

Abstract
During reprogramming of porcine mesenchymal cells with a four-factor (POU5F1/SOX2/KLF4/MYC) mixture of vectors, a fraction of the colonies had an atypical phenotype and arose earlier than the recognizable porcine induced pluripotent stem (iPS) cell colonies. Within days after each passage, patches of cells with an epithelial phenotype formed raised domes, particularly under 20% O2 conditions. Relative to gene expression of the iPS cells, there was up-regulation of genes for transcription factors associated with trophoblast (TR) lineage emergence, e.g., GATA2, PPARG, MSX2, DLX3, HAND1, GCM1, CDX2, ID2, ELF5, TCFAP2C, and TEAD4 and for genes required for synthesis of products more typical of differentiated TR, such as steroids (HSD17B1, CYP11A1, and STAR), pregnancy-associated glycoproteins (PAG6), and select cytokines (IFND, IFNG, and IL1B). Although POU5F1 was down-regulated relative to that in iPS cells, it was not silenced in the induced TR (iTR) cells over continued passage. Like iPS cells, iTR cells did not senesce on extended passage and displayed high telomerase activity. Upon xenografting into immunodeficient mice, iTR cells formed nonhemorrhagic teratomas composed largely of layers of epithelium expressing TR markers. When cultured under conditions that promoted embryoid body formation, iTR cells formed floating spheres consisting of a single epithelial sheet whose cells were tethered laterally by desmosome-like structures. In conclusion, reprogramming of porcine fibroblasts to iPS cells generates, as a by-product, colonies composed of self-renewing populations of TR cells, possibly containing TR stem cells.
Keywords: cell culture, differentiation, porcine/pig, reprogramming, trophoblast
Reprogramming of porcine fibroblasts to iPS cells generates, as a by-product, colonies that are composed of self-renewing populations of trophoblast cells.
INTRODUCTION
Trophoblast (TR) stem cells (TSC) have been described in rodents [1, 2] and possibly in monkeys [3]. They have been best characterized in the mouse, where they can be derived as outgrowths of polar trophectoderm of blastocysts and microdissected explants of extra-embryonic ectoderm [4]. Both sources of cells are dependent upon the addition of FGF4, a supporting layer of irradiated mouse embryonic fibroblasts, and supplementary fetal bovine serum [5]. Authenticated TSC have not been generated from ungulate species, although primary TR cell lines have been established from sheep and goat [6], pig [7–10], and cattle conceptuses [11–13]. Many of these cell lines grow continuously in culture without apparent senescence and display genes characteristic of TR, but they likely represent a stage of differentiation beyond the TSC.
Not only has the derivation of TSC from ungulate embryos proved to be problematic, there have also been difficulties in generating embryonic stem cells (ESC) from such species. As an alternative, several teams of investigators have created induced pluripotent stem (iPS) cells from pigs based on the method first described by Takahashi and Yamanaka [14], by reprogramming somatic cells with “stemness-promoting” genes incorporated into integrating lentiviral vectors [15–18]. In the course of our experiments [15], we noticed that a significant fraction of the lines that we generated displayed unusual colony morphology. Here we describe the phenotype of such cells and show that reprogramming generated not only iPS cells but cells with the features of TR as well.
MATERIALS AND METHODS
Cell Culture and Lentiviral Transduction
Cell culture, virus production, and the reprogramming protocols were carried out as described elsewhere [15]. Progenitor cells were porcine fetal fibroblasts derived from whole conceptuses at day 34 of pregnancy, which were thawed from frozen stocks [19, 20]. All colonies generated, whether authentic iPS cells or cells that manifested properties of TR (iTR) cell lines, were cultured initially under standard conditions for maintaining human ESC. This medium (referred to here as F2 medium) contains 80% Dulbecco modified Eagle medium-F12 (Thermo HyClone, IL) supplemented with 20% Knockout serum replacement (KOSR) (Invitrogen, CA), 1 mM l-glutamine, 0.1 mM 2-mercaptoethanol, 1% nonessential amino acids (Sigma-Aldrich, MO), and 4 ng/ml recombinant human FGF2 (prepared in house) and was changed daily. All cultures were maintained in six-well tissue culture plates (Nunc, Sigma-Aldrich) containing a monolayer of γ-irradiated (∼7000 cGy) mouse embryonic fibroblast (MEF) feeder cells. Cell passage was carried out every 4–6 days by splitting physically dissociated colonies approximately 1:10. However, dissociation with Accutase (Invitrogen) can also be used to disperse iTR cell colonies.
Alkaline Phosphatase Staining and Immunocytochemistry
Alkaline phosphatase (AP) staining of whole colonies was performed with an AP detection kit (Chemicon/Millipore, MA). For immunocytochemical localization, cells were grown on MEF feeder cells over coverslips coated with poly-d-lysine and placed in six-well tissue culture plates. Coverslips were fixed in 4% paraformaldehyde in PBS for 10 min at room temperature, washed, and exposed to a solution of 5% goat serum (Sigma), 5% bovine serum albumin (Jackson-Immunoresearch, PA), and 0.1% Triton X-100 in PBS for 30 min at room temperature. They were then incubated with primary rabbit polyclonal antibody directed against SOX2 (1:600 dilution; product AB5603; Millipore-Chemicon) and mouse monoclonal KRT7 antibody (1:100 dilution; clone OV-TL; DAKO, CA) overnight at 4°C. After cells were washed, they were incubated with secondary antibodies Alexa546-conjugated goat anti-rabbit immunoglobulin G (IgG), and Alexa488-conjugated goat anti-mouse IgG (both 1:500 dilution; both from Invitrogen) in the dark. A secondary antibody-only staining of the coverslips was used as a control in these experiments. Nuclei were stained with 6 ng/ml 4′,6-diamidino-2-phenylindole (DAPI; Invitrogen). Epifluorescent images were obtained with an Olympus Provis AX-70 model inverted microscope with a U-MBC multicontrol box (Melville, NY) equipped with a cooled, charge-coupled device camera (Photometrics CoolSnap-ES, Tucson, TX). Controls were performed with the omission of primary antibodies for all experiments.
