

Abstract
Background and Aims
Manipulation of plant structure can strongly affect light distribution in the canopy and photosynthesis. The aim of this paper is to find a plant ideotype for optimization of light absorption and canopy photosynthesis. Using a static functional structural plant model (FSPM), a range of different plant architectural characteristics was tested for two different seasons in order to find the optimal architecture with respect to light absorption and photosynthesis.
Methods
Simulations were performed with an FSPM of a greenhouse-grown tomato crop. Sensitivity analyses were carried out for leaf elevation angle, leaf phyllotaxis, leaflet angle, leaf shape, leaflet arrangement and internode length. From the results of this analysis two possible ideotypes were proposed. Four different vertical light distributions were also tested, while light absorption cumulated over the whole canopy was kept the same.
Key Results
Photosynthesis was augmented by 6 % in winter and reduced by 7 % in summer, when light absorption in the top part of the canopy was increased by 25 %, while not changing light absorption of the canopy as a whole. The measured plant structure was already optimal with respect to leaf elevation angle, leaflet angle and leaflet arrangement for both light absorption and photosynthesis while phyllotaxis had no effect. Increasing the length : width ratio of leaves by 1·5 or increasing internode length from 7 cm to 12 cm led to an increase of 6–10 % for light absorption and photosynthesis.
Conclusions
At high light intensities (summer) deeper penetration of light in the canopy improves crop photosynthesis, but not at low light intensities (winter). In particular, internode length and leaf shape affect the vertical distribution of light in the canopy. A new plant ideotype with more spacious canopy architecture due to long internodes and long and narrow leaves led to an increase in crop photosynthesis of up to 10 %.
Keywords: Plant architecture, 3-D light distribution, daily assimilation, tomato, Solanum lycopersicum, functional–structural plant modelling, virtual canopy
INTRODUCTION
Light absorption is an important factor for determining crop yield, being one of the driving forces behind plant photosynthesis, and at the same time is highly dependent on single-plant architecture as well as on overall canopy structure (Niinemets, 2007). Plant architectural characteristics (such as the number and geometry of organs, i.e. their shape and position within the plant and the canopy), are genotype specific, while at the same time highly dependent on the climatic conditions at the time of their initiation and development (Godin, 2000). Falster and Westoby (2003) have shown that steeper elevation angles in a number of species improve absorption at higher sun elevations and, therefore, carbon gain through assimilation as it allows more light to penetrate to the lower leaves. While the importance of leaf elevation angles for an improved light-absorption strategy at the level of the whole plant has been shown in a number of studies (Pearcy and Yang, 1998; Sinoquet et al., 2005), reports about the importance of leaf phyllotaxis are contradictory as some studies did and some did not observe effects on the light absorption of the canopy (Brites and Valladares, 2005). Furthermore, aspects such as the elevation angles of the leaflets of composite leaves have not been, to the best of our knowledge, previously investigated. Both leaf shape and size are important aspects of leaf morphology affecting mutual shading of leaves and light absorption of the canopy (Falster and Westoby, 2003).
The quantitative exploration of the specific effects of each plant architectural characteristic on light absorption and photosynthesis was hardly possible until the introduction of spatially explicit models considering plant architecture at the organ level (Vos et al., 2010). General crop models are powerful tools towards a better understanding of plant processes and for testing case scenarios (Marcelis et al., 1998; Vos et al., 2007). More specifically, functional–structural plant models (FSPM) have been introduced as a relatively recent paradigm in plant modelling where physiological processes are coupled with an explicit 3-D plant structure (Vos et al., 2010), often supplied with a mutual feedback between physiology and structure. Modelling on the basis of a 3-D structure gives the opportunity to investigate more in-depth the effect of specific architectural characteristics such as leaf angle, leaf length or leaf shape (Vos et al., 2010). Sinoquet et al. (2005) used FSPMs to show the effect of leaf and branch distribution on light absorption of trees. Sarlikioti et al. (2011) developed a static tomato FSPM to explore the spatial distribution of light absorption and photosynthesis in a tomato canopy. They demonstrated the importance of an explicitly described leaf angle distribution for simulating light absorption and photosynthesis. Accurately calibrated FSPMs can convey a better understanding of the light distribution inside the canopy and also provide us with a tool to define the optimal set of architectural characteristics for maximizing canopy photosynthesis, allocation of assimilates to growing organs and ultimately crop yield.
