

Abstract
The presence of new neurons in the adult hippocampus indicates that this structure incorporates new neurons into its circuitry and uses them for some function related to learning and/or related thought processes. Their generation depends on a variety of factors ranging from age to aerobic exercise to sexual behavior to alcohol consumption. However, most of the cells will die unless the animal engages in some kind of effortful learning experience when the cells are about one week of age. If learning does occur, the new cells become incorporated into brain circuits used for learning. In turn, some processes of learning and mental activity appear to depend on their presence. In this review, we discuss the now rather extensive literature showing that new neurons are kept alive by effortful learning, a process that involves concentration in the present moment of experience over some extended period of time. As these thought processes occur, endogenous patterns of rhythmic electrophysiological activity engage the new cells with cell networks that already exist in the hippocampus and at efferent locations. Concurrent and synchronous activity provides a mechanism whereby the new neurons become integrated with the other neurons. This integration allows the present experience to become integrated with memories from the recent past in order to learn and predict when events will occur in the near future. In this way, neurogenesis and learning interact to maintain a fit brain.
Introduction
Thousands of new neurons are added into the adult hippocampus each day. However, most of the new cells do not survive. In fact, over half of them, if not more, die within just a few weeks of their birth. One of the most effective ways to keep these cells from dying is by learning. Animals that learn a new and difficult task retain more of these new cells than animals that do not learn or learn a very easy task [1–4]. Thus, effortful and successful learning keeps new neurons alive. Moreover, once rescued, the new neurons survive for months at least [5]. These new cells establish anatomical connections with other neurons, which not only affect neuronal activity within the hippocampus but also presumably affect synaptic and neuronal activity at afferent locations throughout the brain.
The evidence that new neurons might be influenced by learning and perhaps even involved in learning was not initially well accepted [1, 6, 7]. Of course, we do not fully understand how learning occurs much less which neuroanatomical and neurophysiological processes are necessary. However, prior to the discovery of neurogenesis, it seemed prudent to assume that learning and the consequent establishment of memory would use already existing neurons and then change the synaptic connections between those neurons as a result of learning – at the so-called Hebbian synapse [8]. This was and still is the prevailing notion behind most neurobiological theories of learning [9]. It is probably in large part correct. However, we must now incorporate the idea of new neurons into the system. Are they involved or just casual by-standers? Are they necessary or simply modulators? Why does the brain produce these new neurons and how does it optimize the number of new ones so that they are useful but not disruptive? In this review, we will address these questions and propose a positive feedback system between neurogenesis and learning. Within this system, learning increases the survival of the new neurons in order that they may then be used to learn more efficiently in the future.
How many is enough?
The number of new cells produced in the hippocampus is estimated to be about 10,000 per day in young adult male rats [10] and around 3,500 in older adult male rats [11]. Similar numbers or more are expected in humans [12, 13]. This seems like a rather large number in isolation. However, one rat dentate gyrus (DG) possesses about one million mature granule cells [14]. In this context, several thousand each day is not so many. As they continually accrue, the adult-born new neurons still only represent about 3% of the granule cell layer present at any given time point [15]. These numbers are gross estimates because the actual number of cells produced varies considerably as a function of species and strain and even sex (i.e. gender). Probably the largest factor is age with numbers decreasing by half or more between puberty and adulthood [16]. Still, thousands of new neurons each day is a significant number.
The new cells are amazingly responsive to their environment. More are produced in response to exercise, antidepressant treatment and even sexual activity. Fewer are produced in response to stressful experience, specific drugs of abuse, alcohol and sleep deprivation [17, see for review]. Thus, it would appear that new neurons tend to be produced in response to experiences that are often considered “healthy” whereas fewer are produced in response to unhealthy behaviors. This relationship may be coincidental but it seems unlikely. Even if it is, one could make a good argument for engaging in activities that enhance neurogenesis, if only to encourage positive healthy behaviors in humans.
Use Them or Lose Them
The good news is that new neurons are produced throughout adulthood. The bad news is that most of them do not survive. More than half of the new cells die within just a few weeks of being born. The death of so many new cells seems like a tremendous waste of energy and leaves one wondering about the functional significance of new neurons. Why would they be produced if only to die weeks later? What happens to the ones that do not die and how can more be encouraged to survive? It turns out that many, if not most, of the new neurons can be rescued from death by learning. This is a prime example of “use it or lose it.”
Cells that are already born and present during the learning experience are more likely to survive than cells that are present in an animal that is not learning [1, 2, 18, 19]. The types of training experiences that increase cell survival include associative learning of a trace conditioned eyeblink response as well as spatial navigation learning in which animals learn to find a hidden platform using spatial cues in the environment [1, 17, 18]. Other forms of learning, such as social transmission of food preference or learning a skilled reaching task can also increase the percentage of cells that survive [20, 21]. The surviving cells then mature into fully functional granule cells that integrate into the existing hippocampal neural circuitry [22–24]. It is important to realize that the training experience, per se, does not rescue the cells from death. Instead, it is the learning, which occurs during training that is critical. This distinction between learning versus training has been repeatedly observed in studies using trace conditioning as well as spatial learning. In these studies, animals that successfully acquire the conditioned response show an increase in the number of surviving cells, whereas those that fail to acquire the conditioned response display no such increase [Figure 1; 1, 2, 3, 5, 25].
Figure 1. Successful and effortful learning increases the survival of adult-born hippocampal neurons.

A) Animals that successfully learned the conditioned response (Good Learners) retained significantly more cells than those that failed to learn (Poor Learners), or those that were not trained with paired stimuli (Unpaired). Animals that failed to learn displayed no increase in the number of surviving cells. Thus, learning only increases the survival of new neurons when that learning is successful. B) Of those animals that learned, those that took longer to do so emitted fewer conditioned responses (CRs) over the course of training. However, those animals that took longer to learn retained more new cells than those that learned in relatively few trials. Representative BrdU-labeled cells from the granule cell layer of (C) an animal that successfully learned trace eyeblink conditioning and (D) an animal that failed to learn (Curlik & Shors, 2011).
Interestingly, a recent study showed that spatial learning might affect the rate of maturation, differentiation and also activation of adult-born new cells in the DG differently in different strains of rats [26]. This suggests that there are multiple ways in which experiences shape the structure and function of the dentate gyrus by means of affecting adult-born cells. For example, it might be that slower maturation rate extends the time period during which the new neurons are most susceptible to be recruited into existing neuronal networks. Further studies on this matter are needed.
With that said, learning only increases the survival of new neurons during a critical period in their development. Neurons that are one to two weeks of age at the time of the training experience can be rescued from death. However, learning does not seem to increase the survival of new neurons that are older or younger than this critical period [27, 28]. In fact, it has been reported that learning can actually decrease the likelihood that cells that are produced at the time of the training experience will mature into fully functional neurons [27, 29]. These cells are very young at the time of training – from minutes to just a few days, whereas those that survive are between one and two weeks of age during training and have differentiated into young neurons by the end of training. Thus, at the same time that some cells are being rescued from death by learning, younger cells are dying. This type of modulated response to learning may be used to maintain the total number of granule cells in the DG at an optimal level [27, 29].
Learning with ease
While some forms of learning can increase neuronal survival, others seem to have no such effect. For example, training with delay eyeblink conditioning does not increase the number of neurons that survive [1]. Why is it that the acquisition of trace, but not delay, eyeblink conditioning increases neuronal survival? Until quite recently, we hypothesized that tasks that require an intact hippocampus would rescue new neurons from death whereas tasks that do not depend on the hippocampus would not. Indeed, our initial studies seemed to support this hypothesis, as acquisition of trace, but not delay, conditioning requires an intact hippocampus [30]. Furthermore, acquisition of the hidden platform version of the Morris water maze increases neuronal survival, but learning the hippocampal-independent version of the task with a visible platform does not [1, 25, 31].
In recent years it has been determined that the hippocampal dependence of a task is neither a necessary nor sufficient factor in determining whether the acquisition of that task will increase neuronal survival. For example, learning a hippocampal-independent form of eyeblink conditioning, known as contiguous trace conditioning, does rescue new cells from death [32]. Conversely, learning a trace-conditioning task with a very short trace interval still requires the hippocampus but does not rescue new neurons from death [33]. Given these dissociations, it is clear that hippocampal dependence is not the key factor that determines whether learning will keep new cells alive or let them die.
