

Abstract
E1210 is a new antifungal compound with a novel mechanism of action and broad spectrum of antifungal activity. We investigated the in vitro antifungal activities of E1210 compared to those of fluconazole, itraconazole, voriconazole, amphotericin B, and micafungin against clinical fungal isolates. E1210 showed potent activities against most Candida spp. (MIC90 of ≤0.008 to 0.06 μg/ml), except for Candida krusei (MICs of 2 to >32 μg/ml). E1210 showed equally potent activities against fluconazole-resistant and fluconazole-susceptible Candida strains. E1210 also had potent activities against various filamentous fungi, including Aspergillus fumigatus (MIC90 of 0.13 μg/ml). E1210 was also active against Fusarium solani and some black molds. Of note, E1210 showed the greatest activities against Pseudallescheria boydii (MICs of 0.03 to 0.13 μg/ml), Scedosporium prolificans (MIC of 0.03 μg/ml), and Paecilomyces lilacinus (MICs of 0.06 μg/ml) among the compounds tested. The antifungal action of E1210 was fungistatic, but E1210 showed no trailing growth of Candida albicans, which has often been observed with fluconazole. In a cytotoxicity assay using human HK-2 cells, E1210 showed toxicity as low as that of fluconazole. Based on these results, E1210 is likely to be a promising antifungal agent for the treatment of invasive fungal infections.
INTRODUCTION
Invasive fungal infections have become increasingly common among immunocompromised or immunosuppressed patients, including solid-organ or hematopoietic stem cell transplant recipients and individuals who are on immunosuppressive drug regimens (6, 12, 18). There is still a high rate of morbidity and mortality associated with invasive fungal infections, because the currently available antifungal drugs are limited in terms of cost, toxicity, pharmacokinetic variability, and complex drug interactions. Amphotericin B is a fungicidal drug with broad-spectrum activity, but it is exclusively a parenteral agent and has significant toxicity (1). Azole antifungal drugs, such as fluconazole, itraconazole, voriconazole, and posaconazole, are now widely used for the treatment of serious fungal infections due to their broad spectrum of activity and more favorable safety profile. However, isolates resistant to azoles are increasingly being reported (16). Echinocandin derivatives, such as caspofungin, micafungin, and anidulafungin, are broad-spectrum antifungal agents with favorable safety properties but are exclusively parenteral agents and are not active against Cryptococcus neoformans (5). Hence, there is a critical need for new antifungal compounds that have broad-spectrum activity, different mechanisms of action, and no cross-resistance to the currently available antifungal drugs.
Glycosylphosphatidylinositol (GPI)-anchored mannoproteins are one of the major cell wall components of eukaryotic microorganisms, including fungi. In fungi, more than half of GPI-anchored proteins have an unknown function, but it is noted that some of them are involved in and are likely vital to cell wall regeneration and adhesion to host cells (20). Therefore, Eisai Co., Ltd., focused on the inhibition of GPI-anchored-protein biosynthesis as a therapeutic target for the treatment of fungal infections. By screening chemical libraries that inhibited fungal cell wall assembly via GPI-anchored proteins in yeast, we found an inhibitor, 1-(4-butylbenzyl)isoquinoline, and its target molecule, the protein GWT1, which is required for inositol acylation at an early step of the GPI biosynthetic pathway (25, 26). In our efforts to optimize the resultant hit compounds by using a medicinal chemistry approach (11, 23), we discovered a novel drug candidate compound, E1210, i.e., 3-(3-{4-[(pyridin-2-yloxy)methyl]benzyl}isoxazol-5-yl)pyridin-2-amine (Fig. 1), as a potential drug candidate based on its expected pharmacological, toxicological, and pharmacokinetic properties (7, 9, 15). The purpose of this study was to examine the in vitro antifungal activities of E1210 against clinically relevant fungal pathogen isolates compared to those of approved, control antifungal agents.
Fig. 1.
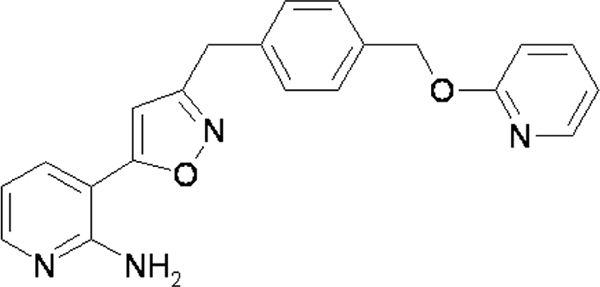
Chemical structure of E1210.
(This work was presented in part at the 50th Interscience Conference on Antimicrobial Agents and Chemotherapy, Boston, MA, 12 to 15 September 2010.)
MATERIALS AND METHODS
Compounds.
E1210 was synthesized at Eisai Co., Ltd. (Tokyo, Japan). Fluconazole and voriconazole were obtained from Pfizer Japan, Inc. (Tokyo, Japan). Micafungin was obtained from Astellas Pharma, Inc. (Tokyo, Japan). Itraconazole and amphotericin B were purchased from Sigma-Aldrich Co., Ltd. (St. Louis, MO). All compounds were dissolved in and diluted with dimethyl sulfoxide (DMSO) before use.
Organisms.
In total, 246 strains of yeast and filamentous fungi were studied. They were clinical pathogens isolated mainly in Japan and were provided by the Medical Mycology Research Center, Chiba University (Chiba, Japan), Graduate School of Medicine, Gifu University (Gifu, Japan), and the Institute for Fermentation, Osaka (Osaka, Japan). A quality control strain, Candida parapsilosis ATCC 22019, was obtained from the American Type Culture Collection (Manassas, VA).
