

Abstract
The floor plate plays a key role in patterning axonal trajectory in the embryonic spinal cord by providing both long-range and local guidance cues that promote or inhibit axonal growth toward and across the ventral midline of the spinal cord, thus acting as an intermediate target for a number of crossing (commissural) and noncrossing (motor) axons. F-spondin, a secreted adhesion molecule expressed in the embryonic floor plate and the caudal somite of birds, plays a dual role in patterning the nervous system. It promotes adhesion and outgrowth of commissural axons and inhibits adhesion of neural crest cells. In the current study, we demonstrate that outgrowth of embryonic motor axons also is inhibited by F-spondin protein in a contact-repulsion fashion. Three independent lines of evidence support our hypothesis: substrate-attached F-spondin inhibits outgrowth of dissociated motor neurons in an outgrowth assay; F-spondin elicits acute growth cone collapse when applied to cultured motor neurons; and challenging ventral spinal cord explants with aggregates of HEK 293 cells expressing F-spondin, causes contact-repulsion of motor neurites. Structural–functional studies demonstrate that the processed carboxyl-half protein that contains the thrombospondin type 1 repeats is more prominent in inhibiting outgrowth, suggesting that the processing of F-spondin is important for enhancing its inhibitory activity.
One of the most remarkable features of the developing nervous system is the ability of neuronal cells to migrate to distant locations and to innervate distant targets (1). Recent studies have shown that guidance molecules can steer axons and migrating cells either by promoting or inhibiting outgrowth and by cell motility. These outgrowth modulators can act either as short-range cues in the form of membrane-attached or extracellular matrix-bound proteins, or as long-range cues in the form of diffusible molecules. It is the relative balance between attractive and repulsive forces that regulates the directionality of axonal outgrowth and cell migration during development (1).
The ventral midline (the floor plate) plays a key role in patterning the ventral central nervous system by guiding axons to and away from it. The floor plate appears to provide both long-range and local guidance cues that promote or inhibit axonal growth toward and across the ventral midline of the spinal cord, acting as an intermediate target for a number of crossing (commissural) axons (2, 3) and noncrossing (motor) axons (4). These signals are integrated by the extending axons and respond, depending on their identity, by extending either toward or away from the midline. The dual functional role of the floor plate is partly mediated by bipotential molecules. Netrin-1, expressed in the floor plate, can either attract the commissural axons (5, 6) or repel a subset of hindbrain motor neurons (7, 8).
F-spondin, a secreted adhesion molecule expressed in the embryonic floor plate of vertebrates (9–13), and the caudal somite of birds (14), plays a similar dual role in patterning the nervous system. F-spondin promotes adhesion and outgrowth of commissural axons and inhibits adhesion of neural crest cells. Consistent with this, in vivo perturbation experiments demonstrate that F-spondin is required both for accurate guidance of commissural axons at the floor plate (13) and for restriction of migration of neural crest cells (14). Blocking of the endogenous F-spondin in ovo causes: (i) lateral drifting of commissural axons at the contralateral floor plate boundary; (ii) neural crest cell migration into otherwise inhibitory domains such as the caudal somite, the ventral medial sclerotome, and the dermomytome; and (iii) widening of the ventral root at the sclerotome level. This last result suggests that somite-derived F-spondin is a contact-repulsion molecule for motor axons and raises the possibility that the floor plate derived F-spondin is participating in repelling motor axons away from the midline spinal cord.
In the current study, we demonstrate that outgrowth of embryonic motor axons is inhibited by F-spondin protein. Three independent lines of evidence support our hypothesis: substrate-attached F-spondin inhibits outgrowth of dissociated motor neurons in an outgrowth assay; F-spondin elicits acute growth cone collapse when applied to cultured motor neurons; and challenging ventral spinal cord explants with aggregates of HEK 293 cells, expressing F-spondin, causes contact-repulsion of motor neuritis. Hence, F-spondin is likely to play a dual role in patterning axonal trajectory in the spinal cord by promoting outgrowth of commissural neurons and inhibiting outgrowth of motor neurons.
Materials and Methods
DNA Constructs.
The F-spondin protein was extracted from HEK 293 cells expressing the pSecF-spo-His plasmid (15). The thrombospondin type 1 repeats (TSR) protein was extracted from HEK 293 cells expressing the pSec1–5a-His plasmid (13). The reespo-His protein was extracted from HEK 293 cells expressing the pSecTS0-His plasmid (13).
