

Abstract
In tissue engineering fields, recent interest has been focused on stem cell therapy to replace or repair damaged or worn-out tissues due to congenital abnormalities, disease, or injury. In particular, the repair of articular cartilage degeneration by stem cell-based tissue engineering could be of enormous therapeutic and economic benefit for an aging population. Bone marrow-derived mesenchymal stem cells (MSCs) that can induce chondrogenic differentiation would provide an appropriate cell source to repair damaged cartilage tissues; however, we must first understand the optimal environmental conditions for chondrogenic differentiation. In this review, we will focus on identifying the best combination of MSCs and functional extracellular matrices that provides the most successful chondrogenesis.
1. Introduction
Tissue loss or degeneration caused by congenital abnormalities, disease, or injury is of great consequence given human tissue's limited intrinsic potential for healing [1]. In particular, articular cartilage shows little or no intrinsic capacity for repair in response to injury or disease, and even minor lesions or injuries may lead to progressive damage and joint degeneration. Currently, frequent treatments, such as surgical intervention, to repair articular cartilage are less than satisfactory and rarely restore full function. One strategy for repairing articular cartilage degeneration via tissue engineering technologies is to create constructs of cells placed or injected onto or with matrices [2]. The underlying principle of tissue engineering (Figure 1(a)) involves the utilisation of biocompatible and mechanically conductive scaffolds, productive cell sources, and inductive molecules for the optimal differentiation and proliferation of the cell type of interest [3]. In this method, constructs of autologous, allogeneic, or xenogeneic cells seeded in scaffolds, that is, synthetic extracellular matrices designed to support cell growth and tissue development, are implanted at a repair site in the body to promote the differentiation and maturation of the cell type of interest (Figure 1(b)) [4]. In practice, tissue-specific cells are often seeded into the scaffold ex vivo prior to transplantation, and with time, the cells synthesise a new extracellular matrix (ECM) as the scaffold produces new, properly functioning tissue.
Figure 1.
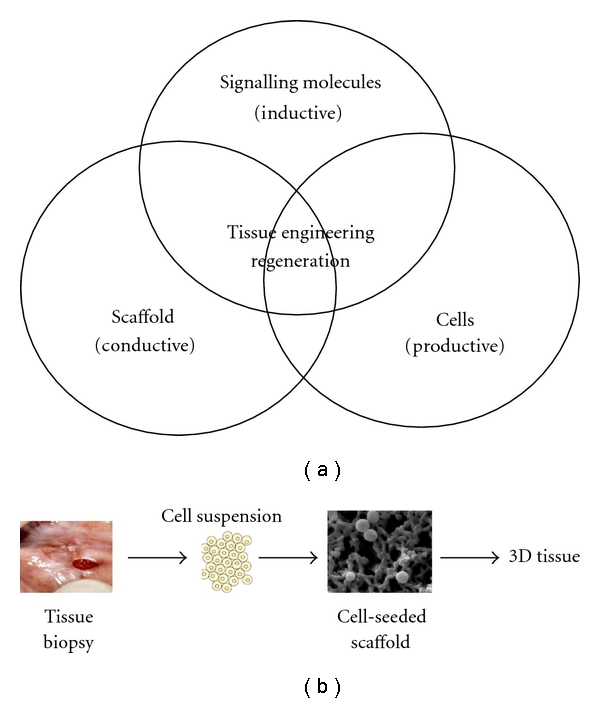
Tissue engineering strategy. (a) Basic principles of tissue engineering. (b) General methods of cell culture using a scaffold.
For this reason, the appropriate selection of cells and materials as scaffolds is one of the most important factors for successful, cell-based cartilage tissue engineering because the reconstruction and regeneration of damaged tissues occurs via an ordered pathway of cellular events affected by biological and mechanical factors [5, 6]. For clinical applications, autologous or allogeneic cell grafts are generally used. Autologous grafts are ideal, but they are often limited by the availability of donors [7]. Moreover, xenogeneic grafts are frequently subject to rejection as antigens present may elicit an immune reaction in the recipient and are further limited by pathogens found in the donor tissue. However, the use of allogeneic grafts is clinically routine due to the development of immunosuppressive drug therapies, such as cyclosporine, FK506, and rapamycin. Stem cells have the potential to be applied as a prepared allogeneic graft, thereby avoiding the need for tissue harvesting of prospective recipients, an extraordinary therapeutic advantage for many cell types. They have the capacity for self-renewal and the ability to generate differentiated cells. Recently, the field of stem cell biology has attracted more attention because of the isolation of human embryonic stem cells (ESCs), and the suggestion that adult stem cells may have a broader potential, that is, plasticity, than was previously thought [8].
