

Abstract
Phosducin (Pd), a small protein found abundantly in photoreceptors, is widely assumed to regulate light sensitivity in the rod outer segment through interaction with the heterotrimeric G protein transducin. But, based on histochemistry and Western blot analysis, Pd is found almost entirely in the inner segment in both light and dark, most abundantly near the rod synapse. We report a second small protein, 14-3-3, in the rod with a similar distribution. By immunoprecipitation, phospho-Pd is found to interact with 14-3-3 in material from dark-adapted retina, and this interaction is markedly diminished by light, which dephosphorylates Pd. Conversely, unphosphorylated Pd binds to inner segment G protein(s) in the light. From these results and reported functions of 14-3-3, we have constructed a hypothesis for the regulation of light sensitivity at the level of rod synapse. By dissociating the Pd/14-3-3 complex, light enables both proteins to function in this role.
The vertebrate rod is evolution's quintessential example of an optimized visual cell. The rod functions as a low noise, single-photon detector that can adjust sensitivity over 3 log units. The molecular mechanism of rod excitation is well studied (1–9). Photoisomerization of rhodopsin (Rho) in the rod outer segment (ROS) produces conformational changes that favor the binding of the heterotrimeric G protein (transducin, Gt). The resulting exchange of GTP for GDP promotes the dissociation of α and βγ subunits. Gtα/GTP can rapidly activate the phosphodiesterase that hydrolyzes cGMP, thus causing cGMP-dependent cation channels to close (10). The resulting membrane hyperpolarization manifests at the rod inner segment (RIS) synapse by preventing release of the inhibitory neurotransmitter glutamate. This interruption in glutamate release disinhibits the bipolar cell, which, together with other retinal neurons, communicates the photon signal to the optic nerve. Meanwhile, the rod recovers through a set of restorative metabolic activities that reopen the cation channels. What is missing from this scenario is a mechanism to more completely explain the light regulation of rod sensitivity: as ambient light increases more light is required to inhibit the “dark” current. There is experimental evidence that the rod itself contributes a major term in the equation of light adaptation (11–13). Several likely molecular mechanisms have been identified, most linked to the 10-fold decrease in Ca2+ level that follows ROS cation channel closure (12, 14). For example, both activation of Rho kinase by recoverin and activation of guanylyl cyclase depend on light-induced falls in ROS [Ca2+] (15–18). However, these explanations and other Ca2+-regulated ROS pathways cannot fully account for light adaptation because of insufficient magnitude and/or mismatched kinetics.
Enter phosducin (Pd), a monomeric 28-kDa phospho-protein, found abundantly in rods, cones, and Rho-expressing pineal cells (19, 20). The putative expression of trace amounts of Pd in other parts of the nervous system, and indeed in non-neuronal cell types remains controversial (21–23). First isolated as a phosphoprotein from dark-adapted rat retinal extracts (24), Pd was found to gradually dephosphorylate on light exposure. Both phosphorylated and unmodified forms of Pd show high affinity binding to G protein βγ subunits (25, 26), but the ability of Pd to compete with or displace Gtα is markedly diminished by phosphorylation at Ser-73 (25, 26). Analyses of structures and binding energies of the two forms of Pd show that phosphorylation perturbs that part of the Pd binding domain that can induce a conformational change in Gtβγ (27, 28). The phosphorylation state of Pd can be regulated by Ca2+ through adenylyl cyclase and protein kinase A (PKA) (25, 26, 29). Hence, the current paradigm for Pd function:photon capture leads to a fall in Ca2+ that deactivates PKA, allowing Pd to dephosphorylate and sequester Gtβγ, preventing reassembly of the trimeric Gt that activated Rho needs to propagate its signal (29).
The perception that Pd functions within the ROS is based largely on the conviction that Pd interacts in vivo with Gt. That the yield of Pd from purified ROS is poor has been explained by the idea that Pd moves to the inner segment (IS) after light exposure and only assumes an outer segment location after dark adaptation. Alternatively, the ROS becomes leaky during preparation and rapidly loses its Pd. However, the assumption that Pd is primarily situated and functions within the ROS was undermined by the data of Lee and coworkers (30) whose histochemical analyses demonstrate an abundance of Pd in the RIS with especially heavy staining in the region of the synapse. Kuo and others (19, 30, 31) also conclude that there is abundant Pd in the RIS. In this report we provide information supporting a primary role for Pd in the RIS. We have quantified the distribution of Pd between the IS (83%) and outer segments (17%) as well as the ratio of Pd to Gt in the outer segment (1 to 60). This is simply too low for the Pd role suggested by the Gt sequestration model.
