

Abstract
The novel object recognition task is gaining popularity for its ability to test a complex behavior which relies on the integrity of memory and attention systems without placing undue stress upon the animal. While the task places few requirements upon the animal, it traditionally requires the experimenter to observe the test phase directly and record behavior. This approach can severely limit the number of subjects which can be tested in a reasonable period of time, as training and testing occur on the same day and span several hours. The current study was designed to test the feasibility of automation of this task for adolescent rats using standard activity chambers, with the goals of increased objectivity, flexibility, and throughput of subjects.
Keywords: Object recognition, adolescence, activity chambers
The novel object recognition task has been developed (Ennaceur & Delacour, 1988, Ennaceur et al., 1989) as a task which can be configured to measure working memory, attention, anxiety, and preference for novelty in rodents. This behavioral measure is gaining popularity as an indicator of susceptibility to drug abuse in adolescent populations (Ciccocioppo et al., 2002; Heyser et al., 2004; Morrow, Elsworth, & Roth, 2002; O'Shea et al., 2004; Piper, Fraiman, & Meyer, 2005; Stanfield & Kirstein, 2005b). The novel object recognition task is particularly attractive for several reasons: first, it requires no external motivation, reward or punishment; second, little training or habituation is required; and third, the task can be completed in a relatively short time. For these reasons, the novel object recognition task is excellent for testing animals which have received previous treatments which might alter the reward system, food and water intake or general stress levels.
Different populations of rats have been shown to vary in their reactions to novelty. Within the Roman rat strain, Roman High Avoidance animals showed increased reactivity to novel spatial arrangements in a familiarized environment relative to Roman Low Avoidance animals. Within the High Avoidance group, males were more reactive to novelty than females (Pisula, 2003). Within the Sprague Dawley strain, comparisons among males have shown adolescents to be more reactive to novelty than male adults. Specifically, adolescents displayed higher activity levels in a novel environment, more rapidly approached a novel object in a familiar environment, and spent more time with a novel object relative to adults (Stanfield & Kirstein, 2005a). Collectively, these findings suggest inherent differences in preference for novelty, dependent upon strain, sex, and age.
The novel object recognition task, while depending on preference for novelty, requires more cognitive skills from the subject, relative to tasks measuring exploration of novel environments or a single novel object. In order to discriminate between a novel and a familiar object, the subject must first attend to two identical objects and keep the two objects in working memory. Upon replacement of one of the familiarized objects with a novel object, if the animal can recognize that one object is novel, the animal will typically display differential behavior directed towards the novel object. Scoring of the task has often involved experimenter recording of the time spent with a novel object versus time spent with a familiar object, and calculation of a novelty or discrimination index based on these measures (Ennaceur, Neave, & Aggleton, 1997). In order to fully describe behavior in this task and to capture all variables of interest, the test session must be recorded in some manner. Recent studies have used videotaping and software assisted experimenter observation (Belcher et al., 2005, 2006; Ennaceur et al., 2005; He et al., 2006). Experimenter observation of each animal is time-consuming, and can hinder the ability to use many subjects, particularly in studies requiring exact timing (such as following a pharmaceutical or lesion treatment or a developmental exposure) or many treatment groups (such as dose-response studies).
The current study explores another method for recording behavior in this task. Adolescent male Sprague-Dawley rats were tested for novel object recognition using commercially available activity chambers and software (MotorMonitor, Hamilton-Kinder, San Diego, CA), as well as observational scoring by the experimenter. Activity chambers, a common instrument in behavioral laboratories, hold promise for providing additional data relevant to the task, allowing for assessment of habituation to the environment as well as movement in relation to the objects. This study addresses the possibility of automation of the task for adolescent rats using activity chambers.
Methods
Subjects
Subjects were 7 adolescent male rats. Sprague-Dawley animals were born in a breeding approved animal colony at the University of South Carolina, weaned on postnatal day 21, and group housed in same sex pairings until testing on postnatal day 35. The mean weight at testing was 128g. Only one male per litter was used in the experiment. Rodent food (Pro-Lab Rat, Mouse Hamster Chow #3000) and water were provided ad lib. The colony was maintained at 21 ± 2 °C, 50 ± 10% relative humidity and a 12L: 12D cycle with lights on at 0700 h (EST). Behavioral testing began at ∼1100 h. The protocol for this research was approved by the Institutional Animal Care and Use Committee (IACUC) of the University of South Carolina, Columbia.