Prior to testing, each yeast isolate, except for Malassezia furfur, was subcultured in Sabouraud dextrose broth (SDB; Becton, Dickinson and Company, Sparks, MD) and incubated at 35°C for 1 to 2 days. All of the filamentous fungi and dermatophytes except Fusarium solani were subcultured on potato dextrose agar (PDA; Eiken Chemical Co., Ltd., Tokyo, Japan) plates and incubated at 35°C for 1 to 3 weeks. F. solani was subcultured on PDA and incubated at 35°C for 2 to 3 days and then at 25°C for 4 to 5 days. M. furfur was subcultured on modified Dixon's agar plates (3.6% malt extract, 0.6% peptone, 1.5% agar, 2% ox bile, 1% Tween 40, 0.2% glycerol, and 0.2% oleic acid) and incubated at 32°C for 3 days (21, 27).
For cytotoxicity assay, HK-2 cells, a human proximal tubular cell line from healthy kidney tissue (ATCC CRL-2190), were obtained from the American Type Culture Collection.
Antifungal susceptibility studies.
The MICs of E1210 and the control antifungal agents were determined consistently using the broth microdilution methods developed by the Clinical and Laboratory Standards Institute and published in documents M27-A3 (2) and M38-A2 (3).
For all organisms except M. furfur, RPMI 1640 medium with 0.165 M morpholinepropanesulfonic acid (MOPS) buffer (pH 7.0) was used as the test medium. The medium for M. furfur was further supplemented with 1% glycerol, 1% peptone, 1.8% glucose, and 0.05% Tween 80 (modified RPMI 1640 medium) (21). The final concentrations of the antifungal agents studied ranged from 0.008 to 32 μg/ml for E1210, 0.06 to 64 μg/ml for fluconazole, 0.004 to 8 μg/ml for itraconazole and voriconazole, 0.016 to 16 μg/ml for amphotericin B, and 0.004 to 32 μg/ml for micafungin. For all organisms except M. furfur, the cell numbers in the yeast cultures or conidial suspensions were counted using a hemocytometer as a modification to the CLSI method. M. furfur cells were harvested using sterile saline containing 0.05% Tween 80 and vortexed with sterile glass beads to disperse Malassezia clumps. The cell densities were adjusted to a turbidity of 0.5 McFarland standard (Remel, Inc., Lenexa, KS) (27). The cell suspensions were diluted with the test medium to obtain an inoculum size of 1.5 × 103 to 3.8 × 103 cells/ml for the yeasts or 5 × 103 to 2.5 × 104 cells/ml for the filamentous fungi and dermatophytes. MICs were determined after incubation at 35°C for all isolates except M. furfur, which was incubated at 32°C. The microplates were incubated for 22 to 26 h for Candida tropicalis, Rhizopus spp., Mucor racemosus, and Cunninghamella bertholletiae, 70 to 74 h for Cryptococcus neoformans, Pseudallescheria boydii, and Paecilomyces lilacinus, 72 to 96 h for M. furfur, and 5 to 10 days for dermatophytes. For the other yeasts, Aspergillus spp., and Fusarium solani, the microplates were incubated for 46 to 50 h, and for the other filamentous fungi, the microplates were incubated for 3 to 6 days.
For all of the yeasts except M. furfur, growth reductions were assessed visually and based on the optical density at 660 nm. For the filamentous fungi, dermatophytes, and M. furfur, the growth reductions were graded visually with a numerical score, ranging from 0 to 4 using the scale established by the CLSI procedures. The MICs of all of the antifungal agents except E1210 were defined by the criteria of the CLSI procedures. For all of the yeasts except M. furfur, the MIC endpoints of E1210 were defined as the lowest concentrations resulting in a prominent decrease in turbidity (that is, a 50% reduction in growth determined spectrophotometrically) relative to that in a control well, like those of the azoles. For Aspergillus spp. and Pseudallescheria spp., the MIC of E1210 was determined by the minimum effective concentration (MEC) endpoints of echinocandin derivatives, because a typical MIC endpoint was not determinable when E1210 was tested against these species, as was also the case with micafungin. For the other filamentous fungi and M. furfur, the MIC endpoints of E1210 were defined as the lowest concentration at which a score of 2 was observed. For black molds, the MICs of itraconazole and voriconazole were defined as the lowest concentration at which a score of 2 was observed.
Time-growth inhibition assay.
Candida albicans IFM49971 was cultured in RPMI 1640 medium containing E1210 (0.001 to 0.06 μg/ml), fluconazole (0.03 to 2 μg/ml), or vehicle at 35°C with an inoculum size of 104 CFU/ml. After 2, 4, 8, and 24 h of incubation, a fraction was obtained from each culture tube, and that fraction was serially diluted in sterile saline and plated on Sabouraud dextrose agar plates. After 24 h of incubation at 35°C, the number of colonies was counted, and the viable cell count was expressed in CFU. The maximal fungistatic activity was defined as less than a 200% increase in the number of CFU per milliliter from the starting inoculum count.
In parallel, the MICs of E1210 and fluconazole for the test strain were determined using a broth macrodilution method (2). The MICs were read 1 day after incubation at 35°C and were defined as the lowest concentrations at which a prominent decrease in turbidity (a 50% reduction in growth) could be visually determined relative to that of the control tube.
Effects of culture conditions on MICs.