As a component of the IS emphasis, we introduce 14-3-3 in a previously unrecognized role, as a protein that copurifies and interacts with Pd. The presence of 14-3-3 is demonstrated in rods, where it is confined to the IS. This phosphoprotein with a molecular mass of about 29 kDa occurs in seven highly conserved vertebrate isoforms of ancient lineage. In contrast with Pd, it is expressed in many different cells and tissues, and it is especially prevalent in neurons. 14-3-3 is known to participate in different signaling pathways, including those that use either Raf kinases or heterotrimeric G proteins. It also has been associated with synaptic structures and has been implicated in the regulation of exocytosis. Its biochemical properties and physiological interactions are summarized in a number of reviews (32, 33).
Finally, we demonstrate that light disrupts the complex formed between Pd and 14-3-3, an interaction that must be occurring within the RIS. When taken together, these findings are consistent with the hypothesis that Pd and 14-3-3, in concert with other as-yet-unidentified IS proteins, provide a major pathway for the light regulation of rod sensitivity.
Materials and Methods
Copurification of Pd and 14-3-3.
Bovine Pd and its companion protein 14-3-3 were purified from fresh, dark-adapted bovine retinas (W.L. Lawson, Lincoln, NE) under IR illumination, essentially by the method of Yoshida et al. (26). In the final stage of purification, Pd was chromatographed on Poros Q/M equilibrated with 50 mM Tris (pH 8.0), containing 0.1 mM PMSF and 1 mM DTT. Pd was eluted by a linear Tris gradient (50–750 mM). Aliquots of fractions in loading buffer were incubated at 80°C (10 min) and analyzed on SDS/PAGE (NOVEX 4–12% Bis⋅Tris gels with Mes running buffer).
MS.
Gel bands containing Coomassie-stained proteins were excised, minced, thrice destained, and vacuum dried. Gel fragments were rehydrated with freshly prepared sequencing grade trypsin (Sigma) (0.025 mg/ml in 25 mM NH4HCO3). Proteins were digested for 20 h at 37°C, and the gel fragments were sedimented at 12,000 × g. Tryptic peptides were twice more extracted from the gel fragments with 2% trifluoroacetic acid (TFA) in 80% aqueous acetonitrile, combined with the earlier supernatants, and dried in a Speed Vac. Samples were redissolved in aqueous TFA, desalted with C-18 peptide tips (Millipore), and analyzed by matrix-assisted laser desorption ionization–time of flight (MALDI-TOF)-MS using α-cyano-4-hydroxycinnamic acid as the matrix (Voyager DE, Applied Biosystems) (34). The protein database was searched through the ExPASy web site (www.expasy.ch/tools) using the peptident mass matching program.
Cloning, Expression, and Antibody Production.
A full-length cDNA fragment of bovine 14-3-3ɛ from a bovine retinal library (Stratagene) was amplified by PCR, using pfuTurbo DNA polymerase (Stratagene). After amplification and sequence verification, DNA was ligated into plasmid pET15b (Novagen), which had been cut with NdeI–XhoI. Full-length rat Pd cDNA subcloned in pET15b was a gift from Barry M. Willardson, Brigham Young University, Provo, UT. Proteins were expressed in Escherichia coli BL21 (DE3). When cells had reached an OD of 0.6 at 600 nm, expression was induced with isopropyl β-d-thiogalactoside, and the cells were incubated for an additional 4 h. Cells then were collected, resuspended in 0.1 volume of 20 mM Tris⋅HCl (pH 7.9), 0.5 M NaCl, 5 mM imidazole, and 1 mM PMSF and sonicated five times at ice temperature for 30 sec with 2-min chilling intervals. Insoluble material was removed by centrifugation, and the proteins were purified on an Ni-NTA column (Qiagen, Chatsworth, CA). Proteins were eluted with 20 mM Tris⋅HCl, 0.5 M NaCl, 1 M imidazole, and 1 mM PMSF buffer (pH 7.9), dialyzed against PBS + protease inhibitor mixture (general use, Sigma), and concentrated with a Centriprep 10 (Millpore). Polyclonal antibodies to Pd and 14-3-3 were prepared and affinity-purified with epitope-conjugated CNBr Sepharose by Strategic Biosolutions, Ramona, CA. Antibodies against conserved and type-specific peptides from Gβ1, Gβ2, and Gβ3 were purchased from Santa Cruz Biotechnology; they are reactive to human, mouse, and rat proteins.
Immunohistochemistry.
Long Evans rats (Charles River Breeding Laboratories) maintained in 12-h day/night cycles were light- or dark-adapted for more than 2 h and euthanized by cervical dislocation under CO2 anesthesia. The eye was enucleated and 0.5 ml of 4% formaldehyde was injected into the space between the lens and retina; fixation was continued for 5 min. The cornea and lens were removed with a scalpel, and the remaining structures were immersed in fixative for 30–60 min. The eye then was debrided of extraocular tissue, embedded in OCT compound (Tissuetek, Torrance, CA), and frozen: 14-μm sections were prepared with a cryostat and mounted on glass slides. Sections were washed with 0.1 M sodium phosphate (pH 7.4), blocked for 1 h, then incubated overnight at 4°C with primary antibody. After washing, goat anti-rabbit antibody labeled with fluorescein (Molecular Probes) was applied for 2 h, and the slides were washed, covered with glycerol, and photographed in a confocal microscope (Olympus). FITC fluorescence was analyzed with National Institutes of Health image software.