To generate plasmids containing the TSR repeats, PCR fragments containing these regions were constructed by using the primer GAGAGATCTTGCATCTACTCCAACTGGTC (encoding amino acids 443–449) and a set of primers GAAACCTGCATCTACTCCAA (for TSR1, encoding amino acids 500–506); AGGAGAGCACTCCTCGTTGA (for TSR1–2, encoding amino acids 557–563); GGTATGGCACTCAGGCATCA (for TSR1–3, encoding amino acids 612–618); GGGGCACTCTGGCAGCATAC (for TS1–4, encoding amino acids 666–672); and GAACTGCTCTCCATCTGACT (for TS1–5, encoding amino acids 722–728). The PCR fragments were subcloned into BamHI and XbaI sites of pSecTagB (Invitrogen).
Neurite Outgrowth Assays.
The dorsal and ventral parts of embryonic day 13 (E13) rat spinal cords were dissected and cultured as described (13). F-spondin protein and the domain proteins, all equipped with 6 His residues at their carboxyl terminus, were affinity-purified on a Talon affinity column (CLONTECH). Affinity-purified proteins were absorbed onto poly(dl-lysine)-coated wells. The E13 spinal cord neurons were plated on immobilized protein substrates and grown for 40 h. Cultures were then fixed in 4% paraformaldehyde, permeabilized with 0.1% Triton X-100, and stained by using either anti-tubulin III mAb (TUJ-1), anti-TAG-1 mAb 1C12 (16), or the anti-DCC mAb (17), which can recognize proteins expressed specifically by commissure axons. Neurite lengths were measured as the distance from the edge of the soma (sharply defined by the obtained fluorescence) to the tip of its longest neurite. Neurite lengths were measured only if the entire length of neurite could be unambiguously identified.
Growth Cone Collapse Assay.
E4.5 chick ventral spinal cord neurons were dissociated and cultured in 35-mm dishes on poly(dl-lysine) and laminin substrate for 24 h. Culture showing substantial growth, but with few migratory nonneural cells, were selected for growth cone collapse assay as described (18, 19). Then 25–100 μl of conditioned media of transfected HEK 293 was added directly to the cultures (1/25 and 1/10 dilution) for 1 h. Cultures were fixed with 4% paraformaldehyde and stained with anti-erm antibodies (Abs). Only axon termini that made no contact with other axons or cells were included in the analysis. Growth cones were photographed and analyzed with NIH image software.
Motor Neuron Repulsion Assay.
Dissection and culture of E11 rat spinal cord was performed essentially as described (20), except that explants and HEK 293 cell aggregates were cultured in a 50:50 mix of rat tail collagen and Matrigel (Collaborative Biomedical Products, Bedford, MA) and were cultured for 30 h. Cultures were fixed in 4% paraformaldehyde and stained with mAb 2H3 against neurofilament and an anti-mouse IgG indocarbocyanine (Cy3)-conjugated secondary Ab in PBS/0.1% Triton X-100/1% heat-inactivated normal goat serum. The cells' nuclei were stained with propidium iodide.
Results
F-Spondin Is Deposited in the Spinal Cord Neuroepithelium.
F-spondin mRNA and protein are expressed in regions that are inhibitory for neural crest migration and motor axon elongation (14). In birds, F-spondin protein is deposited in the caudal somite and the ventral medial scelrotome (14). In both rodents and birds, F-spondin is expressed in the floor plate (9, 13). Immunohistochemical studies demonstrate that there is a difference in the proteins deposition along the caudal-rostral axis of the central nervous system. At the spinal cord level, the protein accumulates between the floor plate cells and the meninges (Fig. 1A). In the rostral part of the central nervous system, the midbrain and the hindbrain, F-spondin immunoreactivity was evident in the extracellular matrix underlying the floor plate and, in addition, in the neuroepithelium lateral to the floor plate (13) (Fig. 1B). It is conceivable that the Ab recognizes only the regions where high levels of protein accumulate and thus F-spondin is better detected in the rostral central nervous system, where expression levels are higher than in the trunk region. To study whether F-spondin protein is deposited in neuroepithelium at the spinal cord level, we examined the protein levels by Western blotting. Rat embryonic spinal cords (omitting the meninges) were dissected and subjected to Western blotting analysis. A major band of ≈60 kDa was detected with R8 (Fig. 2A, anti-reelin domain Ab) in spinal cord extract of E11–E14 (Fig. 2B). The R3 Ab (Fig. 2A, anti-TSR domain) detected two bands of 40 kDa and 50 kDa (Fig. 2C). Similar size proteins were detected in a whole spinal cord extract (with the meninges) of chick (13) and rat (not shown). Thus, the processed fragments of F-spondin are present at the neuroepithelium. Judging by the localization of the protein in the hindbrain, it is reasonable to assume that the protein is deposited laterally to the floor plate, along the entire neural tube.
Figure 1.