Novelty Testing
Preliminary studies in this laboratory suggested the following protocols for achieving positive response to novelty in the adolescent rats. Each animal received one 6 minute period of habituation to the empty activity chamber on postnatal day 34 and was immediately returned to the home cage. The activity monitors (Figures 1a, 2a) were square (40 × 40 cm) chambers (Flex-Field, San Diego Instruments, San Diego, CA) that detected free movement of animals by infrared photocell interruptions. This equipment used an infrared photocell grid (32 emitter/detector pairs) to measure total locomotor activity. All activity monitors were located in an isolated room under dim light conditions, in the absence of direct overhead lighting (< 10 lx).
Figure 1.
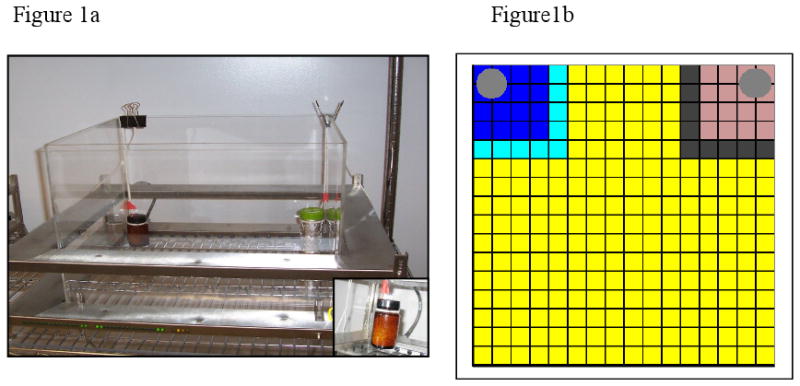
(Panel A) The automated activity monitor as configured for adolescent novelty testing. The inset shows an enlarged view of one object. Objects are suspended to avoid obscuring of the photobeams. The objects also block the animals' access to the rear corners. Lighting conditions in the photograph are not indicative of testing conditions. (Panel B) The MotorMonitor software produced zone map for the activity chamber. The dark blue and light purple squares represent the 5 × 5 photobeam grids around the objects. The light blue and dark purple squares represent the 6 × 6 photobeam grids. The grey circles represent the objects.
Figure 2.

Comparison of the novelty index scores for the adolescent animals produced by different methods of data collection. All indexes are positive, indicating recognition and preference for the novel object over the familiar object. No significant difference was found between the indices for the adolescent animals (n = 7). Error bars denote S.E.M.
Twenty-four hours following the first habituation session, each animal was returned to the activity chamber for 4 habituation and testing sessions, each 6 minutes long, separated by one-hour intervals. Session 1 consisted of habituation to the empty activity chamber. For session 2, two identical objects were placed into the chamber. The objects were placed in the rear corners of the chamber and suspended 0.5 cm above the floor to avoid obscuring of photobeams by the objects. Objects were selected for similarity of size and complexity (For review of object selection, see Bevins & Besheer, 2007; Ennaceur & Delacour, 1988). In the current study, the familiarized object was an aluminum cylinder, approximately 12 cm tall and 4 cm wide, or a black glass bottle, approximately 8 cm tall and 6 cm wide (Figure 1a). The animals were unable to move the objects or climb onto or under the objects. For session 3, the configuration of objects used in session two was repeated. For session 4, the test session, one familiarized object was replaced with a novel object (whichever object had not been previously used). Both choice of object and side of presentation were alternated.
