Angeles, and Will Rogers State Park, in Los Angeles, in May to September 2008 (Table II). Leaves were collected from mature trees and shrubs of nine species. Leaves from sunflower (Helianthus annuus var Sunspot; Botanical Interests) were collected from greenhouse plants grown from seeds in 3.6-L pots (average minimum, mean, and maximum values for temperature, 21.1°C, 23.2°C, and 26.0°C; for humidity, 44%, 51%, and 59%). Sunflowers were irrigated every 2 d, with 200 to 250 μL L−1 20:20:20 nitrogen:phosphorus:potassium; the light availability measured at midday on a sunny day was up to 550 μmol photons m−2 s−1 and on average 300 μmol photons m−2 s−1 (LI-250 light meter; LI-COR Biosciences). Species spanning a wide range of drought sensitivity were selected across nine families to include phylogenetic diversity. Five species were native to dry habitats and five species to moist habitats (Table II).Mature, healthy leaves were excised from sun-exposed branches rehydrated overnight. We used the evaporative flux method (EFM) to determine leaf vulnerability curves. We measured Kleaf as the light-acclimated steady-state transpirational flow rate for excised leaves (E; mmol m−2 s−1) divided by the water potential driving force (ΔΨleaf; MPa; Sack et al., 2002; Scoffoni et al., 2008). The EFM was modified to allow determination of Kleaf at low Ψleaf. Shoots were cut into segments of three or more leaves under ultrapure water and dehydrated with a fan for different periods of time to achieve a range of Ψleaf values. Shoots were allowed to equilibrate for at least 30 min, and then two leaves were excised and measured for initial Ψleaf (Ψo) using a pressure chamber (Plant Moisture Stress model 1000; PMS Instrument Co.). The third leaf (typically the middle leaf) was used to determine Kleaf with the EFM. The vulnerability curve was obtained by plotting Kleaf against which ever was lowest, Ψo or Ψfinal (“Ψlowest”), assuming this to be the Ψleaf associated with the strongest dehydration experienced during the experiment (bench drying and Kleaf measurement). Species show different shapes in their vulnerability curve trajectories, as expected given variations in the importance of multiple mechanisms for the decline of Kleaf with dehydration (Brodribb and Holbrook, 2006). Thus, we determined each species’ vulnerability curve, selecting among four functions applied in the previous literature by maximum likelihood (Burnham and Anderson, 2002) using the optim function in R 2.9.2 (http://www.r-project.org; Burnham and Anderson, 2004; Sack et al., 2006; our scripts are available on request): linear (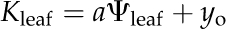 ), sigmoidal (
), sigmoidal (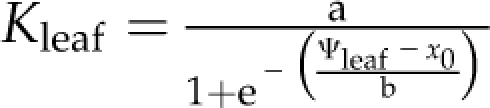 ), logistic (
), logistic (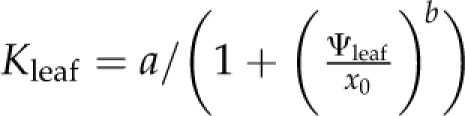 ), and exponential (
), and exponential (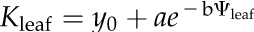 ). We used the best-fit function for each species to estimate the maximum Kleaf for the hydrated leaf (Kmax) and the Ψleaf at 50% and 80% loss of Kleaf (P50 and P80, respectively).
). We used the best-fit function for each species to estimate the maximum Kleaf for the hydrated leaf (Kmax) and the Ψleaf at 50% and 80% loss of Kleaf (P50 and P80, respectively).