If hippocampal dependence is not the key factor, what is? Recent evidence indicates that learning will rescue new neurons from death only when learning is difficult. We have operationally defined difficulty as an increase in the number of trials required to reach asymptotic performance. Over the past several years, our laboratory has repeatedly observed strong positive correlations between the number of trials an individual animal requires to learn a task, and the number of surviving neurons in that animal’s dentate gyrus [Figure 2; 2, 3, 32]. Since trace eyeblink conditioning requires many more trials to master than does delay eyeblink conditioning, this theory would correctly predict that trace, but not delay conditioning would increase neuronal survival. Furthermore, recent evidence indicates that when delay conditioning is made more challenging, more of the new neurons are retained [34]. Conversely, when trace conditioning is rendered easier to learn, the new neurons are not rescued from death [33]. In summary, it appears that only tasks that are sufficiently difficult to learn are successful in the rescue. In other words, the organism must exert effort, in this case mental effort, in order to keep the new cells from dying. These findings take the concept behind “use it or lose it” into a slightly more complex and yet more intriguing realm.
Figure 2. Suppressing neurogenesis with an anti-mitotic agent suppresses associative learning of a trace memory but does not suppress spatial learning in the Morris water maze.
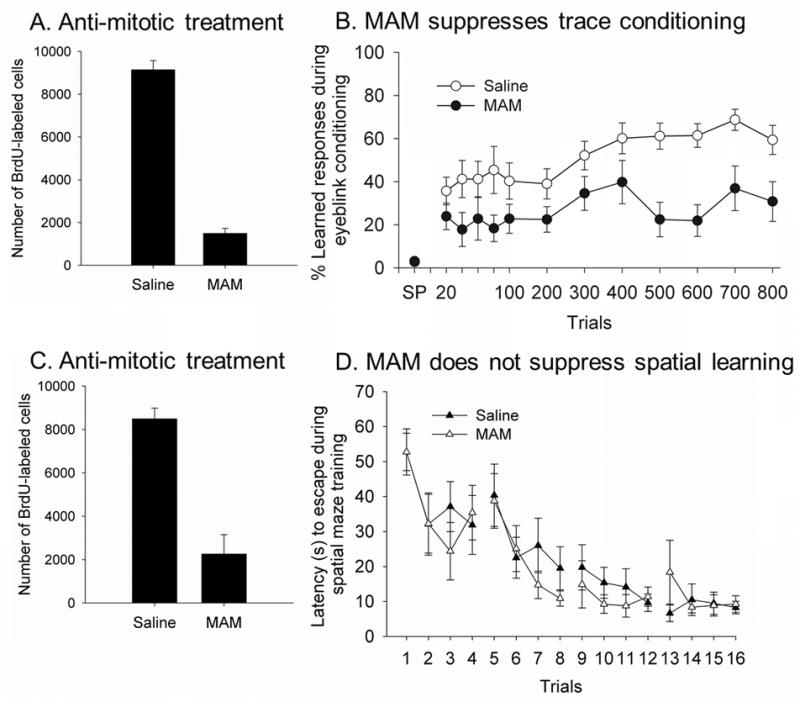
A) Animals treated with the anti-mitotic agent MAM generated very few new neurons and B) most of them were unable to learn the trace memory (Shors et al., 2001). C) Animals treated with the anti-mitotic agent generated very few new neurons but D) readily learned to find the hidden platform using spatial cues in the environment (Shors et al., 2002). Therefore, suppression of neurogenesis is associated with deficits in some but not all types of learning processes.
Currently the mechanism by which learning rescues new neurons is unknown. One potential candidate is gamma-aminobutyric acid (GABA). GABA is a neurotransmitter that is traditionally thought of as having an inhibitory effect; however, GABA activity depolarizes immature adult-born neurons [35]. Furthermore, it has been reported that GABAergic depolarization facilitates the formation of synapses by these new neurons [36]. Therefore, it is possible that learning-induces changes in GABAergic activity which may contribute to the increase in cell survival. This is probably the most parsimonious hypothesis, although difficult to test experimentally because manipulations that alter GABA tend to alter learning.
N-methyl D-aspartate, or NMDA, is another neurotransmitter that modulates the effect of learning on neuronal survival. NMDA receptor activation is necessary for trace eyeblink conditioning [3, 37], as well as most types of spatial navigation learning including the hidden platform version of the Morris water maze [38]. Moreover, survival of two-week old hippocampal neurons requires activation of NMDA receptors [39]. We recently examined the effects of NMDA receptor antagonism on cell survival after learning. Antagonism of these receptors each day before training prevented learning and thereby prevented the increase in cell survival. Antagonism each day after training did not prevent learning but also did not prevent the increase in cell survival [3]. In line with our results, another recent study showed that infusions of an NMDA antagonist during the training period disrupted learning a delayed matching-to-place task and abolished learning-induced changes in neurogenesis [40]. We have also used a drug, d-cycloserine to enhance activity of NMDA receptors during learning [3]. In this case, more animals learned and as a consequence, more cells were rescued from death. Although these data once again associate the increase in cell survival with learning and not training, they do not indicate that NMDA receptor activation is necessary for the increase in cell survival. Clearly, it is important to identify the mechanisms that keep new neurons alive after learning. However, this is no easy task. Most of the likely candidates – GABA, NMDA, BDNF, ACh, etc. – are intimately tied into learning. As such, it is difficult to manipulate or eliminate one without affecting the other. Of course, this is how a unified system works – one component cannot occur without the other. They are positively related to one another and oftentimes feedback on each other.
Using Them
From the moment of their discovery, new neurons were implicated in learning. This was in part because the hippocampus is involved in many processes related to learning and even necessary for a subset of those processes. The hippocampus is most often associated with spatial learning because removal of the hippocampus results in deficits in learning about spatial cues in the environment. The most often used task to assess spatial learning is the Morris water maze task [41]. During training, the animal is placed in a pool of water from which it cannot see a hidden platform. The animal eventually bumps into the platform and then learns to escape onto the platform by visualizing the cues that localize the platform. Animals without a hippocampus cannot learn this escape response and even have difficulty remembering the location of the platform. Since the new neurons are located within the hippocampus, one might predict that they are used to learn about spatial cues. However, many studies imply that they might not. Animals with virtually no new neurons after administration of an antimitotic agent readily learn to find the hidden platform [42]. Similarly, Madsen and colleagues reported no spatial learning deficit in animals with virtually no new neurogenesis after irradiation [43]. This same type of result has been reported using different techniques and protocols to reduce cell number [42–49]. Therefore, it seems that new neurons at the age of around 1 to 2 weeks are not necessary for initial spatial learning – at least as expressed during training on the standard Morris water maze task. However, deficits in spatial memory have been seen when tested after several days or weeks after successful initial training [see for example, 49]. In addition, one study showed subtle deficits in using flexible learning strategies when 4 to 8 weeks old adult-born cells were absent in mice [50], while yet another demonstrated difficulties in a task requiring spatial relational memory [48]. Finally, reducing the functionality of the new adult-born neurons using transgenic manipulations in mice led to deficits in spatial learning in the standard Morris water maze and eight arm radial maze [51]. As is evident, the relationship between adult hippocampal neurogenesis and spatial learning is complex.
In humans, the hippocampus is most often associated with what is referred to as declarative memory – or in some circles, episodic memory [52, 53]. These learning processes are typically verified through verbal recollection and awareness of an experience. For example, humans trained with a classical conditioning task of eyeblink conditioning are more likely to learn the task if they are aware of (or able to consciously recollect) the stimuli, even though the stimuli are separated in time by a trace interval [54]. These types of learning require an animal to maintain a memory of an event over some period of time in the absence of that event and then use that memory to predict the occurrence of a different event in the future. This type of learning, often referred to as trace conditioning, can also be examined in rodent species. In rodents, it has been shown that learning this response keeps new neurons alive that would otherwise die [1, 17, 18].
The obvious next question is: are new neurons necessary to learn a trace memory? This does appear to be the case. In the first study of this sort, animals were treated each day with an antimitotic agent [7]. After two weeks, they generated very few new cells and in the absence of the drug, they had great difficulty learning to associate two stimuli across time. Most animals showed minimal evidence of learning the association. As shown in Figure 2, the variability among treated animals was greater than among those untreated. Therefore, some animals were still able to learn with few new neurons while others did not. Given our data with cell survival and task difficulty, one might propose that some animals – those that can learn with ease – do not depend on the new neurons to learn. Still, other animals are unable to learn with so few new cells. Now one might argue that these animals are unable to learn due to other side effects of the drug and this argument is difficult to refute completely. However, the animals were able to learn just about any other task that we gave them – learning about the context, spatial learning, visible cue learning, simple associations, etc. [42].