Reverse Transcription-PCR Analysis
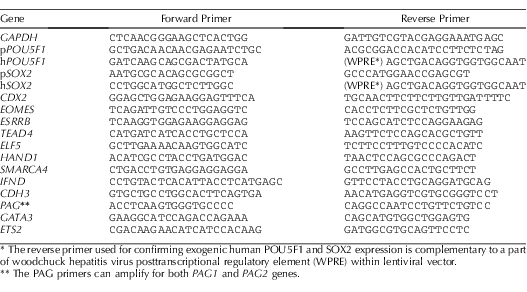
RNA from the initiating fibroblast lines and iPS and iTR cells was isolated with STAT-60 reagent (Tel-Test, TX), treated with DNaseI (Ambion, TX), and reverse transcribed by using oligo(dT) and Superscript III reverse transcriptase (RT; Invitrogen). Amplification was carried out with PCR Master Mix (Fermentas, MD). The primer sets used are listed in Table 1. All PCR reactions were initiated at 94°C for 3 min, followed by 40 cycles of 95°C for 15 sec, 60°C for 15 sec for the porcine POU5F1 (pPOU5F1) and human POU5F1 ortholog (hPOU5F1) and the GAPDH, PAG, pSOX2, SMARCA4, and IFND genes. The temperature was 58°C for amplifying the TEAD4, HAND1, ESRRB, ETS2, GATA3, ELF5, and CDH3 transcripts, 57°C for the CDX2 transcripts, 56°C for the EOMES transcripts, and 62°C for the hSOX2 transcripts for 15 sec and 72°C for 30 sec. Reactions were terminated by 1 min at 72°C.
TABLE 1.
Primer sets for PCR.
Transcriptional Profiling by Microarray
Cells were grown in 6-well plates, lysed with 1 ml of STAT-60, and kept at −80°C. RNA from one iTR, three iPS cell, and two porcine fetal fibroblasts (PFF) lines were hybridized to Affymetrix GeneChip porcine genome arrays, as described in detail elsewhere [15].
Telomerase Activity Assay
Telomerase activity in 0.2 μg of cell protein from representative cell lines was determined with Trapeze RT telomerase detection kit (Chemicon) by using Platinum Taq DNA polymerase (Invitrogen) according to the manufacturer's instructions.
Estradiol Immunoassays
Media samples collected from iTR and iPS cells were stored at −20°C until assayed. Estradiol was measured using an estradiol EIA kit (product no. 582251; Cayman Chemical, Ann Arbor, MI). Standards were supplied by the manufacturer. For all assays, samples fell within the recommended ranges of the standard curves and were assayed in duplicate. Data are presented as means ± SEM in pg/ml.
Teratoma Formation
iTR cells (106) from a 35-mm culture dish were injected subcutaneously into the dorsal flank of a CD1 nude mouse (Charles River, MA). Eleven to 13 weeks after injection, tumors were cut into pieces. Some pieces were fixed in 4% paraformaldehyde, and sections (5 μm) were cut from paraffin-embedded tissue and subsequently stained with hematoxylin and eosin.
Cell Sphere Formation
Colonies were detached from the MEF feeder layer by manual dispersion and transferred into medium without FGF2 (differentiation medium) in low-attachment plates (Corning, NY). After 2 days of culture, cell spheres were either photographed with an Olympus CKX41 model inverted microscope or fixed for electron microscopy.
Population Doubling Time
iTR1 and iTR3 cells were seeded onto 12-well plates (∼5 × 104 cells/well and ∼5 × 103 cells/well, respectively) precoated with Matrigel after dispersal with trypsin under standard culture conditions. Numbers of cells in each well (triplicates for each time point) were counted at 24, 48, 72, and 96 h (iTR1) or at 48, 96, 144, and 192 h (iTR3), with a daily exchange of fresh medium [21, 22].
Karyotyping
Standard G-banding chromosome analysis was carried out at Cell Line Genetics (Madison, WI). Cytogenetic analysis was performed with 20 G-banded metaphase spreads for each cell line.
Electron Microscopy
Cells were cultured on Matrigel-coated Thermanox plastic coverslips (Nunc, Thermo Scientific) and fixed in a solution of 2% glutaraldehyde and 2% paraformaldehyde in 0.1 M cacodylate buffer at room temperature and stained with 1% osmium tetroxide. Samples were dehydrated through a graded ethanol series and placed in acetone prior to infiltration with Epon-Spurr resin. A second fixation, dehydration, and infiltration process was performed at Pelco Microwave Sample Processing Systems (Ted Pella, CA). Thin sections (85 nm) were collected using an Ultracut UCT microtome (Leica, IL) and a 45° Diatome diamond knife and stained with 5% uranyl acetate and Sato triple lead stain. Sections were examined with a JEOL 1400-EX model transmission microscope.
Animal Care and Use
All experiments with mice (generation of embryonic fibroblasts and teratoma formation) were conducted in accordance with the National Research Council publication Guide for Care and Use of Laboratory Research Involving Animals under protocols (4125 and 4467, respectively), approved by the University of Missouri's Animal Care and Use Committee.
RESULTS
Generation of iTR Cells During Reprogramming of Fibroblasts
During the conversion of PFF to iPS cells with the standard four-factor (POU5F1/SOX2/KLF4/MYC, abbreviated OSKM) mixture of vectors, approximately 17% of the colonies picked possessed an abnormal phenotype that was distinct from that of the typical porcine iPS cells generated in the same culture plates (Fig. 1A). In a later experiment in which we reprogrammed porcine umbilical cord mesenchyme cells with a similar OSKM combination of transgenes, an even larger proportion of such colonies appeared (79%), outnumbering the more typical iPS colonies from the same culture dish. However, we have only made a thorough characterization of lines derived from PFF, and it is a description of these cells that follows.
FIG. 1.