Quantification of Leaf Form and Venation Architecture
We determined venation traits from one leaf on an exposed branch from each of three individuals per species from the same individuals measured for hydraulic vulnerability. Leaves were collected in May to September 2007 and fixed in formalin-acetic acid solution (37% aqueous formaldehyde solution, 50% ethanol, and 13% glacial acetic acid solution). Leaves were chemically cleared with 5% NaOH in ethanol, stained with safranin, and counterstained with fast-green (Berlyn and Miksche, 1976). Leaves were mounted with water in transparency film (CG5000; 3M Visual Systems Division) and scanned (flatbed scanner; Canon Scan Lide 90; 1,200 pixels per inch). The leaf area, length, width, perimeter, and numbers and lengths of 1° and 2° veins were measured using ImageJ software (version 1.42q; National Institutes of Health). Two indices of leaf shape were calculated: the length:width ratio and the perimeter squared:area ratio (a size-independent index of edge relative to size; Sack et al., 2003b). The 3° vein lengths were measured for three rectangles per leaf (10–300 mm2, depending on leaf size) located centrally in the top, middle, and bottom thirds of the leaf. For each vein order, the vein density was calculated as length divided by leaf area; for 3° veins, the vein densities were averaged across the three subsampled rectangles. Vein diameters, excluding the bundle sheath, were measured for each vein order by averaging six measurements (two made centrally in veins in the top, middle, and bottom thirds of the leaf).
Measurements of the minor vein system were made with a light microscope (DMRB; Leica Microsystems) with a 5× or 10× objective and digital camera (14.2 Color Mosaic; Diagnostic Instruments). Three rectangles were imaged (areas of 1.5 mm2 or 6 mm2) centrally in the top, middle, and bottom thirds of the leaf, and the number of vein orders, density of minor veins (length per area), number of free vein endings per area, and vein diameters measured centrally in six segments were averaged across the rectangles. The major vein density was determined as the sum of 1°, 2°, and 3° vein densities and the minor vein density as the sum for 4° and higher order veins. The ratio of major to minor vein density was calculated for each leaf for every species.
Quantifying Other Key Traits Related to Leaf Drought Tolerance
Morphological and physiological traits related to leaf drought tolerance were measured for six leaves taken from three to six individuals per species. Measurements were made of leaf area and of dry mass after oven drying at least 48 h at more than 70°C to allow calculation of LMA (g m−2). Leaf thickness was determined using digital calipers (Fowler), and leaf density was calculated as LMA divided by leaf thickness (Witkowski and Lamont, 1991). Cuticular conductance, the minimum conductance to vapor diffusion across the epidermis when the stomata are closed, was measured by weighing leaves as they dehydrated (Sack et al., 2003b; Sack and Scoffoni, 2010). Parameters were determined from pressure-volume curves constructed by measuring leaf water potential and relative water content as leaves dehydrated (Tyree and Hammel, 1972; Sack et al., 2003b), including osmotic potential at full turgor (MPa) and at turgor loss point (πTLP; MPa), saturated water content (g g−1), modulus of elasticity (MPa), and capacitances at full turgor and at turgor loss point (MPa−1; Sack, 2010).
Statistical Analyses of Trait Differences and Correlations across Species
To test trait differences between moist and dry habitat species (Table II; Supplemental Table S1), we performed ANOVAs with species nested within habitat type (Minitab release 15). Prior to tests, data were log transformed to improve normality and heteroscedasticity (Sokal and Rohlf, 1995). We performed t tests for leaf density, maximum Kleaf, P50, and P80 where only species mean values were available (Supplemental Table S1).
A correlation matrix was determined to reveal the intercorrelative structure of hydraulic parameters, leaf size, venation architecture, and other traits putatively related to drought tolerance. For a conservative estimation, correlations were considered significant only at P < 0.05 for both Spearman and Pearson coefficients (rs and rp, respectively). Because many relationships were nonlinear, we determined Pearson correlations for both raw and log-transformed data.
When three variables of interest were intercorrelated across species, we performed partial correlation analysis (Sokal and Rohlf, 1995), testing the relationship between two variables while holding the third variable constant (corpcor package in R; Schaefer et al., 2007).
Supplemental Data
The following materials are available in the online version of this article.
Supplemental Table S1. Species means ± se for 24 morphological, anatomical, and physiological traits and results of ANOVA testing for species differences and for differences between moist and dry habitat species.