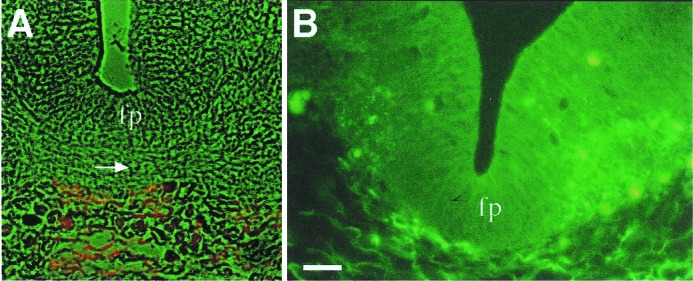
Immunolocalization of F-spondin protein in E13 rat embryo. Sections were labeled with the anti-reespo domain Ab R5 (15) [tetramethylrhodamine B isothiocyanate (TRITC) in A and FITC in B]. (A) Cross section at the spinal cord level. F-spondin protein is localized in the basal membrane that underlies the floor plate (fp). Note that the crossing fibers of the commissural axons (arrow) are in contact with the F-spondin protein. (B) Cross section at the midbrain level. F-spondin protein is localized at the extracellular matrix that underlies the brain, as well as in the neuroepithelium lateral to the floor plate. (Scale bar = 25 μm; A is at the same magnification.)
Figure 2.

F-spondin processed fragments are deposited in the spinal cord neuroepithelium. (A) Schematic representation of the F-spondin domain structure and the Abs used in the Western blot. The black box represents the signal sequence; the gray box, the reelin domain; the dotted box, the spondin domain, and the hatched boxes, the TSR repeats. The broken lines represent the regions used to generate the Abs. (B) Western blot analysis of the F-spondin protein with the R8 Ab. Equal amounts of protein (4 μg) were loaded onto SDS/PAGE. A 60-kDa protein, corresponding to the reelin/spondin domain, is detected in protein extract of E11–E14 rat spinal cords. (C) Western blot analysis of the F-spondin protein with the R3 Ab. The 50-kDa and 40-kDa proteins, corresponding to the TSR domain, are detected within protein extracts E12 and E13 rat spinal cords.
F-Spondin Promotes Outgrowth of Commissural Axons but Not Motor Axons.
We have previously demonstrated that blocking the endogenous somite-derived F-spondin in the quail embryo, leads to an expansion of the ventral root (14). Motor axons that normally avoid the caudal somite are capable of extending through this inhibitory domain when F-spondin activity is blocked. The floor plate-derived F-spondin may act in a similar manner, repelling motor axons away from the ventral midline of the spinal cord. To test whether F-spondin inhibits outgrowth of motor axons, we analyzed the outgrowth-promoting activity of F-spondin and its domains. Purified F-spondin and F-spondin domain proteins were immobilized on poly(dl-lysine)-coated plates. Laminin was used as a control. E13 rat ventral and dorsal spinal neurons were dissociated and plated on the substrate. More than 90% of the dorsal spinal cord neurons expressed the commissural axon cell surface molecules TAG-1 (16) and DCC (17). Also, >90% of the ventral spinal cord neurons expressed the motor neuron nuclear factor isl1 (21) (data not shown). F-spondin, the TSR protein domain (Fig. 3B), and the reespo protein domain (data not shown) did not promote adhesion and outgrowth of motor axons.
Figure 3.

F-spondin proteins inhibit outgrowth of motor axons. (A) Outgrowth of rat E13 commissural neurons on a substrate of laminin, F-spondin, and TSR. Purified proteins (40 μg/ml) were immobilized on poly(dl-lysine). The neurons were stained with the anti-tubulin III mAb (TUJ-1). For each TUJ-1-positive neuron, the neurite length was measured, or, if no neurite was seen, considered to be zero. The percentage of neurons (ordinate) with neurites longer than a given length (in μm; abscissa) is presented. All of the proteins promote outgrowth to a similar extent. (B) Outgrowth of rat E13 motor neurons on a substrate of laminin F-spondin and TSR. The neurons were stained with the anti-neurofilament mAb 2H3. Laminin promotes outgrowth, whereas F-spondin and the TSR domain protein do not. (C) Outgrowth of rat E13 motor neurons on a substrate of laminin (40 μg), laminin + F-spondin (40 μg), and laminin + TSR domain (40 μg). F-spondin and the TSR domain proteins inhibit the outgrowth of motor axons on laminin. (D) The inhibition of the TSR domain is dose-dependent. Then 40, 20, and 10 μg of the TSR domain protein were added to 40 μg of laminin. The degree of inhibition correlates to the concentration of TSR domain protein because 10 μg of TSR protein is less inhibitory than 20 μg and 40 μg. (E) Outgrowth of rat E13 motor neurons on a substrate of laminin (40 μg) and laminin + reespo (40 μg). The reespo domain protein inhibits the outgrowth of motor axons on laminin. (F) The inhibitory effect of the TSR domain protein is not dependent on the number of TSR repeats. Proteins (40 μg) containing 1 repeat, 2 repeats (1–2), 4 repeats (1–4), or 5 repeats (1–5) were mixed with 40 μg of laminin and immobilized on poly(dl-lysine)-coated plates. The outgrowth of motor neurites on the various mixed substrates is inhibited in a similar manner.