Features of iTR cell colonies are shown. A) Emerging colony on original reprogramming plate at Day 22 after viral transduction is shown. The iTR colony, which provided the iTR1 line, was broken up physically and transferred fresh MEF feeder in stem cell medium containing KOSR and FGF2 (F2 medium), i.e., regular stem cell medium. B) During culture after the second passage (p2), we noted that the colonies contained a mixture of cell types, including areas that lifted from the substratum and formed domes. Areas forming domes were roughly ovoid and consisted exclusively of epithelial sheets. C) Small iTR1 colonies contain a heterogeneous mixture of morphologically distinct cell types, including ones with an epithelial appearance. D) A larger iTR1 cell colony stained for alkaline phosphatase (AP). Although epithelial cells with larger cytoplasm were negative for AP, smaller cells are weakly positive. The colony shown had possessed a centrally placed ovoid dome that shrank and collapsed during fixation. E) If colonies are cultured beyond 10 days in F2 medium under 20% O2, iTR colonies became largely epithelial, with most of the surface covered with well-developed “dome-like” structures that give rise to floating spheres. F) In parallel experiments in which iTR cells had been generated under 4% O2 and then maintained under these conditions, iTR colonies showed a more modest development of domes (images captured at Day 14 of culture at p1). Bars = 500 μm in A–F. Immunostaining of regions of an iTR1 colony (G, H) and iPS cell colonies (line ID6 [I]) for KRT7 (green [G, H, I]) and SOX2 (red [G and I]). Nuclei have been counterstained with DAPI (blue [G, H, I]). KRT7 staining was evident in the epithelioid cells and was prominent in the outer regions of the colony (G). The domes also stained intensely for KRT7 and frequently rounded off to form floating vesicles (H). H) Focus was adjusted to the protruding tip of a dome so that the cells attached to the coverslip were out of focus. All cells in iTR and iPS colonies stained positively for SOX2, although immunostaining could not distinguish between the endogenous gene product and SOX2 expressed from the reprogramming gene (G, I). KRT7 expression was observed only in iTR cells. Bars = 200 μm in G–I.
The abnormal colonies first appeared approximately 3 weeks after exposure to the transducing viruses and approximately 1 week earlier than the colonies that displayed the typical properties of iPS cells (Fig. 1A). They were heterogeneous in terms of component cell morphologies but were always characterized by the presence of areas of relatively flattened, epithelium-like cells, as well as regions of smaller cells (Fig. 1B). Henceforth, these atypical colonies are called induced TR (iTR) cells, because, as shown below, they expressed the signatures of porcine TR. The initiating cultures also contained extensive numbers of small, round “granular” cells that tended to attach loosely to both types of colonies, as well as to the MEF feeders. Washing the plates with 1× PBS and carefully selecting colonies for propagation reduced contamination by these granular cells.
iTR colonies were picked individually from the culture plates in which they had been generated and cultured on standard ESC medium containing KOSR and FGF2 (F2 medium) above feeder layers of MEF under a 20% O2 and 5% CO2 gas atmosphere. After passage, the colonies continued to show heterogeneity with a mixture of cell types, including cells that had an epithelioid appearance (Fig. 1, B and C). Another difference that distinguished iTR colonies from iPS colonies was that the former demonstrated only “patchy” and relatively weak staining for the pluripotent marker, AP (Fig. 1D). The epithelial-like cells, in particular, appeared not to be positive for AP. If the colonies were not subcultured frequently, they could reach several millimeters in diameter. They also began to form domes from which vesicles formed and detached into the medium (Fig. 1, B and E). Such domes frequently occupied the central areas of the colonies (Fig. 1E). If the colonies were generated and then maintained under a gas atmosphere containing lower concentrations of O2 (4%), rather than 20% O2, the dome-like structures and associated vesicles formed more slowly over time (compare Fig. 1E with F). Dome formation was not observed with iPS cell colonies generated under identical reprogramming conditions (not shown). Like the iPS cells, iTR cultures showed no tendency to senesce over time in culture. One line (iTR1) had a cell doubling time of ∼21 h, slightly longer than that of iPS cells (17 h) [15] and survived routine passaging every 5–6 days at a dilution of ∼1:10 for 57 passages (>150 doublings), but with increasing passage, colonies became more uniform in appearance, with the epithelium-like cell type predominating early after passage and dome formation that began within 3 days. A second line, iTR2, had a growth speed similar to that of iTR1 and survived for 40 passages. The third line, iTR3, had a longer cell doubling time of 40 h and survived routine passaging every 10–11 days at a dilution of ∼1:10 for 15 passages.
Expression of TR Markers in iTR Cells
Colonies of iTR expressed the TR marker KRT7. Staining was particularly intense in the cells near the edges of the colonies (Fig. 1G) and in the domes and vesicles that formed when culture was prolonged (Fig. 1H), suggesting that the colonies were composed largely, if not entirely, of cells of the TR lineage. In contrast, porcine iPS cells failed to stain for KRT7 (Fig. 1I).
Microarray analysis of RNA extracted from the TR cells was initially performed with cell line iTR1 at passage 2. Figure 2, A and B, compares the global gene expression profiles of line iTR1 with two PFF lines, including the one from which it was derived, and three stable iPS cell lines obtained in the same reprogramming experiment (Gene Expression Omnibus accession no. GSE15472). The iTR clustered separately from both the iPS cells and the originating PFF. Moreover, the spread of normalized gene expression was much broader in the TR than in the iPS cell line (Fig. 2B), and many of the highly expressed genes of the iPS cell lines were down-regulated, while weakly expressed genes were up-regulated in the iTR cells. Specifically, there was up-regulation of genes for transcription factors associated with TR lineage emergence and differentiation, e.g., the GATA2, PPARG, MSX2, DLX3, HAND1, GCM1, CDX2, ID2, ELF5, TCFAP2C, TEAD4 genes, and genes required for steroid synthesis (HSD17B1, CYP11A1, STAR) and some unique porcine TR markers, e.g., the PAG6 and PAG10 genes (Fig. 2C). Inflammatory cytokine IFNG and IL1B transcripts, expressed during the early stages of porcine TR emergence [23, 24], and the imprinted H19 gene were greatly up-regulated compared to those in both PFF and iPS cells. Of the cytokeratins, the KRT7 gene, which is generally regarded as a diagnostic TR marker, was up-regulated in the TR colonies compared to those in both the PFF and iPS cells, while the KRT19, KRT18, and KRT8 genes were overexpressed relative to those in PFF cells but had levels of expression that were similar to those observed in iPS cells. In contrast, three genes important in prostaglandin synthesis, PTGES (PGE synthase), PTGS1 (COX1), and PTGS2 (COX2), were strongly up-regulated in the iTR cells compared to those in iPS cells but not to PFF (GEO accession no. GSE15472).
FIG. 2.