Supplemental Table S2. Correlation matrix for the relationship of leaf hydraulic vulnerability traits with venation architecture and other traits related to leaf morphology and drought tolerance.
Supplementary Material
Acknowledgments
We thank Weimin Dang and the University of California, Los Angeles, Plant Growth Facility, Art Gibson, and Phil Rundel for logistical assistance.
References
- Ackerly D. (2009) Conservatism and diversification of plant functional traits: evolutionary rates versus phylogenetic signal. Proc Natl Acad Sci USA (Suppl 2) 106: 19699–19706 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ackerly DD. (2003) Community assembly, niche conservatism, and adaptive evolution in changing environments. Int J Plant Sci 164: S165–S184 [Google Scholar]
- Ackerly DD. (2004) Adaptation, niche conservatism, and convergence: comparative studies of leaf evolution in the California chaparral. Am Nat 163: 654–671 [DOI] [PubMed] [Google Scholar]
- Ackerly DD, Knight CA, Weiss SB, Barton K, Starmer KP. (2002) Leaf size, specific leaf area and microhabitat distribution of chaparral woody plants: contrasting patterns in species level and community level analyses. Oecologia 130: 449–457 [DOI] [PubMed] [Google Scholar]
- Auge RM, Duan XG, Croker JL, Witte WT, Green CD. (1998) Foliar dehydration tolerance of twelve deciduous tree species. J Exp Bot 49: 753–759 [Google Scholar]
- Ball MC, Canny MJ, Huang CX, Egerton JJG, Wolfe J. (2006) Freeze/thaw-induced embolism depends on nadir temperature: the heterogeneous hydration hypothesis. Plant Cell Environ 29: 729–745 [DOI] [PubMed] [Google Scholar]
- Ball MC, Canny MJ, Huang CX, Heady RD. (2004) Structural changes in acclimated and unacclimated leaves during freezing and thawing. Funct Plant Biol 31: 29–40 [DOI] [PubMed] [Google Scholar]
- Baltzer JL, Davies SJ, Bunyavejchewin S, Noor NSM. (2008) The role of desiccation tolerance in determining tree species distributions along the Malay-Thai peninsula. Funct Ecol 22: 221–231 [Google Scholar]
- Barbour MM, Farquhar GD. (2004) Do pathways of water movement and leaf anatomical dimensions allow development of gradients in H218O between veins and the sites of evaporation within leaves? Plant Cell Environ 27: 107–121 [Google Scholar]
- Berlyn GP, Miksche JP. (1976) Botanical Microtechnique and Cytochemistry. Iowa State University Press, Ames [Google Scholar]
- Blackman CJ, Brodribb TJ, Jordan GJ. (2010) Leaf hydraulic vulnerability is related to conduit dimensions and drought resistance across a diverse range of woody angiosperms. New Phytol 188: 1113–1123 [DOI] [PubMed] [Google Scholar]
- Bragg JG, Westoby M. (2002) Leaf size and foraging for light in a sclerophyll woodland. Funct Ecol 16: 633–639 [Google Scholar]
- Brodribb TJ, Cochard H. (2009) Hydraulic failure defines the recovery and point of death in water-stressed conifers. Plant Physiol 149: 575–584 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brodribb TJ, Feild TS, Jordan GJ. (2007) Leaf maximum photosynthetic rate and venation are linked by hydraulics. Plant Physiol 144: 1890–1898 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brodribb TJ, Feild TS, Sack L. (2010) Viewing leaf structure and evolution from a hydraulic perspective. Funct Plant Biol 37: 488–498 [Google Scholar]