ESCs derived from totipotent cells of an early mammalian embryo can proliferate indefinitely and can give rise to virtually any cell type. Therefore, the use of ESCs to replace damaged cells and tissues promises future hope for the treatment of many diseases. However, many countries now face complex ethical and legal questions as a result of the research to develop these cell therapies [9]. To circumvent these problems, many attempts have been made to isolate adult stem cells from mammalian tissues [10]. In particular, the adult bone marrow contains mesenchymal stem cells (MSCs), which contribute to the regeneration of mesenchymal tissues, such as bone, cartilage, muscle, ligament, tendon, adipose, bone marrow stroma, and other connective tissues [11, 12] and may be obtained from patients using minimally invasive techniques such as a bone marrow biopsy.
In this review, we will focus on the use of synthetic polymeric scaffolds in articular cartilage therapy and discuss the strategies for specific targeting. In particular, we will describe the potential use of MSCs to deliver these scaffolds.
2. MSCs for Cartilage Repair
In cell-based tissue engineering fields, selection of the source cells is required for consideration of several criteria, including ease of access and availability, a capacity for differentiation, and a lack of minimal immunogenic or tumourigenic ability. For cartilage repair in cell-based tissue engineering applications, source cells have included committed chondrocytes, ESCs, and adult stem cells. Each cell type has its limitations and advantages due to its intrinsic biological properties. However, chondrocytes have shown limited redifferentiation capability after in vivo expansion in clinical trials and in tissue engineering applications. Moreover, ESCs and their unwanted differentiations, such as tumour formations, are associated with ethical and legal concerns and are thereby an unsuitable cell source in basic research and clinical applications, despite the infinite pluripotentiality of ESCs. However, adult stem cells derived from various adult tissues have emerged as promising cell sources [11].
Among the adult stem cells, specifically multipotent adult stem cells, MSCs are considered to be the cell type of choice for cell-based cartilage tissue engineering because of (1) the ease with which they can be isolated and expanded and (2) their multilineage differentiation capabilities [13]. The isolation of these cells from adult tissues raises opportunities for the development of novel cellular therapies without the ethical considerations associated with ESC usage. Because of their multipotentiality and capacity for self-renewal, unlike ESCs, MSCs may represent units of active regeneration for damaged cartilage [14].
Although MSCs have shown great promise in cartilage repair and regeneration, several requirements should be examined to allow them to effectively differentiate into chondrocytes and maintain this differentiated phenotype prior to implantation or delivery. These would involve the methods and materials for culture conditions of MSCs to repair or restore full functions of damaged cartilage. Upon proper culture conditions containing certain exogenous factors, MSCs can be directed towards chondrogenic differentiation. Growth factors that promote chondrogenesis or demonstrate a chondrogenic effect both in vivo and in vitro include bone morphogenetic proteins (BMPs), transforming growth factor-β (TGF-β), and insulin-like growth factors [15–17]. BMPs are secreted molecules of the TGF-β superfamily of growth and differentiation factors that were originally detected in and purified from demineralised bone [18]. BMPs have been shown to function as key regulators in cartilage and bone development [19–22] and to function in repair and remodelling of the adult skeletal system [23–25]. These findings also provide crucial insights into cartilage repair and regeneration as the progression of osteoarthritis is always accompanied by damage to the subchondral bone and the formation of osteophytes. Despite many advances in proper culture conditions for MSCs, most of the methods are limited to a two-dimensional (2D) culture, and most of them provide little information about the proper chondrogenic induction of MSCs in three-dimensional (3D) culture.
3. MSC-Based Cartilage Tissue Engineering
The 3D culture system for cell differentiation and proliferation may improve our understanding of the structure-function relationship under both normal and pathological conditions. With regard to cell-based cartilage tissue engineering, successfully reconstructed cartilage tissue formation would be structurally reunited with the peripheral cartilage and would suggest biomechanical properties necessary for permanence and efficacy under 3D environmental conditions [26, 27]. It is currently accepted that 3D behaviours of specific cells, including MSCs, are quite different from 2D behaviours, indicating that 3D in vitro culture systems can mimic the in vivo situation more closely than 2D cultures [28–30].