There was a difference in the extent of adhesion of motor and commissural neurons on laminin. Approximately twice as many commissural neurons as motor neurons adhered to laminin (data not shown). In addition, only 60% of the motor neurons, as opposed to close to 100% of commissural neurons, that adhered to laminin extended neurites. Nevertheless, the extent of outgrowth of the neurite-bearing neurons, of both cell populations on laminin was similar with 50% of the neurite-bearing neurons extending axons longer than 50 μm (Fig. 3 A and B). This difference might be attributed to different adhesion properties of the cells. Whereas commissural neurons adhered and projected neurites on F-spondin and the TSR domain to a similar extent as they did on laminin (Fig. 3A), motor neurons projected neurites only on laminin.
F-Spondin Is Inhibitory for Motor Neuron Outgrowth.
To determine whether F-spondin actively inhibits motor axon outgrowth, we generated mixed substrates of laminin and F-spondin. Additions of TSR domain protein to laminin abolished the outgrowth of motor axons (Fig. 3C). The effect of the TSR protein was dose-dependent (Fig. 3D). The full-length F-spondin (Fig. 3C) and the reespo protein domain (Fig. 3E) also inhibited the outgrowth-promoting activity of laminin, but to a lesser extent. Thus, it can be concluded that the TSR domain of F-spondin is a potent inhibitor of motor axon outgrowth.
The inhibitory activity of the TSR domain might rely on a combination of multiple inhibitory domains. It is conceivable that each TSR domain has an inhibitory activity. The combination of several TSR domains could exacerbate the inhibiting activity through either an additive or a synergistic effect. Alternatively, one TSR repeat might be sufficient to inhibit outgrowth. To differentiate between these two possibilities, we generated deletion constructs that contained five repeats (TSR1–5), four repeats (TSR1–4), two repeats (TSR1–2), or one repeat (TSR1). The deletion proteins were mixed with laminin and immobilized on poly(dl-lysine). Motor axon outgrowth was inhibited to a similar extent on all of the deletion proteins (Fig. 3F). Thus, each TSR domain had the capacity to inhibit outgrowth of motor axons. It is possible to conclude that only the first repeat is required for the inhibition because this repeat is present in each construct. However, outgrowth assays with other isolated TSR domains do not support this theory (data not shown).
F-Spondin Can Act Directly on Growth Cones.
We next tested whether F-spondin can act directly on the growth cones of motor axons. Chick E4.5 ventral spinal cord neurons were grown overnight on laminin substrate, to permit neurite extension, exposed to supernatant from F-spondin, TSR, or mock-transfected HEK 293 cells for 1 h, and then fixed. Neurons were labeled with anti-erm (22) to reveal growth cone morphology.
In cultures exposed to conditioned media of mock-transfected HEK 293 cells (Fig. 4 A and B), >75% of the neurons had growth cones (growth cone defined as larger than 25 μm2) with an average area of 40.8 ± 22.7 μm2 (Fig. 4E). In contrast, after exposure to 0.1 and 0.5 μg/ml F-spondin (Fig. 4 C and D) or the TSR domain protein, only 20% or 15%, respectively, of the neurites had growth cones, with an average area of 16.8 ± 12.5 and 15 ± 9 μm2 (Fig. 4E).
Figure 4.

F-spondin causes collapse of chick ventral spinal cord growth cones. Ventral spinal cord neurons of E4.5 chicks were dissociated and plated on poly(dl-lysine)- and laminin-coated 35-mm dishes overnight. Conditioned medium of HEK 293 cells was added for 1 h. The amount of proteins was measured by comparison to purified F-spondin in Western blotting. (A and B) Growth cones after exposure to mock-transfected conditioned media and stained with anti-erm mAb. (Scale bar = 10 μm.) (C and D) Growth cones after exposure to F-spondin-transfected conditioned media. (E) Box plot analysis. Each dot represents the area occupied by the growth cones. The boxes enclose 50% of the results; the horizontal lines above and under the boxes enclose 90% of the results. The horizontal lines within the boxes are the medians. A comparison of the growth cones area obtained from the TSR (n = 48) and F-spondin (n = 50) with the control group; mock-transfected conditioned media (n = 43) shows a significant difference (P value < 0.05). This comparison was applied by using Dunnett's method (34).