C. albicans IFM49971, Aspergillus fumigatus IFM51126, Aspergillus flavus IFM50915, F. solani IFM50956, and Rhizopus oryzae IFO5442 were used to study the effects of culture conditions on MICs. The effects of the inoculum size (5 × 102 to 5 × 104 cells/ml), the test medium (RPMI 1640, RPMI 1640 supplemented with 2% glucose, SDB, or antibiotic medium 3 [Becton, Dickinson and Company, Sparks, MD]), and the addition of mouse or human serum (0 to 90%) on MICs were evaluated with a broth microdilution method. The test fungi were cultured in a medium containing E1210 (0.001 to 4 μg/ml) or in vehicle at 35°C for 46 h for C. albicans, A. fumigatus, A. flavus, and F. solani and for 22 h for R. oryzae. The MICs were defined as the lowest concentrations at which a prominent decrease in turbidity (a 50% reduction in growth) could be visually determined relative to that of the control tube. Each experiment was repeated three times. The MICs of E1210 were defined as the median values.
Cytotoxicity assay.
HK-2 cells (5 × 103 cells/well) were cultured in Dulbecco's modified Eagle's medium (Invitrogen Corp., Carlsbad, CA) supplemented with 10% fetal bovine serum for 2 days at 37°C in the presence of 5% CO2. Cell viability was assayed with cell counting kit-8 (Dojindo Laboratories, Kumamoto, Japan), and drug concentrations reducing cell viability by 50% (IC50s) were determined.
RESULTS
MICs for clinical isolates of fungi.
Tables 1, 2, and 3 show the MICs of E1210 and the other antifungal compounds for the fungal isolates tested. E1210 showed potent in vitro antifungal activity against almost all Candida species tested. In particular, E1210 potently inhibited the growth of C. albicans (MIC90 of ≤0.008 μg/ml). Its activity was greater than those of fluconazole, itraconazole, amphotericin B, and micafungin and was comparable to that of voriconazole. Against Candida glabrata and Candida tropicalis, E1210 showed potent activities, with MIC90 values of 0.06 and 0.03 μg/ml, respectively; it was more active than fluconazole, itraconazole, voriconazole, and amphotericin B and was comparable to micafungin. E1210 also showed potent antifungal activity against Candida parapsilosis, with a MIC90 of 0.016 μg/ml. Its activity was 32 to 64 times higher than those of fluconazole, amphotericin B, and micafungin, 8 times higher than that of itraconazole, and comparable to that of voriconazole. E1210 also showed potent antifungal activities against the other Candida spp., Candida dubliniensis (MICs of ≤0.008 μg/ml), Candida guilliermondii (MIC of ≤0.008 μg/ml), and Candida lusitaniae (MIC of 0.06 μg/ml). However, the activity of E1210 against Candida krusei was moderate to poor, with a MIC range of 2 to >32 μg/ml, and was lower than those of the other compounds tested, except for fluconazole. Fluconazole resistance (MIC of ≥64 μg/ml) was observed in 18 of the 158 Candida strains tested, including 15 strains of C. tropicalis, two strains of C. albicans, and one strain of C. krusei. E1210 showed equally potent activities against fluconazole-resistant and fluconazole-susceptible Candida strains (MIC90 of 0.03 and 0.06 μg/ml, respectively), and no cross-resistance with azoles was observed (Table 3). Trailing growth was observed when azole antifungals were tested against Candida spp., while E1210 completely inhibited the growth of Candida spp. E1210 showed good activities against C. neoformans (MIC range of 0.13 to 0.5 μg/ml) and Trichosporon asahii (MIC range of 0.5 to 1 μg/ml), and its activities were greater than those of fluconazole and micafungin but were comparable to those of itraconazole, voriconazole, and amphotericin B. E1210 also showed good activity against M. furfur (MIC range of 0.13 to 0.25 μg/ml), and its activity was comparable to those of voriconazole and amphotericin B but was lower than that of itraconazole.
Table 1.
In vitro antifungal activities of E1210 against Candida spp. and Aspergillus fumigatus
| Organism (no. of strains) | Compound | MIC (μg/ml) |
||
|---|---|---|---|---|
| Range | MIC50 | MIC90 | ||
| Candida albicans (52) | E1210 | ≤0.008–0.016 | ≤0.008 | ≤0.008 |
| Fluconazole | ≤0.06–>64 | 0.13 | 0.25 | |
| Itraconazole | 0.008–>4 | 0.016 | 0.016 | |
| Voriconazole | ≤0.004–>4 | ≤0.004 | ≤0.004 | |
| Amphotericin B | 0.25–0.5 | 0.25 | 0.5 | |
| Micafungin | 0.008–0.03 | 0.016 | 0.016 | |
| Candida glabrata (44) | E1210 | ≤0.008–0.06 | 0.06 | 0.06 |
| Fluconazole | 4–32 | 4 | 16 | |
| Itraconazole | 0.25–2 | 0.5 | 1 | |
| Voriconazole | 0.06–1 | 0.13 | 0.25 | |
| Amphotericin B | 0.13–1 | 1 | 1 | |
| Micafungin | 0.008–0.03 | 0.03 | 0.03 | |
| Candida tropicalis (23) | E1210 | ≤0.008–0.03 | 0.016 | 0.03 |
| Fluconazole | 1–>64 | >64 | >64 | |
| Itraconazole | 0.03–>4 | 1 | >4 | |
| Voriconazole | 0.03–>4 | >4 | >4 | |
| Amphotericin B | 0.25–0.5 | 0.5 | 0.5 | |
| Micafungin | 0.016 | 0.016 | 0.016 | |
| Candida parapsilosis (26) | E1210 | ≤0.008–0.016 | ≤0.008 | 0.016 |
| Fluconazole | 0.13–1 | 0.5 | 1 | |
| Itraconazole | 0.03–0.25 | 0.13 | 0.13 | |
| Voriconazole | ≤0.004–0.03 | 0.016 | 0.016 | |
| Amphotericin B | 0.5 | 0.5 | 0.5 | |
| Micafungin | 0.13–1 | 1 | 1 | |
| Aspergillus fumigatus (20) | E1210 | 0.03–0.13 | 0.06 | 0.13 |
| Fluconazole | >64 | >64 | >64 | |
| Itraconazole | 0.5 | 0.5 | 0.5 | |
| Voriconazole | 0.13–0.5 | 0.25 | 0.5 | |
| Amphotericin B | 0.25–1 | 0.5 | 0.5 | |
| Micafungin | 0.008–0.06 | 0.03 | 0.06 | |
Table 2.