3.1. Design of ECM
The successful outcome of cell-based cartilage tissue engineering using a 3D culture of MSCs ultimately depends on the design of synthetic artificial ECMs for the proper differentiation of MSCs into chrondrocytes [31] because specific stem cells alone face obstacles in the construction of cartilage formation. For the development of viable cartilage formation, synthetic ECMs should be designed considering a number of requirements, namely, mechanical properties such as a capability to withstand the large contact stresses and strains of an articulating joint, allow functional tissue growth, and provide appropriate cell-matrix interactions to stimulate tissue growth [32, 33]. One challenge for these solutions is the delivery of stem cells to the targeted tissue without cell loss. Delivery of stem cells alone may not be sufficient to restore damaged tissues as a result of enormous cell loss after delivery. Furthermore, little is known regarding the optimal delivery strategy for stem cells. Increasing the efficiency of MSC delivery and targeting the infused cells to specific tissue locations could have a large impact on the therapeutic uses of MSCs to treat diseases [12]. The delivery of MSCs can be achieved using injectable matrices, soft scaffolds, membranes, solid load-bearing scaffolds, or immunoprotective macroencapsulation. Thus, to expand their clinical potential, next generation therapies will depend on smart delivery concepts that make use of the regenerative potential of MSCs, morphogenetic growth factors, and biomimetic materials.
Other challenges are associated with the biomaterial scaffolds designed to guide tissue growth and differentiation. These biomaterials must meet several criteria to maximise the chances of a successful repair, including biodegradability and/or biocompatibility, facilitating functional tissue growth, and appropriate biomechanical properties [34–36]. Biomaterials used for cartilage tissue engineering can have the form of cell-entrapped scaffolds with nano- or microstructures [30, 37, 38].
3.2. Biomaterial Scaffolds
Polymercross-linked scaffolds have been used for cell entrapment in cell-based tissue engineering applications, due to their 3D networks, tissue-like water content, structure stability, and biocompatibility [30, 40]. There are a number of candidate scaffolding materials that include natural polysaccharides and proteins, such as alginate and collagen, and synthetic polymers, such as polyethylene glycol (PEG), poly(ε-caprolactone) (PCL), polyglycolic acid (PGA), polylactic acid (PLA), and poly (D,L-lactide-co-glycolide) (PLGA) [41]. Although tissue-engineered constructs are well designed for 3D culture, maintaining the chondrogenic phenotype is problematic when culturing MSCs alone on the scaffolds, that is, without other local factors in vivo. To overcome these problems, a number of novel biomaterials, innovative cell culture techniques, and newly discovered growth factors should be utilised according to directions from cell-based tissue engineering applications. In particular, growth factors are local factors that are key regulators for proper differentiation of MSCs in research and clinical applications.
For long-term retention of cells in vivo, the cells should be contained within the inner structures of the scaffold. With a proper fabrication method, porous scaffolds can help cells to penetrate into the scaffold when implanted into the body [42, 43]. One of the basic problems from a scaffold design point of view is that to achieve significant strength, the scaffold material must have sufficiently high interatomic and intermolecular bonding, but must have at the same time a physical and chemical structure which allows for hydrolytic attack and breakdown. For example, PCL as the scaffold material degrades relatively slowly and possesses an appropriately high bulk stiffness to facilitate MSC differentiation toward skeletal lineages [44]. The PCL scaffold was used after loading TGF-β3 physically complexed with chondroitin sulfate (CS) [45], because it was designed to maintain an interconnected pore network for at least 6 months, and the neocartilage would have sufficient time to mature without biomechanical overload [46].