This result suggests that the collapse of growth cones in this experiment was evoked by F-spondin. Thus F-spondin is likely to act directly on motor axons to elicit rapid change in cytoskeletal organization and growth cone collapse.
F-Spondin Is a Contact Repellent Molecule for Motor Axons.
We have previously shown that F-spondin is required locally at the floor plate to attract the commissural axons to the contralateral floor plate boundary (13). Thus, it can be postulated that F-spondin is a short-range contact attractant for commissural axons. The outgrowth inhibitory activity of F-spondin on motor axons might be mediated by either a short-range contact-repellent activity or a long-range chemorepellent activity. To distinguish between these two mechanisms we used the three-dimensional collagen assay. F-spondin expressing aggregates of HEK 293 cells were cocultured either in contact or at a distance from explants of ventral spinal cord from E11 rat embryos. Tissues were cultured in a matrix consisting of collagen and Matrigel (a partially purified extracellular matrix molecule containing collagen and laminin). Motor axons grew out of the explant profusely. When ventral explants were cultured at a distance (100–200 μm) from the aggregates, motor axons had approximately equal outgrowth on the proximal side (facing the aggregates) and the distal side (opposite the aggregates) (data not shown). There was no apparent difference among mock-transfected HEK 293 cells, and F-spondin, TSR, and reespo-transfected cells. When ventral explants were cultured in contact with the expressing aggregates, a clear inhibition of motor axon outgrowth was observed (Fig. 5B). Motor axons completely avoided growing into aggregates expressing the TSR domain protein. Axons did grow along the boundary of the aggregates, but they did not invade it. This behavior suggests that the TSR domain of F-spondin is a contact-repellent molecule for motor axons. In a control experiment, motor axons grew over mock-transfected aggregates (Fig. 5A). The reespo protein had a moderate inhibitory activity (Fig. 5C) whereas the full-length F-spondin did not inhibit motor axon outgrowth (Fig. 5D).
Figure 5.

E11 ventral spinal cord explants were cultured in contact with F-spondin or F-spondin domain-protein transfected HEK 293 cells, as well as mock-transfected HEK 293 cells, for 30 h in a three-dimensional collagen gel matrix and then stained with anti-neurofilament mAb 2H3 (FITC) to visualize axons. At this stage, only motor axons are expected to be found in these explants (20). The cell bodies were stained with propidium iodide. The broken white line demarcates the boundaries of the HEK 293 aggregate. (A) Mock-transfected cells. The neurites grow over the explant. (B) TSR-transfected cells. The neurites extend along the explant boundary but do not invade it. (C) Reespo transfected cells. The neurites extend along the explant boundary but do not invade it. (D) F-spondin-transfected cells. The neurites grow over the explant. (Scale bar in C = 50 μm for A–D.) (E) Quantitative analysis of the observed changes. NIH image software was used to measure the area occupied by the axons over the cell aggregate (aggregate outgrowth) and the total area occupied by axons. A comparison of the percentage of the aggregate outgrowth between the various proteins with the control group (mock-transfected) is presented. Applying Dunnett's method (34) on the ranks shows significant differences among the TSR, reespo, and the control group (mock) but no difference between the F-spondin and the control group under a significant level of α = 0.05.
Cultures from representative experiments were stained with the anti-neurofilament 2H3 mAb to label axons. The extent of outgrowth on the aggregates of expressing cells was calculated as the ratio between the outgrowth over the aggregates and the total outgrowth. The outgrowth was measured as the area occupied by the neurites (Fig. 5E). In the mock-transfected experiment, an average of 14.4 ± 2.6% (n = 9) outgrowth was scored on the aggregates. The average outgrowth on TSR and reespo aggregates was only 2.74 ± 2.2% (n = 9) and 7 ± 2.1% (n = 10), respectively. Both are statistically different from the mock-transfected assay. The average extent of outgrowth on the F-spondin-transfected cells, 14.3 ± 2.6% (n = 7), was not statistically different from the mock-transfected. Thus, it can be concluded from this experiment that both the TSR and the reespo domains of F-spondin are contact-repellent molecules for motor axons. The extent of repulsion correlates with the inhibitory activity detected in the outgrowth assay, where the TRS domain is more inhibitory than the reespo domain.
The failure of the F-spondin full-length protein to repel motor axons in the collagen assay might indicate that cleavage of the protein is required to reinforce its inhibitory activity. The full-length protein may possess a weaker repulsive activity sufficient enough to overcome the outgrowth promoting activity of laminin in the outgrowth and the collapse assays, but less effective when confronted with substrate containing collagen and Matrigel.