ROS Preparation and Quantification of Pd and Gtα.
Four freshly dissected rat retinas were transferred to 500 μl of ice-cold Hepes/Ringer's buffer (35). ROS were detached in 3-sec bursts of vortexing (×15) and allowed to settle for 3 min. The crude ROS supernatant was collected (repeated twice), layered on top of a 30/40% (wt/vol) discontinuous sucrose gradient in Hepes/Ringer's buffer, and centrifuged for 45 min at 150,000 × g. The interface was collected, diluted 2-fold in buffer, and pelleted at 3,000 × g for 20 min.
ROS integrity was assessed by examining the entry of [γ-33P]ATP (NEN) with or without disruption of ROS with a 22-gauge needle (×50). Phosphorylation of Rho by Rho kinase was induced by exposing ROS to ambient light and temperature for 10 min. The reaction was terminated with SDS/PAGE loading buffer, and proteins were separated by 4–12% SDS/PAGE. Radioactivity was measured by scanning a phospho-image plate with the Storm System (Molecular Dynamics) and quantifying bands by imagequant (Molecular Dynamics). Pd and Gtα concentrations in ROS were determined by quantitative Western blot analysis (36).
Immunoprecipitation.
Retinas were solubilized by 5 sec of sonication (×2) in 50 mM Tris⋅HCl (pH 8.00), 150 mM NaCl, 1% NP-40, 0.1 mM PMSF, protease inhibitor mixture, phosphatase inhibitor mixture I, H89 (Sigma) at 4°C. Lysates were incubated with 20 μl of protein A agarose (Roche Molecular Biochemicals) pre-equilibrated in this buffer for 1 h at 4°C to remove nonspecific binding proteins. Supernatants were divided into 150-μl aliquots and incubated with 5 μg of antibody and 50 μl of protein A agarose for 1 h at 4°C, and centrifuged for 20 sec at 10,000 × g, and the supernatants were removed. Beads were washed for 20 min five times with 1 ml of solubilizing buffer. Immune complexes were boiled with SDS/PAGE loading buffer and analyzed on 4–12% SDS/PAGE. Gels were transferred to poly(vinylidene difluoride) membranes (NEN) for Western blot analysis. Membranes were developed with ECL (Amersham Pharmacia) and imaged with Kodak XB-1 film.
Results
Copurification of Pd and 14-3-3.
Despite the many steps of purification (26) leading to a major fraction containing ≈85% Pd (Fig. 1A, lane 2), the trailing shoulder of this peak contained a companion band of similar size on SDS/PAGE (Fig. 1A, lane 3). To identify this protein, it was in-gel digested with trypsin and analyzed by MALDI-TOF-MS. A profile search of the Swiss-Prot database using the peptident program (ExPASy) showed that Pd's copurifier was part of the highly conserved 14-3-3 protein family, perhaps the ζ isoform (Table 1). A second attempt gave similar results.
Figure 1.

Pd and 14-3-3 were copurifed from bovine retinas. (A) Eluted proteins from the Poros Q/M 4.6/100 column were resolved on 4–12% SDS/PAGE and stained with Coomassie blue. Lane 1, molecular mass standard; lane 2, Pd peak; lane 3, trailing shoulder. Bands A, B, and C and background gel were digested with trypsin, and tryptic peptides from bands A, B, and C were characterized by MALDI-TOF-MS. Band A was identified as Pd, B, as a 14-3-3 protein, and C as a mixture of Pd and 14-3-3. (B) Antibody identifications. Lanes 1 and 2 were probed with Pd Ab; lanes 3 and 4 were probed with 14-3-3 Ab. Lanes 1 and 3 contained 10 μg rat retina protein; lanes 2 and 4 contained 10 μg rat brain protein.
Table 1.