The data showing deficits in trace eyeblink conditioning have been substantiated with a different measure of trace conditioning – that of fear. During this task, an animal learns that a conditioned stimulus such as a tone predicts the occurrence of a foot shock later in time, after a temporal gap of about 30 seconds. The animal learns to fear the trace interval, especially the last few seconds during the gap. Trace fear learning just like that of trace eyeblink conditioning requires an intact hippocampus [55]. It should be noted that trace fear conditioning has not been declared “declarative” but there is little reason to think that it would not be – because trace eyeblink conditioning is related to declarative memory and/or thinking in humans [54]. Of course, these distinctions between learning systems are somewhat arbitrary and may not map directly onto neurobiological processes [56]. Certainly, it seems clear that the acquisition of this particular type of information is easily disrupted by a disruption in neurogenesis. When either an antimitotic or irradiation was used to ablate neurogenesis, animals did not express fear of the trace interval [42, 57]. In another study, animals were genetically manipulated to produce virtually no new neurons in their dentate throughout life. As adults, these animals actually expressed more fear during the trace interval. Learning other tasks was unaffected [45]. Thus, a disruption of neurogenesis tends to disrupt trace fear conditioning although some animals express more rather than less fear. From these various reports, one might conclude that neurogenesis is somehow involved in learning to fear an event in the future when the cues associated with the event are no longer present. This would suggest that the cells are somehow being used to integrate experiences in the present moment with those of the recent past to predict events in the near future.
The role of neurogenesis in contextual fear conditioning has also been assessed. Contextual fear conditioning involves the presentation of shocks, usually to the foot through a grid floor, in a specific context. Antimitotic and some irradiation studies indicate that neurogenesis is not necessary to express a learned fear of the context [42, 57–59]. A wide variety of genetic ablation studies have confirmed this conclusion [45, 48, 59–61]; however, some studies provide data that suggest otherwise [44, 62–67]. Given that the same method used to ablate neurogenesis has produced both positive and negative evidence for the argument that contextual fear conditioning depends on the production of new neurons, it seems that differences in conditioning protocols may explain discrepancies in the data. Drew and colleagues found that ablation via irradiation only produced deficits in context fear conditioning with mice if a tone was not present during a single presentation of a shock in a context [66]. Interestingly, the deficit was rescued if a tone preceded the presentation of the shock. In addition, the deficit was rescued if the animals were pre-exposed to the context or if multiple shocks were given. One could argue that the pre-exposure and multiple shocks enabled the shock and context to be more easily associated in this particular situation. If our hypothesis about task difficulty were true, then easier versions of context fear conditioning would not require adult neurogenesis. Snyder and colleagues found that rats had deficits in learning to fear a context while mice had no deficits when training involved multiple tone-shock presentations following irradiation [67]. Given the data, it was suggested that neurogenesis in rats has a more significant role in learning compared to mice likely because more newly made cells survive in the rat. They also found that the learning deficit was only produced if irradiation occurred four or eight but not three weeks before context fear conditioning. However, this contrasts data produced by Anchanta and colleagues that show that 90 days after irradiation, there was no learning deficits in context fear conditioning when multiple tone-shock pairings were used [57]. Obviously, differing species, techniques, protocols and possibly complexities of the contexts complicates the matter. However, given the connection between cell survival and task difficulty one might propose that increased context complexity or reduced context saliency, might make the task more difficult. If so, the neurons may be more likely to be required for discrimination in a spatial environment. Indeed, a recent paper by Tronel and colleagues found that genetic ablation of neurogenesis reduced the ability of mice to discriminate between two similar and complex contexts [68]. Typically, their genetic approach for reducing the number of new neurons produced does not result in deficits in context fear conditioning when training involves a single shock presentation and no tone in mice.
Along these same lines, it has been reported that a reduction in neurogenesis impaired spatial learning when the training conditions rendered the task more difficult to learn. Dupret and colleagues trained transgenic mice with genetically ablated neurogenesis on a task where the platform location was changed every few trials [48]. In wild-type mice, acquisition of this task was retarded suggesting that this task was more difficult to learn compared to the traditional spatial water maze where the hidden location remains unchanged. Transgenic mice were unable to learn this task to the same extent as wild-type mice, even though they were able to perform as well in the traditional spatial maze task. In addition, irradiation directed at the new cells impaired performance in a radial maze task that was difficult to learn [69]. Furthermore, Kempermann and colleagues report that an antimitotic agent did not alter performance in a spatial maze at the time of testing but animals were unable to use information learned for search strategies in the future when the location of the platform was changed [50]. They conclude that new cells provide flexibility to learning processes [see also 48]. Overall, the data to date suggest that tasks that critically engage new neurons in the hippocampus are those that are most challenging for the animal to learn. This would not preclude some tendency for tasks that depend on the hippocampus to be more involved than others because in general tasks that depend on the hippocampus are difficult to learn when compared to those that are not. Obviously, the tasks that depend on neurogenesis in the hippocampus are going to depend on the hippocampus but tasks that depend on the hippocampus would not necessarily depend on neurogenesis in the hippocampus.
Learning to learn
In isolation, the new cells may not seem that distinct from those generated during early gestation and postnatal development [70, 71]. However, there are several key differences. Perhaps most importantly, they mature within a population of mature cells with established connections. As a consequence, they can receive synaptic input almost as soon as they are generated and can therefore do so as they are developing [39, 72, 73]. This input affects their survival and fate. As discussed, learning determines how many of the new cells mature but only if the task is sufficiently difficult to learn and only if animals learn it well [5, 32, 74]. Those animals that fail to learn display no increase in the number of surviving new cells [Figure 1; 1, 2, 3, 5, 19, 25]. When animals take more time to learn, it tends to result in more neurons that survive [2]. It also appears that neurogenesis is required for difficult learning, i.e. tasks that require more persistent activation of the hippocampus. It may be that new neurons are able to integrate their current activity with that of mature neurons to predict events in the future. This would occur when the task is difficult to learn and therefore when more trials are required to learn. Ultimately, this integration produces more complex yet more efficient circuits for learning in the future. This would ease up learning of arduous tasks in the future. These notions are supported by Aimone and colleagues whose data suggest that newly generated neurons serve to integrate information and later as they mature and improve the efficiency of encoding in the future [75].
Learning quickly is of great advantage to the organism. Those animals that quickly learn to find food and avoid predation are more likely to survive to reproduce and pass on their genes. Those with the most diverse set of neurons and synapses may be at an advantage. It should be noted that just because the cells have reached maturity (i.e. stain with NeuN and can produce action potentials), they are not necessarily similar in their responses as mature neurons in the hippocampus. Just as adult organisms differ as a function of chronological age, so do cells. Newly-generated cells that have recently matured are still different from the longer residing neurons. For example, their spines are more mobile [23] and they are more likely to express activity-dependent genes [76]. Interestingly, cells rescued from death by learning a spatial water maze task were preferentially reactivated during re-exposure to the training task [77]. As such, these cells are presumably making fine adjustments to their synapses, which may facilitate learning about similar situations that arise in the future.
Critical periods of time
Adult-born neurons tend to follow the same stages of development, as do cells in the young, immature brain. However, the emergence of synapses requires a bit more time [78]. Adult born cells in the DG extend their axons into the CA3 by the end of the second week of their life and produce dendritic spines and functional synapses with other cells in about 3 weeks [78]. Presumably, these connections are guided by pre-existing synapses within the surrounding local neuronal network in the DG [23]. Most of the new neurons make connections to pyramidal neurons in area CA3 but they can also synapse on interneurons in the hilus, as well as back onto granule cells in the dentate gyrus. When the cells are slightly older – 4–6 weeks of age – long-term potentiation (LTP) is more easily induced and it is of a higher amplitude than that expressed by more mature neurons in the DG [79, 80]. When, and if, the new cells reach the age of 8 weeks, their basic physiological properties have become much like those of mature granule cells.
That said, around one week of age, many of the adult-born new cells start to die through apoptosis. As discussed, adult-born cells in the hippocampus that are 1–2 weeks of age during training are more likely to survive if the animal is trained on a task requiring the formation of associative [27] or spatial memory [28]. In addition, new cells rescued from death by learning at this early stage of maturation are preferentially reactivated during re-exposure to the training task [77]. During this time period the cells are establishing synapses and, more interestingly, play a critical part in some forms of learning [see for example 7] and memory [49]. Training in turn in a spatial memory task increases dendritic growth in 1 to 2 week old cells depending on task complexity [40].