In vitro antifungal activities of E1210 against various clinical fungal isolates
| Organism (no. of strains) | MIC(s) (μg/ml) for individual isolate(s) (no. of isolates with MIC) |
|||||
|---|---|---|---|---|---|---|
| E1210 | Fluconazole | Itraconazole | Voriconazole | Amphotericin B | Micafungin | |
| Yeasts | ||||||
| Candida krusei (4) | 2 (1), 4 (2), >32 (1) | 32 (3), 64 (1) | 0.13 (2), 0.25 (1), 1 (1) | 0.25 (3), 0.5 (1) | 0.5 (1), 1 (3) | 0.06 (2), 0.13 (2) |
| Candida dubliniensis (7) | ≤0.008 (7) | ≤0.06 (1), 0.13 (2), 0.25 (4) | 0.016 (6), 0.03 (1) | ≤0.004 (1), 0.008 (3), 0.016 (2), 0.03 (1) | 0.13 (6), 0.25 (1) | 0.016 (6), 0.06 (1) |
| Candida guilliermondii (1) | ≤0.008 | 4 | 0.25 | 0.03 | 0.25 | 0.25 |
| Candida lusitaniae (1) | 0.06 | 0.5 | 0.016 | 0.008 | 0.25 | 0.06 |
| Cryptococcus neoformans (4) | 0.13 (1), 0.25 (2), 0.5 (1) | 2 (4) | 0.06 (3), 0.13 (1) | 0.03 (4) | 0.25 (4) | >32 (4) |
| Malassezia furfur (2) | 0.13 (1), 0.25 (1) | 2 (1), 8 (1) | ≤0.008 (2) | 0.03 (1), 0.25 (1) | 0.25 (1), 0.5 (1) | 16 (1), >32 (1) |
| Trichosporon asahii (3) | 0.5 (1), 0.25 (1), 1 (1) | 1 (3) | 0.5 (3) | 0.06 (2), 0.13 (1) | 1 (1), 2 (2) | >32 (3) |
| Filamentous fungi | ||||||
| Aspergillus flavus (4) | 0.016 (2), 0.06 (1), 0.13 (1) | >64 (4) | 0.25 (3), 0.5 (1) | 0.5 (3), 2 (1) | 1 (3), 2 (1) | 0.016 (1), 0.03 (3) |
| Aspergillus niger (2) | 0.016 (1), 0.03 (1) | >64 (2) | 2 (2) | 1 (1), 2 (1) | 0.13 (2) | 0.016 (1), 0.03 (1) |
| Aspergillus terreus (1) | 0.13 | >64 | 0.13 | 0.5 | 1 | 0.016 |
| Fusarium solani (7) | 0.03 (4), 0.25 (1), 0.13 (1), >32 (1) | >64 (7) | >4 (7) | 4 (2), >4 (5) | 1 (3), 2 (4) | >4 (7) |
| Paecilomyces lilacinus (4) | 0.06 (4) | 32 (2), 64 (2) | 0.25 (1), 1 (3) | 0.13 (2), 0.25 (2) | >16 (4) | >32 (4) |
| Rhizopus oryzae (5) | 1 (1), 4 (1), 8 (3) | 32 (1), >64 (4) | 0.016 (1), 0.13 (1), 2 (3) | 0.5 (1), 2 (1), >8 (3) | 0.13 (1), 0.25 (3), 1 (1) | 16 (1), >32 (4) |
| Rhizopus microsporus var. rhizopodiformis (2) | 4 (1), 8 (1) | >64 (2) | 1 (2) | 4 (1), 8 (1) | 1 (1), 0.5 (1) | >32 (2) |
| Mucor racemosus (1) | 8 | >64 | 1 | >8 | 0.06 | >32 |
| Scedosporium prolificans (1) | 0.03 | >64 | >4 | >4 | >16 | >4 |
| Pseudallescheria boydii (8) | 0.03 (2), 0.06 (5), 0.13 (1) | 16 (4), 32 (4) | 0.5 (6), 2 (2) | 0.13 (2), 0.25 (5), 0.5 (1) | 2 (1), 4 (1), 8 (1), 16 (1), >16 (4) | 0.25 (1), 0.5 (4), 1 (2), 2 (1) |
| Cunninghamella bertholletiae (2) | 2 (1), 4 (1) | >64 (2) | 1 (2) | >8 (2) | 2 (2) | >32 (2) |
| Cladosporium carrionii (2) | ≤0.008 (1), 0.03 (1) | 8 (1), 16 (1) | 0.016 (1), 0.03 (1) | 0.06 (2) | 0.25 (1), 2 (1) | 0.5 (1), 4 (1) |
| Cladosporium trichoides (2) | 0.016 (2) | 64 (2) | 0.03 (1), 0.13 (1) | 0.13 (1), 0.25 (1) | 0.25 (1), 1 (1) | 0.25 (1), 1 (1) |
| Exophiala dermatitidis (3) | ≤0.008 (3) | 4 (1), 8 (1), 16 (1) | 0.06 (1), 0.13 (1), 0.5 (1) | 0.03 (2), 0.06 (1) | 0.25 (1), 0.5 (2) | 1 (1), >4 (2) |
| Fonsecaea pedrosoi (3) | ≤0.008 (3) | 4 (1), 32 (2) | 0.06 (1), 0.13 (2) | 0.016 (1), 0.06 (1), 0.13 (1) | 0.13 (1), 0.5 (1), 1 (1) | 4 (2), >4 (1) |
| Phialophora verrucosa (1) | 0.016 | 64 | 0.06 | 0.25 | 1 | >4 |
| Phialophora repens (1) | 0.06 | 32 | 0.5 | 0.13 | 0.25 | >4 |
| Phialophora parasitica (1) | 0.016 | >64 | >4 | 0.25 | 1 | >4 |
| Phialophora richardsiae (2) | 0.03 (1), 0.06 (1) | 64 (2) | 0.13 (1), 0.5 (1) | 0.25 (1), 0.5 (1) | 0.5 (1), 1 (1) | 1 (1), 2 (1) |
| Rhinocladiella aquaspersa (1) | 0.06 | 64 | 1 | 0.25 | 1 | >4 |
| Dermatophytes | ||||||
| Trichophyton rubrum (3) | 8 (3) | 1 (2), 2 (1) | 0.25 (3) | 0.06 (3) | 0.25 (3) | ≤0.03 (2), >32 (1) |
| Trichophyton mentagrophytes (3) | 4 (2), 8 (1) | 0.5 (1), 4 (1), 8 (1) | 0.06 (1), 0.13 (1), 0.25 (1) | 0.016 (1), 0.13 (2) | 1 (3) | ≤0.03 (3) |
Table 3.