- Brodribb TJ, Holbrook NM. (2003) Stomatal closure during leaf dehydration, correlation with other leaf physiological traits. Plant Physiol 132: 2166–2173 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brodribb TJ, Holbrook NM. (2005) Water stress deforms tracheids peripheral to the leaf vein of a tropical conifer. Plant Physiol 137: 1139–1146 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brodribb TJ, Holbrook NM. (2006) Declining hydraulic efficiency as transpiring leaves desiccate: two types of response. Plant Cell Environ 29: 2205–2215 [DOI] [PubMed] [Google Scholar]
- Brodribb TJ, McAdam SAM, Jordan GJ, Feild TS. (2009) Evolution of stomatal responsiveness to CO2 and optimization of water-use efficiency among land plants. New Phytol 183: 839–847 [DOI] [PubMed] [Google Scholar]
- Burnham KP, Anderson DR. (2002) Model Selection and Multimodel Inference, Ed 2. Springer, New York [Google Scholar]
- Burnham KP, Anderson DR. (2004) Multimodel inference: understanding AIC and BIC in model selection. Sociol Methods Res 33: 261–304 [Google Scholar]
- Canny MJ. (2001) Embolisms and refilling in the maize leaf lamina, and the role of the protoxylem lacuna. Am J Bot 88: 47–51 [PubMed] [Google Scholar]
- Canny MJ, Huang CX. (2006) Leaf water content and palisade cell size. New Phytol 170: 75–85 [DOI] [PubMed] [Google Scholar]
- Castillo JM, Leira-Doce P, Carrion-Tacuri J, Munoz-Guacho E, Arroyo-Solis A, Curado G, Doblas D, Rubio-Casal AE, Alvarez-Lopez AA, Redondo-Gomez S, et al. (2007) Contrasting strategies to cope with drought by invasive and endemic species of Lantana in Galapagos. Biodivers Conserv 16: 2123–2136 [Google Scholar]
- Choat B, Lahr EC, Melcher P, Zwieniecki MA, Holbrook NM. (2005) The spatial pattern of air seeding thresholds in mature sugar maple trees. Plant Cell Environ 28: 1082–1089 [Google Scholar]
- Cochard H, Froux F, Mayr S, Coutand C. (2004a) Xylem wall collapse in water-stressed pine needles. Plant Physiol 134: 401–408 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cochard H, Nardini A, Coll L. (2004b) Hydraulic architecture of leaf blades: where is the main resistance? Plant Cell Environ 27: 1257–1267 [Google Scholar]
- Corson F. (2010) Fluctuations and redundancy in optimal transport networks. Phys Rev Lett 104: 048703. [DOI] [PubMed] [Google Scholar]
- Crombie DS, Milburn JA, Hipkins MF. (1985) Maximum sustainable xylem sap tensions in Rhododendron and other species. Planta 163: 27–33 [DOI] [PubMed] [Google Scholar]
- Cunningham SA, Summerhayes B, Westoby M. (1999) Evolutionary divergences in leaf structure and chemistry, comparing rainfall and soil nutrient gradients. Ecol Monogr 69: 569–588 [Google Scholar]
- Delaney KJ, Higley LG. (2006) An insect countermeasure impacts plant physiology: midrib vein cutting, defoliation and leaf photosynthesis. Plant Cell Environ 29: 1245–1258 [DOI] [PubMed] [Google Scholar]
- Dolph GE, Dilcher DL. (1980) Variation in leaf size with respect to climate in the tropics of the Western-hemisphere. Bull Torrey Bot Club 107: 154–162 [Google Scholar]
- Domec JC, Noormets A, King JS, Sun G, McNulty SG, Gavazzi MJ, Boggs JL, Treasure EA. (2009) Decoupling the influence of leaf and root hydraulic conductances on stomatal conductance and its sensitivity to vapour pressure deficit as soil dries in a drained loblolly pine plantation. Plant Cell Environ 32: 980–991 [DOI] [PubMed] [Google Scholar]
