

Abstract
Obesity has received much attention worldwide in association with an increased risk of cardiovascular diseases, diabetes, and cancer. At present, bariatric surgery is the only effective treatment for obesity in which long-term weight loss is achieved in patients. By contrast, pharmacological interventions for obesity are usually followed by weight regain. Although the exact mechanisms of long-term weight loss following bariatric surgery are yet to be fully elucidated, several gut hormones have been implicated. Gut hormones play a critical role in relaying signals of nutritional and energy status from the gut to the central nervous system, in order to regulate food intake. Cholecystokinin, peptide YY, pancreatic polypeptide, glucagon-like peptide-1, and oxyntomodulin act through distinct yet synergistic mechanisms to suppress appetite, whereas ghrelin stimulates food intake. Here, we discuss the role of gut hormones in the regulation of food intake and body weight.
1. Introduction
Obesity is one of the major health challenges throughout the world, due to its association with an array of vascular, metabolic, and psychosocial complications [1, 2]. Obesity is traditionally associated with populations in Europe and North America; however Asian countries such as Japan have recently reported increasing prevalences of obesity, which may reflect changes in dietary patterns and lifestyles [3, 4].
Obesity is a state in which energy intake chronically exceeds energy expenditure. Body weight is tightly regulated by complex homeostatic mechanisms involving the hypothalamus and brainstem which integrate inputs from higher cortical centres with peripherally derived signals of the body's nutritional and energy status. In the hypothalamic arcuate nucleus (ARC), there are two neuronal populations with opposing effects on food intake: neurons which coexpress neuropeptide Y (NPY) and agouti-related peptide (AgRP) which stimulate food intake, whereas neurons coexpressing proopiomelanocortin (POMC) and cocaine- and amphetamine-regulated transcript (CART) suppress food intake (see Figure 1). Within the brainstem, the dorsal vagal complex (DVC) consisting of the dorsal motor nucleus of vagus (DVN), area postrema (AP), and the nucleus of the tractus solitarius (NTS) plays a pivotal role in relaying of peripheral signals such as vagal afferents from the gut to the hypothalamus [5]. In human, higher cortical centres are implicated in psychological and emotional factors which can drive food intake beyond homeostatic requirements. In addition, the corticolimbic pathways are responsible for reward-associated feeding behaviour.
Figure 1.
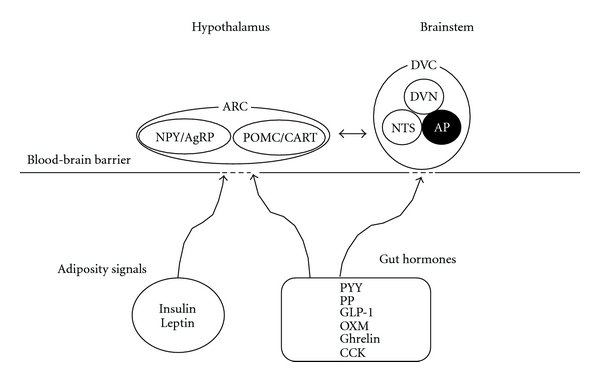
Humoral signals implicated in the physiological regulation of food intake. Diagram summarising the major signalling pathways which converge on the hypothalamus and brainstem in order to regulate food intake. ARC, arcuate nucleus; NPY/AgRP, neuropeptide Y and agouti-related peptide; POMC/CART, proopiomelanocortin and cocaine- and amphetamine-regulated transcript; DVC, dorsal vagal complex; DVN, the dorsal motor nucleus of vagus; NTS, the nucleus of the tractus solitarius; AP, area postrema; GLP-1, glucagon-like peptide-1; CCK, cholecystokinin; PP, pancreatic polypeptide; PYY, peptide YY; OXM, oxyntomodulin.
This article summarises our current understanding of the role of gut hormones in appetite regulation and its potential as therapeutic targets for obesity.
2. Gastrointestinal Tract
More than 30 gut hormone genes are known to be expressed, and more than 100 bioactive peptides are distributed in the gastrointestinal tract, which is thus regarded as the largest endocrine organ in the body [6]. Meal anticipation and the presence of food in the upper gastrointestinal tract stimulate the release of gut hormones and neurotransmitters from the gut. These neurohumoral signals are involved in the initiation and maintenance of food intake as well as termination of meals. The satiating effect of stomach distension is revealed by observing that infusion of either saline or nutrients into the rat stomach results in same reduction in food intake [7]. In humans, the effects of intragastric balloon insertion on body weight and food intake are conflicting [8, 9]; this may reflect differences in the types of balloon used during these studies.
Both meal duration and size are markedly increased during sham feeding, where ingested food is prevented from distending the stomach or small intestine by surgical intervention, whereas intraluminal gastrointestinal (GI) infusion of macronutrient before food access reduces subsequent meal size in a dose-dependent manner. These findings suggest that the upper GI tract has an important role in negative feedback regulation of food intake, and the upper intestine is critical for nutrient absorption [10]. The vagus nerve is closely implicated in the transmission of the food-induced negative feedback signals which are critical for determining meal size. Transection of all gut sensory vagal fibres results in increased meal size and meal duration but does not block gastric preload-induced feeding suppression; this implies that vagal afferent signals contribute to satiety during spontaneous meals [10, 11].
Perfusion of nutrients into the colon inhibits upper gastrointestinal secretion, motility, and transit; this negative feedback mechanism has been called the “ileal brake” [12]. Fat is the most potent trigger of the ileal brake, and glucagon-like peptide-1 (GLP-1) and peptide YY (PYY) may be among the mediators of this phenomenon [13].
3. Bariatric Surgery and Weight Loss
Almost all pharmacological and behavioural treatments for obesity result in weight loss followed by weight regain [14]. In contrast, gastric bypass surgery is an established and effective treatment for obesity which provides sustained weight loss maintenance for at least 15 years [15, 16].
Bariatric surgery may be classified into malabsorptive surgery and restrictive surgery. Malabsorption-based techniques include the jejuno-ileal bypass, allowing nutrients to pass directly from the proximal jejunum to the terminal ileum, and roux-en-Y gastric bypass (RYGB), combining restrictive and malabsorptive procedures. In some cases, the former procedure may be associated with severe complications [17].
Restrictive bariatric surgery involves the laparoscopic application of an adjustable gastric banding (LAGB). LAGB procedures result in slightly less weight loss compared with RYGB but have been reported to be safer [18]. Intriguingly, the mechanisms of long-term weight loss following bariatric surgery are yet to be determined; however several gut hormones have been implicated as contributory factors; a decrease in ghrelin and an increase in PYY and GLP-1 levels have been found following bypass surgery [19–21]. Recent evidence suggests that an increase in energy expenditure may play a role in part in weight loss after gastric bypass surgery as an additional factor [22]. Gastric bypass surgery also improves glycaemic control in patients with type 2 diabetes, and this often occurs prior to observable weight loss [23]. Thus emulating the altered gut hormone signals associated with bypass surgery may offer promising novel treatments for obesity.
4. Gut Microbiota
Recently, a potential link between gut microbiota and obesity has emerged [24]. The human gut harbours a large number of bacterial microorganisms collectively termed gut microbiota. Bäckhed et al. [25] have observed that adult germ-free mice had 40% less total body fat than mice with normal microbiota; furthermore replacing the microbiota in adult germ-free mice produced a 60% increase in body fat content and insulin resistance within 14 days of replacement. In addition, germ-free mice may be protected against high fat diet-induced metabolic changes [26]. There has also been considerable interest in the potential benefits of prebiotic drinks containing bacterial cultures. The study by Cani et al. [27] evaluated the effect of prebiotics on plasma levels of gut hormones in healthy subjects. After two weeks of prebiotic treatment, they observed increased gut microbiota fermentation, decreased appetite, and improved postprandial glucose responses. Furthermore plasma levels of GLP-1 and PYY were increased in subjects following prebiotic treatment. Current data therefore suggest that gut microbiota may promote the development of obesity and that manipulation of gut microbiota using probiotics may alter gut endocrine function. Further studies are required in order to further investigate the pathophysiological basis of the association between gut microbiota and energy homeostasis.