The period of enhanced LTP at 4 to 6 weeks of age probably also makes new granule cells more susceptible to external events and facilitates their recruitment into neuronal networks. In fact, when the new cells are 4 weeks of age or older, they are more likely than mature cells in the dentate gyrus to be recruited into memory circuits by training on a spatial task [76]. The new cells are also more likely to survive if they are exposed to tetanic stimulation sufficient to induce LTP around this same time period [73, 81, 82]. In conclusion, it seems that recruitment of the new dentate granule cells into neuronal networks by exposure to external events occurs primarily when the cells are still immature and starting to form synapses as well as expressing more synaptic plasticity.
Out with the old, in with the new – the role of new neurons in learning
To date, there have been several theories related to how new neurons get incorporated into the existing neuronal networks and participate in processes of learning. Early on, we proposed that they are used to encode the timing of events or the timing of responses [7]. This idea arose in part because the new cells are so responsive to learning the temporal relationship between very closely occurring stimuli: Exposure to a training regime that requires the animal to learn to time its responses within just a few tens of milliseconds in the framework of stimuli that last up to seconds increases the number of new adult-born cells that survive [27]. Moreover, with few of these new cells animals have great difficulties in timing the learned response adaptively and most can not acquire an adaptive conditioned response at all [7]. It should be noted that the idea that neurogenesis may relate to learning about timing actually has its roots in the avian world [see 83]. Nottebohm has long suggested that the new cells are used to time events and perhaps to learn new songs – again a very finely timed motor response.
Theoretically, the idea of new neurons being involved in learning about timing is plausible. New cells are continuously being generated and therefore may simply provide an endogenous template for marking events as they occur in time. One theory suggests that new cells get incorporated into the existing neuronal networks without the older, more mature cells being abolished [75, 84]. In this view, new cells are used to encode events in the present as they become associated with the context. Although increasing the likelihood of interference, such an additive process would be useful in order to link temporally and/or spatially related events together. Interference with existing representations, a clear drawback of this view, could be overcome with selective synaptic plasticity to new neurons [84]. Related to this idea, some have suggested that the new cells are used for pattern separation [for a review on hippocampus as a pattern separator, see 85 for example]. In addition to empirical data, the anatomical connections within the hippocampus are consistent with its putative role in discriminating patterns. For example, the dentate has five to ten times as many neurons as the entorhinal cortex, which feeds sensory and motor information into the hippocampus [86]. In fact, recently it was shown that a reduction in neurogenesis reduced the ability to separate patterns of visual information [69]. If the cells become more engaged as the task becomes more difficult to solve, then one might presume that they would be involved in separating very similar patterns more than separating very different ones.
Another theory about how new cells become incorporated into the existing neuronal networks and participate in processes of learning suggests that the new cells are used to replace existing mature cells in the dentate gyrus [86]. With this idea, the new cells might be used to encode events as they occur without necessarily integrating present information with past information. This idea is support by recent data published by Alme and colleagues [87]. This would require that the DG is only working as an encoding station, not as storage for representations of events and not as a relay station for memory retrieval either. A benefit of this view is that recall of past events would be shielded from interference because different sets of cells would encode events at different times. These ideas are consistent with the time-limited role of the hippocampus in trace conditioning suggested by studies showing that the structure is necessary for initial acquisition but not for long term expression of the memory [88, 89]. For spatial learning, however, it appears that the hippocampus is critically involved also in the long-term expression of memory [90]. Therefore, it may be that the new neurons play a critical role in memory for some aspects of memory retrieval and/or expression. In any case, it seems unlikely that new neurons adhere to some dichotomous view of learning and memory, i.e. learning versus memory, hippocampal-dependent or not, declarative versus procedural, working versus reference, spatial versus nonspatial, conscious versus unconscious, etc.
Communication is key
When new neurons were first discovered, it was necessary to evaluate them exclusively. This approach provided us with lots of information about the cells and their development. However, now that we know as much as we do, it is time to consider them in their context. To process external input, the brain works as a whole, based on spatially distributed but functionally related networks of cell assemblies. One fairly convincing view of how cell assemblies scattered throughout the brain communicate to encode events is through synchronized electrophysiological oscillatory activity [91–94]. Synchrony can be observed both over several cycles as well as within single cycles, for example when faster oscillations are nested within slower oscillations, or when specific cells show firing at a specific phase of another neural oscillation. As a mechanism, oscillatory synchrony enables flexible yet very effective communication: By synchronizing and desynchronizing their activity in relation to each other cell assemblies can make sure that signals are sent and received at an optimal time point leading to maximal effects, excitatory or inhibitory, with very little wasted effort or chances for error.
The hippocampus engages in oscillations that are related to the acquisition of new information. One of the most studied is the theta oscillation that has a frequency of 4–12 Hz [95, for a review see 96]. The relative power of theta predicts learning rate and increases early in learning in animals that initially show a low level of theta and learn slow [97]. Hippocampal theta oscillation synchronizes with theta in the cerebellum [98, 99] especially in response to external stimulation and also modulates oscillatory activity in the neocortex [100, 101]. Synchronized slow oscillations (<1 Hz) common to the hippocampus and the neocortex relate to and may even control the timing and content of communication from the hippocampus to the entorhinal cortex and further to the neocortex [102]. Moreover, high frequency ripples (~200 Hz) in the hippocampus have been implicated in memory consolidation [103, 104]. These events represent the synchronous firing of tens of thousands of neurons along the hippocampo-entorhinal axis [105] and might reflect the replay and consolidation of recently acquired information. Last but not least, gamma oscillations (30–80 Hz) riding on theta waves have been proposed to reflect a mechanism of representing active memories in a neocortical multi-item buffer from which the items would then be stored to hippocampal long-term memory by means of LTP [106]. This theta-gamma –synchrony could be especially useful in encoding ordered sequences of event or items into long-term memory [107, 108].
Due to their increased excitability, immature adult-born granule cells play a crucial part in the synchronous electrophysiological oscillatory activity of the DG. Indeed, their removal either through irradiation or antimitotic agents disrupts ongoing oscillations in the dentate gyrus [109; Nokia & Shors, in preparation]. Namely, abolishing neurogenesis in mice using irradiation induces an increase in the amplitude of spontaneous gamma bursts in the dentate gyrus, as well as increases synchronization of dentate neuron firing to these bursts [109]. Our preliminary data further indicates that immature adult-born neurons contribute to spontaneous as well as stimulus-induced theta-band oscillatory responses in the dentate gyrus. The disruption in neurogenesis and oscillatory activity of the DG is accompanied by a profound deficit in learning a difficult trace conditioning task in which animals must associate stimuli that are separated by a temporal gap [7; Nokia & Shors, in preparation]. They have no difficulty whatsoever learning a simple task in which the stimuli are contiguous in time. Thus, the role of the newly-born cells might be related to learning about the temporal relationship between stimuli as they occur in real time. Obviously, temporal accuracy of cell firing in response to stimuli, and its subsequent fine-tuning is part of the learning process. Indeed, it is critical for a system that learns to predict events at precisely the right time in the future.
Use and misuse
It is of course encouraging to know that our brains continue to produce new neurons throughout our lives. It is even more encouraging to know that we can make more of them by engaging in healthy behaviors and we can keep more of them by engaging in serious mental activity. However, the pendulum can swing the other way – with bad behaviors and minimal mental activity, fewer cells will be produced and even fewer will survive. One or two days or even one or two months may not be enough to affect the overall integrity of the dentate gyrus, much less the entire hippocampus. But day after day, year after year, such a reduction in physical and mental activity would produce a smaller granule cell layer – with fewer axonal connections to cells in CA3 and then in CA1 et cetera. Not only would the anatomical integrity be compromised but presumably so would neuronal activity and synchronicity that normally develops between these cells and the older residing neurons (Figure 3). Such a lifestyle, if maintained over years, would ultimately compromise thought processes that are necessary to integrate the present moment with memories from the past and anticipate events in the future. Unfortunately, a positive feedback system can quickly become one of negative consequence for the organism. Luckily, most humans have their share of good and bad days of thoughts and behaviors, which in the long run probably works out just fine for maintaining a state of mental and physical equilibrium.