- Dunbar-Co S, Sporck MJ, Sack L. (2009) Leaf trait diversification and design in seven rare taxa of the Hawaiian Plantago radiation. Int J Plant Sci 170: 61–75 [Google Scholar]
- Ellis B, Daly DC, Hickley LJ, Johnson KR, Mitchell JD, Wilf P, Wing SL. (2009) Manual of Leaf Architecture. Cornell University Press/New York Botanical Garden, New York [Google Scholar]
- Fonseca CR, Overton JM, Collins B, Westoby M. (2000) Shifts in trait-combinations along rainfall and phosphorus gradients. J Ecol 88: 964–977 [Google Scholar]
- Fricke W. (2002) Biophysical limitation of cell elongation in cereal leaves. Ann Bot (Lond) 90: 157–167 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gibson AC. (1998) Photosynthetic organs of desert plants. Bioscience 48: 911–920 [Google Scholar]
- Givnish TJ. (1987) Comparative studies of leaf form: assessing the relative roles of selective pressures and phylogenetic constraints. New Phytol 106: 131–160 [Google Scholar]
- Gonzalez N, De Bodt S, Sulpice R, Jikumaru Y, Chae E, Dhondt S, Van Daele T, De Milde L, Weigel D, Kamiya Y, et al. (2010) Increased leaf size: different means to an end. Plant Physiol 153: 1261–1279 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hacke UG, Sperry JS. (2001) Functional and ecological xylem anatomy. Perspect Plant Ecol Evol Syst 4: 97–115 [Google Scholar]
- Hacke UG, Sperry JS, Pockman WT, Davis SD, McCulloch KA. (2001) Trends in wood density and structure are linked to prevention of xylem implosion by negative pressure. Oecologia 126: 457–461 [DOI] [PubMed] [Google Scholar]
- Hao GY, Hoffmann WA, Scholz FG, Bucci SJ, Meinzer FC, Franco AC, Cao KF, Goldstein G. (2008) Stem and leaf hydraulics of congeneric tree species from adjacent tropical savanna and forest ecosystems. Oecologia 155: 405–415 [DOI] [PubMed] [Google Scholar]
- Hao GY, Sack L, Wang AY, Cao KF, Goldstein G. (2010) Differentiation of leaf water flux and drought tolerance traits in hemiepiphytic and non-hemiepiphytic Ficus tree species. Funct Ecol 24: 731–740 [Google Scholar]
- Huve K, Remus R, Luttschwager D, Merbach W. (2002) Water transport in impaired leaf vein systems. Plant Biol 4: 603–611 [Google Scholar]
- Jacobsen AL, Pratt RB, Ewers FW, Davis SD. (2007) Cavitation resistance among 26 chaparral species of southern California. Ecol Monogr 77: 99–115 [Google Scholar]
- Johnson DM, Meinzer FC, Woodruff DR, McCulloh KA. (2009a) Leaf xylem embolism, detected acoustically and by cryo-SEM, corresponds to decreases in leaf hydraulic conductance in four evergreen species. Plant Cell Environ 32: 828–836 [DOI] [PubMed] [Google Scholar]
- Johnson DM, Woodruff DR, McCulloh KA, Meinzer FC. (2009b) Leaf hydraulic conductance, measured in situ, declines and recovers daily: leaf hydraulics, water potential and stomatal conductance in four temperate and three tropical tree species. Tree Physiol 29: 879–887 [DOI] [PubMed] [Google Scholar]
- Katifori E, Szöllosi GJ, Magnasco MO. (2010) Damage and fluctuations induce loops in optimal transport networks. Phys Rev Lett 104: 048704. [DOI] [PubMed] [Google Scholar]
- Kehr J, Buhtz A. (2008) Long distance transport and movement of RNA through the phloem. J Exp Bot 59: 85–92 [DOI] [PubMed] [Google Scholar]
- Kikuta SB, LoGullo MA, Nardini A, Richter H, Salleo S. (1997) Ultrasound acoustic emissions from dehydrating leaves of deciduous and evergreen trees. Plant Cell Environ 20: 1381–1390 [Google Scholar]