5. Gut Hormones
5.1. PP-Fold Protein
The PP-fold family consists of neuropeptide Y (NPY), peptide YY (PYY), and pancreatic polypeptide (PP). PYY and PP are secreted from gastrointestinal tract, whereas NPY is predominantly distributed within central nervous system [28]. The members of PP-fold family act via G protein-coupled receptors: Y1, Y2, Y4, Y5, and Y6 [29].
5.2. Peptide Tyrosine Tyrosine (PYY)
PYY was first isolated as a 36-amino acid peptide from porcine upper small intestine [28]. PYY is released by L cells of the distal gut. There are two circulating forms of PYY: PYY (1–36) and PYY (3–36). PYY (3–36), the major circulating form, is produced by cleavage of the N-terminal Tyrosine-Proline residues from PYY (1–36) by the enzyme dipeptidyl-peptidase IV (DPPIV) [30]. PYY (1–36) has affinity to all Y receptors, while PYY (3–36) binds with highest affinity to the hypothalamic Y2 receptor, suppressing food intake. Circulating PYY concentrations are low in fasted state and rise rapidly following a meal with a peak at 1-2 hours and remain elevated for several hours [31]. Ingestion of fat results in greater release of PYY than observed with ingestion of carbohydrate or protein meals with a similar calorie content [31]. Peripheral PYY administration shows a decrease in food intake and body weight gain in rats [32]. In both lean and obese humans, intravenous injection of PYY reduces appetite and food intake [32, 33], suggesting that, unlike leptin, the sensitivity of PYY is preserved in obese subjects. PYY levels are found to be elevated in patients with gastrointestinal disorders including inflammatory bowel disease and steatorrhea [34, 35].
In addition to its effect on food intake, PYY may regulate energy expenditure [36, 37], delay gastric emptying, and reduce acid secretion [38, 39]. In obese subjects circulating PYY levels are low [33, 40], and PYY levels are reported to be higher in patients with anorexia nervosa when compared with control subjects [41]. Studies of circulating levels of PYY in obese and lean people have yielded conflicting results [42, 43]. However a blunted postprandial rise in PYY is observed in obese people, which suggests its association with impaired satiety [39].
The anorectic effects of PYY (3–36) may be mediated centrally via a direct action in the ARC, through an indirect action involving the vagus and brainstem, or by a combination of both pathways. Peripheral administration of PYY (3–36) increases c-fos expression (a marker of neuronal activation) in the ARC, and direct injection of PYY (3–36) into the ARC inhibits food intake. This effect is most likely mediated through the Y2 receptor since the anorectic effect of peripheral PYY (3–36) administration is blocked in Y2 receptor-null mice and intra-arcuate injection of a Y2 receptor selective agonist also reduces food intake [32]. Although there have been conflicting results [44], the importance of vagal-brainstem signalling to the actions of PYY on food intake is suggested by observing that bilateral subdiaphragmatic vagotomy and brainstem-hypothalamic pathway transectioning abolish the anorectic effect of peripheral PYY (3–36) administration in rats [45, 46].
In contrast to peripheral and intra-arcuate PYY (3–36) administration, PYY (3–36) results in an increase in food intake when administered directly into the third ventricle of the brain [47] or directly into the paraventricular nucleus (PVN) [48]. This apparently confusing observation may be explained by considering that such effects might be endogenously mediated by the CNS-distributed peptide, NPY, through an action on Y1 receptor and Y5 receptors [49].
PYY may also act in areas of the brain other than the hypothalamus and brainstem. Batterham et al. [50] suggested that PYY (3–36) infusion modulates neural activity within corticolimbic and higher cortical brain using functional magnetic resonance imaging. Under conditions of high circulating PYY (3–36) designed to mimic the postprandial state, changes in neural activity within the caudolateral orbital frontal cortex predicted subsequent feeding behaviour. By contrast, hypothalamic activation correlated with food intake under conditions of low circulating PYY (3–36).
5.3. Pancreatic Polypeptide (PP)
PP is secreted from PP cells in the pancreatic islets of Langerhans. PP appears to reduce food intake directly through the Y4 receptor in the brainstem and hypothalamus. The anorectic effects of PP are abolished by vagotomy in rodents, suggesting that PP may also act via the vagus nerve to reduce food intake [51]. Y4 receptor expression is found in the AP, NTS, DVN, ARC, and PVN [52]. An autoradiography study also identified saturable PP binding sites at the interpeduncular nucleus, AP, NTS, and DVN [53], thus suggesting the brainstem as the major site of action for PP. Like PYY, differential effects on food intake are observed following PP injection, depending on the route of administration. In contrast to the anorectic effects observed with peripheral PP administration, its central administration stimulates food intake [54]. These differential effects may be explained by activation of distinct receptor populations, although the exact mechanism is not yet clear.
Circulating PP concentrations rise after a meal in proportion to the calorific load. Although reported differences in circulating levels of PP between lean and obese people have been conflicting [55, 56], some studies have demonstrated significantly reduced levels in obese subjects [57, 58]. Levels of PP are elevated after a test meal in anorexia [59]. Patients with Prader-Willi syndrome (PWS) have been reported to show reduced PP release both basally and postprandially when compared with age- and weight-matched control subjects [60].
The anorectic effects of PP have been demonstrated in a number of experimental models. In mice, acute and chronic peripheral administration of PP reduces food intake [47, 57]. In leptin-deficient ob/ob mice, repeated intraperitoneal injection of PP decreases body weight gain and ameliorates insulin resistance and hyperlipidaemia [51]. Furthermore, transgenic mice overexpressing PP are lean and demonstrate reduced food intake compared with wild-type controls [61]. In normal-weight human subjects, intravenous infusion of PP results in a 25% reduction in 24-h food intake [62]. Furthermore twice-daily infusion of PP in volunteers with PWS caused a 12% reduction in food intake [63].
5.4. Proglucagon-Derived Peptides
GLP-1, GLP-2, oxyntomodulin (OXM), and glucagon are proglucagon-derived peptides. Proglucagon is expressed in the pancreas, L-cells of the small intestine, and in the NTS of the brainstem [64, 65]. Glucagon is produced in the pancreas, whereas OXM, GLP-1, and GLP-2 are the major products in the brain and intestine [66].
5.5. Glucagon-Like Peptide-1
GLP-1 is cosecreted with PYY from L cells in the intestine in response to nutrient intake. Enzymatic degradation by DPPIV and renal clearance rapidly inactivate and remove GLP-1 from plasma circulation, respectively [67, 68], thus accounting for its short plasma half-life of 1-2 minutes [69]. GLP-1 has two biologically active forms, GLP-1 (7–37) and GLP-1 (7–36) amide, the latter being the major circulating form in humans [70]. GLP-1 exerts its effect at the GLP-1R to stimulate adenylyl cyclase activity and thereby cAMP production [71]. GLP-1R is widely distributed particularly in the brain, GI tract, and pancreas [71, 72]. Circulating GLP-1 levels rise after a meal and fall in the fasted state. Recent evidence also suggests that levels rise in anticipation of a meal [73]. GLP-1 reduces food intake, suppresses glucagon secretion, and delays gastric emptying [74]. Intravenous infusion of GLP-1 results in a dose-dependent reduction of food intake in both normal weight and obese subjects [75]; however obese subjects have a blunted postprandial GLP-1 response compared to lean subjects [71].