In vitro antifungal activities of E1210 against fluconazole-sensitive and fluconazole-resistant Candida spp.
| Fluconazole susceptibility of Candida spp. (no. of strains) | Compound | MIC (μg/ml) |
||
|---|---|---|---|---|
| Range | MIC50 | MIC90 | ||
| Susceptible (140) | E1210 | ≤0.008–4 | ≤0.008 | 0.06 |
| Fluconazole | ≤0.06–32 | 0.5 | 8 | |
| Itraconazole | 0.008–4 | 0.13 | 0.5 | |
| Voriconazole | ≤0.004–>4 | 0.016 | 0.13 | |
| Amphotericin B | 0.13–1 | 0.5 | 1 | |
| Micafungin | 0.008–1 | 0.016 | 1 | |
| Resistant (18) | E1210 | ≤0.008–>32 | 0.016 | 0.03 |
| Fluconazole | 64–>64 | >64 | >64 | |
| Itraconazole | 0.06–>4 | 2 | >4 | |
| Voriconazole | 0.03–>4 | >4 | >4 | |
| Amphotericin B | 0.25–1 | 0.5 | 0.5 | |
| Micafungin | 0.008–0.13 | 0.016 | 0.03 | |
E1210 also showed potent activities against Aspergillus spp. and Scedosporium spp., but its inhibitory effects were incomplete, as was the case with micafungin (Fig. 2). As the drug concentration increased, E1210 and micafungin induced white and small colonies attached to the bottom of the microtiter plate. The size of colonies treated with E1210 was smaller than those observed after incubation with micafungin, and the colony size became smaller as the E1210 concentration increased.
Fig. 2.

Microdilution plate used for antifungal susceptibility studies with E1210 (A) and micafungin (B) against A. fumigatus IFM51126. E1210 and micafungin were serially diluted 2-fold, from the highest concentration in column 2 (8 μg/ml) to the lowest concentration in column 12 (0.008 μg/ml). Column 1 of panel A was a drug-free growth control. The MICs of E1210 and micafungin were 0.06 μg/ml (column 9) and 0.016 μg/ml (column 11), respectively.
E1210 showed potent activity against A. fumigatus (MIC90 of 0.13 μg/ml). Its activity was 4 times higher than those of itraconazole, voriconazole, and amphotericin B and was 2 times lower than that of micafungin. Against A. flavus and Aspergillus terreus, E1210 also showed potent activities, with a MIC range of 0.016 to 0.13 μg/ml. E1210 was more active than fluconazole, voriconazole, and amphotericin B and comparable to itraconazole but less active than micafungin. E1210 showed potent activity against Aspergillus niger (MIC range of 0.016 to 0.03 μg/ml). Its activity was higher than those of the azoles tested and amphotericin B and was equal to that of micafungin. E1210 showed the greatest activity against Paecilomyces lilacinus (MICs of 0.06 μg/ml) among the compounds tested. E1210 showed the greatest activity against most Fusarium solani strains (MIC range of 0.03 to 0.25 μg/ml) among the compounds tested but showed less activity against one F. solani strain (MIC of >32 μg/ml). Regarding black molds, E1210 showed the greatest activities against Pseudallescheria boydii (MIC range of 0.03 to 0.13 μg/ml) and Scedosporium prolificans (MIC of 0.03 μg/ml) among the compounds tested. E1210 was potently active against some less common black molds, such as Cladosporium carrionii (MIC range of ≤0.008 to 0.03 μg/ml), Cladosporium trichoides (MIC of 0.016 μg/ml), Exophiala dermatitidis (MIC of ≤0.008 μg/ml), Fonsecaea pedrosoi (MIC of ≤0.008 μg/ml), and Phialophora verrucosa (MIC of 0.016 μg/ml). Regarding the Zygomycetes, E1210 had moderate activities against Rhizopus oryzae and Rhizopus microsporus var. rhizopodiformis (MIC ranges of 1 to 8 and 4 to 8 μg/ml, respectively). Its activities were greater than those of fluconazole and micafungin and comparable to those of voriconazole but lower than those of itraconazole and amphotericin B. E1210 also showed moderate activities against Mucor racemosus and Cunninghamella bertholletiae, with MICs of 8 and 2 to 4 μg/ml, respectively.