Donald (1968) defined the ‘crop ideotype’ in the context of cereal breeding as an idealized plant type with a specific combination of characteristics favourable for photosynthesis, growth and grain production based on knowledge of plant and crop physiology and morphology. He argued that it would be more efficient to define a plant type that was theoretically efficient and then breed for this. The crop ideotype is thus an idealized crop consisting of a plant type with a specific combination of characteristics based on the detailed knowledge of morphological and physiological plant traits (Peng et al., 2008) as well as mutual interactions among plants of the canopy. These traits often are also contributing to plant architecture. Modifications of the arrangement and size of leaves can affect light availability, especially in the lower parts of the canopy, and alter leaf photosynthetic activity by adjusting light-harvesting efficiency (Werner et al., 2001). As an example, a reduction in leaf clustering can increase light absorption and enhance photosynthetic productivity at canopy level (De Castro and Fetcher, 1999). Morphological characteristics such as leaf inclination and leaf shape are often inherited as simple traits (i.e. under the influence of one or a few major genes) in the plant (Thurling, 1991) and can be used to create a more open canopy structure. These breeding traits can be strongly affected by the environment under stress conditions (Valladares and Niinemets, 2007). Tomato (Solanum lycopersicum) is a species that exhibits a high variability in vegetative morphology (Peralta and Spooner, 2000) ranging from small leaves with a few leaflets to big ones with many leaflets. Lately, studies of the genetic basis of this variation at the leaf level have shown that there are leaf-specific genes that control its shape and morphology (Frary et al., 2004). This genetic background knowledge in combination with the detailed information on the effect of leaf topology and geometry on light absorption and photosynthesis could help to identify or approximate the theoretical optimum of plant architecture.
The objective of this study is to define a plant ideotype for greenhouse-grown tomato with respect to optimization of light absorption and photosynthesis at the canopy level. A range of different plant architectural characteristics were tested under light conditions of a typical summer and winter day in order to define the ideal for each case. We tested two hypotheses: (1) the manipulation of plant structure of a greenhouse-grown tomato crop can lead to substantial improvement in crop photosynthesis even when leaf area index and leaf photosynthetic characteristics remain unaltered; and (2) a more spacious canopy architecture improves crop photosynthesis. For this purpose we used the static functional structural tomato model developed by Sarlikioti et al. (2011).
MATERIALS AND METHODS
Model description
Simulations were performed with a functional–structural model of tomato (Solanum lycopersicum). This model was built as a parametric open L-system using the cpfg language within the platform L-Studio (Prusinkiewicz, 1999) consisting of three modules (model structure and parameterization have been described in detail in Sarlikioti et al., 2011):
Architectural module
This is essentially a static 3-D reconstruction of tomato plant architecture, in which each plant consists of 27 phytomers with the first phytomer being the one at the top of the plant (basipetal ranking). A phytomer is the basic architectural unit consisting of a leaf and an internode, the leaf itself being made up of a leaf rachis, a central midrib and 13 individual leaflets, each one of them consisting of a blade and a petiole, of which one is terminal and 12 are lateral. The blade of each leaflet is described as a flat polygon instead of one with a curvature for reasons of calculation efficiency. Characteristics are: leaf elevation angle (defined as the angle between the leaf rachis, at its insertion point to the stem, and the horizontal plane), leaf length (defined as the distance from the leaf insertion point at the stem to the tip of the terminal leaflet) and leaf width (defined as the distance between the tips of the two longest lateral leaflets). The construction of the leaf leads to a 3-D object. The architectural model was parameterized during a summer period for a fully grown tomato crop (Solanum lycopersicum ‘Aranca’) grown in a high-wired greenhouse system in Bleiswijk, The Netherlands (52°01′N, 4°32′E) with a density of 4·1 stems m−2 (see Sarlikioti et al., 2011). LAI was equal to 3·6 m2 of leaf area per square metre of floor area. Every week the lowest leaves were removed, the plants were lowered and kept at the same height throughout the season as it is common practice in The Netherlands. Therefore plant architecture remained ‘static’ at least during each season (summer, winter). For that reason a static modelling approach was used.
Light module
Light calculations are based on a nested radiosity approach developed by Chelle and Andrieu (1998). The module requires as an input the amount of light absorbed by each plant organ, the leaf transmittance and reflectance coefficients for the upper and lower side of the leaf (upper side reflectance = 0·17, upper side transmittance = 0·06, lower side reflectance = 0·12, lower side transmittance = 0·03) and the light from the light sources that were used to simulate the sky. In The Netherlands, greenhouses are usually larger than 1 ha. In order to approach real greenhouse cultivation conditions better, an infinite canopy was assumed in which the basic unit is theoretically reproduced in all directions, thus avoiding a border effect during the calculations. The basic unit consisted of 20 plants arranged in two double rows with five plants per row. The distance between the double rows was 1·2 m (path), the distance between each row of the double row (within the row distance) was 0·4 m and the distance between plants within the row was 0·3 m.
Photosynthesis module
Photosynthesis calculations are based on the biochemical model of Farquhar et al. (1980). This model requires as input the computed light absorbed per leaflet in the model and photosynthetic parameters that were derived from the experimental data (Sarlikioti et al., 2011). All photosynthetic parameters (Vmax, α, θ, etc.) were assumed invariate with canopy depth, except for Jmax, which followed a logistic pattern from the top to the bottom of the canopy (Jmax at the top = 265 µmol e− m−2 s−1, Jmax at the bottom = 180 µmol e− m−2 s−1).
Case studies
In this study we attempted to define the importance of each architectural component for optimum light absorption and canopy photosynthesis by performing a sensitivity analysis of the most important components as they appear in the literature. Each component is described in detail below.