GLP-1 possesses a potent incretin effect in addition to its anorectic action; it stimulates insulin secretion in a glucose-dependent manner following ingestion of carbohydrate. Continuous subcutaneous infusion of GLP-1 to patients with type 2 diabetes for 6 weeks reduces appetite, body weight and improves glycaemic control [76]. Exendin-4, a naturally occurring peptide from the saliva of the Gila monster lizard, is a DPPIV-resistant GLP-1R agonist [77]. Exendin-4 (exenatide, Byetta) has been approved for type 2 diabetes in conjunction with either metformin, a sulphonylurea, or both drugs. Twice-daily subcutaneous injection of exendin-4 to type 2 diabetes patients failing to achieve glycaemic control with maximal doses of metformin improves glycaemic control and decreases body weight [78]. Once-weekly subcutaneous injection of a long-acting exenatide preparation and once-daily subcutaneous injection of the GLP-1 analogue called liraglutide demonstrate greater improvements in glycaemic control than twice-daily exenatide administration [79, 80].
GLP-1 possesses trophic effects on pancreatic beta cells in animal models [81]. Most recently, GLP-1 and exendin-4 have been shown to promote cellular growth and reduce apoptosis in nervous tissues [82]. GLP-1 receptor stimulation has been shown to have neuroprotective effect in models of Parkinson's disease [83–85], Alzheimer's disease [86, 87], cerebrovascular stroke [85], and peripheral neuropathy [88]. Further work is needed to determine if GLP-1 analogues could be potential novel therapies for patients with these neurological and neurodegenerative diseases.
5.6. Glucagon-Like Peptide-2
GLP-2 is released from enteroendocrine cells in a nutrient-dependent manner, like GLP-1. Acute or chronic administration with GLP-2 has no effect on food intake in either rodents or humans [89, 90]. However, GLP-2 has an intestinal trophic effect [91, 92], and chronic subcutaneous administration of GLP-2 stimulates crypt cell proliferation. As such, GLP-2 analogues have been developed for use in patients with inflammatory bowel disease [93]. In addition, some studies have demonstrated a reduction in gastric emptying in humans by GLP-2, although the effect is not as potent as GLP-1 [94].
5.7. Oxyntomodulin
OXM is another product of the proglucagon gene and is released from L-cells of the intestine in response to ingested food and in proportion to caloric intake [95]. Administration of OXM reduces food intake and increases energy expenditure in both rodents and humans [96–98]. The anorectic effect of OXM is blocked by the GLP-1R antagonist exendin 9–39 [99] and is abolished in GLP-1R null mice [100]; this suggests that OXM mediates its effects via the GLP-1R. However OXM has relatively low in vitro affinity for the GLP-1R which is 50 fold lower than the affinity of GLP-1 for GLP1R. This raises the possibility that a further receptor through which OXM mediates its anorectic effect has yet to be identified. Indeed, several actions of OXM appear to be independent of the GLP-1R [97, 101, 102]. For example, the cardiovascular effects of OXM are preserved in GLP-1R knockout mice [101]. Like GLP-1, OXM is inactivated by DPPIV; hence OXM analogues resistant to DPPIV degradation are being developed as potential obesity treatments [103].
5.8. Ghrelin
Ghrelin is the only known orexigenic gut hormone and was identified as an endogenous ligand for the growth hormone secretagogue receptor (GHS-R) in rat stomach [104]. Lower levels of ghrelin are also localized to the hypothalamic ARC. Levels of circulating ghrelin increase preprandially and fall rapidly in the postprandial period [105]. Both central and peripheral administrations of ghrelin increase food intake and body weight with a reduction in fat utilisation in rodents [106, 107]. In human, negative correlations between circulating ghrelin levels and body mass index are found. Fasting plasma levels of ghrelin are high in patients with anorexia nervosa [108] and in subjects with diet-induced weight loss [19]. By contrast, obese subjects display a less marked drop in plasma ghrelin after meal injection [109]. Plasma ghrelin levels are elevated in cachectic patients with heart failure as compared with noncachectic patients with heart failure and control subjects [110]. Furthermore elevated circulating ghrelin levels are observed in patients with PWS compared with individuals with nonsyndromic forms of obesity [111]. Dysregulation of ghrelin secretion is also implicated in the mechanism through which sleep disturbance contributes to obesity. Subjects with short sleep duration have elevated ghrelin levels, reduced leptin, and high body mass index compared with subjects with normal sleep duration [112].
Evidence suggests that ghrelin mediates its orexigenic action via stimulation of NPY/AgRP coexpressing neurons within the ARC of hypothalamus. Peripheral administration of ghrelin increases c-fos expression in ARC NPY/AgRP neurons [113], and ablation of both AgRP and NPY neurons completely abolishes the orexigenic effect of ghrelin [114]. Brainstem and vagus nerve may also contribute to the effects of ghrelin on food intake. Intracerebroventricular injection of ghrelin induces c-fos expression in NTS and AP [115]. GHS-R is expressed in the vagus nerve. Furthermore blockade of gastric vagal afferents in rats abolishes ghrelin-induced feeding and prevents the ghrelin-induced rise in c-fos expression within the ARC [116].
Ghrelin may promote food intake in part by enhancing the hedonic responses to food cues. The recent study by Malik et al. [117] using functional magnetic resonance imaging during exposure to food pictures revealed increased activation in the amygdala, orbitofrontal cortex, anterior insula, and striatum, during intravenous infusion of ghrelin. Furthermore the effects of ghrelin on the response of amygdala and orbitofrontal cortex were correlated with self-rated hunger ratings.
5.9. Cholecystokinin
CCK was the first gut hormone found to play a role in food intake [118]. CCK is secreted postprandially by the I cell of the small intestine into circulation [119], with a short plasma half-life of a few minutes. Plasma CCK levels rise within 15 minutes after meals [119]. CCK is reported to reduce food intake in human and rodents [119, 120]. There are two CCK receptor subtypes; CCK1 and CCK2 receptors, previously classified as CCK A and CCK B. The anorectic action of CCK appears to be mostly mediated via CCK1 receptors on the vagal nerve [121, 122]. CCK 1 and 2 receptors are widely distributed in brain including the brainstem and hypothalamus [123]. Intermittent prandial CCK infusion reduces meal size in rats but provokes a compensatory increase in meal frequency [124]. In addition, a 2-week continuous intraperitoneal infusion of CCK failed to suppress food intake at any time point [125].
5.10. Peripheral Adiposity Signals: Insulin and Leptin
Adiposity signals are involved in the long-term regulation of energy balance, while gut peptides modulate food intake on a meal-by-meal basis. Circulating levels of insulin and leptin are positively correlated with adipose tissue mass within the body and are implicated in the long-term regulation of energy balance. Insulin is synthesized in the β cells of the pancreas and secreted rapidly after a meal, with well-characterised hypoglycaemic effects [126]. However intracerebroventricular administration of insulin also results in a dose-dependent suppression of food intake and body weight gain in baboons and rodents [127, 128]. Insulin may therefore have an anorectic action in addition to its hypoglycaemic effects. Leptin is secreted by adipocytes with circulating levels proportional to fat mass [129] with a diurnal and pulsatile pattern, peaking at night [130]. Leptin administration alleviates the hyperphagia associated with congenital leptin deficiency. However obese subjects are resistant to leptin, which may account for its lack of effectiveness in such individuals [131, 132].
6. Conclusion
Obesity, the metabolic syndrome, and their associated risk factors of cardiovascular disease and diabetes mellitus are among the most important health issues facing modern economies regardless of geographical location. Recent work has revealed that energy homeostasis is maintained by an array of complex pathways. The presence of multiple, overlapping feeding mechanisms reflects the vital nature of feeding behaviour for survival. However these homeostatic feeding mechanisms may be viewed as maladaptive in obese individuals exposed to diets with a high calorific content. Several gut hormones are thought to play a role in the sustained weight loss observed following bypass surgery; hence mimicking bypass surgery by administration of gut hormone-derived therapies could offer a promising treatment for obesity. A further understanding of the pathogenesis of obesity and the role of gut hormones in appetite regulation is imperative.
Conflict of Interests
The authors declare that there is no conflict of interests.