Transcriptome analysis of porcine iTR cells by microarray analysis is shown. A) Hierarchical clustering of the microarray data of iTR1 and three iPS cells (IC1, ID4, and ID6) and parental fibroblast lines (PFF) reported previously [15] show clear branching of the iTR1 line from the pluripotent iPS cell lines and the somatic cells (PFF). B) Normalized intensities of the genes reveal striking differences in gene expression of the iTR1 line in comparison to the iPS and PFF cells. Log2-fold change in TR (C) and pluripotent (D) gene expression in iTR1 cells compared to iPS cells (red bars) and PFF (blue bars). E, F) Candidate gene expression analysis of iTR1 cells and teratomas. E) Semiquantitative RT-PCR analysis was conducted for expression of selected pluripotency and TR marker genes from cells at passage (p) 6 cultured in F2 medium. The “RT” lanes indicate control reactions without reverse transcriptase. F) Similar PCR analysis of reverse transcribed RNA from a teratoma derived from iTR1 at p7 is shown.
iTR cells also differed from iPS cells in expression of some but not all genes commonly associated with pluripotency (Fig. 2D). As expected, the POU5F1 gene was down-regulated (approximately 80%) compared to that in the iPS cells but was up-regulated relative to that in PFF cells. In other words, the gene was expressed in the TR cells but at lower levels than in the iPS cells. In contrast, the UTF1, PODXL, GCNF, and CDH2 genes showed quite similar levels of expression in iTR and iPS cells and were highly up-regulated in iTR compared to PFF progenitors. The CD9 gene showed a similar trend. The expression of several other well known “pluripotency genes,” including SOX2 and NANOG, could not be assessed by microarray, either because of inadequate annotation of the microarrays or because appropriate diagnostic probes were absent from the arrays.
The expression levels of a number of marker genes for TR were also examined by semiquantitative RT-PCR with RNA extracted from the iTR1 line at passage 6 (Fig. 2E). In general, these data were consistent with those of the microarray study performed at the earlier passage. Transcripts for the definitive gene markers for porcine TR, IFND and PAG, were strongly expressed, as were a number of more general TR markers, including CDX2, EOMES, and GATA3. The experiment also confirmed that the colonies expressed the pPOU5F1 gene relatively strongly and the pSOX2 gene relatively weakly and that the two human orthologs of these genes, hPOU5F1 and hSOX2, had not been silenced after reprogramming.
Telomerase Activity
Telomerase activity, which was uniformly high in iPS cells and low in MEF and founding PFF cells, was expressed in both the iTR1 and iTR2 cell lines (Fig. 3A). In the latter, activity was comparable to that of the iPS cell lines, while the former had lower telomerase activity, but this level of activity was still comparable to that of the well-studied human ESC H9 line.
FIG. 3.

Telomerase activity of iTR cells is shown. A) Telomerase activity was measured enzymatically in two iTR lines (iTR1 passage [p] 6, iTR2 p3), three iPS cells (IC1, ID4, and ID6, p10); parental PFF, p10; MEF, p4; and H9 hES cells, p41. The assay was performed in triplicate samples with 0.2 μg of total cell protein by using Trapeze-RT telomerase detection kit (Chemicon). Activities represent relative units of telomerase assessed by the quantity of primers (amole/sample) extended with telomeric repeats. Data are presented as means ± SEM. B) Relative production of estradiol by iPS cells and iTR cells. Values show the relative amounts of estradiol released into the medium after culturing two iPS cell lines (ID6 and IB3) and two iTR lines (iTR1 and iTR2) in F2 medium for 48 h (from Day 4 until Day 6 after passage when the cells were harvested). Bars (±SEM) represent estradiol concentrations (pg/ml) normalized to the total RNA amount (μg). ID6 and IB3 cells were at passage 38 and 35, respectively; iTR1 and iTR2 were at passage 35 and 19, respectively.
Production of Estradiol
Two iPS lines (ID6 and IB3) and two iTR lines (iTR1 and iTR2) were examined for production of estradiol. Daily production at Day 6 of culture was higher for the iTR than for the iPS lines (Fig. 3B).
Teratoma Formation
WheniTR1 cells were injected subcutaneously into the upper flanks of one nude and one NOD SCID mouse, both animals gave rise to solid tumors that resembled teratomas (Fig. 4, A and E). The central regions of the tumors were relatively uniform in cross-section (Fig. 4, B and F) and consisted largely of packed layers of what appeared to be epithelium-like cells (Fig. 4, C and H). Differentiated structures of identifiable tissue types were rarer, but islands of striated muscle could be observed, especially near the margins of the tumor close to where they attached to the body wall (Fig. 4, D and G). Tissue samples that had been obtained from small pieces of teratomas expressed genes of porcine and not murine origin and a range of markers for TR (Fig. 2F), including all of those detected in the original iTR cells. Coexpression of the pCDX2 and pPOU5F1 genes was again evident.
FIG. 4.

Teratoma formation from injected iTR1 cells is shown. A–D) Tumor derived from iTR1 p7 cells; (E to H) iTR1 p11 cells. CD-1 nude mouse 11 weeks after subcutaneous injection (A); a tumor 13 weeks after injection of iTR1 p11 cells into a NOD SCID mouse (in millimeters) (E). Cross-section of encapsulated tumors (in inches) (B, F); most tumors consisted of packed layers of epithelial-like cells (C, G, H), but islands of striated muscle were observed near the margins of the tumor close to where it attached to the peritoneum (D, G). Bars = 100 μm in C; 50 μm in D; 250 μm in G; and 100 μm in H.
Cell Sphere Formation
When iTR1 and iTR2 cells were passaged into culture dishes that possessed a nonadherent surface, as used with pluripotent cells to test their ability to form embryoid bodies, the cells formed floating spheres that often attained a large diameter (0.7–1.8 mm) (Fig. 5, A and B). Such spheres were quite unlike embryoid bodies in appearance and size (diameters of embryoid bodies from ID6 iPS cells were 0.12–0.36 mm) (Fig. 5C) and also in the fact that they consisted of a single layer of cells with an epithelioid appearance whose points of attachment were highly enriched in desmosome-like structures that resembled tight junctions (Fig. 5, D and E). Both the inner (i.e., vacuole-facing) and the outer surfaces of the cells were populated with microvilli (Fig. 5D).