Data Analysis
Using Motor Monitor (Hamilton-Kinder, San Diego, CA) software, activity in the chamber was measured. This software allows for post-hoc definition of zones of interest. A zone of any size can be defined by selecting grid squares of photobeam intersections. With the MotorMonitor interface, a small zone (5 × 5 photobeam grid) and a larger zone (6 × 6 photobeam grid) were designated around each object (Figures 1b). The software collected the total activity, number of entries into each zone (object approaches), and the time in each zone. Simultaneously, time spent with the objects was also recorded by the experimenter with a stopwatch. Observational time included only time during which the animal was in close proximity or actively touching or sniffing the object. A novelty index was calculated using the formula NI = (n-f)/(n+f), where n = time with novel object, f = time with familiar object. This index ranges from -1 to 1, with a -1 signifying complete preference for the familiar object, 0 signifying no preference, and 1 signifying complete preference for the novel object. Statistical evaluations were made using Anaylsis of Variance (ANOVA) and planned comparisons. An α level of 0.05 was considered significant for all statistical tests employed.
Results
Observational data indicated that each animal spent more time with the novel than the familiar object, producing a positive mean novelty discrimination index of 0.62 ± 0.11. Overall, automated data produced novelty indexes comparable to those produced by observational data. Zones of two sizes were examined around the novel and familiar objects in order to compare automated data and observational data. Both automated zone sizes produced index scores which were positive and indicated preference for the novel object over the familiar object (Figure 2). Analysis of Variance revealed no significant differences between the methods of data collection (observational, small zone, large zone) in index data. However, for the total raw time (Table 1) data, a 2 (method: observation, small zone automated) × 2 (object; novel, familiar) ANOVA revealed a significant interaction between method and object [F (1, 24) = 5.62, p < 0.05]. Post hoc analysis with Bonferroni test revealed that automated method recorded the animals spending significantly more time with the novel object [t (24) = 3.32, p < 0.05]. Total time spent with either the novel or familiar object was not significantly increased by increasing the size of the zones.
Table 1.
Mean Values ± SEM for variables of interest for the novel and familiar objects as measured by each method of data collection. Total time, resting time, and moving time are presented in seconds.
| Observational | Small Zone | Large Zone | ||||
|---|---|---|---|---|---|---|
| Novel | Familiar | Novel | Familiar | Novel | Familiar | |
| Total Time | 28.01±10.01 | 6.53±1.84 | 137.42±45.41 | 5.48±1.95 | 165.74±45.24 | 13.35±3.94 |
| Resting | 103.8±38.95 | 1.44±0.84 | 112.10±39.61 | 3.68±1.76 | ||
| Moving | 33.63±7.45 | 4.04±1.19 | 53.63± 9.74 | 9.67± 2.48 | ||
| Entries | 17.00±2.58 | 6.00±1.43 | 30.86±5.53 | 13.71±3.29 | ||
Time spent with the object was divided into time resting or moving within the zone (Table 1). ANOVA revealed significantly more time spent resting in the zone around the novel object than the zone around the familiar object [F (1, 24) = 14.38, p < 0.01] regardless of zone size. Significantly more time was spent moving in the zone around the novel object than the zone around the familiar object [F (1, 24) = 34.21, p < 0.01] regardless of zone size. ANOVA comparing the observational total time data to the moving time data (small zone) X the two objects (novel, familiar) revealed no effect of method of collection, with both methods showing more time with the novel than the familiar object [F (1, 24) = 16.25, p < 0.01]. Cronbach's alpha reliability coefficient comparing observational total time to moving time (small zone) was 0.70 for the novel object and 0.69 for the familiar object.
The number of entries into the zones around the objects also revealed preferential approaches to the novel object. In the number of entries into the zones around the objects (object approaches), ANOVA revealed no interaction between zone size and object. Both zone sizes detected more approaches to the novel object than the familiar [F (1, 24) = 15.78, p < 0.01], and the larger zone size detected more approaches to both objects than the smaller zone [F (1, 24) = 9.270, p < 0.01].
Discussion
The current study investigated use of the novel object recognition task in automated activity chambers in adolescent male rats. All subjects demonstrated the ability to detect the novel object following habituation. Observational and automated data produced similar novelty indexes. Automated data collection significantly increased the total amount of time spent with the novel object, but not time spent with the familiar object. This increase in time spent near the novel object was insufficient to significantly alter the novelty index. Because the total time measure produced generated by the automated data is a measure of proximity to the novel object, it indicated that the animals spent time in the zone around the novel object that was not considered exploratory by the experimenter observation. Analyzing time spent moving and time spent resting in the object zones (measures provided by MotorMonitor) allowed for more accurate approximation of the time data. The time spent moving in the zones around the objects closely resembled the observational total time. While the time spent resting in the object zones did not resemble the observational data, it too could be considered a behavioral response to novelty, further indicating that the animal preferred the novel to the familiar.