References
- 1.Field AE, Coakley EH, Must A, et al. Impact of overweight on the risk of developing common chronic diseases during a 10-year period. Archives of Internal Medicine. 2001;161(13):1581–1586. doi: 10.1001/archinte.161.13.1581. [DOI] [PubMed] [Google Scholar]
- 2.Must A, Spadano J, Coakley EH, Field AE, Colditz G, Dietz WH. The disease burden associated with overweight and obesity. Journal of the American Medical Association. 1999;282(16):1523–1529. doi: 10.1001/jama.282.16.1523. [DOI] [PubMed] [Google Scholar]
- 3.Matsuzawa Y, Nakamura T, Takahashi M, et al. New criteria for ’obesity disease’ in Japan. Circulation Journal. 2002;66(11):987–992. doi: 10.1253/circj.66.987. [DOI] [PubMed] [Google Scholar]
- 4.Matsushita Y, Takahashi Y, Mizoue T, Inoue M, Noda M, Tsugane S. Overweight and obesity trends among Japanese adults: a 10-year follow-up of the JPHC Study. International Journal of Obesity. 2008;32(12):1861–1867. doi: 10.1038/ijo.2008.188. [DOI] [PubMed] [Google Scholar]
- 5.Bailey EF. A tasty morsel: the role of the dorsal vagal complex in the regulation of food intake and swallowing. Focus on “BDNF/TrkB signaling interacts with GABAergic system to inhibit rhythmic swallowing in the rat,” by Bariohay et al. American Journal of Physiology. 2008;295(4):R1048–R1049. doi: 10.1152/ajpregu.90701.2008. [DOI] [PubMed] [Google Scholar]
- 6.Rehfeld JF. The new biology of gastrointestinal hormones. Physiological Reviews. 1998;78(4):1087–1108. doi: 10.1152/physrev.1998.78.4.1087. [DOI] [PubMed] [Google Scholar]
- 7.Phillips RJ, Powley TL. Gastric volume rather than nutrient content inhibits food intake. American Journal of Physiology. 1996;271(3):R766–R779. doi: 10.1152/ajpregu.1996.271.3.R766. [DOI] [PubMed] [Google Scholar]
- 8.Martin CK, Bellanger DE, Rau KK, Coulon S, Greenway FL. Safety of the Ullorex oral intragastric balloon for the treatment of obesity. Journal of Diabetes Science and Technology. 2007;1(40):574–581. doi: 10.1177/193229680700100416. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Rigaud D, Trostler N, Rozen R, Vallot T, Apfelbaum T. Gastric distension, hunger and energy intake after balloon implantation in severe obesity. International Journal of Obesity. 1995;19(7):489–495. [PubMed] [Google Scholar]
- 10.Schwartz GJ. The role of gastrointestinal vagal afferents in the control of food intake: current prospects. Nutrition. 2000;16(10):866–873. doi: 10.1016/s0899-9007(00)00464-0. [DOI] [PubMed] [Google Scholar]
- 11.Schwartz GJ, Salorio CF, Skoglund C, Moran TH. Gut vagal afferent lesions increase meal size but do not block gastric preload-induced feeding suppression. American Journal of Physiology. 1999;276(6):R1623–R1629. doi: 10.1152/ajpregu.1999.276.6.R1623. [DOI] [PubMed] [Google Scholar]
- 12.Spiller RC, Trotman IF, Higgins BE. The ileal brake—inhibition of jejunal motility after ileal fat perfusion in man. Gut. 1984;25(4):365–374. doi: 10.1136/gut.25.4.365. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Wen J, Phillips SF, Sarr MG, Kost LJ, Holst JJ. PYY and GLP-1 contribute to feedback inhibition from the canine ileum and colon. American Journal of Physiology. 1995;269(6):G945–G952. doi: 10.1152/ajpgi.1995.269.6.G945. [DOI] [PubMed] [Google Scholar]
- 14.Yanovski SZ, Yanovski JA. Obesity. New England Journal of Medicine. 2002;346(8):591–602. doi: 10.1056/NEJMra012586. [DOI] [PubMed] [Google Scholar]
- 15.Brolin RE. Bariatric surgery and long-term control of morbid obesity. Journal of the American Medical Association. 2002;288(22):2793–2796. doi: 10.1001/jama.288.22.2793. [DOI] [PubMed] [Google Scholar]
- 16.Cummings DE, Overduin J, Foster-Schubert KE. Gastric bypass for obesity: mechanisms of weight loss and diabetes resolution. Journal of Clinical Endocrinology and Metabolism. 2004;89(6):2608–2615. doi: 10.1210/jc.2004-0433. [DOI] [PubMed] [Google Scholar]
- 17.Tadross JA, Le Roux CW. The mechanisms of weight loss after bariatric surgery. International Journal of Obesity. 2009;33(1):S28–S32. doi: 10.1038/ijo.2009.14. [DOI] [PubMed] [Google Scholar]
- 18.Favretti F, Ashton D, Busetto L, Segato G, De Luca M. The gastric band: first-choice procedure for obesity surgery. World Journal of Surgery. 2009;33(10):2039–2048. doi: 10.1007/s00268-009-0091-6. [DOI] [PubMed] [Google Scholar]
- 19.Cummings DE, Weigle DS, Scott Frayo R, et al. Plasma ghrelin levels after diet-induced weight loss or gastric bypass surgery. New England Journal of Medicine. 2002;346(21):1623–1630. doi: 10.1056/NEJMoa012908. [DOI] [PubMed] [Google Scholar]
- 20.Korner J, Inabnet W, Febres G, et al. Prospective study of gut hormone and metabolic changes after adjustable gastric banding and Roux-en-Y gastric bypass. International Journal of Obesity. 2009;33(7):786–795. doi: 10.1038/ijo.2009.79. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Le Roux CW, Welbourn R, Werling M, et al. Gut hormones as mediators of appetite and weight loss after Roux-en-Y gastric bypass. Annals of Surgery. 2007;246(5):780–785. doi: 10.1097/SLA.0b013e3180caa3e3. [DOI] [PubMed] [Google Scholar]
- 22.Bueter M, Löwenstein C, Olbers T, et al. Gastric bypass increases energy expenditure in rats. Gastroenterology. 2010;138(5):1845–1853.e1. doi: 10.1053/j.gastro.2009.11.012. [DOI] [PubMed] [Google Scholar]
- 23.Laville M, Disse E. Bariatric surgery for diabetes treatment: why should we go rapidly to surgery. Diabetes and Metabolism. 2009;35(6):562–563. doi: 10.1016/S1262-3636(09)73466-4. [DOI] [PubMed] [Google Scholar]
- 24.Musso G, Gambino R, Cassader M. Obesity, diabetes, and gut microbiota: the hygiene hypothesis expanded? Diabetes Care. 2010;33(10):2277–2284. doi: 10.2337/dc10-0556. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Bäckhed F, Ding H, Wang T, et al. The gut microbiota as an environmental factor that regulates fat storage. Proceedings of the National Academy of Sciences of the United States of America. 2004;101(44):15718–15723. doi: 10.1073/pnas.0407076101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Bäckhed F, Manchester JK, Semenkovich CF, Gordon JI. Mechanisms underlying the resistance to diet-induced obesity in germ-free mice. Proceedings of the National Academy of Sciences of the United States of America. 2007;104(3):979–984. doi: 10.1073/pnas.0605374104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Cani PD, Lecourt E, Dewulf EM, et al. Gut microbiota fermentation of prebiotics increases satietogenic and incretin gut peptide production with consequences for appetite sensation and glucose response after a meal. American Journal of Clinical Nutrition. 2009;90(5):1236–1243. doi: 10.3945/ajcn.2009.28095. [DOI] [PubMed] [Google Scholar]