Identification by MALDI-TOF-MS of 14-3-3 isolated from rat retina
| Peptide mass (Da), experimental | In best match (human/bovine ζ)
|
||
|---|---|---|---|
| Theoretical | Sequence | Position | |
| 615.2 | 615.36 | NVVGAR | 50–55 |
| 629.2 | |||
| 657.1 | |||
| 673.1 | 673.29 | GDYYR | 123–127 |
| 684.1 | |||
| 747.3 | |||
| 760.2 | 760.42 | IETELR | 86–91 |
| 785.1 | |||
| 816.3 | 816.42 | LAEQAER | 12–18 |
| 907.4 | 907.52 | NLLSVAYK | 42–49 |
| 1080.4 | |||
| 1108.2 | 1108.56 | EMQPTHPIR | 159–167 |
| 1124.3 | |||
| 1205.3 | |||
| 1256.4 | |||
A tryptic digest of the PAGE isolated protein copurifying with Pd was analyzed by MALDI-TOF-MS (Voyager-DE) using alpha-cyano-4-hydroxycinnamic acid as the matrix and external calibration. All the prominent masses above 500 were used to search the Swiss-Prot database using peptident at the ExPASy web site. Matches were restricted to proteins of 26.4–39.6 kDa, with a peptide mass tolerance of +/− 1 Da. The best match showed six hits and several showed five; all were isoforms of 14-3-3. Although form ζ was a slightly better match in this first experiment, later runs and manual analysis gave a slight edge to η. In any case, almost all the matched peptides were from conservative regions where most 14-3-3 isoforms were identical.
Antibodies Against Rat Pd and 14-3-3.
We chose the rat to achieve total dark adaptation as well as the freshness required for histochemical analyses. Polyclonal antibodies against recombinant Pd (rat) and 14-3-3ɛ (bovine) were raised in rabbit. Specificity of Pd antibody was examined by Western blot (shown in Fig. 1B). No cross-reactivity was observed with whole rat brain extract (Fig. 1B, lane 2). Furthermore, the Pd antibody had similar reactivity toward nonphosphorylated and Pd phosphorylated by protein kinase A (not shown). 14-3-3 antibody identified a single band of 29 kDa in both retina and brain extracts from rat (Fig. 1B).
Immunocytochemical Localization of Pd and 14-3-3 in Rat Retina.
To characterize the abundance and subcelluar localization of Pd, immunocytochemical samples were prepared from light- and dark-adapted rat retinas (Fig. 2 A and B). Adjacent to the photomicrograph, we show integrated fluorescence intensity profiles for the section of retina shown. The amount of Pd calculated from the integrated intensities was 83 ± 6% for RIS and 17 ± 6% for ROS (n = 8). Comparison of light and dark sections gave no evidence of IS/outer segment translocations, although, in the dark, Pd is slightly more intense at the synapse itself. These observations are not consistent with light-induced Pd translocations between ROS and RIS (31, 37).
Figure 2.

Immunohistological localization of Pd and 14-3-3 in rat retina. Retina sections were incubated with polyclonal Pd Ab (A and B) and 14-3-3ɛ Ab (C) separately, followed with fluorescein-labeled secondary antibody and analysis by fluorescent confocal microscopy. (A) Pd in light-adapted retina. (B) Pd in dark-adapted retina. (C) 14-3-3 in light-adapted retina. Integrated FITC intensity profiles are shown at the right. OS, outer segment; ONL, outer nuclear layer; INL, inner nuclear layer; IPL, inner plexiform layer; GCL, ganglion cell layer.
14-3-3 stained heavily in the photoreceptor IS, outer plexiform layer (OPL), inner plexiform layer (IPL), and ganglion cell layer (GCL) (Fig. 2C). In rods, the 14-3-3 data support an exclusive RIS localization with ROS staining at background levels. Thus, Pd and 14-3-3 colocalize in the RIS.
Quantification of Pd in ROS and RIS.
ROS were isolated from dark-adapted rat retina. The leakiness of rat ROS membrane was estimated by penetration of [γ-33P]ATP and subsequent labeling of Rho, which was 15 ± 4% (relative to “disrupted” ROS) in four trials. Abundance of Gtα and Pd in intact ROS was quantified by Western blotting (Fig. 3). Gtα and Pd were used as the internal reference protein (Fig. 3 A and B, respectively), and the amount of each protein was estimated with calibration curves obtained with recombinant Pd or purified bovine Gtα. This procedure gave a relative abundance for Pd of 0.052 ± 0.015 pmol/μg and for Gtα of 3.12 ± 0.98 pmol/μg of total ROS protein (n = 4). The two measurements give a ratio of ≈1 Pd for every 60 Gtα in rat ROS.
Figure 3.

Quantification of Pd and Gtα in ROS using Western blot analysis. (A) Lanes 1–4 are 0.5, 1, 2, and 4 μg of ROS protein. Lanes 5–8 are 0.5, 1, 2, and 5 pmol Gtα. (B) Lanes 1–4 are 0.2, 0.5, 1, and 2 μg of ROS protein. Lanes 5–9 are recombinant Pd, 0.05, 0.1, 0.2, 0.4, and 0.8 pmol. Blots were probed with Pd and Gtα antibodies. Concentrations of Pd and Gtα in ROS were calculated by comparing the slopes of the linear regions of the ROS chemilluminescence curves with similar data with pure proteins.