Against dermatophytes, E1210 was less active than the other antifungal compounds, with MICs of 8 μg/ml for Trichophyton rubrum and 4 to 8 μg/ml for Trichophyton mentagrophytes.
Time-growth inhibition assay.
The inhibitory effects of E1210 and fluconazole on fungal growth of C. albicans IFM49971 were evaluated comparatively (Fig. 3). E1210 and fluconazole inhibited C. albicans growth in a concentration-dependent manner, and the inhibition was fungistatic against the strain tested. E1210 at its MIC inhibited the growth of C. albicans, which was approximately 1 log10 CFU/ml lower than the growth of the control. The cell numbers of C. albicans treated with E1210 at concentrations four times the MIC (0.016 μg/ml) or greater (≥0.016 μg/ml) were approximately 2 log10 CFU/ml lower at 8 h and 3 log10 CFU/ml lower at 24 h than those of the control. The maximal fungistatic activity of E1210 was observed at eight times the MIC (0.03 μg/ml). However, with fluconazole, the maximal fungistatic activity was not observed even at 8 times its MIC (2 μg/ml).
Fig. 3.

Time-growth inhibition curves of C. albicans exposed to E1210 (A) and fluconazole (B). C. albicans IFM49971 was cultured in RPMI 1640 medium containing E1210 (0.001 to 0.06 μg/ml), fluconazole (0.03 to 2 μg/ml), or vehicle with the inoculum size of 104 CFU/ml. The inhibitory effect on fungal growth was evaluated after 2, 4, 8, and 24 h of incubation by measuring the number of CFU in duplicate.
Effects of culture conditions on E1210 MICs.
The effects of the inoculum size, the test medium, and the addition of mouse or human serum on the MICs of E1210 were evaluated (Table 4). The inoculum size (5 × 102 to 5 × 104 cells/ml) and the test medium did not greatly affect the MICs of E1210 for C. albicans, A. fumigatus, and A. flavus within a range of 1- to 4-fold the MICs. In contrast, the inoculum size affected the MICs of E1210 for F. solani and R. oryzae, which were increased 8-fold to 16-fold in an inoculum size-dependent manner.
Table 4.
Effect of culture conditions on MICs of E1210
| Organism | MIC (μg/ml) under indicated culture conditiona |
||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Inoculum size (cells/ml) |
Medium |
Addition of mouse serum (%) |
Addition of human serum (%) |
||||||||||||
| 5 × 102 | 5 × 103 | 5 × 104 | RPMI 1640 | RPMI 1640–2% glucose | SDB | AM3 | 0 | 10 | 30 | 90 | 0 | 10 | 30 | 90 | |
| C. albicans IFM49971 | 0.008 | 0.008 | 0.008 | 0.008 | 0.016 | 0.016 | 0.008 | 0.008 | 0.03 | 0.13 | 0.5 | 0.008 | 0.016 | 0.06 | 0.5 |
| A. fumigatus IFM51126 | 0.03 | 0.03 | 0.06 | 0.03 | 0.03 | 0.06 | 0.03 | 0.03 | 0.25 | 1 | 2 | 0.03 | 0.25 | 1 | 2 |
| A. flavus IFM50915 | 0.016 | 0.016 | 0.03 | 0.016 | 0.016 | 0.06 | 0.016 | 0.016 | 0.25 | 0.5 | 1 | 0.016 | 0.25 | 0.5 | 1 |
| F. solani IFM50956 | 0.03 | 0.06 | 0.25 | 0.06 | NT | NT | NT | 0.06 | NT | NT | NT | 0.06 | NT | NT | NT |
| R. oryzae IFO5442 | 0.25 | 0.5 | 2 | 0.5 | NT | NT | NT | 0.5 | NT | NT | NT | 0.5 | NT | NT | NT |
The results are the median MICs of E1210 from three independent experiments. For determining the effects of the test medium and the addition of serum, the inoculum sizes were 1 × 103 cells/ml for C. albicans and 1 × 104 cells/ml for the filamentous fungi. AM3, antibiotic medium 3; NT, not tested.
The addition of serum (10 to 90%) greatly affected the MICs of E1210 for the one Candida and two Aspergillus strains tested. The MICs of E1210 increased 64-fold in a serum concentration-dependent manner.
Cytotoxicity assay.
The cytotoxicity of E1210 was compared with those of fluconazole and amphotericin B using human HK-2 cells. The cytotoxicities of both E1210 and fluconazole were low (IC50s of >32 μg/ml) and were significantly lower than that of amphotericin B (IC50 of 3.7 μg/ml). As shown in Table 4, E1210 showed potent antifungal activities against C. albicans (MICs of 0.03 to 0.016 μg/ml), A. fumigatus (MIC of 0.25 μg/ml), and A. flavus (MIC of 0.25 μg/ml) in the presence of 10% serum, and its activity was >128-fold higher than its toxicity.
DISCUSSION
In this study, we examined the antifungal activities of E1210 against clinical fungal isolates compared to those of control antifungal agents and showed the unique antifungal profile of E1210. First, E1210 exhibited potent broad-spectrum antifungal activities against clinically important fungi, including Candida spp. and Aspergillus spp. Of note, E1210 inhibited P. boydii, S. prolificans, and P. lilacinus with lower MICs than the other compounds tested and also showed low MICs for F. solani and some black molds.