In addition to porous scaffolds, the biofunctional constructs have been developed for better chondrogenesis of MSCs. In the application of cartiliage formation in the research field, for instance, fully thermoreversible gelling polymers have attracted considerable attention for use as scaffold materials to hold cells in situ [47]. Na et al. developed poly(N-isopropylacrylamide) (PNIPAAm)-based hydrogel scaffolds to investigate their capability to deliver a mixture of MSCs and growth factors for the better induction of chondrogenic differentiation [48, 49]. These thermoreversible hydrogel scaffolds can revert from solid to liquid state and from liquid to solid state without abrogating their intrinsic properties. These scaffolds were completely soluble in aqueous solutions at temperatures below their lower critical solution temperature (LCST), but they solidify at temperatures above their LCST, forming a hydrated gel [49, 50]. However, its clinical application is limited due to its nonbiodegradability. Therefore, the design of biodegradable scaffolds, which have biofunctions as well as mechanical strength for effective chondrogenesis of MSCs, is required to overcome the issues. The ideal scaffold has sufficient strength to protect cells from compression and shearing forces, while still having injury site anchoring potential and porosity to allow nutrient and differentiation factors to diffuse through it. The scaffold must also degrade at a rate that optimizes cellular growth and tissue regeneration. Such ideal scaffolds have not yet been designed. The optimal time point for evaluation of a scaffold-based treatment is also critical, which depends on the scaffold, cells, and tissue in question [51].
Biodegradable micro- and nanocomposite materials that can provide the appropriate strength, integrate the desirable biological cues, and provide for the controlled sequential delivery of multiple growth factors would help fulfill the promise of regenerative medicine. Also the development of relevant scaffold design using suitable biomaterials and incorporation of appropriate biomolecules and the selection of cell types plays a vital role in tissue repair. Recent conceptual advances, which have taken advantage of new and practical techniques for size distribution and stabilisation control, have created novel routes for the synthesis of nanoparticle-based materials, in which nanoparticle building blocks can be spatially ordered in a controlled manner [37]. Polyionic complexed nanoparticles are composed of heparin and poly(l-lysine) as a stem cell-delivery system, as depicted in Figure 2(a). Heparin-functionalised hydrogel supported MSC viability and induced chondrogenic differentiation. Negatively charged heparin is widely used in the biomaterial field because it can interact with a variety of proteins that have heparin-binding domains, including various growth factors that enable the growth factors to cross-link their receptors. Additionally, poly(l-lysine) has been commonly used as a model cationic polymer to fabricate the polyionic complexes with anionic polymers. Heparin/poly(l-lysine) nanoparticles formed a polyelectrostatic layer-by-layer assembly and were sequentially immobilised on PLGA microspheres as microcarriers of MSCs. The heparin/poly(l-lysine) polyelectrolyte complex is highly distributed on the PLGA microspheres. PLGA is known as the scaffold material to easily form nano- or microstructured particles which can entrap cells or load small molecules due to its excellent biocompatibility, degradability, and processibility [52]. The specific binding activity of heparin in the bioconjugate is not reduced in the immobilisation process results, which may be due to the presence of heparin within the outer shell of the nanoparticles on the surfaces of the PLGA microspheres (Figure 2(b)). As shown in the SEM images, the heparin/poly(l-lysine) electrolyte complex is heavily distributed on the PLGA microspheres.
Figure 2.
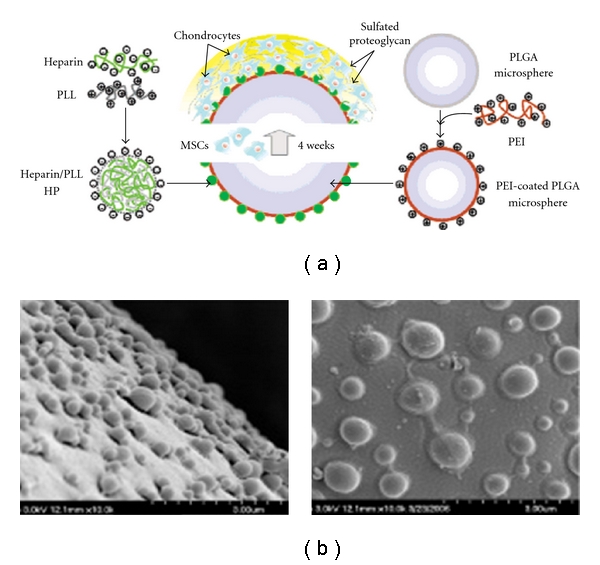
(a) Diagram of heparinized nanoparticles coated onto PLGA microsphere for stem cell delivery. (b) The SEM images of PLGA microspheres (upper (left) and bottom (right)) fabricated with heparin/poly(l-lysine) nanoparticles [37].