- 28.Tatemoto K, Mutt V. Isolation of two novel candidate hormones using a chemical method for finding naturally occurring polypeptides. Nature. 1980;285(5764):417–418. doi: 10.1038/285417a0. [DOI] [PubMed] [Google Scholar]
- 29.Lin S, Boey D, Herzog H. NPY and Y receptors: lessons from transgenic and knockout models. Neuropeptides. 2004;38(4):189–200. doi: 10.1016/j.npep.2004.05.005. [DOI] [PubMed] [Google Scholar]
- 30.Eberlein GA, Eysselein VE, Schaeffer M, et al. A new molecular form of PYY: structural characterization of human PYY(3-36) and PYY(1-36) Peptides. 1989;10(4):797–803. doi: 10.1016/0196-9781(89)90116-2. [DOI] [PubMed] [Google Scholar]
- 31.Adrian TE, Ferri GL, Bacarese-Hamilton AJ. Human distribution and release of a putative new gut hormone, peptide YY. Gastroenterology. 1985;89(5):1070–1077. doi: 10.1016/0016-5085(85)90211-2. [DOI] [PubMed] [Google Scholar]
- 32.Batterham RL, Cowley MA, Small CJ, et al. Gut hormone PYY3-36 physiologically inhibits food intake. Nature. 2002;418(6898):650–654. doi: 10.1038/nature00887. [DOI] [PubMed] [Google Scholar]
- 33.Batterham RL, Cohen MA, Ellis SM, et al. Inhibition of food intake in obese subjects by peptide YY3-36. New England Journal of Medicine. 2003;349(10):941–948. doi: 10.1056/NEJMoa030204. [DOI] [PubMed] [Google Scholar]
- 34.Adrian TE, Savage AP, Bacarese-Hamilton AJ. Peptide YY abnormalities in gastrointestinal diseases. Gastroenterology. 1986;90(2):379–384. doi: 10.1016/0016-5085(86)90936-4. [DOI] [PubMed] [Google Scholar]
- 35.Wahab PJ, Hopman WPM, Jansen JBMJ. Basal and fat-stimulated plasma peptide YY levels in celiac disease. Digestive Diseases and Sciences. 2001;46(11):2504–2509. doi: 10.1023/a:1012344424300. [DOI] [PubMed] [Google Scholar]
- 36.Sloth B, Holst JJ, Flint A, Gregersen NT, Astrup A. Effects of PYY1-36 and PYY3-36 on appetite, energy intake, energy expenditure, glucose and fat metabolism in obese and lean subjects. American Journal of Physiology. 2007;292(4):E1062–E1068. doi: 10.1152/ajpendo.00450.2006. [DOI] [PubMed] [Google Scholar]
- 37.Boey D, Lin S, Enriquez RF, et al. PYY transgenic mice are protected against diet-induced and genetic obesity. Neuropeptides. 2008;42(1):19–30. doi: 10.1016/j.npep.2007.11.003. [DOI] [PubMed] [Google Scholar]
- 38.Talsania T, Anini Y, Siu S, Drucker DJ, Brubaker PL. Peripheral exendin-4 and peptide YY3-36 synergistically reduce food intake through different mechanisms in mice. Endocrinology. 2005;146(9):3748–3756. doi: 10.1210/en.2005-0473. [DOI] [PubMed] [Google Scholar]
- 39.Ashby D, Bloom SR. Recent progress in PYY research—an update report for 8th NPY meeting. Peptides. 2007;28(2):198–202. doi: 10.1016/j.peptides.2006.10.019. [DOI] [PubMed] [Google Scholar]
- 40.Bartolomé MA, Borque M, Martinez-Sarmiento J, et al. Peptide YY secretion in morbidly obese patients before and after vertical banded gastroplasty. Obesity Surgery. 2002;12(3):324–327. doi: 10.1381/096089202321088084. [DOI] [PubMed] [Google Scholar]
- 41.Misra M, Miller KK, Tsai P, et al. Elevated peptide YY levels in adolescent girls with anorexia nervosa. Journal of Clinical Endocrinology and Metabolism. 2006;91(3):1027–1033. doi: 10.1210/jc.2005-1878. [DOI] [PubMed] [Google Scholar]
- 42.Kim BJ, Carlson OD, Jang HJ, Elahi D, Berry C, Egan JM. Peptide YY is secreted after oral glucose administration in a gender-specific manner. Journal of Clinical Endocrinology and Metabolism. 2005;90(12):6665–6671. doi: 10.1210/jc.2005-0409. [DOI] [PubMed] [Google Scholar]
- 43.Pfluger PT, Kampe J, Castaneda TR, et al. Effect of human body weight changes on circulating levels of peptide YY and peptide YY3-36. Journal of Clinical Endocrinology and Metabolism. 2007;92(2):583–588. doi: 10.1210/jc.2006-1425. [DOI] [PubMed] [Google Scholar]
- 44.Halatchev IG, Cone RD. Peripheral administration of PYY3-36 produces conditioned taste aversion in mice. Cell Metabolism. 2005;1(3):159–168. doi: 10.1016/j.cmet.2005.02.003. [DOI] [PubMed] [Google Scholar]
- 45.Abbott CR, Monteiro M, Small CJ, et al. The inhibitory effects of peripheral administration of peptide YY 3-36 and glucagon-like peptide-1 on food intake are attenuated by ablation of the vagal-brainstem-hypothalamic pathway. Brain Research. 2005;1044(1):127–131. doi: 10.1016/j.brainres.2005.03.011. [DOI] [PubMed] [Google Scholar]
- 46.Koda S, Date Y, Murakami N, et al. The role of the vagal nerve in peripheral PYY3-36-induced feeding reduction in rats. Endocrinology. 2005;146(5):2369–2375. doi: 10.1210/en.2004-1266. [DOI] [PubMed] [Google Scholar]
- 47.Morley JE, Levine AS, Grace M, Kneip J. Peptide YY (PYY), a potent orexigenic agent. Brain Research. 1985;341(1):200–203. doi: 10.1016/0006-8993(85)91490-8. [DOI] [PubMed] [Google Scholar]
- 48.Stanley BG, Daniel DR, Chin AS, Leibowitz SF. Paraventricular nucleus injections of peptide YY and neuropeptide Y preferentially enhance carbohydrate ingestion. Peptides. 1985;6(6):1205–1211. doi: 10.1016/0196-9781(85)90452-8. [DOI] [PubMed] [Google Scholar]
- 49.Kanatani A, Mashiko S, Murai N, et al. Role of the Y1 receptor in the regulation of neuropeptide Y-mediated feeding: comparison of wild-type, Y1 receptor-deficient, and Y5 receptor-deficient mice. Endocrinology. 2000;141(3):1011–1016. doi: 10.1210/endo.141.3.7387. [DOI] [PubMed] [Google Scholar]
- 50.Batterham RL, Ffytche DH, Rosenthal JM, et al. PYY modulation of cortical and hypothalamic brain areas predicts feeding behaviour in humans. Nature. 2007;450(7166):106–109. doi: 10.1038/nature06212. [DOI] [PubMed] [Google Scholar]
- 51.Asakawa A, Inui A, Yuzuriha H, et al. Characterization of the effects of pancreatic polypeptide in the regulation of energy balance. Gastroenterology. 2003;124(5):1325–1336. doi: 10.1016/s0016-5085(03)00216-6. [DOI] [PubMed] [Google Scholar]
- 52.Parker RMC, Herzog H. Regional distribution of Y-receptor subtype mRNAs in rat brain. European Journal of Neuroscience. 1999;11(4):1431–1448. doi: 10.1046/j.1460-9568.1999.00553.x. [DOI] [PubMed] [Google Scholar]
- 53.Whitcomb DC, Taylor IL, Vigna SR. Characterization of saturable binding sites for circulating pancreatic polypeptide in rat brain. American Journal of Physiology. 1990;259(4):G687–G691. doi: 10.1152/ajpgi.1990.259.4.G687. [DOI] [PubMed] [Google Scholar]
- 54.Clark JT, Kalra PS, Crowley WR, Kalra SP. Neuropeptide Y and human pancreatic polypeptide stimulate feeding behavior in rats. Endocrinology. 1984;115(1):427–429. doi: 10.1210/endo-115-1-427. [DOI] [PubMed] [Google Scholar]