Figure 3. A model of integration of adult-born immature neurons into functional networks within the dentate gyrus (DG).
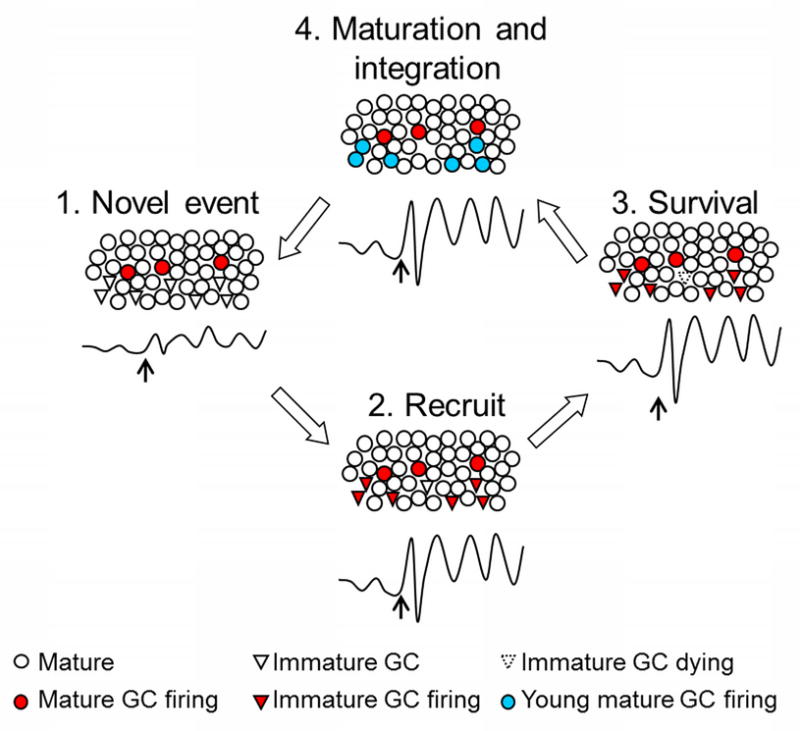
We propose that when a novel event occurs, oscillatory activity, that is, synchronous firing of existing mature neurons in response to that event engages immature neurons to also fire in synchrony. These cells are thus recruited into the network. Immature neurons might be especially receptive because of their heightened excitability. At the level of the local field potential, joint activity of both mature and immature neurons would be manifested as increased synchrony and response magnitude. The new neurons might thereby enhance the quality of the output by improving both short (milliseconds) and long (days, months, years) term temporal coding. Eventually, the new neurons would survive because of their participation in the network activity. Once rescued from death, they would mature into neurons and participate in new learning related to the initial learning experience.
From muscles to memories
The phrase “use it or lose it” is used most often to refer to the relationship between exercise and muscle mass. In this case, the cells themselves become larger after exposure to physical stimulation and strenuous activity. Once enlarged, they integrate more readily with other muscles to form functional muscle circuits. As a consequence, motor activities and skills that were once difficult if not impossible to do can now be accomplished with ease. In this way, the analogy to neurogenesis is perhaps not far-fetched. Once generated, new neurons are stimulated to survive by engaging in neurophysiological activity related to new and challenging learning experiences (Figure 3). New thoughts and memories arise which become integrated with past experience encoded by older more established neurons. As such, these integrated thoughts are represented within dynamic neuronal networks, which use oscillations to mark the present moment and predict how and when events will occur in the future. Critically, once the once-new neurons become assimilated into the network, similar thoughts now arise with ease as new neurons are generated and more complicated thoughts and insights become possible.
Research highlights.
Neurogenesis and learning interact to maintain a fit brain
Learning increases the survival of new neurons if it is difficult to achieve and successful
Learning increases the survival of new neurons only during a critical period in cell maturation
Immature neurons are needed for some types of complex learning
Immature neurons might be recruited into existing neural networks via oscillatory activity
Acknowledgments
This work was supported by the National Institutes of Health (grant nos. MH-59970 and ARRA-3R01MH059970-10S1) and the National Science Foundation (grant nos. IOB-0444364 and IOS-0914386) to T.J.S. This work was also supported by grants from the Academy of Finland, Emil Aaltonen Foundation, and Jenny and Antti Wihuri Foundation to M. S. N.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Gould E, Beylin A, Tanapat P, Reeves A, Shors TJ. Learning enhances adult neurogenesis in the hippocampal formation. Nature Neuroscience. 1999;2:260–5. doi: 10.1038/6365. [DOI] [PubMed] [Google Scholar]
- 2.Waddell J, Shors TJ. Neurogenesis, learning and associative strength. European Journal of Neuroscience. 2008;27:3020–8. doi: 10.1111/j.1460-9568.2008.06222.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Curlik DM, II, Shors TJ. Learning increases the survival of newborn neurons provided that learning is difficult to achieve and successful. Journal of Cognitive Neuroscience. 2011 doi: 10.1162/jocn.2010.21597. In press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Dalla C, Papachristos EB, Whetstone AS, Shors TJ. Female rats learn trace memories better than male rats and consequently retain a greater proportion of new neurons in their hippocampi. Proceedings of the National Academy of Sciences of the United States of America. 2009;106:2927–32. doi: 10.1073/pnas.0809650106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Leuner B, Mendolia-Loffredo S, Kozorovitskiy Y, Samburg D, Gould E, Shors TJ. Learning enhances the survival of new neurons beyond the time when the hippocampus is required for memory. The Journal of Neuroscience. 2004;24:7477–81. doi: 10.1523/JNEUROSCI.0204-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Nottebohm F. Neuronal replacement in the adult brain. Brain Research Bulletin. 2002;57:737–49. doi: 10.1016/s0361-9230(02)00750-5. [DOI] [PubMed] [Google Scholar]
- 7.Shors TJ, Miesegaes G, Beylin AV, Zhao M, Rydel T, Gould E. Neurogenesis in the adult is involved in the formation of trace memories. Nature. 2001;410:372–6. doi: 10.1038/35066584. [DOI] [PubMed] [Google Scholar]
- 8.Hebb DO. The organization of behavior: a neuropsychological theory. New York: Wiley; 1949. [Google Scholar]
- 9.Shors TJ, Matzel LD. Long-term potentiation: what’s learning got to do with it? The Behavioral and Brain Sciences. 1997;20:597–614. doi: 10.1017/s0140525x97001593. [DOI] [PubMed] [Google Scholar]
- 10.Cameron HA, McKay RDG. Adult neurogenesis produces a large pool of new granule cells in the dentate gyrus. The Journal of Comparative Neurology. 2001;435:406–17. doi: 10.1002/cne.1040. [DOI] [PubMed] [Google Scholar]
- 11.Rao MS, Shetty AK. Efficacy of doublecortin as a marker to analyse the absolute number and dendritic growth of newly generated neurons in the adult dentate gyrus. European Journal of Neuroscience. 2004;19:234–46. doi: 10.1111/j.0953-816x.2003.03123.x. [DOI] [PubMed] [Google Scholar]
- 12.Coras R, Siebzehnrubl FA, Pauli E, Huttner HB, Njunting M, Kobow K, et al. Low proliferation and differentiation capacities of adult hippocampal stem cells correlate with memory dysfunction in humans. Brain. 2010;133:3359–72. doi: 10.1093/brain/awq215. [DOI] [PubMed] [Google Scholar]
- 13.Knoth R, Singec I, Ditter M, Pantazis G, Capetian P, Meyer RP, et al. Murine features of neurogenesis in the human hippocampus across the lifespan from 0 to 100 years. PloS One. 2010;5:e8809. doi: 10.1371/journal.pone.0008809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Bayer SA, Yackel JW, Puri PS. Neurons in the rat dentate gyrus granular layer substantially increase during juvenile and adult life. Science. 1982;216:890–2. doi: 10.1126/science.7079742. [DOI] [PubMed] [Google Scholar]
- 15.Witter MP, Amaral DG. Hippocampal Formation. In: Paxinos G, editor. The Rat Nervous System. 2. San Diego: Academic Press; 1995. pp. 443–93. [Google Scholar]
- 16.Hodes GE, Yang L, Van Kooy J, Santollo J, Shors TJ. Prozac during puberty: distinctive effects on neurogenesis as a function of age and sex. Neuroscience. 2009;163:609–17. doi: 10.1016/j.neuroscience.2009.06.057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Shors TJ. Saving new brain cells. Scientific American. 2009;300:46–52. doi: 10.1038/scientificamerican0309-46. [DOI] [PubMed] [Google Scholar]
- 18.Shors TJ. From stem cells to grandmother cells: How neurogenesis relates to learning and memory. Cell Stem Cell. 2008;3:253–8. doi: 10.1016/j.stem.2008.08.010. [DOI] [PubMed] [Google Scholar]