FIG. 5.
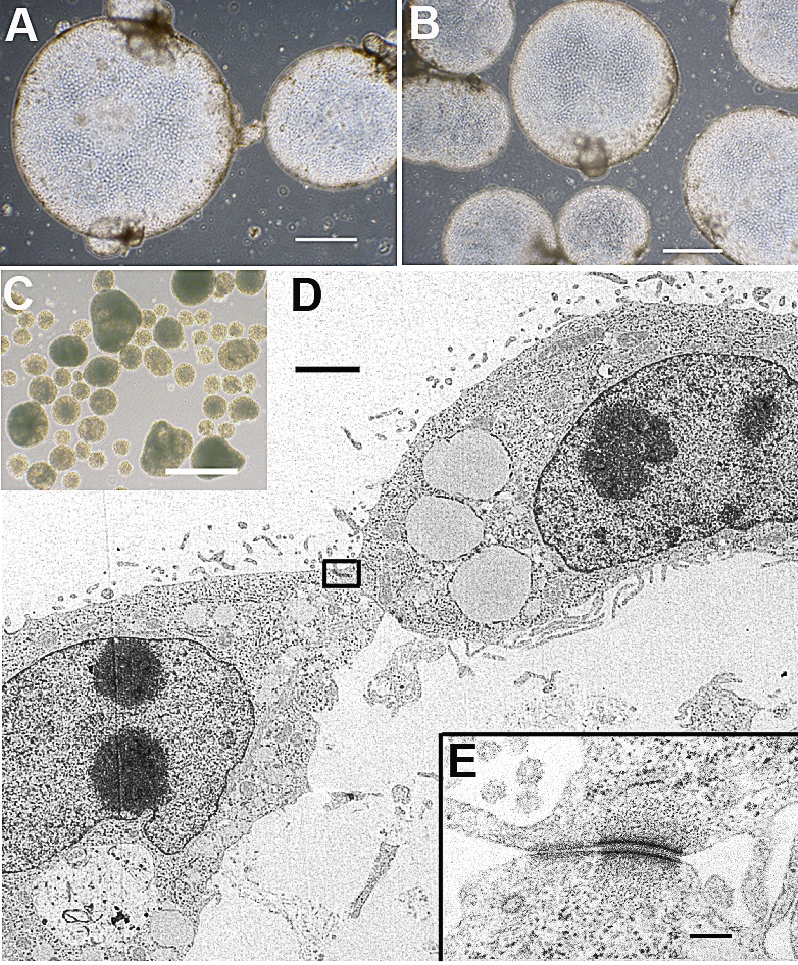
Cell sphere formation from iTR cells is shown. Spheres from iTR1 p42 (A) and iTR2 p7 cells are shown (B). C) Embryoid body formation from an iPS cell line (ID6, p5) is shown. Bars in A–C = 500 μm. D) Ultrastructure of a portion of a cell sphere from iTR2 cells (85-nm-thick section; bar = 2 μm). E) Cell junction (area boxed in D) shows a desmosome-like structure. Bar = 0.2 μm.
Karyotype of iTR Cell Lines
Cytogenetic analyses were performed with three iTR cell lines (iTR1, -2, and -3). Of the three, only iTR3 (38, XY) appeared to have a normal male karyotype as diagnosed by G-banding. The iTR1 line exhibited trisomy of chromosome 18 (39, XY + 18), while iTR2 had apparent addition of unknown genetic material on the long arm of chromosome 2 [38, XY, add(2)(q29)].
DISCUSSION
Here we have shown that when porcine mesenchymal cells, whether from fetal connective tissue or from umbilical cord, were subjected to standard reprogramming protocols, a significant fraction of the emergent colonies cultured on KOSR/FGF2 medium had features of TR. These colonies became apparent approximately a week earlier than the iPS cell colonies that were described previously [15] and were readily distinguishable in terms of their colony morphology. However, despite surfacing early, it is unclear whether the reprogrammed cells first passed through a transient, metastable, iPS cell stage that could not be maintained under the culture conditions employed or whether the conversion to TR was more direct and without such an intermediate step. Whatever the route of reprogramming of the PFF cells to TR, even well-established lines of human ESC tend to default to TR spontaneously under standard culture conditions, particularly if the culture is prolonged and carried out under a gas environment that features high, i.e., 20% O2, content [25, 26]. TR may therefore provide an alternative default state to pluripotency when the latter shows signs of being unsustainable. In the case of the iTR cell lines described here, it seems possible that genetic imbalance, evident as abnormal karyotypes in two of the lines, biased cell fate decisions and prompted lineage switching that favored TR. It is known, for example, that in chimeric mouse embryos, polyploid cells are shunted to the trophectoderm and subsequent TR lineages [27, 28]. Human ESC lines can also convert rapidly to TR upon exposure to BMP4 and to related growth factors [29–31] and when activin/nodal signaling is reduced [32]. Indeed, in the presence of BMP4 and absence of FGF2, transition to TR appears to be unidirectional and is relatively rapid, ultimately giving rise to more differentiated sublineages, including extravillous TR and syncytioTR [30], but whether this BMP-driven transition involves an intermediate but unstable TSC state is unclear. The fact that the iTR colonies contained a mixture of cells in the early passages and that as passage number was increased the flattened epithelial cells began to predominate suggests that the phenotype was not stable and that the more differentiated cells gradually out-competed the smaller cells. We hypothesize, but have not yet proved, that the latter cell populations are precursors of the former and may be composed of porcine TSC.