Simulations were performed under summer and winter light conditions and for two distinctive days (21 December and 21 June) using ten-yearly (2000–2010) averages, with an hourly time step, of direct and diffuse light incident at these dates under Dutch conditions. Daily global radiation outside the greenhouse was equal to 3·5 MJ m−2 d−1 in winter and 26 MJ m−2 d−1 in summer. Transmissivity of the greenhouse was 60 %. For direct light conditions, light sources were given the same Cartesian co-ordinates as the solar trajectory. To simulate diffuse light, light sources were positioned on a virtual hemisphere around the architectural mock-up. In total, 48 directional light sources were used, with daily intensity equal to 1·5 MJ m−2 d−1 in winter and 7·5 MJ m−2 d−1 in summer. The fraction of diffuse light was equal to 0·3 for winter and 0·22 for summer. Leaf temperature was 23 °C and calculations were performed for CO2 levels equal to 400 ppm. Computed hourly assimilation rates were integrated to daily amount of assimilates produced.
Vertical light distribution scenario
To understand the effect of vertical light distribution on canopy photosynthesis, we constructed four light absorption curves (Fig. 1A) that were based on the light absorption of the reference structure for each date. Light absorption was increased and reduced from the first to the 8th phytomer by 10 % and 25 %, respectively, while the light absorption of the canopy as a whole remained the same. Canopy photosynthesis was calculated for each curve.
Fig. 1.
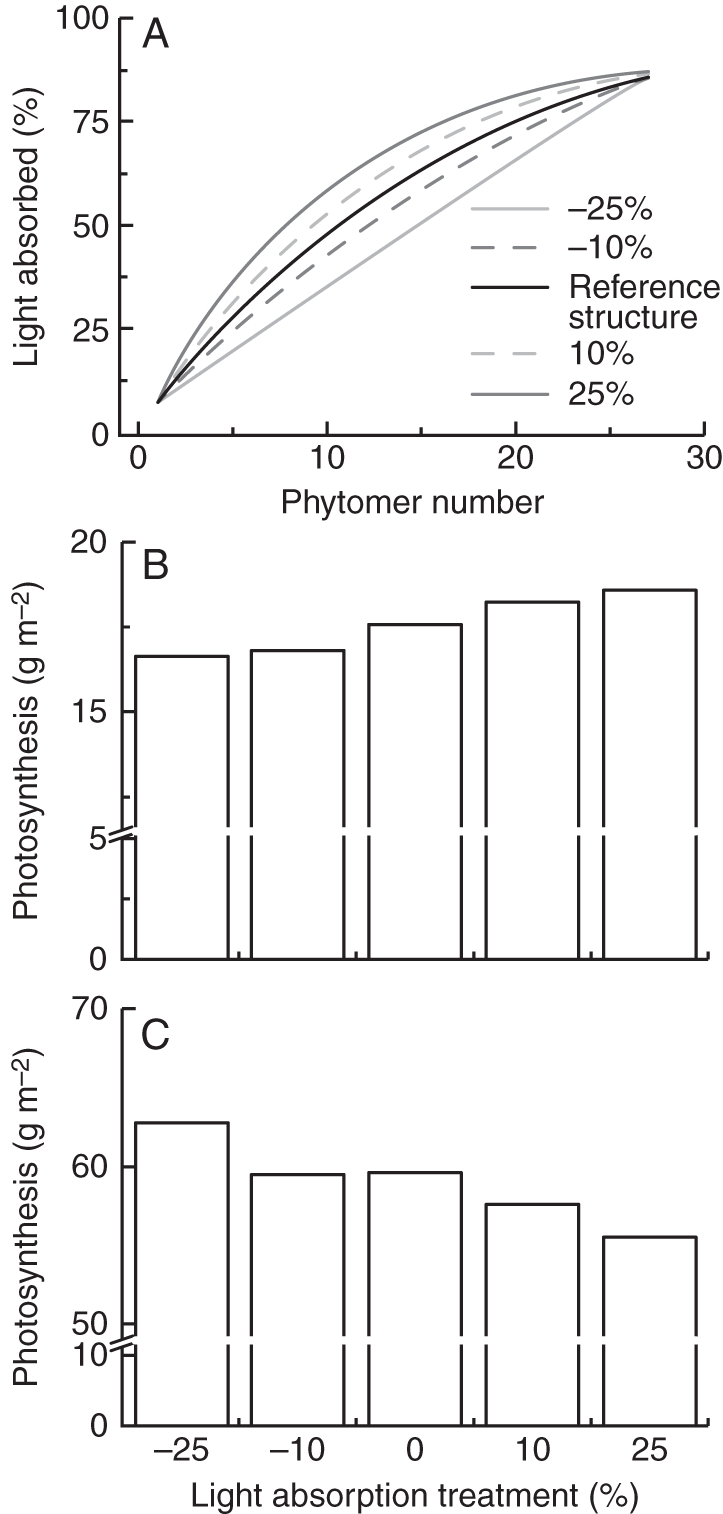
Five different vertical light absorption profiles (A) were imposed as treatments for studying the optimal profile for maximum crop photosynthesis. The absorbed light is the cumulated value from top (phytomer 1) to the bottom of the canopy. Photosynthesis at 1200 h (g CH2O m−2 h−1) is presented for each treatment for winter (B) and summer (C). Treatments for absorption profile are –25 %, –10 %, 0 %, +10 % and +25 %. These values refer to the percentage change in light absorption in the upper eight phytomers of the canopy while light absorption cumulated over the whole canopy was kept constant.