- 19.Drapeau E, Mayo W, Aurousseau C, Le Moal M, Piazza PV, Abrous DN. Spatial memory performances of aged rats in the water maze predict levels of hippocampal neurogenesis. Proceedings of the National Academy of Sciences of the United States of America. 2003;100:14385–90. doi: 10.1073/pnas.2334169100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Olariu A, Cleaver KM, Shore LE, Brewer MD, Cameron HA. A natural form of learning can increase and decrease the survival of new neurons in the dentate gyrus. Hippocampus. 2005;15:750–62. doi: 10.1002/hipo.20097. [DOI] [PubMed] [Google Scholar]
- 21.Wurm F, Keiner S, Kunze A, Witte OW, Redecker C. Effects of skilled forelimb training on hippocampal neurogenesis and spatial learning after focal cortical infarcts in the adult rat brain. Stroke. 2007;38:2833–40. doi: 10.1161/STROKEAHA.107.485524. [DOI] [PubMed] [Google Scholar]
- 22.van Praag H, Schinder AF, Christie BR, Toni N, Palmer TD, Gage FH. Functional neurogenesis in the adult hippocampus. Nature. 2002;415:1030–4. doi: 10.1038/4151030a. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Toni N, Teng EM, Bushong EA, Aimone JB, Zhao C, Consiglio A, et al. Synapse formation on neurons born in the adult hippocampus. Nature Neuroscience. 2007;10:727–34. doi: 10.1038/nn1908. [DOI] [PubMed] [Google Scholar]
- 24.Toni N, Laplagne DA, Zhao C, Lombardi G, Ribak CE, Gage FH, et al. Neurons born in the adult dentate gyrus form functional synapses with target cells. Nature Neuroscience. 2008;11:901–7. doi: 10.1038/nn.2156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Sisti HM, Glass AL, Shors TJ. Neurogenesis and the spacing effect: learning over time enhances memory and the survival of new neurons. Learning & Memory. 2007;14:368–75. doi: 10.1101/lm.488707. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Epp JR, Scott NA, Galea LA. Strain differences in neurogenesis and activation of new neurons in the dentate gyrus in response to spatial learning. Neuroscience. 2011;172:342–254. doi: 10.1016/j.neuroscience.2010.10.025. [DOI] [PubMed] [Google Scholar]
- 27.Anderson ML, Sisti HM, Curlik DM, II, Shors TJ. Associative learning increases adult neurogenesis during a critical period. European Journal of Neuroscience. 2011;33:175–81. doi: 10.1111/j.1460-9568.2010.07486.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Epp JR, Spritzer MD, Galea LA. Hippocampus-dependent learning promotes survival of new neurons in the dentate gyrus at a specific time during cell maturation. Neuroscience. 2007;149:273–85. doi: 10.1016/j.neuroscience.2007.07.046. [DOI] [PubMed] [Google Scholar]
- 29.Dupret D, Fabre A, Döbrössy MD, Panatier A, Rodríguez JJ, Lamarque S, et al. Spatial learning depends on both the addition and removal of new hippocampal neurons. PLoS Biology. 2007;5:e214. doi: 10.1371/journal.pbio.0050214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Beylin AV, Gandhi CC, Wood GE, Talk AC, Matzel LD, Shors TJ. The role of the hippocampus in trace conditioning: Temporal discontinuity or task difficulty? Neurobiology of Learning and Memory A celebration of the scientific contributions of Richard F Thompson. 2001;76:447–61. doi: 10.1006/nlme.2001.4039. [DOI] [PubMed] [Google Scholar]
- 31.Drapeau E, Montaron MF, Aguerre S, Abrous DN. Learning-induced survival of new neurons depends on the cognitive status of aged rats. The Journal of Neuroscience. 2007;27:6037–44. doi: 10.1523/JNEUROSCI.1031-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Dalla C, Bangasser DA, Edgecomb C, Shors TJ. Neurogenesis and learning: Acquisition and asymptotic performance predict how many new cells survive in the hippocampus. Neurobiology of Learning and Memory. 2007;88:143–8. doi: 10.1016/j.nlm.2007.02.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Waddell J, Anderson ML, Shors TJ. Changing the rate and hippocampal dependence of trace eyeblink conditioning: Slow learning enhances survival of new neurons. Neurobiology of Learning and Memory. 2011;95:159–65. doi: 10.1016/j.nlm.2010.09.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Leuner B, Gould E, Shors TJ. Is there a link between adult neurogenesis and learning? Hippocampus. 2006;16:216–24. doi: 10.1002/hipo.20153. [DOI] [PubMed] [Google Scholar]
- 35.Overstreet-Wadiche L, Bromberg DA, Bensen AL, Westbrook GL. GABAergic signaling to newborn neurons in dentate gyrus. Journal of Neurophysiolology. 2005;94:4528–32. doi: 10.1152/jn.00633.2005. [DOI] [PubMed] [Google Scholar]
- 36.Ge S, Goh EL, Sailor KA, Kitabatake Y, Ming GL, Song H. GABA regulates synaptic integration of newly generated neurons in the adult brain. Nature. 2006;439:589–93. doi: 10.1038/nature04404. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Leuner B, Falduto J, Shors TJ. Associative memory formation increases the observation of dendritic spines in the hippocampus. The Journal of Neuroscience. 2003;23:659–65. doi: 10.1523/JNEUROSCI.23-02-00659.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Bannerman DM, Good MA, Butcher SP, Ramsay M, Morris RG. Distinct components of spatial learning revealed by prior training and NMDA receptor blockade. Nature. 1995;378:182–6. doi: 10.1038/378182a0. [DOI] [PubMed] [Google Scholar]
- 39.Tashiro A, Sandler VM, Toni N, Zhao C, Gage FH. NMDA-receptor-mediated, cell-specific integration of new neurons in adult dentate gyrus. Nature. 2006;442:929–33. doi: 10.1038/nature05028. [DOI] [PubMed] [Google Scholar]
- 40.Tronel S, Fabre A, Charrier V, Oliet SH, Gage FH, Abrous DN. Spatial learning sculpts the dendritic arbor of adult-born hippocampal neurons. Proceedings of the National Academy of Sciences of the United States of America. 2010;107:7963–8. doi: 10.1073/pnas.0914613107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Morris RG, Garrud P, Rawlins JN, O’Keefe J. Place navigation impaired in rats with hippocampal lesions. Nature. 1982;297:681–3. doi: 10.1038/297681a0. [DOI] [PubMed] [Google Scholar]
- 42.Shors TJ, Townsend DA, Zhao M, Kozorovitskiy Y, Gould E. Neurogenesis may relate to some but not all types of hippocampal-dependent learning. Hippocampus. 2002;12:578–84. doi: 10.1002/hipo.10103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Madsen TM, Kristjansen PE, Bolwig TG, Wörtwein G. Arrested neuronal proliferation and impaired hippocampal function following fractionated brain irradiation in the adult rat. Neuroscience. 2003;119:635–42. doi: 10.1016/s0306-4522(03)00199-4. [DOI] [PubMed] [Google Scholar]
- 44.Saxe MD, Battaglia F, Wang JW, Malleret G, David DJ, Monckton JC, et al. Ablation of hippocampal neurogenesis impairs contextual fear conditioning and synaptic plasticity in the dentate gyrus. Proceedings of the National Academy of Sciences of the United States of America. 2006;103:17501–6. doi: 10.1073/pnas.0607207103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Jaholkowski P, Kiryk A, Jedynak P, Ben Abdallah NM, Knapska E, Kowalczyk A, et al. New hippocampal neurons are not obligatory for memory formation; cyclin D2 knockout mice with no adult brain neurogenesis show learning. Learning & Memory. 2009;16:439–51. doi: 10.1101/lm.1459709. [DOI] [PubMed] [Google Scholar]
- 46.Snyder JS, Hong NS, McDonald RJ, Wojtowicz JM. A role for adult neurogenesis in spatial long-term memory. Neuroscience. 2005;130:843–52. doi: 10.1016/j.neuroscience.2004.10.009. [DOI] [PubMed] [Google Scholar]