Light-Dependent Interaction of Pd and 14-3-3: Immunoprecipitation.
Pd is phosphorylated in dark-adapted retina and dephosphorylated in light-exposed retina (24). Furthermore, 14-3-3 interacts with Pd in vitro only when Pd is phosphorylated.† Because Pd and 14-3-3 colocalize in RIS by immunohistochemistry, the possibility of direct interaction in vivo was examined with immunoprecipitation. Dark- or light-adapted retinal extracts were mixed with Pd antibody bound to protein A agarose, and captured antibody/antigen complexes were eluted from protein A agarose and probed with 14-3-3 antibody by Western blotting (Fig. 4A, lanes 1 and 2). A significantly larger quantity (≈five times) of 14-3-3 formed a complex with Pd and was captured by Pd antibody in dark-exposed (Fig. 4A, lane 2) than in light-exposed retina (Fig. 4A, lane 1). Similar results were obtained when Pd/14-3-3 complexes were captured by 14-3-3 antibody bound to protein A agarose (Fig. 4A, lanes 4 and 3). These data support previous observations that Pd dephosphorylates in light-exposed retinal extracts and that the phosphorylated form of Pd binds to 14-3-3 with much higher avidity.
Figure 4.

Coprecipitation of Pd/14-3-3 (A) and Pd/Gβ (B). Dark- and light-adapted Long-Evans rat retinas were solubilized and mixed with rat Pd or bovine 14-3-3ɛ polyclonal antibodies. Immunocomplexes were precipitated with Protein A agarose. Western blots were probed with bovine 14-3-3ɛ, rat Pd or Gβ peptide polyclonal Ab. Retinal extracts were mixed with Protein A agarose-Pd Ab (A1, A2, B1, B2) or Protein A agarose-14-3-3ɛ Ab (A3, A4). Captured complexes were electrophoresed and probed by Western blot analysis. Light-adapted (A1, A3, B1) and dark-adapted (A2, A4, B2) extracts were analyzed with 14-3-3 Ab (A1, A2), Pd Ab (A3, A4), or Gβ Ab (B1, B2). IP, Immunoprecipitation; WB, Western blot.
Reciprocal Effects of Light on the Interaction of Pd with Gβγ and 14-3-3.
Gβγ is known to form a tight complex with dephosphorylated Pd (26, 38). We next examined light-dependent formation of Gtβγ/Pd complexes by immunoprecipitation. We exposed anti-Pd/protein A agarose to extracts prepared from both dark- and light-adapted rat retinas (Fig. 4B). When the captured complex was probed with anti-Gβ (not type-specific), Gβ was found in complex with Pd in light-adapted retina (Fig. 4B, lane 1). When a similar experiment was performed in dark-adapted retinal extracts, the amount of captured Gβ was markedly decreased (Fig. 4B, lane 2). These data show opposite effects of light on Gβγ/Pd binding compared with those found for 14-3-3/Pd binding.
Immunohistological Localization of Gβ1, Gβ2, and Gβ3 in Rat Retina.
Sections from light-adapted rat retina were probed with antibodies specific for Gβ1, Gβ2, and Gβ3. Sections were visualized with a FITC-conjugated secondary antibody (Fig. 5). In confirmation of previous data (39), the Gβ1 subtype (which includes Gt) was found virtually exclusively in ROS (Fig. 5A). Gβ2 was localized to RIS and the rest of the retina. Gβ3 was localized in cone outer segments as reported (39) as well as in OPL (which includes the axon of the RIS) and the inner retina.
Figure 5.
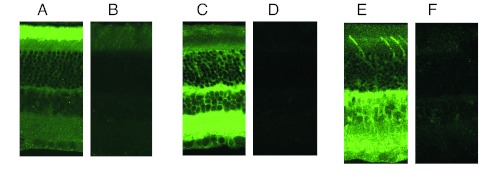
Immunohistochemical localization of Gβ subtypes in rat retina. (A) FITC localization of Gβ1 heavily stains the outer segment of photoreceptors, with weak staining elsewhere. (C) Gβ2 stains heavily in the outer plexiform layer and inner retina. (E) Gβ3 localizes in the cone outer segment, outer plexiform layer, and selectively stained bipolar cells, amacrine cells and ganglion cell bodies. Specificity of staining was demonstrated by preincubating primary Ab with 10-fold excess of blocking peptides (shown in B, D, and F).
Discussion
Subcellular Distribution of Rod Proteins.
Results from this and other laboratories (19, 30) support the conclusion that Pd is primarily an IS protein.
First, Pd is seldom purified from ROS because the yields are miniscule (26, 39). The explanation that Pd can escape from leaky ROS is not compelling because labeled ATP, a smaller molecule with higher diffusivity, can access only about 15% of the Rho molecules in freshly prepared dark-adapted rat ROS. By Western blot analysis the ratio of Pd to Gtα is 1:60. This is a maximal value because of the inevitable contamination of ROS by RIS. The true ratio may be much less.