- Kim YX, Steudle E. (2007) Light and turgor affect the water permeability (aquaporins) of parenchyma cells in the midrib of leaves of Zea mays. J Exp Bot 58: 4119–4129 [DOI] [PubMed] [Google Scholar]
- Klos RJ, Wang GG, Bauerle WL, Rieck JR. (2009) Drought impact on forest growth and mortality in the southeast USA: an analysis using Forest Health and Monitoring data. Ecol Appl 19: 699–708 [DOI] [PubMed] [Google Scholar]
- Layton BE, Boyd MB, Tripepi MS, Bitonti BM, Dollahon MNR, Balsamo RA. (2010) Dehydration induced expression of a 31-kDa dehydrin in Polypodium polypodioides (Polypodiaceae) may enable large, reversible deformation of cell walls. Am J Bot 97: 535–544 [DOI] [PubMed] [Google Scholar]
- Lewis AM, Boose ER. (1995) Estimating volume flow-rates through xylem conduits. Am J Bot 82: 1112–1116 [Google Scholar]
- Marenco RA, Siebke K, Farquhar GD, Ball MC. (2006) Hydraulically based stomatal oscillations and stomatal patchiness in Gossypium hirsutum. Funct Plant Biol 33: 1103–1113 [DOI] [PubMed] [Google Scholar]
- Marks CO, Lechowicz MJ. (2006) Alternative designs and the evolution of functional diversity. Am Nat 167: 55–66 [DOI] [PubMed] [Google Scholar]
- McDonald PG, Fonseca CR, Overton JM, Westoby M. (2003) Leaf-size divergence along rainfall and soil-nutrient gradients: is the method of size reduction common among clades? Funct Ecol 17: 50–57 [Google Scholar]
- McKown AD, Cochard H, Sack L. (2010) Decoding leaf hydraulics with a spatially explicit model: principles of venation architecture and implications for its evolution. Am Nat 175: 447–460 [DOI] [PubMed] [Google Scholar]
- Metcalfe DJ. (2005) Hedera helix L. J Ecol 93: 632–648 [Google Scholar]
- Nardini A, Raimondo F, Lo Gullo MA, Salleo S. (2010) Leafminers help us understand leaf hydraulic design. Plant Cell Environ 33: 1091–1100 [DOI] [PubMed] [Google Scholar]
- Nardini A, Salleo S. (2003) Effects of the experimental blockage of the major veins on hydraulics and gas exchange of Prunus laurocerasus L. leaves. J Exp Bot 54: 1213–1219 [DOI] [PubMed] [Google Scholar]
- Nardini A, Salleo S, Raimondo F. (2003) Changes in leaf hydraulic conductance correlate with leaf vein embolism in Cercis siliquastrum L. Trees Structure and Function 17: 529–534 [Google Scholar]
- Nardini A, Tyree MT, Salleo S. (2001) Xylem cavitation in the leaf of Prunus laurocerasus and its impact on leaf hydraulics. Plant Physiol 125: 1700–1709 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nicotra AB, Cosgrove MJ, Cowling A, Schlichting CD, Jones CS. (2008) Leaf shape linked to photosynthetic rates and temperature optima in South African Pelargonium species. Oecologia 154: 625–635 [DOI] [PubMed] [Google Scholar]
- Niinemets U, Portsmuth A, Tobias M. (2006) Leaf size modifies support biomass distribution among stems, petioles and mid-ribs in temperate plants. New Phytol 171: 91–104 [DOI] [PubMed] [Google Scholar]
- Niinemets U, Portsmuth A, Tobias M. (2007) Leaf shape and venation pattern alter the support investments within leaf lamina in temperate species: a neglected source of leaf physiological differentiation? Funct Ecol 21: 28–40 [Google Scholar]
- Niklas KJ. (1999) A mechanical perspective on foliage leaf form and function. New Phytol 143: 19–31 [Google Scholar]
- Nobel PS. (1976) Photosynthetic rates of sun versus shade leaves of Hyptis emoryi Torr. Plant Physiol 58: 218–223 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nobel PS. (2010) Physicochemical and Environmental Plant Physiology, Ed 4. Academic Press, San Diego [Google Scholar]