In another example using the nanoconstructs composed of fibrin hydrogels containing MSCs mixed with heparinised BMP-2, the bioactivities of entrapped MSCs mixed with growth factors were maintained for long term [30]. Moreover, PLGA microspheres can be used as cell delivery vehicles for controlled release of cells mixed with small molecules, which can help MSCs enhance their bioactivities. Park et al. reported the dual delivery of TGF-β3 and dexamethasone from transplanted PLGA constructs in vivo to engineer inflammation-free and cartilage-associated tissue [38].
Although PLGA has proved to be an excellent material for cartilage tissue engineering due to its biodegradable properties, mechanical strength, and ease of fabrication into a considerably complex formation, the principle usage of PLGA as a delivery vehicle has problems as it does not offer a desirable environment for cell adhesion due to its limitation of binding sites mediated by biological recognition and high hydrophobicity. Based upon an early fundamental step in which positive cell-substrate interactions enable cell proliferation, migration, and differentiation on the surface of materials, many studies have focused on modifying the matrix surface in an effort to increase cell-substrate interaction for cell delivery [53–55]. Binding sites in fibronectin, osteopontin, collagens, fibrinogen, and thrombospondin that contain the tripeptide Arg-Gly-Asp (RGD) are easily recognised by mammalian cells. The RGD sequences of the adhesive proteins are recognised by a structurally related receptor family, that is, integrins, which bind to RGD on the surface of cells, allowing cells to adhere [42, 56, 57]. The incorporation of bioactive motifs such as RGD may be the best adapted strategy to enhance cell adhesion [58–60]. As 3D scaffolds have a larger surface area and highly interconnected porous structures with suitable porosity and pore size, modification of the scaffold surface to improve the interaction between cells and the surface would have a greater potential for tissue engineering [39, 61–65]. A promising strategy is to immobilise RGD peptides on scaffold surfaces by evaluating embedded MSC behaviours, including attachment, cellular distribution, signal transduction, and survival on the modified surface. For instance, PLGA microscaffolds conjugated with RGD peptides were constructed as an MSC-delivery vehicle (Figure 3). The regulation of stem cell differentiation by adhesion molecules and growth factors has the potential to enable the formation of therapeutic vehicles for the delivery of MSCs that are easily fabricated, less expensive, and more easily controlled than currently available delivery systems. The embedded MSCs easily adhered onto PLGA microspheres mediated by the RGD peptide, proliferated well onto the scaffolds and differentiated to perform distinct functions [66].
Figure 3.
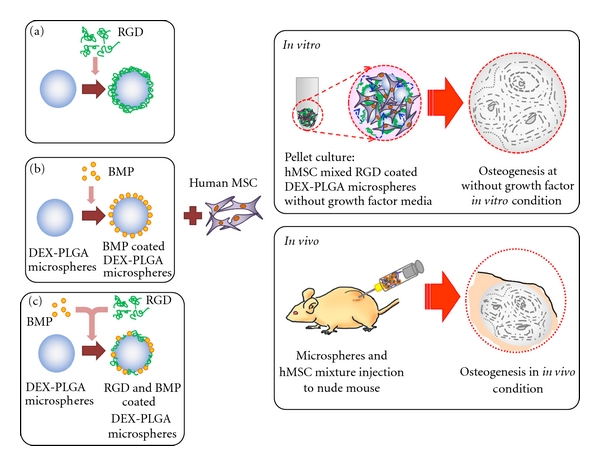
Schematic diagram of RGD-modified PLGA microspheres containing growth factors and dexamethasone. Using a layer-by-layer (LBL) technique, positively charged, PEI precoated PLGA microspheres coated with negatively charged RGD molecules were fabricated for hMSC delivery and regeneration of injured tissues. The combination of growth factors, DEX, and RGD was an effective scaffold for cell delivery and differentiation of embedded hMSCs [39].