Discussion
F-spondin is expressed in the embryo along the intermediate axonal pathway at the floor plate (9, 13) and the endoneurium of the peripheral nerve (15), and in regions that inhibit cell migration and motor axon outgrowth at the caudal somite and the floor plate (9, 14, 15). In the adult, F-spondin is expressed in regions with active axogenesis: the hippocampus (23) and the distal stump of regenerating axotomized peripheral nerves (15). In accordance with its expression pattern, F-spondin promotes outgrowth of commissural axons that extend along the floor plate; promotes outgrowth of embryonic sensory axons, which form part of the peripheral nerve, and promotes outgrowth of hippocampal neurons and inhibits attachment and migration of neural crest cells (13–15, 23). In the current study, we have demonstrated that F-spondin also inhibits outgrowth of motor neurons. This result is in concurrence with our previous observation that blocking endogenous somite-derived F-spondin activity in the avian embryo leads to an extension of motor axons over an otherwise inhibitory domain: the caudal half somite (14). The erroneous trajectory of the motor axons was evident after using either anti-TSR domain Abs or anti-reespo domain Abs (14). Similarly, in the current study we have demonstrated that the TSR domain protein and the reespo domain protein inhibits outgrowth in both the outgrowth and collagen assay. Analogously to motor neurons, neural crest cells were shown to migrate into otherwise inhibitory domains after blockage of both the TSR and reespo domains (14).
Growth Cone Responds to F-Spondin Directly.
The ability of factors to evoke the morphological changes associated with growth cone collapse appears to correlate with axon-reorienting ability (24). The direct and rapid action of F-spondin on growth cones in vitro supports the possibility that it acts as a repellent to guide motor axons. However, our data do not role out the possibility that F-spondin is sequestering an intrinsic motor axons outgrowth-promoting molecule.
Vermeren et al. (25) have used F-spondin in a similar collapse assay. In contrast to our results, they demonstrated that conditioned media of recombinant F-spondin expressed in COS cells does not induce growth cone collapse of chick sensory, motor, and retinal axons. It is conceivable that the amount of F-spondin protein used by Vermeren et al. (25) was not sufficient to evoke growth cone collapse.
F-Spondin May Play a Dual Role in Controlling Peripheral Nerve Extension.
We have previously proposed that the Schwann cell-derived F-spondin promotes the outgrowth of the peripheral nerve during embryogenesis and during peripheral nerve regeneration (15). In support for this hypothesis, application of F-spondin neutralizing Abs to a cryostat section of axotomized sciatic nerve significantly inhibits the outgrowth promoting potential of the Sciatic nerve distal stump as determined by outgrowth of cultured dorsal root ganglion (DRG) neurons (15). The presence of motor axons in the peripheral nerve poses a potential dilemma. How can motor axons elongate during development and regeneration in an F-spondin-rich milieu? One possibility is that they might change their responsiveness to F-spondin during development. As axons start to extend from the spinal cord, they might be repelled by F-spondin, and as they encounter Schwann cells, they might alter their responsiveness and become indifferent or even attracted to F-spondin. In a similar fashion, it was demonstrated that Sema3A repels rat E14 nerve growth factor-dependent DRG neurons, whereas E18 neurons are not repelled (26). In addition, it was demonstrated that the attraction of commissural axons to Netrin-1 before reaching the floor plate is silenced as they reach the floor plate (27), and the repulsion of commissural axons by Slit and Semaphorin proteins is elevated only after they have crossed the floor plate (28, 29). This behavior may be explained by the use of different receptor complexes. The presentation of a receptor that mediates attraction to F-spondin might be up-regulate on motor axons as they extend out of the spinal cord, as it was demonstrated for the expression of the robo receptor on the longitudinal but not the transverse track of Drosophila commissural axons (28). Alternatively, different forms of F-spondin may be deposited in the repulsive as opposed to the attractive substrate. In support of this hypothesis, analysis of the protein extract of a distal stump of an axotomized sciatic nerves revealed high levels of the full-length F-spondin and the reespo domain protein, whereas the TSR domain protein was undetected (15). It is conceivable that the TSR domain is not anchored tightly to the endoneurium of the distal stump. Thus, the regenerating fibers do not encounter the most prominent repulsive domain, the TSR fragment, but rather the full-length. It is yet to be demonstrated which protein fragments are present along the peripheral nerve during embryogenesis.
F-Spondin Is Acting as a Contact-Repellent Molecule.