E1210 showed potent activities against most Candida spp., such as C. albicans, C. glabrata, C. tropicalis, and C. parapsilosis. As one of the notable features of E1210, it had equally potent activities against fluconazole-resistant and fluconazole-susceptible Candida strains, and it showed no cross-resistance with azoles in Candida spp. However, the antifungal activity of E1210 against C. krusei was lower than those of the other compounds tested. This might result from a structural difference in the E1210 binding site of the GWT1 protein of C. krusei compared to that of E1210-susceptible Candida spp., although its precise mechanism remains unknown.
We also found that E1210 had potent activities against Aspergillus spp., including A. fumigatus, A. flavus, A. niger, and A. terreus. The activities of E1210 were greater than those of the azoles tested and amphotericin B and were almost equal to those of micafungin. In particular, A. terreus was reported to have in vitro and in vivo resistance to amphotericin B, as evidenced by a diminished therapeutic response to amphotericin B treatment for infections due to A. terreus species with elevated MICs to amphotericin B (10). Further investigation using strains with greater levels of resistance to amphotericin B is needed to determine the potential for E1210 to treat infections due to such A. terreus strains. In addition, further investigation regarding its in vivo efficacy will be needed. At concentrations higher than the MIC, E1210 induced white and small colonies attached to the bottom of the microtiter plate, as was the case with micafungin. This phenotype of Aspergillus has been previously reported for the echinocandin derivatives (14, 24). Rennie et al. reported that electron microscopic examination of these colonies induced by anidulafungin revealed that they appeared to be cell wall-deficient microcolonies (19). These colonies became smaller in size as the E1210 concentration increased, but micafungin did not decrease colony size even at higher concentrations than its MIC. This might be due to the difference in their mechanisms of action, i.e., GPI synthesis inhibition versus 1,3-β-d-glucan synthesis inhibition (8). GPI proteins might play a more important role in cell wall assembly than 1,3-β-d-glucan in molds. In the latest published CLSI M38-A2 document, the minimum effective concentration (MEC) is defined as the endpoints for measuring echinocandin activity against molds (3). The MEC is the lowest concentration of drug that leads to the growth of small, rounded, and compact hyphal forms and is equivalent to the MIC defined in this study. This terminology is currently used only with respect to testing of the echinocandins. In this study, we determined the MIC of E1210 by the MEC endpoint, because E1210 induced a morphological change similar to that of micafungin in some molds. The endpoint determination for E1210 activity against molds needs to be further discussed.
Scedosporium spp., R. oryzae, F. solani, and P. lilacinus are emerging filamentous pathogens that cause severe invasive infections in immunocompromised patients (4, 13, 17, 22). The optimal treatment for these infections has not yet been definitively established because of the low susceptibilities to conventional antifungal drugs. E1210 showed the highest activities against P. boydii, S. prolificans, and P. lilacinus among the compounds tested and potent activities against some black molds. Of note, S. prolificans was resistant to all other antifungal compounds tested, including amphotericin B. E1210 was also active against F. solani strains, which were resistant to the azoles and echinocandins tested. In this study, the number of strains tested was limited, so further studies with more strains are needed to confirm the activity of E1210 against rare fungi. The MIC of E1210 for F. solani was significantly altered by the inoculum size. Slight variations in inoculum size might cause great variations in E1210's MICs in the seven F. solani isolates tested. For R. oryzae, the MIC of E1210 was also greatly affected by the inoculum size. The in vivo efficacies of E1210 against infections caused by these strains should be evaluated to clarify the optimal conditions for the determination of MIC endpoints.
GPI anchoring is a highly conserved mechanism within eukaryotes, including pathogenic fungi, but E1210 showed fungus-specific inhibition of GWT1 protein function (28). Consistent with the finding, E1210 showed low cytotoxicity in human cells tested. Although this toxicity study was preliminary, using a single cell line, its results demonstrated that E1210 is not generally toxic for eukaryotic cells and suggested that the therapeutic index of the compound might well be at least that of the relatively nontoxic azole drugs.
In conclusion, E1210 is a promising novel antifungal agent which has potent broad-spectrum antifungal activity against clinically important fungi, including Candida spp., Aspergillus spp., and non-Aspergillus filamentous fungi. Further studies of its in vivo efficacy, its pharmacokinetic profile, and its toxicologic safety are warranted.
ACKNOWLEDGMENTS
We thank the Medical Mycology Research Center, Chiba University (Chiba, Japan), Graduate School of Medicine, Gifu University (Gifu, Japan), and the Institute for Fermentation, Osaka (Osaka, Japan), for providing the fungal strains. We thank Takashi Owa, Eisai Product Creation Systems, Eisai, Inc., NJ, for making constructive suggestions to the antifungal project team. We thank Frederick P. Duncanson, Eisai Product Creation Systems, Eisai, Inc., NJ, for editing and proofreading this article.
Footnotes
Published ahead of print on 8 August 2011.