Culture methods are deeply considerable to improve the chondrogenetic potential of MSCs, because MSCs markedly decrease with the increase of passage number. If cultured in a medium that is not supplemented with factors facilitating the maintenance of plural differentiation potential, MSCs can hardly differentiate into chondrocytes after repeated passages. Some strong inductive signals for chondrogenesis are required to differentiate the passage-cultured MSCs into chondrocytes [67]. Coculture of MSCs with mature chondrocytes is a strategy that both provides inductive signals and solves the cell source problem. Coculture techniques of MSCs and autologous chondrocytes are frequently used to improve induction of the chondrogenic differentiation of MSCs instead of including growth factors in the MSC culture [68, 69], because the chondrogenic differentiation of MSCs induced by growth factors exhibits some defects, including an instability of the chondrocyte phenotype and a lack of ECM secretion [70]. In the 3D hydrogel constructs, Coculture with autologous chondrocytes and MSCs can show a significantly higher number of specific lacunae phenotypes [68, 71]. Chondrocytes express soluble growth factors that can help MSCs selectively promote chondrogenesis, and this selective effect is not mimicked by an exogenously added growth factors.
In addition, transfection of MSCs with growth factor genes has been proposed and practiced. However, the outcomes are not completely desirable not only due to the damage caused by the invasive procedure of transfection but also because long-term overexpression of a growth factor may result in undesirable changes in the transfected cells. To overcome these challenges, effective chondrogenesis in MSCs can be achieved by coculturing them with autologous chondrocytes transfected with growth factor genes [72]. This gene transfected Coculture system can avoid directly transfecting MSCs, but instead transfers growth factor genes to their Cocultured chondrocytes.
4. Final Remarks and Further Researches
The clinical need for cartilage repair technologies is unmistakable. Many people over the age of 40 suffer from degeneration or injury of their cartilage, leading to a reduced workforce and increased medical expenses. Thus, improvements in cartilage repair using a cell-based tissue engineering approach will greatly benefit public health and the economy. Personalised cell therapy for cartilage repair using cell-based tissue engineering technologies would provide clinically practical methods for producing a cartilage tissue equivalent. A number of biomaterials are available as scaffolds, and research continues to help us understand more details about how tissues develop and which cell type should be applied. These studies have provided details of how tissues grow in vitro and in vivo, but clinical applications depend on working with surgeons and the translation of these materials and technologies to in vivo models that are more relevant to patients. When cell-based cartilage tissue engineering technologies are applied to new animal models, we attempted to find better functional compositions for successful applications than were observed in previous studies. Although stem cell-based cartilage tissue engineering systems may demonstrate success even in animal models, there are a number of new challenges when the technologies are applied to humans. Further research on in vivo application must address immunological issues, integration of host and stem cell-based engineered cartilage, and the variability of tissue development in an in vivo environment, depending on surrounding disease processes, age, or physical activity. Therefore, interdisciplinary studies are not only necessary but crucial before cell-based cartilage tissue engineering can reach its full potential in cartilage repair and regeneration.
Acknowledgments
This research was financially supported by the Fundamental R&D Program for Core Technology of Materials (K0006028) Republic of Korea, the Gyeonggi Regional Research Center (GRRC), the Ministry of Knowledge Economy (MKE), and the Korea Industrial Technology Foundation (KOTEF) through the Human Resource Training Project for Strategic Technology, Republic of Korea.