That the colonies at passage 2 were composed predominantly of porcine TR is evident from microarray data (Fig. 2). Moreover, this general aspect of their phenotype was maintained until at least passage 6, as evident from RT-PCR analyses (Fig. 2E). Even at passage 2, however, a pattern of gene expression consistent with differentiated as well as immature TR was evident (Fig. 2, C and D). Production of pregnancy-associated glycoproteins (PAG) [33], trypsin inhibitor UPTI [34], interferon-gamma (IFNG) [35], and interleukin-1β (IL1B) [24], for example, are characteristic of TR from elongating pig conceptuses during the period when placentation has been initiated and TR has begun to assume a transport function and is actively signaling to the mother [36]. Such early colonies also produce estrogen (Fig. 3B), release antiviral activity into the medium (data not shown), and form KRT7-positive domes that lift away from the substratum, all features of the presence of differentiating TR. Dome and vesicle formation from TR outgrowths of pig conceptuses [37] and production of estrogen [38, 39] are well-described phenomena associated with porcine TR. On the other hand, the passage 2 gene expression patterns, which were confirmed at passage 6, also suggested that a TSC population could be present simultaneously, as transcripts representing many genes typically associated with mouse TR stem cells, such as the GATA2, GATA3, TEAD4, ELF5, ETS2, SOX2, and CDX2 genes were all expressed (Fig. 2E). The lack of uniformity of the iTR colonies, their phenotypic instability over time, and the apparent propensity of cells within the colonies to differentiate will represent technical challenges if pure TSC are to be isolated and maintained in a self-renewing, stem-like state. The obvious solution, namely replacing the standard F2 medium with one supplemented with fetal bovine serum and substituting FGF4 with FGF2, conditions that have proven ideal for generating and maintaining mouse TSC [1, 40], have not yet been proven to provide such stabilizing conditions for porcine iTR cells (unpublished data).
One unanticipated feature of the putative porcine iTR cells was their expression of the pSOX2 and pPOU5F1 pluripotency gene markers, whose products are generally believed to be key components of a network that maintains self-renewal and pluripotent properties of ES cells [41, 42]. Of the two, pSOX2 transcript expression was less surprising, as Sox2 expression is known to be necessary for establishment of TR lineages in mouse embryos [43]. On the other hand, in mouse pluripotent cells, POU5F1 is considered to antagonize CDX2 in directing TR emergence, and its gene is thought to be regulated reciprocally relative to that of the Cdx2 gene, such that expression falls as TR differentiates [44, 45]. Either the transcriptional networks maintaining porcine iTR cells do not require complete silencing of the POU5F1 gene, or the coexpression is a reflection of the cellular heterogeneity of the colonies. However, coexpression of POU5F1 and CDX2 has been noted in trophectoderm of porcine blastocysts [46, 47], and TR emergence in bovine embryos is not accompanied by immediate POU5F1 transcriptional down-regulation, as observed in the mouse. This difference between mouse and bovine TR cells has been attributed to a lack of repressive elements that bind TCFAP2 (AP2) in a cis-acting regulatory region of the bovine POU5F1 promoter [48]. These repressive sequences may be unique to the mouse Pou5f1gene, as they are absent in a range of other species, including rats, shrews, and primates [48]. In view of the POU5F1 expression in the porcine TR cell lines, we predict that these AP2-binding motifs will also be absent from the pig gene.
iTR cells also had another feature of stem cells in that they generated solid, relatively uniform teratomas in nude mice. Unexpectedly, these tumors were quite unlike the rapidly forming, hemorrhagic structures containing large blood-filled lacunae that emerged after mouse and vole TSC had been injected into nude mice [49, 50]. One explanation for this difference in tumor type is that the porcine placenta is of the noninvasive, epitheliochorial type and may not contain cells capable of penetrating blood vessels. The solid tumors that formed were relatively uniform in cross section and composed largely of packed epithelial layers (Fig. 4) and expressed markers of differentiated TR. In fact, virtually the same porcine TR markers were evident in the teratomas as in the injected founder cells (Fig. 2F). Our observation that iTR cells gave rise to islands of striated muscle tissue within the teratomas, although not anticipated, is consistent with the observation that human chorionic villi transplanted into SCID/mdx mice can form striated muscle expressing human dystrophin [51] and provides further evidence that the iTR cells generated by reprogramming are multipotent.
The lack of full pluripotent potential of iTR is emphasized by their failure to form embryoid bodies after they had been dissociated and cultured on a nonadherent substratum in the absence of MEF. Instead of giving rise to compact multilayered structures containing cells representative of the three germ lines, the structures that formed consisted of variably sized, single-layered floating vesicles (Fig. 5), not unlike those that appeared after dome formation on the standard F2 medium.
Together, the above data indicate that reprogramming of porcine fibroblasts to iPS cells generates, as a by-product, colonies that are composed of self-renewing populations of TR cells. The number of such colonies relative to those that have the phenotype of fully reprogrammed iPS cells varies from experiment to experiment, emphasizing the empirical nature of the procedure. Although these colonies do not maintain a consistent phenotype over extended passage, they are clearly distinct from iPS cells and, at early passage, may contain cells that are self renewing, apparently multipotent, and in the early commitment stage to TR. It will be of interest if reprogramming of mesenchymal cells from other species through use of the same viral vectors can also be employed to generate an analogous type of TR cell.
ACKNOWLEDGMENTS
We thank Dr. Andrei Alexenko for assistance with derivation of teratomas, Sunilima Sinha for cell culture, Dr. Randall Prather and Lee Spate for providing EGFP-PFF, and the University of Missouri's Electron Microscopy Core Facility for assistance in preparing and imaging specimens.
Footnotes
Supported by National Institutes of Health grant HD-21896 (to R.M.R.), Agriculture and Food Research Initiative competitive grant 2011-67015-20070 from the USDA National Institute of Food and Agriculture (to R.M.R.), and Missouri Life Sciences Research Trust Fund grant 00022147 (to T.E.).