Second, a comparison of ROS Pd with that from RIS by a quantitative analysis of fluorescence intensities reveals that 83% of Pd is localized to the IS with heavy staining in the photoreceptor synapse. The remaining 17% is scattered throughout the ROS. This is again a maximum, because self-quenching at the intensely stained synapse undoubtedly leads to underestimation of synaptic Pd.
Third, the idea that light induces Pd translocation between RIS and ROS (31, 37) is not supported by our data. In our experiments little, if any, induced movement of Pd was found. There are no data that support rapid, reversible Pd movement between RIS and ROS within the time intervals required for Pd to be a significant light-dependent modulator of ROS Gt concentrations. The differences among these studies may reflect different antisera, different animal species, and different times of day and formats by which light is introduced. Adding to the complexity is the fact that Pd changes its phosphorylation state as well as its protein interactions as a function of illumination. Thus for these measurements an antibody must be selected that binds equally to phosphorylated and unmodified Pd, as well as one that can identify Pd whether or not it is bound to Gβγ or 14-3-3. We have verified our polyclonal antibody in these ways, and we find no light-induced movement.
Fourth, when Pd phosphorylation was first shown to be light dependent (24), whole rat retina was the source, not ROS. When Shuster and Farber (11) examined the effect of cyclic nucleotides on protein phosphorylation in purified, dark-adapted ROS, they found no sign of cAMP-dependent phosphorylation of Pd-sized protein. On the other hand, they did report such phosphorylation in retinal preparations that contained RIS.
Fifth, not only is Pd far more abundant in RIS, its highest concentration is found at the rod synapse. Although not excluding other Pd roles (19, 31), this striking synaptic concentration indicates a Pd function at this site.
Pd in ROS.
Although our data support an IS function for Pd, they do not preclude an outer segment function. However, the 1:60 ratio of Pd to Gt does appear to exclude a stoichiometric modulation of Gt activity by Pd. On the other hand, any putative activity of Pd would be constrained by the large excess of Gt. And there is another way to account for the small amount of Pd in ROS: because 1/12th of ROS Gt is replenished daily, perhaps 1 in 60 of these molecules fortuitously traps Pd into the ROS.
Regulation of Pd Phosphorylation.
We have not yet identified the IS kinases and phosphatases responsible for regulating the light-dependent interactions of Pd and 14-3-3. A number of Pd phosphorylation patterns have been generated with Ca2+/calmodulin-dependent protein† and protein kinase A (25, 26) in vitro, both of which are regulated by Ca2+. Identification of the phosphorylation loci involved in the light-dependent interaction of Pd and 14-3-3 will help to identify kinases and phosphatases responsible for regulating these interactions.
Pd Function.
Pd interacts with all known Gβγ subunits (40). One RIS candidate is the G protein associated with the group III metabotropic glutamate receptor (41), which has been implicated as a feedback modulator of glutamate release in other neurons (42). The associated G protein in such neurons is subclass Gi, a negative regulator of adenylyl cyclase. A similar case experimentally demonstrated is Gβγ regulation of release of hexosaminidase containing vesicles from rat peritoneal mast cells (43, 44). Because types 2 and 3 Gβ are found histochemically in the synaptic region, interactions with more than one G protein may occur.
Now that 14-3-3 also has been identified as a Pd interactive partner, the functional possibilities appear more complex. 14-3-3 plays a role in the exocytosis of catecolamine-containing granules in chromaffin cells (45, 46). This leads to the following hypothesis: light dissociation of the Pd/14-3-3 complex has the double consequence whereby free 14-3-3 potentiates the release of glutamate and dephosphorylated Pd diminishes the constraining effect of the glutamate receptor on glutamate release. These combined actions would make it more difficult for light to block glutamate release and thus require ever larger light flashes for effective excitation as background light increases.
Acknowledgments
M.W.B. expresses his profound gratitude to Dr. William H. Miller of Yale University, whose initial insights and skillful teaching were largely responsible for the introduction of G protein-mediated signal transduction into the arena of photoreceptor biology. We thank Dr. Lee Segel of Weizmann Institute for his kind discussions on models of neurotransmitter release mechanisms. We deeply appreciate the contributions of Dr. William Eldred of Boston University, who provided methods and facilities indispensable for accomplishing the immunocytochemistry. This research was supported National Eye Institute Grant 5R01 EY06816–10 to M.W.B.
Abbreviations
- Pd
phosducin
- Rho
rhodopsin
- ROS
rod outer segment
- Gt
transducin
- RIS
rod inner segment
- IS
inner segment
- MALDI-TOF
matrix-assisted laser desorption ionization–time of flight
- OPL
outer plexiform layer
Footnotes
McLaughlin, J. N., Thulin, C. D., Savage, J. R., Truscott, S. M., Ahn, N. G., Resing, K. A., Soderling, T. R., Bitensky, M. W. & Willardson, B. M. (2000) FASEB J. 14, A1485 (abstr.).