Several ventral midline molecules are implicated in motor axon repulsion: Netrin-1, expressed at the floor plate, acts both as an attractant for the commissural (5, 6) and as a repellent for a subset of hindbrain and midbrain motor neurons (7). Sema3A, expressed at the ventral neural tube and the caudal somite halves, repels nerve growth factor-dependent sensory neurons (26), subpopulations of hindbrain motor neurons (8), and migrating neural crest cells (30). Slit-2, expressed at the floor plate, roof plate, ventral spinal cord and motor neurons, repels motor axons and mediates branching of sensory axons (31, 32). The collagen assay results indicate that F-spondin can repel spinal motor axons in a contact-dependent manner in vitro. However, it still remains to be determined whether F-spondin proteins affect motor axon trajectories in a contact-dependent or -independent manner in vivo. In our in vitro assay, the repulsive effect of F-spondin appeared only when the cells were placed in contact with the explants. Furthermore, motor axons elongated along the cell aggregate but did not penetrate it. Thus, F spondin behaves in a profoundly different way to other repulsive molecules such as Netrin-1 and Sema3A, which repel motor axons from a distance (7, 8). There is, however, some resemblance between F-spondin and Slit-2 activities in the collagen assay. Both molecules require contact with the explant to elicit their repulsive activities (31). Nevertheless, motor axons elongate away from Slit-2-expressing cells, rather than avoiding contact with cells expressing F-spondin. In the case of commissural axon guidance, the action of Slit-2 and F-spondin also appears to be contact-mediated, although Slit-2 is repulsive whereas F-spondin is attractive (13, 33).
Acknowledgments
We thank Chaya Kalcheim and Anat Debby-Brufman for suggestions and insights, and Marc Tessier-Lavigne, David Tannahill, Chaya Kalcheim, and Joel Yisraeli for comments on the manuscript. This work was supported by grants to A.K. from the Israel USA Binational Foundation, the Israel Science Foundation, and the Human Frontier Science Foundation.
Abbreviations
- TSR
thrombospondin type 1 repeats
- En
embryonic day n
References
- 1.Tessier-Lavigne M, Goodman C S. Science. 1996;274:1123–1133. doi: 10.1126/science.274.5290.1123. [DOI] [PubMed] [Google Scholar]
- 2.Placzek M, Tessier-Lavigne M, Jessell T M, Dodd J. Development (Cambridge, UK) 1990;110:19–30. doi: 10.1242/dev.110.1.19. [DOI] [PubMed] [Google Scholar]
- 3.Tessier-Lavigne M, Placzek M, Lumsden A G S, Dodd J, Jessell T M. Nature (London) 1988;336:775–778. doi: 10.1038/336775a0. [DOI] [PubMed] [Google Scholar]
- 4.Guthrie S, Pini A. Neuron. 1995;14:1117–1130. doi: 10.1016/0896-6273(95)90260-0. [DOI] [PubMed] [Google Scholar]
- 5.Serafini T, Kennedy T E, Galko M, Mirzayan C, Jessell T M, Tessier-Lavigne M. Cell. 1994;78:409–424. doi: 10.1016/0092-8674(94)90420-0. [DOI] [PubMed] [Google Scholar]
- 6.Kennedy T E, Serafini T, de la Torre J R, Tessier-Lavigne M. Cell. 1994;78:425–435. doi: 10.1016/0092-8674(94)90421-9. [DOI] [PubMed] [Google Scholar]
- 7.Colamarino S A, Tessier-Lavigne M. Cell. 1995;81:621–629. doi: 10.1016/0092-8674(95)90083-7. [DOI] [PubMed] [Google Scholar]
- 8.Varela-Echavarria A, Tucker A, Puschel A W, Guthrie S. Neuron. 1997;18:193–207. doi: 10.1016/s0896-6273(00)80261-5. [DOI] [PubMed] [Google Scholar]
- 9.Klar A, Baldassare M, Jessell T M. Cell. 1992;69:95–110. doi: 10.1016/0092-8674(92)90121-r. [DOI] [PubMed] [Google Scholar]
- 10.Klar A, Jessell T M, Altaba A R I. Cold Spring Harbor Symp Quant Biol. 1992;57:473–482. doi: 10.1101/sqb.1992.057.01.052. [DOI] [PubMed] [Google Scholar]