References
- 1.Griffith LG, Naughton G. Tissue engineering—current challenges and expanding opportunities. Science. 2002;295(5557):1009–1014. doi: 10.1126/science.1069210. [DOI] [PubMed] [Google Scholar]
- 2.Kuo CK, Tuan RS. Tissue engineering with mesenchymal stem cells. IEEE Engineering in Medicine and Biology Magazine. 2003;22(5):51–56. doi: 10.1109/memb.2003.1256272. [DOI] [PubMed] [Google Scholar]
- 3.Song L, Baksh D, Tuan RS. Mesenchymal stem cell-based cartilage tissue engineering: cells, scaffold and biology. Cytotherapy. 2004;6(6):596–601. doi: 10.1080/14653240410005276-1. [DOI] [PubMed] [Google Scholar]
- 4.Rose FRAJ, Oreffo ROC. Bone tissue engineering: hope vs hype. Biochemical and Biophysical Research Communications. 2002;292(1):1–7. doi: 10.1006/bbrc.2002.6519. [DOI] [PubMed] [Google Scholar]
- 5.Patterson TE, Kumagai K, Griffith L, Muschler GF. Cellular strategies for enhancement of fracture repair. The Journal of Bone and Joint Surgery. 2008;90(1):111–119. doi: 10.2106/JBJS.G.01572. [DOI] [PubMed] [Google Scholar]
- 6.Kloss FR, Gassner R, Preiner J, et al. The role of oxygen termination of nanocrystalline diamond on immobilisation of BMP-2 and subsequent bone formation. Biomaterials. 2008;29(16):2433–2442. doi: 10.1016/j.biomaterials.2008.01.036. [DOI] [PubMed] [Google Scholar]
- 7.Lee KH. Tissue-engineered human living skin substitutes: development and clinical application. Yonsei Medical Journal. 2000;41(6):774–779. doi: 10.3349/ymj.2000.41.6.774. [DOI] [PubMed] [Google Scholar]
- 8.Ramalho-Santos M, Yoon S, Matsuzaki Y, Mulligan RC, Melton DA. ‘Stemness’: transcriptional profiling of embryonic and adult stem cells. Science. 2002;298(5593):597–600. doi: 10.1126/science.1072530. [DOI] [PubMed] [Google Scholar]
- 9.Robertson JA. Human embryonic stem cell research: ethical and legal issues. Nature Reviews Genetics. 2001;2(1):74–78. doi: 10.1038/35047594. [DOI] [PubMed] [Google Scholar]
- 10.Toma JG, Akhavan M, Fernandes KJL, et al. Isolation of multipotent adult stem cells from the dermis of mammalian skin. Nature Cell Biology. 2001;3(9):778–784. doi: 10.1038/ncb0901-778. [DOI] [PubMed] [Google Scholar]
- 11.Pittenger MF, Mackay AM, Beck SC, et al. Multilineage potential of adult human mesenchymal stem cells. Science. 1999;284(5411):143–147. doi: 10.1126/science.284.5411.143. [DOI] [PubMed] [Google Scholar]
- 12.Caplan AI. Cell delivery and tissue regeneration. Journal of Controlled Release. 1990;11(1–3):157–165. [Google Scholar]
- 13.Bruder SP, Jaiswal N, Haynesworth SE. Growth kinetics, self-renewal, and the osteogenic potential of purified human mesenchymal stem cells during extensive subcultivation and following cryopreservation. Journal of Cellular Biochemistry. 1997;64(2):278–294. doi: 10.1002/(sici)1097-4644(199702)64:2<278::aid-jcb11>3.0.co;2-f. [DOI] [PubMed] [Google Scholar]
- 14.Weissman IL. Translating stem and progenitor cell biology to the clinic: barriers and opportunities. Science. 2000;287(5457):1442–1446. doi: 10.1126/science.287.5457.1442. [DOI] [PubMed] [Google Scholar]
- 15.Linkhart TA, Mohan S, Baylink DJ. Growth factors for bone growth and repair: IGF, TGFβ and BMP. Bone. 1996;19(1):S1–S12. doi: 10.1016/s8756-3282(96)00138-x. [DOI] [PubMed] [Google Scholar]
- 16.O’Driscoll SW. Articular cartilage regeneration using periosteum. Clinical Orthopaedics and Related Research. 1999;(367):S186–S203. doi: 10.1097/00003086-199910001-00020. [DOI] [PubMed] [Google Scholar]
- 17.Kil SJ, Carlin C. Isolation, characterization, and chondrogenic potential of human bone marrow-derived multipotential stromal cells. Journal of Cellular Physiology. 2000;185(1):98–106. doi: 10.1002/1097-4652(200010)185:1<98::AID-JCP9>3.0.CO;2-1. [DOI] [PubMed] [Google Scholar]
- 18.Wang EA, Rosen V, Cordes P, et al. Purification and characterization of other distinct bone-inducing factors. Proceedings of the National Academy of Sciences of the United States of America. 1988;85(24):9484–9488. doi: 10.1073/pnas.85.24.9484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Kingsley DM. What do BMPs do in mammals? Clues from the mouse short-ear mutation. Trends in Genetics. 1994;10(1):16–21. doi: 10.1016/0168-9525(94)90014-0. [DOI] [PubMed] [Google Scholar]