- Nobel PS, Jordan PW. (1983) Transpiration stream of desert species: resistances and capacitances for a C3, a C4 and a CAM plant. J Exp Bot 34: 1379–1391 [Google Scholar]
- Ogburn RM, Edwards EJ. (2009) Anatomical variation in Cactaceae and relatives: trait lability and evolutionary innovation. Am J Bot 96: 391–408 [DOI] [PubMed] [Google Scholar]
- Parkhurst DF, Loucks OL. (1972) Optimal leaf size in relation to environment. J Ecol 60: 505–537 [Google Scholar]
- Roth-Nebelsick A, Uhl D, Mosbrugger V, Kerp H. (2001) Evolution and function of leaf venation architecture: a review. Ann Bot (Lond) 87: 553–566 [Google Scholar]
- Sack L, Cowan PD, Holbrook NM. (2003a) The major veins of mesomorphic leaves revisited: tests for conductive overload in Acer saccharum (Aceraceae) and Quercus rubra (Fagaceae). Am J Bot 90: 32–39 [DOI] [PubMed] [Google Scholar]
- Sack L, Cowan PD, Jaikumar N, Holbrook NM. (2003b) The ‘hydrology’ of leaves: co-ordination of structure and function in temperate woody species. Plant Cell Environ 26: 1343–1356 [Google Scholar]
- Sack L, Dietrich EM, Streeter CM, Sánchez-Gómez D, Holbrook NM. (2008) Leaf palmate venation and vascular redundancy confer tolerance of hydraulic disruption. Proc Natl Acad Sci USA 105: 1567–1572 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sack L, Frole K. (2006) Leaf structural diversity is related to hydraulic capacity in tropical rain forest trees. Ecology 87: 483–491 [DOI] [PubMed] [Google Scholar]
- Sack L, Grubb PJ, Maranon T. (2003c) The functional morphology of juvenile plants tolerant of strong summer drought in shaded forest understories in southern Spain. Plant Ecol 168: 139–163 [Google Scholar]
- Sack L, Holbrook NM. (2006) Leaf hydraulics. Annu Rev Plant Biol 57: 361–381 [DOI] [PubMed] [Google Scholar]
- Sack L, Melcher PJ, Liu WH, Middleton E, Pardee T. (2006) How strong is intracanopy leaf plasticity in temperate deciduous trees? Am J Bot 93: 829–839 [DOI] [PubMed] [Google Scholar]
- Sack L, Melcher PJ, Zwieniecki MA, Holbrook NM. (2002) The hydraulic conductance of the angiosperm leaf lamina: a comparison of three measurement methods. J Exp Bot 53: 2177–2184 [DOI] [PubMed] [Google Scholar]
- Sack L. (2010) Leaf pressure-volume curve parameters. PrometheusWiki. http://prometheuswiki.publish.csiro.au/tiki-index.php?page=PROTOCOL%3A+Leaf+pressure-volume+curve+parameters (April 18, 2011)
- Sack L, Scoffoni C. (2010) Minimum epidermal conductance (gmin a.k.a. cuticular conductance). PrometheusWiki. http://prometheuswiki.publish.csiro.au/tiki-index.php?page=PROTOCOL%3A+Minimum+epidermal+conductance+%28gmin%2C+a.k.a.+cuticular+conductance%29&highlight=cuticular (April 18, 2011)
- Salleo S, Lo Gullo MA, Raimondo F, Nardini A. (2001) Vulnerability to cavitation of leaf minor veins: any impact on leaf gas exchange? Plant Cell Environ 24: 851–859 [Google Scholar]
- Salleo S, Nardini A, Lo Gullo MA, Ghirardelli LA. (2002) Changes in stem and leaf hydraulics preceding leaf shedding in Castanea sativa L. Biol Plant 45: 227–234 [Google Scholar]
- Salleo S, Nardini A, Pitt F, Lo Gullo MA. (2000) Xylem cavitation and hydraulic control of stomatal conductance in laurel (Laurus nobilis L.). Plant Cell Environ 23: 71–79 [Google Scholar]