Leaf elevation angle
Here, we studied the effect of modifying the leaf elevation angle with respect to the original angular distribution. As a starting value we assumed an explicitly described leaf angle distribution (Sarlikioti et al., 2011). Simulations were carried out by adding or subtracting 15° or 30° to the measured value of each leaf in the canopy. In the reference crop the leaf angle distribution ranged from –23° for bottom leaves to 15° for top leaves.
Leaf phyllotaxis
The phyllotactic angle is defined as the angle between two subsequent leaves along the plant stem. Atherton and Rudich (1986) reported that tomato plants follow a common 135° phyllotaxis. In reality, however, plants tend to rearrange their leaves, thereby deviating from this value. In previous experiments, it was found that the phyllotactic angle on a tomato row crop is about 160° (Sarlikioti et al., 2011). Here we assess the effect of leaf phyllotaxis for values equal to 110°, 135°, 150° and 180°.
Leaflet angle
As leaflet angle we define the angle between the leaflet petiole and the leaf rachis. In a composite leaf, leaflet angles steadily decrease from the proximal to the terminal leaflet. On average, the measured leaflet angle of tomato leaves was 22°, ranging from 35° for the basal leaflets to 0° for the terminal leaflet (Sarlikioti et al., 2011). In the present study this original distribution (reference crop) was compared with leaves having all leaflets oriented at an angle of 22° or 0°.
Leaf shape
The ratio between leaf length and leaf width (LW ratio) was used as a convenient measure of leaf shape. In the model the default value of the LW ratio is equal to 1·02. Leaf ratios of 0·5, 0·75, 1·25, 1·5 and 2 times the original ratio were tested for their effect on light absorption and canopy photosynthesis. These ratios resulted in a range of leaf shapes from wide and short leaves to long and narrow ones. For all simulations, the leaf area index per plant was kept constant at 3·6 m2 leaf area per m−2 floor area.
Leaflet arrangement on a leaf
The area of each leaf is equal to the sum of the areas of each of its leaflets. In the leaf of a typical tomato cultivar, pairs of big and small leaflets alternate. A wide range of leaf types can be found in tomato. We investigated the (crop scale) effect of leaf types with all leaflets having the same area as well as the effect of having fewer but bigger leaflets (Fig. 2). Also the effect of an increase in petiole length by 20 % of the original value was investigated. In all simulations the area per leaf (sum of all leaflets) was kept the same.
Fig. 2.

Schematic representation of leaflet arrangement scenarios. The total area per composite leaf was kept constant.
Internode length
As internode length, we define the distance between the insertion points of two consecutive plant organs (leaves or trusses). Measured average internode length was 7·45 cm (standard deviation = 0·8) (Sarlikioti et al., 2011). Here we investigated the effect of internode lengths of 3 cm, 5 cm, 10 cm and 12 cm. In all simulations leaf number (13) and area per leaf (m2) were kept constant.
Ideotyping scenarios
After the above assessment of architectural characteristics of the tomato plant we found that a number of parameters improve the light absorption as well as canopy photosynthesis. Based on these results, we designed the following two scenarios.
Scenario (A)
For this scenario architectural characteristics were chosen, which by themselves had produced a minimum increase in canopy photosynthesis by 4 %. Internode length was set to 10 cm and the LW ratio to 1·5. All other architectural characteristics were kept the same as in the reference structure.
Scenario (B)
This scenario aimed at reconstructing an open structure of the canopy: The internode length was again set to 10 cm and for leaflet arrangement the leaves with longer petioles (Fig. 2) were chosen. All other architectural characteristics remained the same as in the reference structure.
RESULTS
Vertical light distribution scenario
Imposing five different vertical light absorption profiles, while keeping light absorption cumulated over the whole canopy constant (Fig. 1A), showed some distinct effects on canopy photosynthesis. Under winter light conditions at noon, canopy photosynthesis increased by 6 % when absorption in the top part of the canopy (upper eight phytomers) was increased by 25 % (Fig. 1B). In contrast, under summer light conditions, for the same increase of light absorption canopy photosynthesis was reduced by 7 % (Fig. 1C).
Leaf elevation angle
Highest light absorption was achieved with the original leaf angle distribution (leaf angle decreasing from 15° for top leaves to –23° for bottom leaves) under winter light conditions (Fig. 3A) while under summer conditions it was maximum when elvation angle was changed by 30° (Fig. 3B). Changes in leaf angle of ±15° decreased light absorption by 5 % and an increase of the leaf angle to +30° led to a decrease of 6 % under winter conditions (Fig. 3A). A decrease in leaf angle by 30° distinctively decreased light absorption by 18 %. Under summer light conditions (Fig. 3B), the increase was equal to 3 % for ±30° treatments. Interestingly, in winter simulations, the decrease in elevation angle by 30° did not only decrease light absorption but also changed the vertical distribution of light absorption leading to a higher absorption in the top part of the canopy (Fig. 3).