REFERENCES
- Tanaka S, Kunath T, Hadjantonakis AK, Nagy A, Rossant J. Promotion of trophoblast stem cell proliferation by FGF4. Science 1998; 282: 2072 2075 [DOI] [PubMed] [Google Scholar]
- Roberts RM, Fisher SJ. Trophoblast stem cells. Biol Reprod 2011; 84: 412 421 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vandevoort CA, Thirkill TL, Douglas GC. Blastocyst-derived trophoblast stem cells from the rhesus monkey. Stem Cells Dev 2007; 16: 779 788 [DOI] [PubMed] [Google Scholar]
- Tanaka S. Derivation and culture of mouse trophoblast stem cells in vitro. Methods Mol Biol 2006; 329: 35 44 [DOI] [PubMed] [Google Scholar]
- Guzman-Ayala M, Ben-Haim N, Beck S, Constam DB. Nodal protein processing and fibroblast growth factor 4 synergize to maintain a trophoblast stem cell microenvironment. Proc Natl Acad Sci U S A 2004; 101: 15656 15660 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miyazaki H, Imai M, Hirayama T, Saburi S, Tanaka M, Maruyama M, Matsuo C, Meguro H, Nishibashi K, Inoue F, Djiane J, Gertler A, et al. Establishment of feeder-independent cloned caprine trophoblast cell line which expresses placental lactogen and interferon tau. Placenta 2002; 23: 613 630 [DOI] [PubMed] [Google Scholar]
- Ramsoondar J, Christopherson RJ, Guilbert LJ, Wegmann TG. A porcine trophoblast cell line that secretes growth factors which stimulate porcine macrophages. Biol Reprod 1993; 49: 681 694 [DOI] [PubMed] [Google Scholar]
- Ka H, Jaeger LA, Johnson GA, Spencer TE, Bazer FW. Keratinocyte growth factor is up-regulated by estrogen in the porcine uterine endometrium and functions in trophectoderm cell proliferation and differentiation. Endocrinology 2001; 142: 2303 2310 [DOI] [PubMed] [Google Scholar]
- Flechon JE, Laurie S, Notarianni E. Isolation and characterization of a feeder-dependent, porcine trophectoderm cell line obtained from a 9-day blastocyst. Placenta 1995; 16: 643 658 [DOI] [PubMed] [Google Scholar]
- La Bonnardiere C, Flechon JE, Battegay S, Flechon B, Degrouard J, Lefevre F. Polarized porcine trophoblastic cell lines spontaneously secrete interferon-gamma. Placenta 2002; 23: 716 726 [DOI] [PubMed] [Google Scholar]
- Talbot NC, Caperna TJ, Edwards JL, Garrett W, Wells KD, Ealy AD. Bovine blastocyst-derived trophectoderm and endoderm cell cultures: interferon tau and transferrin expression as respective in vitro markers. Biol Reprod 2000; 62: 235 247 [DOI] [PubMed] [Google Scholar]
- Hashizume K, Shimada A, Nakano H, Takahashi T. Bovine trophoblast cell culture systems: a technique to culture bovine trophoblast cells without feeder cells. Methods Mol Med 2006; 121: 179 188 [PubMed] [Google Scholar]
- Patel OV, Yamada O, Kizaki K, Takahashi T, Imai K, Takahashi S, Izaike Y, Schuler LA, Takezawa T, Hashizume K. Expression of trophoblast cell-specific pregnancy-related genes in somatic cell-cloned bovine pregnancies. Biol Reprod 2004; 70: 1114 1120 [DOI] [PubMed] [Google Scholar]
- Takahashi K, Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006; 126: 663 676 [DOI] [PubMed] [Google Scholar]
- Ezashi T, Telugu BP, Alexenko AP, Sachdev S, Sinha S, Roberts RM. Derivation of induced pluripotent stem cells from pig somatic cells. Proc Natl Acad Sci U S A 2009; 106: 10993 10998 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Esteban MA, Xu J, Yang J, Peng M, Qin D, Li W, Jiang Z, Chen J, Deng K, Zhong M, Cai J, Lai L, et al. Generation of induced pluripotent stem cell lines from Tibetan miniature pig. J Biol Chem 2009; 284: 17634 17640 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu Z, Chen J, Ren J, Bao L, Liao J, Cui C, Rao L, Li H, Gu Y, Dai H, Zhu H, Teng X, et al. Generation of pig induced pluripotent stem cells with a drug-inducible system. J Mol Cell Biol 2009; 1: 46 54 [DOI] [PubMed] [Google Scholar]
- West FD, Terlouw SL, Kwon DJ, Mumaw JL, Dhara SK, Hasneen K, Dobrinsky JR, Stice SL. Porcine induced pluripotent stem cells produce chimeric offspring. Stem Cells Dev 2010; 19: 1211 1220 [DOI] [PubMed] [Google Scholar]
- Lai L, Prather RS. Production of cloned pigs by using somatic cells as donors. Cloning Stem Cells 2003; 5: 233 241 [DOI] [PubMed] [Google Scholar]
- Whitworth KM, Li R, Spate LD, Wax DM, Rieke A, Whyte JJ, Manandhar G, Sutovsky M, Green JA, Sutovsky P, Prather RS. Method of oocyte activation affects cloning efficiency in pigs. Mol Reprod Dev 2009; 76: 490 500 [DOI] [PubMed] [Google Scholar]
- Freshney I. Application of cell cultures to toxicology. Cell Biol Toxicol 2001; 17: 213 230 [DOI] [PubMed] [Google Scholar]
- Freshney RI. Culture of Animal Cells: A Manual of Basic Techniques. Hoboken, NJ: Wiley-Liss. 2005. [Google Scholar]
- Marcelo P, Lefevre F. Induction of the IFN-gamma gene and protein is linked to the establishment of cell polarity in a porcine epithelial cell line. Exp Cell Res 2002; 280: 33 44 [DOI] [PubMed] [Google Scholar]
- Ross JW, Malayer JR, Ritchey JW, Geisert RD. Characterization of the interleukin-1beta system during porcine trophoblastic elongation and early placental attachment. Biol Reprod 2003; 69: 1251 1259 [DOI] [PubMed] [Google Scholar]
- Ezashi T, Das P, Roberts RM. Low O2 tensions and the prevention of differentiation of hES cells. Proc Natl Acad Sci U S A 2005; 102: 4783 4788 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Westfall SD, Sachdev S, Das P, Hearne LB, Hannink M, Roberts RM, Ezashi T. Identification of oxygen-sensitive transcriptional programs in human embryonic stem cells. Stem Cells Dev 2008; 17: 869 881 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nagy A, Gocza E, Diaz EM, Prideaux VR, Ivanyi E, Markkula M, Rossant J. Embryonic stem cells alone are able to support fetal development in the mouse. Development 1990; 110: 815 821 [DOI] [PubMed] [Google Scholar]