- 47.Jessberger S, Clark RE, Broadbent NJ, Clemenson GDJ, Consiglio A, Lie DC, et al. Dentate gyrus-specific knockdown of adult neurogenesis impairs spatial and object recognition memory in adult rats. Learning & Memory. 2009;16:147–54. doi: 10.1101/lm.1172609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Dupret D, Revest JM, Koehl M, Ichas F, De Giorgi F, Costet P, et al. Spatial relational memory requires hippocampal adult neurogenesis. PloS One. 2008;3:e1959. doi: 10.1371/journal.pone.0001959. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Goodman T, Trouche S, Massou I, Verret L, Zerwas M, Roullet P, et al. Young hippocampal neurons are critical for recent and remote spatial memory in adult mice. Neuroscience. 2010;171:769–78. doi: 10.1016/j.neuroscience.2010.09.047. [DOI] [PubMed] [Google Scholar]
- 50.Garthe A, Behr J, Kempermann G. Adult-generated hippocampal neurons allow the flexible use of spatially precise learning strategies. PloS One. 2009;4:e5464. doi: 10.1371/journal.pone.0005464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Farioli-Vecchioli S, Saraulli D, Costanzi M, Pacioni S, Cinà I, Aceti M, et al. The timing of differentiation of adult hippocampal neurons is crucial for spatial memory. PLoS Biology. 2008;6:e246. doi: 10.1371/journal.pbio.0060246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Squire LR, Zola SM. Structure and function of declarative and nondeclarative memory systems. Proceedings of the National Academy of Sciences of the United States of America. 1996;93:13515–22. doi: 10.1073/pnas.93.24.13515. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Dickerson BC, Eichenbaum H. The episodic memory system: Neurocircuitry and disorders. Neuropsychopharmacology. 2010;35:86–104. doi: 10.1038/npp.2009.126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Clark RE, Squire LR. Classical conditioning and brain systems: The role of awareness. Science. 1998;280:77–81. doi: 10.1126/science.280.5360.77. [DOI] [PubMed] [Google Scholar]
- 55.Bangasser DA, Waxler DE, Santollo J, Shors TJ. Trace conditioning and the hippocampus: the importance of contiguity. The Journal of Neuroscience. 2006;26:8702–6. doi: 10.1523/JNEUROSCI.1742-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Buzsáki G. Rhythms of the Brain. New York: Oxford University Press; 2006. [Google Scholar]
- 57.Achanta P, Fuss M, Martinez JL., Jr Ionizing radiation impairs the formation of trace fear memories and reduces hippocampal neurogenesis. Behavioral Neuroscience. 2009;123:1036–45. doi: 10.1037/a0016870. [DOI] [PubMed] [Google Scholar]
- 58.Clark PJ, Brzezinska WJ, Thomas MW, Ryzhenko NA, Toshkov SA, Rhodes JS. Intact neurogenesis is required for benefits of exercise on spatial memory but not motor performance or contextual fear conditioning in C57BL/6J mice. Neuroscience. 2008;155:1048–58. doi: 10.1016/j.neuroscience.2008.06.051. [DOI] [PubMed] [Google Scholar]
- 59.Kitamura T, Saitoh Y, Takashima N, Murayama A, Niibori Y, Ageta H, et al. Adult neurogenesis modulates the hippocampus-dependent period of associative fear memory. Cell. 2009;139:814–27. doi: 10.1016/j.cell.2009.10.020. [DOI] [PubMed] [Google Scholar]
- 60.Zhang CL, Zou Y, He W, Gage FH, Evans RM. A role for adult TLX-positive neural stem cells in learning and behaviour. Nature. 2008;451:1004–7. doi: 10.1038/nature06562. [DOI] [PubMed] [Google Scholar]
- 61.Deng W, Saxe MD, Gallina IS, Gage FH. Adult-born hippocampal dentate granule cells undergoing maturation modulate learning and memory in the brain. The Journal of Neuroscience. 2009;29:13532–42. doi: 10.1523/JNEUROSCI.3362-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Winocur G, Wojtowicz JM, Sekeres M, Snyder JS, Wang S. Inhibition of neurogenesis interferes with hippocampal-dependent memory function. Hippocampus. 2006;16:296–304. doi: 10.1002/hipo.20163. [DOI] [PubMed] [Google Scholar]
- 63.Imayoshi I, Sakamoto M, Ohsuka T, Takao K, Miyakawa T, Yamaguchi M, et al. Roles of continuous neurogenesis in the structural and functional integrity of the adult fore brain. Nature Neuroscience. 2008;11:1153–61. doi: 10.1038/nn.2185. [DOI] [PubMed] [Google Scholar]
- 64.Hernández-Rabaza V, Llorens-Martín M, Velázquez-Sánchez C, Ferragud A, Arcusa A, Gumus HG, et al. Inhibition of adult hippocampal neurogenesis disrupts contextual learning but spares spatial working memory, long-term conditional rule retention and spatial reversal. Neuroscience. 2009;159:59–68. doi: 10.1016/j.neuroscience.2008.11.054. [DOI] [PubMed] [Google Scholar]
- 65.Wojtowicz JM, Askew ML, Winocur G. The effects of running and of inhibiting adult neurogenesis on learning and memory in rats. European Journal of Neuroscience. 2008;27:1494–502. doi: 10.1111/j.1460-9568.2008.06128.x. [DOI] [PubMed] [Google Scholar]
- 66.Drew MR, Denny CA, Hen R. Arrest of adult hippocampal neurogenesis in mice impairs single- but not multiple-trial contextual fear conditioning. Behavioral Neuroscience. 2010;124:446–54. doi: 10.1037/a0020081. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Snyder JS, Choe JS, Clifford MA, Jeurling SI, Hurley P, Brown A, et al. Adult-born hippocampal neurons are more numerous, faster maturing, and more involved in behavior in rats than in mice. The Journal of Neuroscience. 2009;29:14484–95. doi: 10.1523/JNEUROSCI.1768-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Tronel S, Belnoue L, Grosjean N, Revest JM, Piazza PV, Koehl M, et al. Adult-born neurons are necessary for extended contextual discrimination. Hippocampus. 2010 doi: 10.1002/hipo.20895. In press. [DOI] [PubMed] [Google Scholar]
- 69.Clelland CD, Choi M, Romberg C, Clemenson GD, Jr, Fragniere A, Tyers P, et al. A functional role for adult hippocampal neurogenesis in spatial pattern separation. Science. 2009;325:210–3. doi: 10.1126/science.1173215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Overstreet-Wadiche LS, Bensen AL, Westbrook GL. Delayed development of adult-generated granule cells in dentate gyrus. Journal of Neuroscience. 2006;26:2326–34. doi: 10.1523/JNEUROSCI.4111-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Laplagne DA, Espósito MS, Piatti VC, Morgenstern NA, Zhao C, van Praag H, et al. Functional convergence of neurons generated in the developing and adult hippocampus. PLoS Biology. 2006;4:2349–60. doi: 10.1371/journal.pbio.0040409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Overstreet Wadiche L, Bromberg DA, Bensen AL, Westbrook GL. GABAergic signaling to newborn neurons in dentate gyrus. Journal of Neurophysiolology. 2005;94:4528–32. doi: 10.1152/jn.00633.2005. [DOI] [PubMed] [Google Scholar]
- 73.Tonuka Y, Fuduka S, Namba T, Seki T, Hisatsune T. GABAergic excitation promotes neuronal differentiation in adult hippocampal progenitor cells. Neuron. 2005;47:803–15. doi: 10.1016/j.neuron.2005.08.023. [DOI] [PubMed] [Google Scholar]
- 74.Leuner B, Waddell J, Gould E, Shors TJ. Temporal discontiguity is neither necessary nor sufficient for learning-induced effects on adult neurogenesis. Journal of Neuroscience. 2006;26:13437–42. doi: 10.1523/JNEUROSCI.2781-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Aimone JB, Wiles J, Gage FH. Computational influence of adult neurogenesis on memory encoding. Neuron. 2009;61:187–202. doi: 10.1016/j.neuron.2008.11.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Kee N, Teixeira CM, Wang AH, Frankland PW. Preferential incorporation of adult-generated granule cells into spatial memory networks in the dentate gyrus. Nature Neuroscience. 2007;10:355–62. doi: 10.1038/nn1847. [DOI] [PubMed] [Google Scholar]