Fig. 3.

Effect of leaf elevation angle on cumulated light absorption from top (phytomer 1) to bottom of the canopy under winter (A) and summer (B) conditions. 0° refers to the reference structure with an angle distribution ranging from 15° (top leaves) to –23° (bottom leaves).
In general a change in elevation angle reduced canopy photosynthesis by 6–7 % in winter, which is similar to the reduction in light absorption (Table 1). However, when the angle was reduced by 30°, canopy photosynthesis decreased only by 11 % while absorption decreased by 18 %.
Table 1.
Canopy photosynthesis (assimilation rate, g CH2O m−2 d−1) in response to leaf elevation angle, phyllotaxis, leaflet angle and arrangement, leaf length : width ratio, internode length scenarios as well as the optimized architectures
| Winter | Summer | |
|---|---|---|
| Leaf elevation angle | ||
| –30° | 4·7 | 40·5 |
| –15° | 5·0 | 40·5 |
| 0 (reference structure) | 5·3 | 42·0 |
| + 15° | 5·0 | 40·5 |
| + 30° | 4·9 | 41·7 |
| Leaf azimuth angle | ||
| 110 | 5·2 | 42·3 |
| 135 | 5·3 | 41·8 |
| 160 (reference structure) | 5·3 | 42·0 |
| 180 | 5·1 | 41·7 |
| Leaflet angle | ||
| 0° | 5·1 | 37·3 |
| 22° | 5·2 | 40·5 |
| From 35° to 0° (reference structure) | 5·3 | |
| 42° | ||
| Leaflet arrangement | ||
| Longer leaflet petioles | 5·2 | 39·4 |
| Same area leaflets | 5·0 | 39·3 |
| Fewer/bigger leaflets | 4·9 | 41·8 |
| Reference structure | 5·3 | 42·0 |
| Leaf length : width ratio | ||
| 0·5 | 4·7 | 34·3 |
| 0·75 | 4·8 | 39·8 |
| 1·02 (reference structure) | 5·3 | 42·0 |
| 1·25 | 5·5 | 44·2 |
| 1·5 | 5·6 | 44·6 |
| Internode length | ||
| 3 cm | 4·3 | 37·0 |
| 5 cm | 5·0 | 38·8 |
| 7 cm (reference structure) | 5·3 | 42·0 |
| 10 cm | 5·6 | 44·2 |
| 12 cm | 5·7 | 44·6 |
| Optimized canopy architecture | ||
| Scenario A | 5·6 | 45·2 |
| Scenario B | 5·9 | 46·7 |
Leaf phyllotaxis
Changes in phyllotactic angle hardly affected light absorption (data not shown) and canopy photosynthesis (Table 1).
Leaflet angle
An increase in leaflet angle from completely horizontal leaflets (0°) to an angle of 22° resulted in an increase in light absorption by 2 % in winter (Fig. 4A) and 8 % (Fig. 4B) in summer. The subsequent increase in canopy photosynthesis was 2 % under winter and 8 % under summer light conditions (Table 1). When all leaflet angles were 22°, light absorption was 2 % lower under winter and 4 % under summer light conditions compared with the reference plants (angle decreasing from 35° for basal leaves to 0° for the terminal leaflet) (Fig. 4), while canopy photosynthesis exhibited the same increase for both seasonal conditions.
Fig. 4.

Effect of leaflet angle on cumulated light absorption from top (phytomer 1) to bottom of the canopy under winter (A) and summer (B) conditions. In the reference structure the angle is decreasing from 35° for basal leaflets to 0° for the terminal leaflet.
Leaf shape
The simulations showed that longer and narrower leaves (LW > 1) increased cumulative light absorption of the canopy in comparison to shorter and wider leaves when leaf area was kept constant. The LW ratio was positively correlated with light absorption (Fig. 5). Light absorption was reduced by 8 % and 12 %, respectively, in winter (Fig. 5A), and 14 % and 23 %, respectively, in summer for a leaf ratio reduction of 0·25 and 0·5 (Fig. 5B). An increase in the LW ratio to 1·25 of the reference value resulted in an increase in light absorption by 5 % in winter and 4 % in summer. When the ratio was 1·5 times as high as the reference ratio, light absorption was increased by 8 % in winter and 10 % in summer. An increase in the LW ratio to 2 did not further increase light absorption (data not shown). The effect of the different treatments on canopy photosynthesis followed the trends observed in light absorption, but the total increase was 1–2 % smaller compared with the increase in light absorption (Table 1).
Fig. 5.

Effect of leaf shape on cumulated light absorption from top (phytomer 1) to bottom of the canopy under winter (A) and summer (B) conditions. Leaf shape is referring to the LW ratio with reference structure having a ratio of 1.