- 55.Jorde R, Burhol PG. Fasting and postprandial plasma pancreatic polypeptide (PP) levels in obesity. International Journal of Obesity. 1984;8(5):393–397. [PubMed] [Google Scholar]
- 56.Wisen O, Bjorvell H, Cantor P, Johansson C, Theodorsson E. Plasma concentrations of regulatory peptides in obesity following modified sham feeding (MSF) and a liquid test meal. Regulatory Peptides. 1992;39(1):43–54. doi: 10.1016/0167-0115(92)90007-h. [DOI] [PubMed] [Google Scholar]
- 57.Glaser B, Zoghlin G, Pienta K, Vinik AI. Pancreatic polypeptide response to secretin in obesity: effects of glucose intolerance. Hormone and Metabolic Research. 1988;20(5):288–292. doi: 10.1055/s-2007-1010817. [DOI] [PubMed] [Google Scholar]
- 58.Lassmann V, Vague P, Vialettes B, Simon MC. Low plasma levels of pancreatic polypeptide in obesity. Diabetes. 1980;29(6):428–430. doi: 10.2337/diab.29.6.428. [DOI] [PubMed] [Google Scholar]
- 59.Uhe AM, Szmukler GI, Collier GR, Hansky J, O’Dea K, Young GP. Potential regulators of feeding behavior in anorexia nervosa. American Journal of Clinical Nutrition. 1992;55(1):28–32. doi: 10.1093/ajcn/55.1.28. [DOI] [PubMed] [Google Scholar]
- 60.Zipf WB, O’Dorisio TM, Cataland S, Dixon K. Pancreatic polypeptide responses to protein meal challenges in obese but otherwise normal children and obese children with Prader-Willi syndrome. Journal of Clinical Endocrinology and Metabolism. 1983;57(5):1074–1080. doi: 10.1210/jcem-57-5-1074. [DOI] [PubMed] [Google Scholar]
- 61.Ueno N, Inui A, Iwamoto M, et al. Decreased food intake and body weight in pancreatic polypeptide-overexpressing mice. Gastroenterology. 1999;117(6):1427–1432. doi: 10.1016/s0016-5085(99)70293-3. [DOI] [PubMed] [Google Scholar]
- 62.Batterham RL, Le Roux CW, Cohen MA, et al. Pancreatic polypeptide reduces appetite and food intake in humans. Journal of Clinical Endocrinology and Metabolism. 2003;88(8):3989–3992. doi: 10.1210/jc.2003-030630. [DOI] [PubMed] [Google Scholar]
- 63.Berntson GG, Zipf WB, O’Dorisio TM, Hoffman JA, Chance RE. Pancreatic polypeptide infusions reduce food intake in Prader-Willi syndrome. Peptides. 1993;14(3):497–503. doi: 10.1016/0196-9781(93)90138-7. [DOI] [PubMed] [Google Scholar]
- 64.Holst JJ. On the physiology of GIP and GLP-1. Hormone and Metabolic Research. 2004;36(11-12):747–754. doi: 10.1055/s-2004-826158. [DOI] [PubMed] [Google Scholar]
- 65.Tang-Christensen M, Vrang N, Larsen PJ. Glucagon-like peptide containing pathways in the regulation of feeding behaviour. International Journal of Obesity. 2001;25(supplement 5):S42–S47. doi: 10.1038/sj.ijo.0801912. [DOI] [PubMed] [Google Scholar]
- 66.Tucker JD, Dhanvantari S, Brubaker PL. Proglucagon processing in islet and intestinal cell lines. Regulatory Peptides. 1996;62(1):29–35. doi: 10.1016/0167-0115(95)00167-0. [DOI] [PubMed] [Google Scholar]
- 67.Deacon CF. Circulation and degradation of GIP and GLP-1. Hormone and Metabolic Research. 2004;36(11-12):761–765. doi: 10.1055/s-2004-826160. [DOI] [PubMed] [Google Scholar]
- 68.Mentlein R, Gallwitz B, Schmidt WE. Dipeptidyl-peptidase IV hydrolyses gastric inhibitory polypeptide, glucagon-like peptide-1(7-36)amide, peptide histidine methionine and is responsible for their degradation in human serum. European Journal of Biochemistry. 1993;214(3):829–835. doi: 10.1111/j.1432-1033.1993.tb17986.x. [DOI] [PubMed] [Google Scholar]
- 69.Vilsbøll T, Agersø H, Krarup T, Holst JJ. Similar elimination rates of glucagon-like peptide-1 in obese type 2 diabetic patients and healthy subjects. Journal of Clinical Endocrinology and Metabolism. 2003;88(1):220–224. doi: 10.1210/jc.2002-021053. [DOI] [PubMed] [Google Scholar]
- 70.Orskov C, Rabenhoj L, Wettergren A, Kofod H, Holst JJ. Tissue and plasma concentrations of amidated and glycine-extended glucagon-like peptide I in humans. Diabetes. 1994;43(4):535–539. doi: 10.2337/diab.43.4.535. [DOI] [PubMed] [Google Scholar]
- 71.Holst JJ. The physiology of glucagon-like peptide 1. Physiological Reviews. 2007;87(4):1409–1439. doi: 10.1152/physrev.00034.2006. [DOI] [PubMed] [Google Scholar]
- 72.Yamato E, Ikegami H, Takekawa K, et al. Tissue-specific and glucose-dependent expression of receptor genes for glucagon and glucagon-like peptide-1 (GLP-1) Hormone and Metabolic Research. 1997;29(2):56–59. doi: 10.1055/s-2007-978985. [DOI] [PubMed] [Google Scholar]
- 73.Vahl TP, Drazen DL, Seeley RJ, D’Alessio DA, Woods SC. Meal-anticipatory glucagon-like peptide-1 secretion in rats. Endocrinology. 2010;151(2):569–575. doi: 10.1210/en.2009-1002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Cummings DE, Overduin J. Gastrointestinal regulation of food intake. Journal of Clinical Investigation. 2007;117(1):13–23. doi: 10.1172/JCI30227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Verdich C, Flint A, Gutzwiller JP, et al. A meta-analysis of the effect of glucagon-like peptide-1 (7-36) amide on Ad Libitum energy intake in humans. Journal of Clinical Endocrinology and Metabolism. 2001;86(9):4382–4389. doi: 10.1210/jcem.86.9.7877. [DOI] [PubMed] [Google Scholar]
- 76.Zander M, Madsbad S, Madsen JL, Holst JJ. Effect of 6-week course of glucagon-like peptide 1 on glycaemic control, insulin sensitivity, and β-cell function in type 2 diabetes: a parallel-group study. Lancet. 2002;359(9309):824–830. doi: 10.1016/S0140-6736(02)07952-7. [DOI] [PubMed] [Google Scholar]
- 77.Eng J, Kleinman WA, Singh L, Singh G, Raufman JP. Isolation and characterization of exendin-4, an exendin-3 analogue, from Heloderma suspectum venom. Further evidence for an exendin receptor on dispersed acini from guinea pig pancreas. Journal of Biological Chemistry. 1992;267(11):7402–7405. [PubMed] [Google Scholar]
- 78.DeFronzo RA, Ratner RE, Han J, Kim DD, Fineman MS, Baron AD. Effects of exenatide (exendin-4) on glycemic control and weight over 30 weeks in metformin-treated patients with type 2. Diabetes Care. 2005;28(5):1092–1100. doi: 10.2337/diacare.28.5.1092. [DOI] [PubMed] [Google Scholar]
- 79.Drucker DJ, Buse JB, Taylor K, et al. Exenatide once weekly versus twice daily for the treatment of type 2 diabetes: a randomised, open-label, non-inferiority study. The Lancet. 2008;372(9645):1240–1250. doi: 10.1016/S0140-6736(08)61206-4. [DOI] [PubMed] [Google Scholar]
- 80.Buse JB, Rosenstock J, Sesti G, et al. Liraglutide once a day versus exenatide twice a day for type 2 diabetes: a 26-week randomised, parallel-group, multinational, open-label trial (LEAD-6) The Lancet. 2009;374(9683):39–47. doi: 10.1016/S0140-6736(09)60659-0. [DOI] [PubMed] [Google Scholar]