REFERENCES
- 1. Bates D. W., et al. 2001. Mortality and costs of acute renal failure associated with amphotericin B therapy. Clin. Infect. Dis. 32:686–693 [DOI] [PubMed] [Google Scholar]
- 2. Clinical and Laboratory Standards Institute 2008. Reference method for broth dilution antifungal susceptibility testing of yeasts; approved standard. CLSI document M27-A3. CLSI, Wayne, PA [Google Scholar]
- 3. Clinical and Laboratory Standards Institute 2008. Reference method for broth dilution antifungal susceptibility testing of conidium-forming filamentous fungi: approved standard, 2nd ed CLSI document M38-A2. CLSI, Wayne, PA [Google Scholar]
- 4. Cortez K. J., et al. 2008. Infections caused by Scedosporium spp. Clin. Microbiol. Rev. 2008 21:157–197 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Denning D. W. 2003. Echinocandin antifungal drugs. Lancet 362:1142–1151 [DOI] [PubMed] [Google Scholar]
- 6. Enoch D. A., Ludlam H. A., Brown N. M. 2006. Invasive fungal infections: a review of epidemiology and management options. J. Med. Microbiol. 55:809–818 [DOI] [PubMed] [Google Scholar]
- 7. Hata K., Horii T., Miyazaki M., Watanabe N. 2010. Abstr. 50th Intersci. Conf. Antimicrob. Agents Chemother., abstr. F1-842 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Hatano K., Morishita Y., Nakai T., Ikeda F. 2002. Antifungal mechanism of FK463 against Candida albicans and Aspergillus fumigatus. J. Antibiot. (Tokyo) 55:219–222 [DOI] [PubMed] [Google Scholar]
- 9. Horii T., Okubo M., Miyazaki M., Hata K., Watanabe N. 2010. Abstr. 50th Intersci. Conf. Antimicrob. Agents Chemother., abstr. F1-843 [Google Scholar]
- 10. Lass-Flörl C., et al. 2005. Epidemiology and outcome of infections due to Aspergillus terreus: 10-year single centre experience. Br. J. Haematol. 131:201–207 [DOI] [PubMed] [Google Scholar]
- 11. Nakamoto K., et al. 2010. Synthesis and evaluation of novel antifungal agents—quinoline and pyridine amide derivatives. Bioorg. Med. Chem. Lett. 20:4624–4626 [DOI] [PubMed] [Google Scholar]
- 12. Neofytos D., et al. 2009. Epidemiology and outcome of invasive fungal infection in adult hematopoietic stem cell transplant recipients: analysis of multicenter Prospective Antifungal Therapy (PATH) Alliance registry. Clin. Infect. Dis. 48:265–273 [DOI] [PubMed] [Google Scholar]
- 13. Nucci M., Anaissie E. 2007. Fusarium infections in immunocompromised patients. Clin. Microbiol. Rev. 20:695–704 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Oakley K. L., Moore C. B., Denning D. W. 1998. In vitro activity of the echinocandin antifungal agent LY303,366 in comparison with itraconazole and amphotericin B against Aspergillus spp. Antimicrob. Agents Chemother. 42:2726–2730 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Okubo M., Toritsuka N., Horii T., Hata K., Sonoda J. 2010. Abstr. 50th Intersci. Conf. Antimicrob. Agents Chemother., abstr. F1-844 [Google Scholar]
- 16. Ostrosky-Zeichner L., et al. 2003. Antifungal susceptibility survey of 2,000 bloodstream Candida isolates in the United States. Antimicrob. Agents Chemother. 47:3149–3154 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Pastor F. J., Guarro J. 2006. Clinical manifestations, treatment and outcome of Paecilomyces lilacinus infections. Clin. Microbiol. Infect. 12:948–960 [DOI] [PubMed] [Google Scholar]
- 18. Pfaller M. A., Diekema D. J. 2010. Epidemiology of invasive mycoses in North America. Crit. Rev. Microbiol. 36:1–53 [DOI] [PubMed] [Google Scholar]
- 19. Rennie R., Sand C., Sherburne R. 1997. Electron microscopic evidence of the effect of LY303366 on Aspergillus fumigatus. Abstr. 13th Congr. Int. Soc. Hum. Animal Mycol., abstr. P451 [Google Scholar]
- 20. Richard M. L., Plaine A. 2007. Comprehensive analysis of glycosylphosphatidylinositol-anchored proteins in Candida albicans. Eukaryot. Cell 6:119–133 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Rincón S., Cepero de García M. C., Espinel-Ingroff A. 2006. A modified Christensen's urea and CLSI broth microdilution method for testing susceptibilities of six Malassezia species to voriconazole, itraconazole, and ketoconazole. J. Clin. Microbiol. 44:3429–3431 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Roden M. M., et al. 2005. Epidemiology and outcome of zygomycosis: a review of 929 reported cases. Clin. Infect. Dis. 41:634–653 [DOI] [PubMed] [Google Scholar]
- 23. Tanaka K., et al. 2010. An effective synthesis of a (pyridine-3-yl)isoxazole via 1,3-dipolar cycloaddition using ZnCl2: synthesis of a (2-aminopyridin-3-yl)isoxazole derivative and its antifungal activity. Chem. Lett. 39:1033–1035 [Google Scholar]
- 24. Tawara S., et al. 2000. In vitro activities of a new lipopeptide antifungal agent, FK463, against a variety of clinically important fungi. Antimicrob. Agents Chemother. 44:57–62 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Tsukahara K., et al. 2003. Medicinal genetics approach towards identifying the molecular target of a novel inhibitor of fungal cell wall assembly. Mol. Microbiol. 48:1029–1042 [DOI] [PubMed] [Google Scholar]
- 26. Umemura M., et al. 2003. GWT1 gene is required for inositol acylation of glycosylphosphatidylinositol anchors in yeast. J. Biol. Chem. 278:23639–23647 [DOI] [PubMed] [Google Scholar]
- 27. Velegraki A., Alexopoulos E. C., Kritikou S., Gaitanis G. 2004. Use of fatty acid RPMI 1640 media for testing susceptibilities of eight Malassezia species to the new triazole posaconazole and six established antifungal agents by a modified NCCLS M27-A2 microdilution method and Etest. J. Clin. Microbiol. 42:3589–3593 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Watanabe N., Miyazaki M., Horii T., Hata K. 2010. Abstr. 50th Intersci. Conf. Antimicrob. Agents Chemother., abstr. F1-841 [DOI] [PMC free article] [PubMed] [Google Scholar]