- 77.Trouche S, Bontempi B, Roullet P, Rampon C. Recruitment of adult-generated neurons into functional hippocampal networks contributes to updating and strengthening of spatial memory. Proceedings of the National Academy of Sciences of the United States of America. 2009;106:5919–24. doi: 10.1073/pnas.0811054106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Zhao C, Teng EM, Summers RG, Jr, Ming GL, Gage FH. Distinct morphological stages of dentate granule neuron maturation in the adult mouse hippocampus. Journal of Neuroscience. 2006;26:3–11. doi: 10.1523/JNEUROSCI.3648-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Ge S, Yang C, Hsu K, Ming G, Song H. A critical period for enhanced synaptic plasticity in newly generated neurons of the adult brain. Neuron. 2007;54:559–66. doi: 10.1016/j.neuron.2007.05.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Schmidt-Hieber C, Jonas P, Bischofberger J. Enhanced synaptic plasticity in newly generated granule cells of the adult hippocampus. Nature. 2004;429:184–7. doi: 10.1038/nature02553. [DOI] [PubMed] [Google Scholar]
- 81.Bruel-Jungerman E, Davis S, Rampon C, Laroche S. Long-term potentiation enhances neurogenesis in the adult dentate gyrus. The Journal of Neuroscience. 2006;26:5888–93. doi: 10.1523/JNEUROSCI.0782-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Kitamura T, Saitoh Y, Murayama A, Sugiyama H, Inokuchi K. LTP induction within a narrow critical period of immature stages enhances the survival of newly generated neurons in the adult rat dentate gyrus. Molecular Brain. 2010;3:13. doi: 10.1186/1756-6606-3-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Nottebohm F, Liu WC. The origins of vocal learning: New sounds, new circuits, new cells. Brain and Language. 2010;115:3–17. doi: 10.1016/j.bandl.2010.05.002. [DOI] [PubMed] [Google Scholar]
- 84.Wiskott L, Rasch MJ, Kempermann G. A functional hypothesis for adult hippocampal neurogenesis: avoidance of catastrophic interference in the dentate gyrus. Hippocampus. 2006;16:329–43. doi: 10.1002/hipo.20167. [DOI] [PubMed] [Google Scholar]
- 85.Rolls ET, Kesner RP. A computational theory of hippocampal function, and empirical tests of the theory. Progress in Neurobiology. 2006;79:1–48. doi: 10.1016/j.pneurobio.2006.04.005. [DOI] [PubMed] [Google Scholar]
- 86.Becker S. A computational principle for hippocampal learning and neurogenesis. Hippocampus. 2005;15:722–38. doi: 10.1002/hipo.20095. [DOI] [PubMed] [Google Scholar]
- 87.Alme CB, Buzzetti RA, Marrone DF, Leutgeb JK, Chawla MK, Schaner MJ, et al. Hippocampal granule cells opt for early retirement. Hippocampus. 2010;20:1109–23. doi: 10.1002/hipo.20810. [DOI] [PubMed] [Google Scholar]
- 88.Kim JJ, Clark RE, Thompson RF. Hippocampectomy impairs the memory of recently, but not remotely, acquired trace conditioned responses. Behavioral Neuroscience. 1995;109:195–203. doi: 10.1037//0735-7044.109.2.195. [DOI] [PubMed] [Google Scholar]
- 89.Takehara K, Kawahara S, Kirino Y. Time-dependent reorganization of the brain components underlying memory retention in trace eyeblink conditioning. The Journal of Neuroscience. 2003;23:9897–905. doi: 10.1523/JNEUROSCI.23-30-09897.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Riedel G, Micheau J, Lam AG, Roloff EL, Martin SJ, Bridge H, et al. Reversible neural inactivation reveals hippocampal participation in several memory processes. Nature Neuroscience. 1999;2:898–905. doi: 10.1038/13202. [DOI] [PubMed] [Google Scholar]
- 91.Fries P. A mechanism for cognitive dynamics: neuronal communication through neuronal coherence. Trends in Cognitive Sciences. 2005;9:474–80. doi: 10.1016/j.tics.2005.08.011. [DOI] [PubMed] [Google Scholar]
- 92.Klimesch W. Memory processes, brain oscillations and EEG synchronization. INternational Journal of Pyschophysiology. 1996;24:61–100. doi: 10.1016/s0167-8760(96)00057-8. [DOI] [PubMed] [Google Scholar]
- 93.Singer W. Distributed processing and temporal codes in neuronal networks. Cognitive Neurodynamics. 2009;3:189–96. doi: 10.1007/s11571-009-9087-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Uhlhaas PJ, Pipa G, Lima B, Melloni L, Neuenschwander S, Nikolić D, et al. Neural synchrony in cortical networks: History, concept and current status. Frontiers in Integrative Neuroscience. 2009;3:17. doi: 10.3389/neuro.07.017.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Bland BH. The physiology and pharmacology of hippocampal formation theta rhythms. Progress in Neurobiology. 1986;26:1–54. doi: 10.1016/0301-0082(86)90019-5. [DOI] [PubMed] [Google Scholar]
- 96.Buzsáki G. Theta oscillations in the hippocampus. Neuron. 2002;33:325–40. doi: 10.1016/s0896-6273(02)00586-x. [DOI] [PubMed] [Google Scholar]
- 97.Nokia MS, Penttonen M, Coronen T, Wikgren J. Hippocampal theta-band activity and trace eyeblink conditioning in rabbits. Behavioral Neuroscience. 2009;123:631–40. doi: 10.1037/a0015334. [DOI] [PubMed] [Google Scholar]
- 98.Hoffmann LC, Berry SD. Cerebellar theta oscillations are synchronized during hippocampal theta-contingent trace conditioning. Proceedings of the National Academy of Sciences of the United States of America. 2009;106:21371–6. doi: 10.1073/pnas.0908403106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Wikgren J, Nokia MS, Penttonen M. Hippocampo-cerebellar theta band phase synchrony in rabbits. Neuroscience. 2010;165:1538–45. doi: 10.1016/j.neuroscience.2009.11.044. [DOI] [PubMed] [Google Scholar]
- 100.Buzsáki G, Draguhn A. Neuronal oscillations in cortical networks. Science. 2004;304:1926–9. doi: 10.1126/science.1099745. [DOI] [PubMed] [Google Scholar]
- 101.Sirota A, Montgomery S, Fujisawa S, Isomura Y, Zugaro M, Buzsáki G. Entrainment of neocortical neurons and gamma oscillations by the hippocampal theta rhythm. Neuron. 2008;60:683–97. doi: 10.1016/j.neuron.2008.09.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Sirota A, Buzsáki G. Interaction between neocortical and hippocampal networks via slow oscillations. Thalamus & Related Systems. 2005;3:245–59. doi: 10.1017/S1472928807000258. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Eschenko O, Ramadan W, Mölle M, Born J, Sara SJ. Sustained increase in hippocampal sharp-wave ripple activity during slow-wave sleep after learning. Learning & Memory. 2008;15:222–8. doi: 10.1101/lm.726008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Girardeau G, Benchenane K, Wiener SI, Buzsáki G, Zugaro MB. Selective suppression of hippocampal ripples impairs spatial memory. Nature Neuroscience. 2009;12:1222–3. doi: 10.1038/nn.2384. [DOI] [PubMed] [Google Scholar]
- 105.Chrobak JJ, Buzsáki G. High-frequency oscillations in the output networks of the hippocampal-entorhinal axis of the freely behaving rat. The Journal of Neuroscience. 1996;16:3056–66. doi: 10.1523/JNEUROSCI.16-09-03056.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Jensen O, Lisman JE. Hippocampal sequence-encoding driven by a cortical multi-item working memory buffer. Trends in Neurosciences. 2005;28:67–72. doi: 10.1016/j.tins.2004.12.001. [DOI] [PubMed] [Google Scholar]
- 107.Lisman J, Buzsáki G. A neural coding scheme formed by the combined function of gamma and theta oscillations. Schizophrenia Bulletin. 2008;34:974–80. doi: 10.1093/schbul/sbn060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Nyhus E, Curran T. Functional role of gamma and theta oscillations in episodic memory. Neuroscience and Biobehavioral Reviews. 2010;34:1023–35. doi: 10.1016/j.neubiorev.2009.12.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Lacefield CO, Itskov V, Reardon T, Hen R, Gordon JA. Effects of adult-generated granule cells on coordinated network activity in the dentate gyrus. Hippocampus. 2010 doi: 10.1002/hipo.20860. In press. [DOI] [PMC free article] [PubMed] [Google Scholar]