- Salleo S, Raimondo F, Trifilo P, Nardini A. (2003) Axial-to-radial water permeability of leaf major veins: a possible determinant of the impact of vein embolism on leaf hydraulics? Plant Cell Environ 26: 1749–1758 [Google Scholar]
- Santiago LS, Kim SC. (2009) Correlated evolution of leaf shape and physiology in the woody Sonchus alliance (Asteraceae: Sonchinae) in Macaronesia. Int J Plant Sci 170: 83–92 [Google Scholar]
- Schaefer J, Opgen-Rhein R, Strimmer K. (2007) corpcor: efficient estimation of covariance and (partial) correlation. R Package Version 1.4.7. http://cran.r-project.org/web/packages/corpcor/ (April 18, 2011) [Google Scholar]
- Scoffoni C, Pou A, Aasamaa K, Sack L. (2008) The rapid light response of leaf hydraulic conductance: new evidence from two experimental methods. Plant Cell Environ 31: 1803–1812 [DOI] [PubMed] [Google Scholar]
- Shipley B. (2000) Cause and Correlation in Biology. Cambridge University Press, Cambridge, UK [Google Scholar]
- Sokal RR, Rohlf FJ. (1995) Biometry: The Principles and Practice of Statistics in Biological Research, Ed 3. W.H. Freeman, New York [Google Scholar]
- Sperry JS. (2003) Evolution of water transport and xylem structure. Int J Plant Sci 164: S115–S127 [Google Scholar]
- Sperry JS, Hacke UG, Wheeler JK. (2005) Comparative analysis of end wall resistivity in xylem conduits. Plant Cell Environ 28: 456–465 [Google Scholar]
- Sultan SE, Bazzaz FA. (1993) Phenotypic plasticity in Polygonum persicaria. II. Norms of reaction to soil-moisture and the maintenance of genetic diversity. Evolution 47: 1032–1049 [DOI] [PubMed] [Google Scholar]
- Trifilò P, Gascó A, Raimondo F, Nardini A, Salleo S. (2003a) Kinetics of recovery of leaf hydraulic conductance and vein functionality from cavitation-induced embolism in sunflower. J Exp Bot 54: 2323–2330 [DOI] [PubMed] [Google Scholar]
- Trifilo P, Nardini A, Lo Gullo MA, Salleo S. (2003b) Vein cavitation and stomatal behaviour of sunflower (Helianthus annuus) leaves under water limitation. Physiol Plant 119: 409–417 [Google Scholar]
- Tyree MT, Hammel HT. (1972) Measurement of turgor pressure and water relations of plants by pressure bomb technique. J Exp Bot 23: 267–282 [Google Scholar]
- Uhl D, Mosbrugger V. (1999) Leaf venation density as a climate and environmental proxy: a critical review and new data. Palaeogeogr Palaeoclimatol Palaeoecol 149: 15–26 [Google Scholar]
- Vogel S. (1968) “Sun leaves” and “shade leaves”: differences in convective heat dissipation. Ecology 49: 1203–1204 [Google Scholar]
- Vogel S. (1970) Convective cooling at low airspeeds and shapes of broad leaves. J Exp Bot 21: 91–101 [Google Scholar]
- Vogel S. (2009) Leaves in the lowest and highest winds: temperature, force and shape. New Phytol 183: 13–26 [DOI] [PubMed] [Google Scholar]
- Wilf P. (1997) When are leaves good thermometers? A new case for leaf margin analysis. Paleobiology 23: 373–390 [Google Scholar]
- Witkowski ETF, Lamont BB. (1991) Leaf specific mass confounds leaf density and thickness. Oecologia 88: 486–493 [DOI] [PubMed] [Google Scholar]
- Wylie RB. (1938) Concerning the conductive capacity of the minor veins of foliage leaves. Am J Bot 25: 567–572 [Google Scholar]
- Yates MJ, Verboom GA, Rebelo AG, Cramer MD. (2010) Ecophysiological significance of leaf size variation in Proteaceae from the Cape Floristic region. Funct Ecol 24: 485–492 [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.