References
- 1.Miller W H, Gorman R E, Bitensky M W. Science. 1971;174:295–297. doi: 10.1126/science.174.4006.295. [DOI] [PubMed] [Google Scholar]
- 2.Keirns J J, Miki N, Bitensky M W, Keirns M. Biochemistry. 1975;14:2760–2766. doi: 10.1021/bi00683a032. [DOI] [PubMed] [Google Scholar]
- 3.Miki N, Baraban J M, Keirns J J, Boyce J J, Bitensky M W. J Biol Chem. 1975;250:6320–6327. [PubMed] [Google Scholar]
- 4.Wheeler G L, Matuo Y, Bitensky M W. Proc Natl Acad Sci USA. 1977;74:4238–4242. doi: 10.1073/pnas.74.10.4238. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Wheeler G L, Matuo Y, Bitensky M W. Nature (London) 1977;269:822–824. doi: 10.1038/269822a0. [DOI] [PubMed] [Google Scholar]
- 6.Shinozawa T, Sen I, Wheeler G L, Bitensky M W. J Supramol Struct. 1979;10:185–190. doi: 10.1002/jss.400100208. [DOI] [PubMed] [Google Scholar]
- 7.Shinozawa T, Uchida S, Martin E, Cafiso D, Hubbell W, Bitensky M W. Proc Natl Acad Sci USA. 1980;77:1408–1411. doi: 10.1073/pnas.77.3.1408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Yamazaki A, Sen I, Bitensky M W, Casnellie J E, Greengard P J. J Biol Chem. 1980;255:11619–11624. [PubMed] [Google Scholar]
- 9.Baylor D. Proc Natl Acad Sci USA. 1996;93:560–565. doi: 10.1073/pnas.93.2.560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Fesenko E E, Kolesnikov S S, Lyubarsky A L. Nature (London) 1985;313:310–313. doi: 10.1038/313310a0. [DOI] [PubMed] [Google Scholar]
- 11.Shuster T A, Farber D B. Biochemistry. 1984;23:515–521. doi: 10.1021/bi00298a018. [DOI] [PubMed] [Google Scholar]
- 12.Attwell D, Borges S, Wu S M, Wilson M. Nature (London) 1987;328:522–524. doi: 10.1038/328522a0. [DOI] [PubMed] [Google Scholar]
- 13.Pugh E N, Jr, Nikonov S, Lamb T D. Curr Opin Neurobiol. 1999;9:410–418. doi: 10.1016/S0959-4388(99)80062-2. [DOI] [PubMed] [Google Scholar]
- 14.Koutalos Y, Yau K W. Trends Neurosci. 1996;19:73–81. doi: 10.1016/0166-2236(96)89624-x. [DOI] [PubMed] [Google Scholar]
- 15.Wilden U, Hall S W, Kuhn H. Proc Natl Acad Sci USA. 1986;83:1174–1178. doi: 10.1073/pnas.83.5.1174. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Dizhoor A M, Ray S, Kumar S, Niemi G, Spencer M, Brolley D, Walsh K A, Philipov P P, Hurley J B, Stryer L. Science. 1991;251:915–918. doi: 10.1126/science.1672047. [DOI] [PubMed] [Google Scholar]
- 17.Gorodovikova E N, Senin I I, Philippov P P. FEBS Lett. 1994;353:171–172. doi: 10.1016/0014-5793(94)01030-7. [DOI] [PubMed] [Google Scholar]
- 18.Chen C K, Inglese J, Lefkowitz R J, Hurley J B. J Biol Chem. 1995;270:18060–18066. doi: 10.1074/jbc.270.30.18060. [DOI] [PubMed] [Google Scholar]
- 19.Kuo C H, Akiyama M, Miki N. Mol Brain Res. 1989;6:1–10. doi: 10.1016/0169-328x(89)90022-3. [DOI] [PubMed] [Google Scholar]
- 20.Craft C M, Lolley R N, Seldin M F, Lee R H. Genomics. 1991;10:400–409. doi: 10.1016/0888-7543(91)90325-9. [DOI] [PubMed] [Google Scholar]
- 21.Abe T, Nakabayashi H, Tamada H, Takagai T, Sakuragi S, Yamaki K, Shinohara T. Gene. 1990;91:209–215. doi: 10.1016/0378-1119(90)90090-e. [DOI] [PubMed] [Google Scholar]
- 22.Bauer P H, Müller S, Puzicha M, Pippig S, Obermaier B, Helmreich E J M, Lohse M J. Nature (London) 1992;358:73–76. doi: 10.1038/358073a0. [DOI] [PubMed] [Google Scholar]