- 11.Ruiz i Altaba A, Cox C, Jessell T M, Klar A. Proc Natl Acad Sci USA. 1993;90:8268–8272. doi: 10.1073/pnas.90.17.8268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Higashijima S, Nose A, Eguchi G, Hotta Y, Okamoto H. Dev Biol. 1997;192:211–227. doi: 10.1006/dbio.1997.8760. [DOI] [PubMed] [Google Scholar]
- 13.Burstyn-Cohen T, Tzarfaty V, Frumkin A, Feinstein Y, Stoeckli E, Klar A. Neuron. 1999;23:233–246. doi: 10.1016/s0896-6273(00)80776-x. [DOI] [PubMed] [Google Scholar]
- 14.Debby-Brafman A, Burstyn-Cohen T, Klar A, Kalcheim C. Neuron. 1999;22:475–488. doi: 10.1016/s0896-6273(00)80703-5. [DOI] [PubMed] [Google Scholar]
- 15.Burstyn-Cohen T, Frumkin A, Xu Y T, Scherer S S, Klar A. J Neurosci. 1998;18:8875–8885. doi: 10.1523/JNEUROSCI.18-21-08875.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Dodd J, Morton S B, Karagogeos D, Yamamoto M, Jessell T M. Neuron. 1988;1:105–116. doi: 10.1016/0896-6273(88)90194-8. [DOI] [PubMed] [Google Scholar]
- 17.Keino-Masu K, Masu M, Hinck L, Leonardo E D, Chan S S, Culotti J G, Tessier-Lavigne M. Cell. 1996;87:175–185. doi: 10.1016/s0092-8674(00)81336-7. [DOI] [PubMed] [Google Scholar]
- 18.Davis C A, Cook G M W, Stern C D, Keynes R J. Neuron. 1990;2:11–20. doi: 10.1016/0896-6273(90)90439-m. [DOI] [PubMed] [Google Scholar]
- 19.Raper J A, Kapthammer J P. Neuron. 1990;2:21–29. doi: 10.1016/0896-6273(90)90440-q. [DOI] [PubMed] [Google Scholar]
- 20.Ebens A, Brose K, Leonardo E D, Hanson M G, Jr, Bladt F, Birchmeier C, Barres B A, Tessier-Lavigne M. Neuron. 1996;17:1157–1172. doi: 10.1016/s0896-6273(00)80247-0. [DOI] [PubMed] [Google Scholar]
- 21.Ericson J, Thor S, Edlund T, Jessell T M, Yamada T. Science. 1992;256:1555–1560. doi: 10.1126/science.1350865. [DOI] [PubMed] [Google Scholar]
- 22.Birgbauer E, Dinsmore J H, Winckler M, Lander A D, Solomon F. J Neurosci Res. 1991;30:232–241. doi: 10.1002/jnr.490300124. [DOI] [PubMed] [Google Scholar]
- 23.Feinstein Y, Borrell V, Garcia C, Burstyn-Cohen T, Tzarfaty V, Frumkin A, Nose A, Okamoto H, Higashijima S, Soriano A, Klar A. Development (Cambridge, UK) 1999;126:3637–3648. doi: 10.1242/dev.126.16.3637. [DOI] [PubMed] [Google Scholar]
- 24.Fan J, Raper J A. Neuron. 1995;14:263–274. doi: 10.1016/0896-6273(95)90284-8. [DOI] [PubMed] [Google Scholar]
- 25.Vermeren M M, Cook G M W, Johnson A R, Keynes R J, Tannahill D. Dev Biol. 2000;225:241–252. doi: 10.1006/dbio.2000.9820. [DOI] [PubMed] [Google Scholar]
- 26.Messersmith E K, Leonardo E D, Shatz C J, Tessier-Lavigne M, Goodman C S, Kolodkin A L. Neuron. 1995;14:949–959. doi: 10.1016/0896-6273(95)90333-x. [DOI] [PubMed] [Google Scholar]
- 27.Shirasaki R, Katsumata R, Murakami F. Science. 1998;279:105–107. doi: 10.1126/science.279.5347.105. [DOI] [PubMed] [Google Scholar]
- 28.Kidd T, Brose K, Mitchell K J, Fetter R D, Tessier-Lavigne M, Goodman C S, Tear G. Cell. 1998;92:205–215. doi: 10.1016/s0092-8674(00)80915-0. [DOI] [PubMed] [Google Scholar]
- 29.Zou Y, Stoeckli E, Chen H, Tessier-Lavigne M. Cell. 2000;102:363–375. doi: 10.1016/s0092-8674(00)00041-6. [DOI] [PubMed] [Google Scholar]
- 30.Eickholt B J, Mackenzie S L, Graham A, Walsh F S, Doherty P. Development (Cambridge, UK) 1999;126:2181–2189. doi: 10.1242/dev.126.10.2181. [DOI] [PubMed] [Google Scholar]
- 31.Brose B, Bland K S, Wang K H, Arnott D, Henzel W, Goodman C S, Tessier-Lavigne M, Kidd T. Cell. 1999;96:795–806. doi: 10.1016/s0092-8674(00)80590-5. [DOI] [PubMed] [Google Scholar]
- 32.Wang K, Brose K, Arnott D, Kidd T, Goodman C S, Henzel W, Tessier-Lavigne M. Cell. 1999;96:771–784. doi: 10.1016/s0092-8674(00)80588-7. [DOI] [PubMed] [Google Scholar]
- 33.Kidd T, Bland K S, Goodman C S. Cell. 1999;96:785–794. doi: 10.1016/s0092-8674(00)80589-9. [DOI] [PubMed] [Google Scholar]
- 34.Dunnett C W. J Am Stat Assoc. 1955;50:1096–1121. [Google Scholar]