Leaflet arrangement
Changing the arrangement of leaflets within a leaf while keeping leaf area constant (Fig. 2) affected the vertical light distribution as well as the total light absorption. The reference structure showed the maximum absorption under both summer and winter conditions (Fig. 6). Plants with the reference leaves, i.e. exhibiting irregular leaflet size, absorbed more light in the top portion of the canopy (0·5–1 m) than leaves with equally sized leaflets. Leaves with fewer but bigger leaflets absorbed more light than leaves with more but smaller leaflets (Fig. 6). An increase in petiole length slightly diminished both light absorption and canopy photosynthesis in both seasonal conditions. The effect of the difference in vertical light distribution between the scenarios was reflected on canopy photosynthesis where the leaf with leaflets of the same area exhibited 2 % less canopy photosynthesis in winter and 6 % in summer than the other two scenarios (Table 1).
Fig. 6.

Effect of leaflet arrangement on cumulated light absorption from top (phytomer 1) to bottom of the canopy under winter (A) and summer (B) conditions.
Internode length
Light absorption and canopy photosynthesis were strongly affected by internode length. A decrease in the average internode length from 7 cm to 3 cm reduced cumulative light absorption by 14 % in winter and by 12 % in summer. An increase in internode length from 7 cm to 10 cm resulted in an increase in light absorbed of 4 % in winter and 5 % in summer, while a further increase in internode lengths to 12 cm resulted in a further increase in light absorption by 2 % for each date. With respect to canopy photosynthesis, the increase in internode length to 10 cm increased canopy photosynthesis by 5–6 % in summer and winter (Table 1). An increase to 12 cm increased photosynthesis by 8 % and 6 % for winter and summer, respectively.
Ideotyping scenarios
According to the above assessment of architectural characteristics of the tomato plant, two ideotyping scenarios were designed. In scenario A, internode length was set to 10 cm and the LW ratio to 1·5, while all other architectural characteristics were kept the same as in the reference structure. In scenario B, plant structure consisted of the same long internodes (10 cm) and a leaflet arrangement with long leaf petioles was chosen, while all other architectural characters remained the same as in the reference structure. Scenario A improved both light absorption (data not shown) and canopy photosynthesis by 6 % (Table 1) in winter and by 8 % in summer, respectively. In scenario B, the construction of a more open structure of the canopy resulted in an increase in light absorption of 11 % in both winter and summer, with a similar increase in canopy photosynthesis.
DISCUSSION
From the result of this work, we can conclude that both light absorbed and the vertical distribution of light in the canopy are very important for crop photosynthesis. Analysing the effects of vertical light distribution showed that these effects strongly differ between winter and summer light conditions. Deep penetration of light into the canopy has positive effects in summer, while these effects are negative in winter. Under summer light conditions photosynthesis of the upper leaves may be close to saturation, which explains the positive effects of deep penetration of light. This is in contrast to the photosynthesis rate of lower leaves, which remains unsaturated. Under winter light conditions, photosynthesis of the upper leaves is far from saturation. With the same light intensity at leaf level, upper leaves have a higher rate of photosynthesis than lower leaves (e.g. Sarlikioti et al., 2011). This effect dominates in winter, resulting in the lower crop photosynthesis when the light penetrates deeply into the canopy.
Leaf elevation angles are maybe among the best investigated traits. It has been shown that a change in the elevation angles significantly influences light captured in different environments (Valladares and Pearcy, 1998; Niinemets and Fleck, 2002). In a previous study (Sarlikioti et al., 2011), it was shown that changes in leaf angles could have a substantial effect on both light absorption and photosynthesis. In the current study, optimal results were achieved when leaf elevation-angle distribution ranged between 15° (top) and –23° (bottom), indicating that during the cultivation period the plant orientates its leaves in such a way as to maximize light absorption and therefore photosynthesis. Deviations from that range failed to distinctly increase both light absorption and photosynthesis, though the vertical light distribution was affected.
Modifying the phyllotactic angle resulted in no improvement of light absorption and photosynthesis. These findings agree with those of Niklas (1998) who reported that phyllotactic angle in a crop has no effect on light absorption or photosynthesis. In contrast to this, Zotz et al. (2002) reported that a change of leaf phyllotaxis to a golden angle of 137·5° significantly improved the light capture efficiency in an epiphytic plant. Nevertheless, phyllotactic angle is very dependent on light competition. Although a change in the angle as such might have no direct effect on light absorption and photosynthesis, it might be followed by changes in other architectural characteristics that may lead to an increase in plant efficiency with respect to light absorption and photosynthesis.
In our study we found that an increase in LW ratio of leaves by 0·5 boosted light absorption by 8 % in winter and 10 % in summer. In environments where light is not an inhibiting parameter (e.g. because of an excess in radiation), structures that avoid mutual shading of the leaf components as well as the shading between neighbouring leaves can be advantageous for optimization of light absorption. A high leaf LW ratio has been reported to have a positive effect on light capture and crop photosynthesis in many species (Falster and Westoby, 2003). Other aspects of morphology of composite leaves such as shape and number of leaflets have, to our knowledge, hardly been investigated. In forest species it has been shown that the space and degree of overlap of the leaves in leaf clusters on the same branch has a significant effect on light capture (Planchais and Sinoquet, 1998).