The novel object recognition task has been successfully used to demonstrate differences in novelty processing based on rat strain and sex (Ennaceur et al., 2005). The task also has been used successfully to demonstrate alterations in novelty processing based on drug treatments. Prenatal cocaine was shown to disrupt performance in this task in adolescent and adult male rats. Animals exposed to prenatal cocaine displayed preference for the novel object when tested after 20 minutes, but no preference for the novel object after either 1 or 24 hr (Morrow et al., 2002), possibly indicating deficits in short-term memory in the task. Administration of methamphetamine in adult rats decreased the novelty index significantly but not dramatically, possibly indicating altered preference for novelty following the drug (Belcher et al., 2005; 2006). These studies suggest that the novel object recognition task is sensitive enough to detect alterations in behavior in populations of animals expected to differ on measures of attention, motivation, or memory. Drug abuse studies in particular stress the attraction of automation of this task, which would allow for increased cohort size by allowing testing of many subjects on the same postnatal day or post-drug exposure day.
With thorough exploration of experimental parameters, it appears possible to automate the novel object recognition task using activity chambers. Additional studies from this laboratory have been performed with adolescent female rats and young adults (postnatal day 70) with similar reliability of this automated method. Specifically, both adolescent females and adult males demonstrated positive novelty index scores with no significant difference between methods of data collection; however, the scores were too low to indicate strong preference for novelty, unlike that reported in adolescent males (unpublished data). Interpretation of such results has been discussed extensively elsewhere (for example, Bevins & Besheer, 2006; and Ennaceur et al., 2005). One caveat of the current studies is that animal size must be considered. For adult animals, it was necessary to increase the size of the zone defined around the objects in order to closely approximate time spent with the objects. Also, this protocol may not be suitable for larger rats, as their increased body size may allow for proximity to both objects at once.
The currently described protocols were designed for using photo-beam based activity chambers as an alternative to using observational data alone or the use of video-based systems. Video-based systems are attractive for their ability to record the test session permanently for observational scoring and re-analysis. In the currently described system, a permanent recording is made which can be watched by the experimenter; however it is only a representation of the location of the subject, not a video image. As such the recording does not indicate orientation of the animal. The automated activity chambers do offer several advantages over video-based systems. Automation of this task with activity chambers has the potential to dramatically increase throughput. Because the MotorMonitor software allows for designation of zones of interest at the time of analyzing the data, no field has to be defined visually around the objects for detection of zone entry, as with some video-based behavior analysis software. The size of the zone can also be manipulated at any time to investigate proximity to the object, and to aid in judgment as to the exploratory nature of time spent near the object. With consideration of both raw data and index scores, an accurate representation of discriminate behavior in this task can be obtained from automated data. In conclusion, these studies emphasize the importance of careful analysis and interpretation of results from the novel object recognition task and the potential of automated protocols.