- Tarkowski AK, Witkowska A, Opas J. Development of cytochalasin in B-induced tetraploid and diploid/tetraploid mosaic mouse embryos. J Embryol Exp Morphol 1977; 41: 47 64 [PubMed] [Google Scholar]
- Xu RH, Chen X, Li DS, Li R, Addicks GC, Glennon C, Zwaka TP, Thomson JA. BMP4 initiates human embryonic stem cell differentiation to trophoblast. Nat Biotechnol 2002; 20: 1261 1264 [DOI] [PubMed] [Google Scholar]
- Das P, Ezashi T, Schulz LC, Westfall SD, Livingston KA, Roberts RM. Effects of FGF2 and oxygen in the BMP4-driven differentiation of trophoblast from human embryonic stem cells. Stem Cell Research 2007; 1: 61 74 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Golos TG, Pollastrini LM, Gerami-Naini B. Human embryonic stem cells as a model for trophoblast differentiation. Semin Reprod Med 2006; 24: 314 321 [DOI] [PubMed] [Google Scholar]
- Wu Z, Zhang W, Chen G, Cheng L, Liao J, Jia N, Gao Y, Dai H, Yuan J, Xiao L. Combinatorial signals of activin/nodal and bone morphogenic protein regulate the early lineage segregation of human embryonic stem cells. J Biol Chem 2008; 283: 24991 25002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Szafranska B, Xie S, Green J, Roberts RM. Porcine pregnancy-associated glycoproteins: new members of the aspartic proteinase gene family expressed in trophectoderm. Biol Reprod 1995; 53: 21 28 [DOI] [PubMed] [Google Scholar]
- Duffy JY, Stallings-Mann ML, Trout WE, Roberts RM. Expression of a plasmin/trypsin Kunitz inhibitor by pig trophoblast. Mol Reprod Dev 1997; 46: 443 449 [DOI] [PubMed] [Google Scholar]
- Lefevre F, Martinat-Botte F, Guillomot M, Zouari K, Charley B, La Bonnardiere C. Interferon-gamma gene and protein are spontaneously expressed by the porcine trophectoderm early in gestation. Eur J Immunol 1990; 20: 2485 2490 [DOI] [PubMed] [Google Scholar]
- Geisert RD, Ross JW, Ashworth MD, White FJ, Johnson GA, DeSilva U. Maternal recognition of pregnancy signal or endocrine disruptor: the two faces of oestrogen during establishment of pregnancy in the pig. Soc Reprod Fertil Suppl 2006; 62: 131 145 [PubMed] [Google Scholar]
- Whyte A, Wooding FB, Saunders PT. Characteristics of cellular structures derived by tissue culture of pre-implantation blastocysts of the pig. Arch Anat Microsc Morphol Exp 1986; 75: 91 109 [PubMed] [Google Scholar]
- Conley AJ, Christenson LK, Ford SP, Christenson RK. Immunocytochemical localization of cytochromes P450 17 alpha-hydroxylase and aromatase in embryonic cell layers of elongating porcine blastocysts. Endocrinology 1994; 135: 2248 2254 [DOI] [PubMed] [Google Scholar]
- Geisert RD, Yelich JV. Regulation of conceptus development and attachment in pigs. J Reprod Fertil Suppl 1997; 52: 133 149 [PubMed] [Google Scholar]
- Quinn J, Kunath T, Rossant J. Mouse trophoblast stem cells. Methods Mol Med 2006; 121: 125 148 [DOI] [PubMed] [Google Scholar]
- Pan G, Thomson JA. Nanog and transcriptional networks in embryonic stem cell pluripotency. Cell Res 2007; 17: 42 49 [DOI] [PubMed] [Google Scholar]
- Orkin SH, Wang J, Kim J, Chu J, Rao S, Theunissen TW, Shen X, Levasseur DN. The transcriptional network controlling pluripotency in ES cells. Cold Spring Harb Symp Quant Biol 2008; 73: 195 202 [DOI] [PubMed] [Google Scholar]
- Avilion AA, Nicolis SK, Pevny LH, Perez L, Vivian N, Lovell-Badge R. Multipotent cell lineages in early mouse development depend on SOX2 function. Genes Dev 2003; 17: 126 140 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen L, Yabuuchi A, Eminli S, Takeuchi A, Lu CW, Hochedlinger K, Daley GQ. Cross-regulation of the Nanog and Cdx2 promoters. Cell Res 2009; 19: 1052 1061 [DOI] [PubMed] [Google Scholar]
- Strumpf D, Mao CA, Yamanaka Y, Ralston A, Chawengsaksophak K, Beck F, Rossant J. Cdx2 is required for correct cell fate specification and differentiation of trophectoderm in the mouse blastocyst. Development 2005; 132: 2093 2102 [DOI] [PubMed] [Google Scholar]
- Kirchhof N, Carnwath JW, Lemme E, Anastassiadis K, Scholer H, Niemann H. Expression pattern of Oct-4 in preimplantation embryos of different species. Biol Reprod 2000; 63: 1698 1705 [DOI] [PubMed] [Google Scholar]
- Kuijk EW, Du Puy L, Van Tol HT, Oei CH, Haagsman HP, Colenbrander B, Roelen BA. Differences in early lineage segregation between mammals. Dev Dyn 2008; 237: 918 927 [DOI] [PubMed] [Google Scholar]
- Berg DK, Smith CS, Pearton DJ, Wells DN, Broadhurst R, Donnison M, Pfeffer PL. Trophectoderm lineage determination in cattle. Dev Cell 2011; 20: 244 255 [DOI] [PubMed] [Google Scholar]
- Grigor'eva EV, Shevchenko AI, Mazurok NA, Elisaphenko EA, Zhelezova AI, Shilov AG, Dyban PA, Dyban AP, Noniashvili EM, Slobodyanyuk SY, Nesterova TB, Brockdorff N, et al. FGF4 independent derivation of trophoblast stem cells from the common vole. PLoS One 2009; 4: e7161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kibschull M, Nassiry M, Dunk C, Gellhaus A, Quinn JA, Rossant J, Lye SJ, Winterhager E. Connexin31-deficient trophoblast stem cells: a model to analyze the role of gap junction communication in mouse placental development. Dev Biol 2004; 273: 63 75 [DOI] [PubMed] [Google Scholar]
- Park TS, Gavina M, Chen CW, Sun B, Teng PN, Huard J, Deasy BM, Zimmerlin L, Peault B. Placental Perivascular Cells for Human Muscle Regeneration. Stem Cells Dev 2011; 20: 451 463 [DOI] [PMC free article] [PubMed] [Google Scholar]