Takenaka et al. (1994) showed that, in general, an increase in internode length causes an increase in light absorption and photosynthetic efficiency in nature; in fact, hormonally mediated internode elongation is among the most important mechanisms of the so-called shade avoidance syndrome, a set of processes that enable a plant to avoid shading by neighbouring plants (Smith and Whitelam, 1997). Our results showed that plants with longer internodes exhibited higher light absorption and photosynthesis both in winter and summer.
The combination of different architectural parameters in the ideotype scenarios resulted in an increase in both light absorption and canopy photosynthesis. Both ideotype scenarios aimed at creating a more open structure with more light absorption. In both cases, this combination of characteristics led to an increase in photosynthesis which, in turn, could potentially result in a yield increase. Modern developments in plant breeding can use this type of information to produce more efficient genotypes in terms of canopy photosynthesis. On the other hand plant architectural characteristics are very plastic and plants are usually dynamically adapting to their environment (Valladares et al., 2007), thereby potentially eluding breeding efforts towards a light absorption ideotype. For example, during the course of the day, leaf movement has been observed (Kao and Forseth, 1992) or plants tend to readjust their position when facing an intense environmental factor such as, for example, shading (Kahlen et al., 2008). Thus, even if this type of static model can give us a good quantification of the effect of each parameter on light absorption and photosynthesis, the ensuing genotype has to be tested under actual cultivation conditions to verify the expectations.
In the simulation model photosynthetic parameters decreased from top to the bottom of the canopy. These parameters were not adjusted with the increase or decrease of light intensities in the middle and lower parts of the canopy. In other words, no dynamic adaptation of these parameters to the new light profiles induced by the case studies was considered. In reality, leaf photosynthetic potential, of course, adapts to the long-term light conditions a leaf has been exposed to (Gonzáles-Real et al., 2007), as well as the short-term changes in light climate during the day (Schurr et al., 2006). Experimental data describing these phenomena could help to improve the model calculations by taking into account these adaptations.
Conclusions
The importance of different plant architectural components for light absorption and photosynthesis was investigated in detail using a static virtual plant. Our simulations lead to the conclusion that the most important architectural traits with respect to the optimization of light absorption and photosynthesis are internode length and leaf shape. We also assessed the importance of vertical light distribution for canopy photosynthesis and showed that the advantage of a deeper penetration of light in the canopy depends on the season.
ACKNOWLEDGEMENTS
The authors would like to thank two anonymous reviewers for providing helpful comments on a previous version of this manuscript. We also thank the Greek State Scholarships Foundation for awarding a scholarship to V.S. and Essent/Powerhouse for financial support to L.F.M.M.
LITERATURE CITED
- Atherton JG, Rudich J. The tomato crop: a scientific basis for improvement. London: Chapman and Hall; 1986. [Google Scholar]
- Brites D, Valladares F. Implications of opposite phyllotaxis for light interception efficiency of Mediterranean woody plants. Trees. 2005;19:671–679. [Google Scholar]
- Chelle M, Andrieu B. The nested radiosity model for the distribution of light within plant canopies. Ecological Modelling. 1998;111:75–91. [Google Scholar]
- De Castro F, Fetcher N. The effect of leaf clustering in the interception of light in vegetal canopies: theoretical considerations. Ecological Modelling. 1999;116:125–134. [Google Scholar]
- Donald CM. The breeding of crop ideotypes. Euphytica. 1968;17:385–403. [Google Scholar]
- Falster DS, Westoby M. Leaf size and angle vary widely across species: what consequences for light absorption? New Phytologist. 2003;158:509–525. doi: 10.1046/j.1469-8137.2003.00765.x. [DOI] [PubMed] [Google Scholar]
- Farquhar GD, von Caemmerer S, Berry JA. A biochemical-model of photosynthetic CO2 assimilation in leaves of C3 species. Planta. 1980;149:78–90. doi: 10.1007/BF00386231. [DOI] [PubMed] [Google Scholar]
- Frary A, Fritz LA, Tanksley SD. A comparative study of the genetic bases of natural variation in tomato leaf, sepal, and petal morphology. Theoretical and Applied Genetics. 2004;109:523–533. doi: 10.1007/s00122-004-1669-x. [DOI] [PubMed] [Google Scholar]
- Godin C. Representing and encoding plant architecture: a review. Annals of Forest Science. 2000;57:413–438. [Google Scholar]
- González-Real MM, Baille A, Gutiérrez Colomer RP. Leaf photosynthetic properties and radiation profiles in a rose canopy (Rosa hybrida L.) with bent shoots. Scientia Horticulturae. 2007;114:177–187. [Google Scholar]