Acknowledgments
Supported by: NIDA grants DA009160, DA013965, DA013137, DA013712, and DA014401
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Belcher AM, O'Dell SJ, Marshall JF. Impaired object recognition memory following methamphetamine, but not p-chloroamphetamine- or d-amphetamine-induced neurotoxicity. Neuropsychopharmacology. 2005;30:2026–34. doi: 10.1038/sj.npp.1300771. [DOI] [PubMed] [Google Scholar]
- Belcher AM, O'Dell SJ, Marshall JF. A sensitizing regimen of methamphetamine causes impairments in a novelty preference task of object recognition. Behav Brain Res. 2006;170:167–72. doi: 10.1016/j.bbr.2006.02.025. [DOI] [PubMed] [Google Scholar]
- Bevins RA, Besheer J. Object recognition in rats and mice: a one-trial non-matching-to-sample learning task to study “recognition memory”. Nat Protoc. 2006;1:1306–1311. doi: 10.1038/nprot.2006.205. [DOI] [PubMed] [Google Scholar]
- Ciccocioppo R, Antonelli L, Biondini M, Perfumi M, Pompei P, Massi M. Memory impairment following combined exposure to delta(9)-tetrahydrocannabinol and ethanol in rats. Eur J Pharmacol. 2002;449:245–52. doi: 10.1016/s0014-2999(02)01999-4. [DOI] [PubMed] [Google Scholar]
- Ennaceur A, Delacour J. A new one-trial test for neurobiological studies of memory in rats. 1: Behavioral data. Behav Brain Res. 1988;31:47–59. doi: 10.1016/0166-4328(88)90157-x. [DOI] [PubMed] [Google Scholar]
- Ennaceur A, Cavoy A, Costa JC, Delacour J. A new one-trial test for neurobiological studies of memory in rats. II: Effects of piracetam and pramiracetam. Behav Brain Res. 1989;33:197–207. doi: 10.1016/s0166-4328(89)80051-8. [DOI] [PubMed] [Google Scholar]
- Ennaceur A, Michalikova S, Bradford A, Ahmed S. Detailed analysis of the behavior of Lister and Wistar rats in anxiety, object recognition and object location tasks. Behav Brain Res. 2005;159:247–66. doi: 10.1016/j.bbr.2004.11.006. [DOI] [PubMed] [Google Scholar]
- Ennaceur A, Neave N, Aggleton JP. Spontaneous object recognition and object location memory in rats: the effects of lesions in the cingulate cortices, the medial prefrontal cortex, the cingulum bundle and the fornix. Exp Brain Res. 1997;113:509–19. doi: 10.1007/pl00005603. [DOI] [PubMed] [Google Scholar]
- He J, Yang Y, Yu Y, Li X, Li XM. The effects of chronic administration of quetiapine on the methamphetamine-induced recognition memory impairment and dopaminergic terminal deficit in rats. Behav Brain Res. 2006;172:39–45. doi: 10.1016/j.bbr.2006.04.009. [DOI] [PubMed] [Google Scholar]
- Heyser CJ, Pelletier M, Ferris JS. The effects of methylphenidate on novel object exploration in weanling and periadolescent rats. Annals of the New York Academy of Sciences. 2004;1021:465–469. doi: 10.1196/annals.1308.066. [DOI] [PubMed] [Google Scholar]
- Morrow BA, Elsworth JD, Roth RH. Prenatal cocaine exposure disrupts non-spatial, short-term memory in adolescent and adult male rats. Behav Brain Res. 2002;129:217–23. doi: 10.1016/s0166-4328(01)00338-2. [DOI] [PubMed] [Google Scholar]
- O'Shea M, Singh ME, McGregor IS, Mallet PE. Chronic cannabinoid exposure produces lasting memory impairment and increased anxiety in adolescent but not adult rats. J Psychopharmacol. 2004;18:502–8. doi: 10.1177/026988110401800407. [DOI] [PubMed] [Google Scholar]
- Piper BJ, Fraiman JB, Meyer JS. Repeated MDMA (“Ecstasy”) exposure in adolescent male rats alters temperature regulation, spontaneous motor activity, attention, and serotonin transporter binding. Dev Psychobiol. 2005;47:145–57. doi: 10.1002/dev.20085. [DOI] [PubMed] [Google Scholar]
- Pisula W. The Roman high- and low-avoidance rats respond differently to novelty in a familiarized environment. Behav Processes. 2003;63:63–72. doi: 10.1016/s0376-6357(03)00032-9. [DOI] [PubMed] [Google Scholar]
- Stansfield KH, Kirstein CL. Effects of novelty on behavior in the adolescent and adult rat. Dev Psychobiol. 2005a;48:10–15. doi: 10.1002/dev.20127. [DOI] [PubMed] [Google Scholar]
- Stansfield KH, Kirstein CL. Neurochemical effects of cocaine in adolescence compared to adulthood. Dev Brain Res. 2005b;159:119–125. doi: 10.1016/j.devbrainres.2005.07.005. [DOI] [PubMed] [Google Scholar]