- 23.Danner S, Lohse M J. Proc Natl Acad Sci USA. 1996;93:10145–10150. doi: 10.1073/pnas.93.19.10145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Lee R H, Lieberman B S, Lolley R N. Biochemistry. 1987;26:3983–3990. doi: 10.1021/bi00387a036. [DOI] [PubMed] [Google Scholar]
- 25.Lee R H, Brown B M, Lolley R N. J Biol Chem. 1990;265:15860–15866. [PubMed] [Google Scholar]
- 26.Yoshida T, Willardson B M, Wilkins J F, Jensen G J, Thornton B D, Bitensky M W. J Biol Chem. 1994;269:24050–24057. [PubMed] [Google Scholar]
- 27.Loew A, Ho Y K, Blundell T, Bax B. Structure (London) 1988;6:1007–1019. doi: 10.1016/s0969-2126(98)00102-6. [DOI] [PubMed] [Google Scholar]
- 28.Gaudet R, Bohm A, Sigler P B. Cell. 1996;87:577–588. doi: 10.1016/s0092-8674(00)81376-8. [DOI] [PubMed] [Google Scholar]
- 29.Willardson B M, Wilkins J F, Yoshida T, Bitensky M W. Proc Natl Acad Sci USA. 1996;93:1475–1479. doi: 10.1073/pnas.93.4.1475. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Lee R H, Whelan J P, Lolley R N, McGinnis J F. Exp Eye Res. 1988;46:829–840. doi: 10.1016/s0014-4835(88)80035-6. [DOI] [PubMed] [Google Scholar]
- 31.Whelan J P, McGinnis J F. J Neurosci Res. 1988;20:263–270. doi: 10.1002/jnr.490200216. [DOI] [PubMed] [Google Scholar]
- 32.Aitken A, Jones D, Soneji Y, Howell S. Biochem Soc Trans. 1995;23:605–611. doi: 10.1042/bst0230605. [DOI] [PubMed] [Google Scholar]
- 33.Aitken A. Trends Cell Biol. 1996;6:341–347. doi: 10.1016/0962-8924(96)10029-5. [DOI] [PubMed] [Google Scholar]
- 34.Neville D C, Rozanas C R, Price E M, Gruis D B, Verkman A S, Townsend R R. Protein Sci. 1997;6:2436–2445. doi: 10.1002/pro.5560061117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Uhl R, Desel H, Ryba N, Wagner R. J Biochem Biophys Methods. 1987;14:127–138. doi: 10.1016/0165-022x(87)90015-7. [DOI] [PubMed] [Google Scholar]
- 36.Thulin C D, Howes K, Driscoll C D, Savage J R, Rand T A, Baehr W, Willardson B M. Mol Vis. 1999;5:40–49. [PubMed] [Google Scholar]
- 37.Kuo C H, Miki N. Neurosci Lett. 1989;103:8–10. doi: 10.1016/0304-3940(89)90476-x. [DOI] [PubMed] [Google Scholar]
- 38.Lee R H, Lieberman B S, Lolley R N. Biochemistry. 1987;26:3983–3990. doi: 10.1021/bi00387a036. [DOI] [PubMed] [Google Scholar]
- 39.Peng Y W, Robishaw J D, Levine M A, Yau K W. Proc Natl Acad Sci USA. 1992;89:10882–10886. doi: 10.1073/pnas.89.22.10882. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Müller S, Straub A, Schröder S, Bauer P H, Lohse M J. J Biol Chem. 1996;271:11781–11786. doi: 10.1074/jbc.271.20.11781. [DOI] [PubMed] [Google Scholar]
- 41.Koulen P, Kuhn R, Wässle H, Brandsätter J H. Proc Natl Acad Sci USA. 1999;96:9909–9914. doi: 10.1073/pnas.96.17.9909. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Yusim K, Parnas H, Segel L A. Bull Math Biol. 2000;62:717–757. doi: 10.1006/bulm.2000.0174. [DOI] [PubMed] [Google Scholar]
- 43.Okano K, Monck J R, Fernandez J M. Neuron. 1993;11:165–172. doi: 10.1016/0896-6273(93)90280-5. [DOI] [PubMed] [Google Scholar]
- 44.Pinxteren J F, O'Sullivan A J, Tatham E R, Gomperts B D. EMBO J. 1998;17:6210–6218. doi: 10.1093/emboj/17.21.6210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Morgan A, Burgoyne R D. Nature (London) 1992;355:833–836. doi: 10.1038/355833a0. [DOI] [PubMed] [Google Scholar]
- 46.Roth D, Morgan A, Burgoyne R D. FEBS Lett. 1993;320:207–210. doi: 10.1016/0014-5793(93)80587-k. [DOI] [PubMed] [Google Scholar]