- Kahlen K, Wiechers D, Stützel H. Modelling leaf phototropism in a cucumber canopy. Functional Plant Biology. 2008;35:876–884. doi: 10.1071/FP08034. [DOI] [PubMed] [Google Scholar]
- Kao WY, Forseth IN. Diurnal leaf movement, chlorophyll fluorescence and carbon assimilation in soybean grown under different nitrogen and water availabilities. Plant, Cell & Environment. 1992;15:703–710. [Google Scholar]
- Marcelis LFM, Heuvelink E, Goudriaan J. Modelling biomass production and yield of horticultural crops: a review. Scientia Horticulturae. 1998;74:83–111. [Google Scholar]
- Niinemets Ü. Photosynthesis and resource distribution through plant canopies. Plant, Cell & Environment. 2007;30:1052–1071. doi: 10.1111/j.1365-3040.2007.01683.x. [DOI] [PubMed] [Google Scholar]
- Niinemets Ü, Fleck S. Petiole mechanics, leaf inclination, morphology, and investment in support in relation to light availability in the canopy of Liriodendron tulipifera. Oecologia. 2002;132:21–33. doi: 10.1007/s00442-002-0902-z. [DOI] [PubMed] [Google Scholar]
- Niklas KJ. Light harvesting ‘fitness landscapes’ for vertical shoots with different phyllotactic patterns. In: Jean RV, Barabé D, editors. Symmetry in plants. Singapore: World Scientific Publishing Co; 1998. pp. 759–773. [Google Scholar]
- Pearcy RW, Yang W. The functional morphology of light capture and carbon gain in the Redwood forest understorey plant Adenocaulon bicolor Hook. Functional Ecology. 1998;12:543–552. [Google Scholar]
- Peng S, Khush GS, Virk P, Tang Q, Zou Y. Progress in ideotype breeding to increase rice yield potential. Field Crops Research. 2008;108:32–38. [Google Scholar]
- Peralta IE, Spooner DM. Classification of wild tomatoes: a review. Kurtziana. 2000;28:45–54. [Google Scholar]
- Planchais I, Sinoquet H. Foliage determinants of light absorption in sunny and shaded branches of Fagus sylvatica (L.) Agricultural and Forest Meteorology. 1998;89:241–253. [Google Scholar]
- Prusinkiewicz P. A look at the visual modelling of plants using L-systems. Agronomie. 1999;19:211–224. [Google Scholar]
- Sarlikioti V, de Visser PHB, Marcelis LFM. Exploring the spatial distribution of light absorption and photosynthesis of canopies by means of a functional–structural plant model. Annals of Botany. 2011;107:875–883. doi: 10.1093/aob/mcr006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schurr U, Walter A, Rascher U. Functional dynamics of plant growth and photosynthesis – from steady-state to dynamics – from homogeneity to heterogeneity. Plant, Cell & Environment. 2006;29:340–352. doi: 10.1111/j.1365-3040.2005.01490.x. [DOI] [PubMed] [Google Scholar]
- Sinoquet H, Sonohat G, Phattaralerphong J, Godin C. Foliage randomness and light absorption in 3-D digitized trees: an analysis from multiscale discretization of the canopy. Plant, Cell & Environment. 2005;28:1158–1170. [Google Scholar]
- Smith H, Whitelam GC. The shade avoidance syndrome: multiple responses mediated by multiple phytochromes. Plant, Cell & Environment. 1997;20:840–844. [Google Scholar]
- Takenaka A. Effects of leaf blade narrowness and petiole length on the light capture efficiency of a shoot. Ecological Research. 1994;9:109–114. [Google Scholar]
- Thurling N. Application of the ideotype concept in breeding for higher yield in the oilseed brassicas. Field Crops Research. 1991;26:201–219. [Google Scholar]
- Valladares F, Pearcy RW. The functional ecology of shoot architecture in sun and shade plants of Heteromeles arbutifolia M. Roem., a Californian chaparral shrub. Oecologia. 1998;114:1–10. doi: 10.1007/s004420050413. [DOI] [PubMed] [Google Scholar]
- Valladares F, Niinemets Ü. The architecture of plant crowns: from design rules to light capture and performance. In: Pugnaire F, Valladares F, editors. Functional plant ecology. New York, NY: Taylor and Francis; 2007. pp. 101–149. [Google Scholar]
- Valladares F, Gianoli E, Gómez JM. Ecological limits to plant phenotypic plasticity. New Phytologist. 2007;176:749–763. doi: 10.1111/j.1469-8137.2007.02275.x. [DOI] [PubMed] [Google Scholar]
- Vos J, Marcelis LFM, Evers JB. Functional–structural plant modelling in crop production: adding a dimension. In: Vos J, Marcelis LFM, De Visser PHB, Struik PC, Evers JB, editors. Functional–structural plant modelling in crop production. Dordrecht: Springer; 2007. pp. 1–12. [Google Scholar]
- Vos J, Evers JB, Buck-Sorlin GH, Andrieu B, Chelle M, de Visser PHB. Functional–structural plant modelling: a new versatile tool in crop science. Journal of Experimental Botany. 2010;61:2101–2115. doi: 10.1093/jxb/erp345. [DOI] [PubMed] [Google Scholar]
- Werner C, Ryel RJ, Correia O, Beyschlag W. Structural and functional variability within the canopy and its relevance for carbon gain and stress avoidance. Acta Oecologica. 2001;22:129–138. [Google Scholar]
- Zotz G, Reichling P, Valladares F. A simulation study on the importance of size and related changes in leaf morphology and physiology for carbon gain in an epiphytic bromeliad. Annals of Botany. 2002;90:437–443. doi: 10.1093/aob/mcf208. [DOI] [PMC free article] [PubMed] [Google Scholar]