- 81.Egan JM, Bulotta A, Hui H, Perfetti R. GLP-1 receptor agonists are growth and differentiation factors for pancreatic islet beta cells. Diabetes/Metabolism Research and Reviews. 2003;19(2):115–123. doi: 10.1002/dmrr.357. [DOI] [PubMed] [Google Scholar]
- 82.Harkavyi A, Whitton PS. Glucagon-like peptide 1 receptor stimulation as a means of neuroprotection. British Journal of Pharmacology. 2010;159(3):495–501. doi: 10.1111/j.1476-5381.2009.00486.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Harkavyi A, Abuirmeileh A, Lever R, Kingsbury AE, Biggs CS, Whitton PS. Glucagon-like peptide 1 receptor stimulation reverses key deficits in distinct rodent models of Parkinson’s disease. Journal of Neuroinflammation. 2008;5, article no. 19 doi: 10.1186/1742-2094-5-19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Kim S, Moon M, Park S. Exendin-4 protects dopaminergic neurons by inhibition of microglial activation and matrix metalloproteinase-3 expression in an animal model of Parkinson’s disease. Journal of Endocrinology. 2009;202(3):431–439. doi: 10.1677/JOE-09-0132. [DOI] [PubMed] [Google Scholar]
- 85.Li Y, Perry T, Kindy MS, et al. GLP-1 receptor stimulation preserves primary cortical and dopaminergic neurons in cellular and rodent models of stroke and Parkinsonism. Proceedings of the National Academy of Sciences of the United States of America. 2009;106(4):1285–1290. doi: 10.1073/pnas.0806720106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Perry T, Lahiri DK, Sambamurti K, et al. Glucagon-like peptide-1 decreases endogenous amyloid-β peptide (Aβ) levels and protects hippocampal neurons from death induced by Aβ and iron. Journal of Neuroscience Research. 2003;72(5):603–612. doi: 10.1002/jnr.10611. [DOI] [PubMed] [Google Scholar]
- 87.Perry TA, Greig NH. A new Alzheimer’s disease interventive strategy: GLP-1. Current Drug Targets. 2004;5(6):565–571. doi: 10.2174/1389450043345245. [DOI] [PubMed] [Google Scholar]
- 88.Perry T, Holloway HW, Weerasuriya A, et al. Evidence of GLP-1-mediated neuroprotection in an animal model of pyridoxine-induced peripheral sensory neuropathy. Experimental Neurology. 2007;203(2):293–301. doi: 10.1016/j.expneurol.2006.09.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Drucker DJ. Glucagon-like peptide 2. Trends in Endocrinology and Metabolism. 1999;10(4):153–156. doi: 10.1016/s1043-2760(98)00136-2. [DOI] [PubMed] [Google Scholar]
- 90.Schmidt PT, Näslund E, Grybäck P, et al. Peripheral administration of GLP-2 to humans has no effect on gastric emptying or satiety. Regulatory Peptides. 2003;116(1–3):21–25. doi: 10.1016/s0167-0115(03)00175-7. [DOI] [PubMed] [Google Scholar]
- 91.Drucker DJ, Ehrlich P, Asa SL, Brubaker PL. Induction of intestinal epithelial proliferation by glucagon-like peptide 2. Proceedings of the National Academy of Sciences of the United States of America. 1996;93(15):7911–7916. doi: 10.1073/pnas.93.15.7911. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Wallis K, Walters JRF, Forbes A. Review article: glucagon-like peptide 2—current applications and future directions. Alimentary Pharmacology and Therapeutics. 2007;25(4):365–372. doi: 10.1111/j.1365-2036.2006.03193.x. [DOI] [PubMed] [Google Scholar]
- 93.Buchman AL, Katz S, Fang JC, Bernstein CN, Abou-Assi SG. Teduglutide, a novel mucosally active analog of glucagon-like peptide-2 (GLP-2) for the treatment of moderate to severe Crohn’s disease. Inflammatory Bowel Diseases. 2010;16(6):962–973. doi: 10.1002/ibd.21117. [DOI] [PubMed] [Google Scholar]
- 94.Nagell CF, Wettergren A, Pedersen JF, Mortensen D, Holst JJ. Glucagon-like peptide-2 inhibits antral emptying in man, but is not as potent as glucagon-like peptide-1. Scandinavian Journal of Gastroenterology. 2004;39(4):353–358. doi: 10.1080/00365520410004424. [DOI] [PubMed] [Google Scholar]
- 95.Ghatei MA, Uttenthal LO, Christofides ND. Molecular forms of human enteroglucagon in tissue and plasma: plasma responses to nutrient stimuli in health and in disorders of the upper gastrointestinal tract. Journal of Clinical Endocrinology and Metabolism. 1983;57(3):488–495. doi: 10.1210/jcem-57-3-488. [DOI] [PubMed] [Google Scholar]
- 96.Cohen MA, Ellis SM, Le Roux CW, et al. Oxyntomodulin suppresses appetite and reduces food intake in humans. Journal of Clinical Endocrinology and Metabolism. 2003;88(10):4696–4701. doi: 10.1210/jc.2003-030421. [DOI] [PubMed] [Google Scholar]
- 97.Dakin CL, Gunn I, Small CJ, et al. Oxyntomodulin inhibits food intake in the rat. Endocrinology. 2001;142(10):4244–4250. doi: 10.1210/endo.142.10.8430. [DOI] [PubMed] [Google Scholar]
- 98.Wynne K, Park AJ, Small CJ, et al. Oxyntomodulin increases energy expenditure in addition to decreasing energy intake in overweight and obese humans: a randomised controlled trial. International Journal of Obesity. 2006;30(12):1729–1736. doi: 10.1038/sj.ijo.0803344. [DOI] [PubMed] [Google Scholar]
- 99.Dakin CL, Small CJ, Batterham RL, et al. Peripheral oxyntomodulin reduces food intake and body weight gain in rats. Endocrinology. 2004;145(6):2687–2695. doi: 10.1210/en.2003-1338. [DOI] [PubMed] [Google Scholar]
- 100.Baggio LL, Huang Q, Brown TJ, Drucker DJ. Oxyntomodulin and glucagon-like peptide-1 differentially regulate murine food intake and energy expenditure. Gastroenterology. 2004;127(2):546–558. doi: 10.1053/j.gastro.2004.04.063. [DOI] [PubMed] [Google Scholar]
- 101.Sowden GL, Drucker DJ, Weinshenker D, Swoap SJ. Oxyntomodulin increases intrinsic heart rate in mice independent of the glucagon-like peptide-1 receptor. American Journal of Physiology. 2007;292(2):R962–R970. doi: 10.1152/ajpregu.00405.2006. [DOI] [PubMed] [Google Scholar]
- 102.Ban K, Kim KH, Cho CK, et al. Glucagon-Like Peptide (GLP)-1(9-36)amide-mediated cytoprotection is blocked by exendin(9-39) yet does not require the known GLP-1 receptor. Endocrinology. 2010;151(4):1520–1531. doi: 10.1210/en.2009-1197. [DOI] [PubMed] [Google Scholar]
- 103.Druce MR, Minnion JS, Field BCT, et al. Investigation of structure-activity relationships of oxyntomodulin (Oxm) using oxm analogs. Endocrinology. 2009;150(4):1712–1721. doi: 10.1210/en.2008-0828. [DOI] [PubMed] [Google Scholar]
- 104.Kojima M, Hosoda H, Date Y, Nakazato M, Matsuo H, Kangawa K. Ghrelin is a growth-hormone-releasing acylated peptide from stomach. Nature. 1999;402(6762):656–660. doi: 10.1038/45230. [DOI] [PubMed] [Google Scholar]
- 105.Cummings DE, Purnell JQ, Frayo RS, Schmidova K, Wisse BE, Weigle DS. A preprandial rise in plasma ghrelin levels suggests a role in meal initiation in humans. Diabetes. 2001;50(8):1714–1719. doi: 10.2337/diabetes.50.8.1714. [DOI] [PubMed] [Google Scholar]
- 106.Nakazato M, Murakami N, Date Y, et al. A role for ghrelin in the central regulation of feeding. Nature. 2001;409(6817):194–198. doi: 10.1038/35051587. [DOI] [PubMed] [Google Scholar]
- 107.Tschop M, Smiley DL, Heiman ML. Ghrelin induces adiposity in rodents. Nature. 2000;407(6806):908–913. doi: 10.1038/35038090. [DOI] [PubMed] [Google Scholar]
- 108.Otto B, Cuntz U, Fruehauf E, et al. Weight gain decreases elevated plasma ghrelin concentrations of patients with anorexia nervosa. European Journal of Endocrinology. 2001;145(5):669–673. [PubMed] [Google Scholar]
- 109.Le Roux CW, Patterson M, Vincent RP, Hunt C, Ghatei MA, Bloom SR. Postprandial plasma ghrelin is suppressed proportional to meal calorie content in normal-weight but not obese subjects. Journal of Clinical Endocrinology and Metabolism. 2005;90(2):1068–1071. doi: 10.1210/jc.2004-1216. [DOI] [PubMed] [Google Scholar]
- 110.Nagaya N, Uematsu M, Kojima M, et al. Elevated circulating level of ghrelin in cachexia associated with chronic heart failure: relationships between ghrelin and anabolic/catabolic factors. Circulation. 2001;104(17):2034–2038. doi: 10.1161/hc4201.097836. [DOI] [PubMed] [Google Scholar]
- 111.Cummings DE, Clement K, Purnell JQ, et al. Elevated plasma ghrelin levels in Prader-Willi syndrome. Nature Medicine. 2002;8(7):643–644. doi: 10.1038/nm0702-643. [DOI] [PubMed] [Google Scholar]
- 112.Taheri S, Lin L, Austin D, Young T, Mignot E. Short sleep duration is associated with reduced leptin, elevated ghrelin, and increased body mass index. PLoS Medicine. 2004;1, article e62 doi: 10.1371/journal.pmed.0010062. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Wang L, Saint-Pierre DH, Taché Y. Peripheral ghrelin selectively increases Fos expression in neuropeptide Y—synthesizing neurons in mouse hypothalamic arcuate nucleus. Neuroscience Letters. 2002;325(1):47–51. doi: 10.1016/s0304-3940(02)00241-0. [DOI] [PubMed] [Google Scholar]
- 114.Chen HY, Trumbauer ME, Chen AS, et al. Orexigenic action of peripheral ghrelin is mediated by neuropeptide Y and agouti-related protein. Endocrinology. 2004;145(6):2607–2612. doi: 10.1210/en.2003-1596. [DOI] [PubMed] [Google Scholar]
- 115.Lawrence CB, Snape AC, Baudoin FMH, Luckman SM. Acute central ghrelin and GH secretagogues induce feeding and activate brain appetite centers. Endocrinology. 2002;143(1):155–162. doi: 10.1210/endo.143.1.8561. [DOI] [PubMed] [Google Scholar]
- 116.Date Y, Murakami N, Toshinai K, et al. The role of the gastric afferent vagal nerve in Ghrelin-induced feeding and growth hormone secretion in rats. Gastroenterology. 2002;123(4):1120–1128. doi: 10.1053/gast.2002.35954. [DOI] [PubMed] [Google Scholar]
- 117.Malik S, McGlone F, Bedrossian D, Dagher A. Ghrelin modulates brain activity in areas that control appetitive behavior. Cell Metabolism. 2008;7(5):400–409. doi: 10.1016/j.cmet.2008.03.007. [DOI] [PubMed] [Google Scholar]
- 118.Gibbs J, Young RC, Smith GP. Cholecystokinin decreases food intake in rats. Journal of Comparative and Physiological Psychology. 1973;84(3):488–495. doi: 10.1037/h0034870. [DOI] [PubMed] [Google Scholar]
- 119.Liddle RA, Goldfine ID, Rosen MS. Cholecystokinin bioactivity in human plasma. Molecular forms, responses to feeding, and relationship to gallbladder contraction. Journal of Clinical Investigation. 1985;75(4):1144–1152. doi: 10.1172/JCI111809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Kissileff HR, Pi-Sunyer FX, Thornton J, Smith GP. C-terminal octapeptide of cholecystokinin decreases food intake in man. American Journal of Clinical Nutrition. 1981;34(2):154–160. doi: 10.1093/ajcn/34.2.154. [DOI] [PubMed] [Google Scholar]
- 121.Moran TH, Baldessarini AR, Salorio CF, Lowery T, Schwartz GJ. Vagal afferent and efferent contributions to the inhibition of food intake by cholecystokinin. American Journal of Physiology. 1997;272(4):R1245–R1251. doi: 10.1152/ajpregu.1997.272.4.R1245. [DOI] [PubMed] [Google Scholar]
- 122.Moran TH, Katz LF, Plata-Salaman CR, Schwartz GJ. Disordered food intake and obesity in rats lacking cholecystokinin A receptors. American Journal of Physiology. 1998;274(3):R618–R625. doi: 10.1152/ajpregu.1998.274.3.R618. [DOI] [PubMed] [Google Scholar]
- 123.Wank SA. Cholecystokinin receptors. American Journal of Physiology. 1995;269(5):G628–G646. doi: 10.1152/ajpgi.1995.269.5.G628. [DOI] [PubMed] [Google Scholar]
- 124.West DB, Fey D, Woods SC. Cholecystokinin persistently suppresses meal size but not food intake in free-feeding rats. The American journal of physiology. 1984;246(5):R776–R787. doi: 10.1152/ajpregu.1984.246.5.R776. [DOI] [PubMed] [Google Scholar]
- 125.Crawley JN, Beinfeld MC. Rapid development of tolerance to the behavioural actions of cholecystokinin. Nature. 1983;302(5910):703–706. doi: 10.1038/302703a0. [DOI] [PubMed] [Google Scholar]
- 126.Polonsky KS, Given BD, Van Cauter E. Twenty-four-hour profiles and pulsatile patterns of insulin secretion in normal and obese subjects. Journal of Clinical Investigation. 1988;81(2):442–448. doi: 10.1172/JCI113339. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Air EL, Benoit SC, Blake Smith KA, Clegg DJ, Woods SC. Acute third ventricular administration of insulin decreases food intake in two paradigms. Pharmacology Biochemistry and Behavior. 2002;72(1-2):423–429. doi: 10.1016/s0091-3057(01)00780-8. [DOI] [PubMed] [Google Scholar]
- 128.Porte D, Jr., Woods SC. Regulation of food intake and body weight by insulin. Diabetologia. 1981;20:274–280. doi: 10.1007/BF00254493. [DOI] [PubMed] [Google Scholar]
- 129.Considine RV, Sinha MK, Heiman ML, et al. Serum immunoreactive-leptin concentrations in normal-weight and obese humans. New England Journal of Medicine. 1996;334(5):292–295. doi: 10.1056/NEJM199602013340503. [DOI] [PubMed] [Google Scholar]
- 130.Saad MF, Riad-Gabriel MG, Khan A, et al. Diurnal and ultradian rhythmicity of plasma leptin: effects of gender and adiposity. Journal of Clinical Endocrinology and Metabolism. 1998;83(2):453–459. doi: 10.1210/jcem.83.2.4532. [DOI] [PubMed] [Google Scholar]
- 131.Farooqi IS, Jebb SA, Langmack G, et al. Effects of recombinant leptin therapy in a child with congenital leptin deficiency. New England Journal of Medicine. 1999;341(12):879–884. doi: 10.1056/NEJM199909163411204. [DOI] [PubMed] [Google Scholar]
- 132.Zelissen PMJ, Stenlof K, Lean MEJ, et al. Effect of three treatment schedules of recombinant methionyl human leptin on body weight in obese adults: a randomized, placebo-controlled trial. Diabetes, Obesity and Metabolism. 2005;7(6):755–761. doi: 10.1111/j.1463-1326.2005.00468.x. [DOI] [PubMed] [Google Scholar]