- 20.Duprez D, De EJ, Richardson MK, et al. Overexpression of BMP-2 and BMP-4 alters the size and shape of developing skeletal elements in the chick limb. Mechanisms of Development. 1996;57(2):145–157. doi: 10.1016/0925-4773(96)00540-0. [DOI] [PubMed] [Google Scholar]
- 21.Macias D, Gañan Y, Sampath TK, Piedra ME, Ros MA, Hurle JM. Role of BMP-2 and OP-1 (BMP-7) in programmed cell death and skeletogenesis during chick limb development. Development. 1997;124(6):1109–1117. doi: 10.1242/dev.124.6.1109. [DOI] [PubMed] [Google Scholar]
- 22.Denker AE, Haas AR, Nicoll SB, Tuan RS. Chondrogenic differentiation of murine C3H10T1/2 multipotential mesenchymal cells: I. Stimulation by bone morphogenetic protein-2 in high-density micromass cultures. Differentiation. 1999;64(2):67–76. doi: 10.1046/j.1432-0436.1999.6420067.x. [DOI] [PubMed] [Google Scholar]
- 23.Lyons KM, Pelton RW, Hogan BL. Patterns of expression of murine Vgr-1 and BMP-2a RNA suggest that transforming growth factor-beta-like genes coordinately regulate aspects of embryonic development. Genes & Development. 1989;3(11):1657–1668. doi: 10.1101/gad.3.11.1657. [DOI] [PubMed] [Google Scholar]
- 24.Zehentner BK, Dony C, Burtscher H. The transcription factor Sox9 is involved in BMP-2 signaling. Journal of Bone and Mineral Research. 1999;14(10):1734–1741. doi: 10.1359/jbmr.1999.14.10.1734. [DOI] [PubMed] [Google Scholar]
- 25.Sellers RS, Zhang R, Glasson SS, et al. Repair of articular cartilage defects one year after treatment with recombinant human bone morphogenetic protein-2 (rhBMP-2) The Journal of Bone and Joint Surgery. 2000;82(2):151–160. doi: 10.2106/00004623-200002000-00001. [DOI] [PubMed] [Google Scholar]
- 26.Block MS, Finger I, Lytle R. Human mineralized bone in extraction sites before implant placement: preliminary results. Journal of the American Dental Association. 2002;133(12):1631–1638. doi: 10.14219/jada.archive.2002.0112. [DOI] [PubMed] [Google Scholar]
- 27.Werner CML, Favre P, Van Lenthe HG, Dumont CE. Pedicled vascularized rib transfer for reconstruction of clavicle nonunions with bony defects: anatomical and biomechanical considerations. Plastic and Reconstructive Surgery. 2007;120(1):173–180. doi: 10.1097/01.prs.0000263537.57701.8b. [DOI] [PubMed] [Google Scholar]
- 28.Valarmathi MT, Yost MJ, Goodwin RL, Potts JD. The influence of proepicardial cells on the osteogenic potential of marrow stromal cells in a three-dimensional tubular scaffold. Biomaterials. 2008;29(14):2203–2216. doi: 10.1016/j.biomaterials.2008.01.025. [DOI] [PubMed] [Google Scholar]
- 29.Jäger M, Urselmann F, Witte F, et al. Osteoblast differentiation onto different biometals with an endoprosthetic surface topography in vitro. Journal of Biomedical Materials Research—Part A. 2008;86(1):61–75. doi: 10.1002/jbm.a.31552. [DOI] [PubMed] [Google Scholar]
- 30.Park KH, Kim H, Moon S, Na K. Bone morphogenic protein-2 (BMP-2) loaded nanoparticles mixed with human mesenchymal stem cell in fibrin hydrogel for bone tissue engineering. Journal of Bioscience and Bioengineering. 2009;108(6):530–537. doi: 10.1016/j.jbiosc.2009.05.021. [DOI] [PubMed] [Google Scholar]
- 31.Chen FH, Rousche KT, Tuan RS. Technology insight: adult stem cells in cartilage regeneration and tissue engineering. Nature Clinical Practice Rheumatology. 2006;2(7):373–382. doi: 10.1038/ncprheum0216. [DOI] [PubMed] [Google Scholar]
- 32.Guilak F, Butler DL, Goldstein SA. Functional tissue engineering: the role of biomechanics in articular cartilage repair. Clinical Orthopaedics and Related Research. 2001;(391):S295–S305. [PubMed] [Google Scholar]
- 33.Butler DL, Goldstein SA, Guilak F. Functional tissue engineering: the role of biomechanics. Journal of Biomechanical Engineering. 2000;122(6):570–575. doi: 10.1115/1.1318906. [DOI] [PubMed] [Google Scholar]
- 34.Hunziker EB. Articular cartilage repair: basic science and clinical progress. A review of the current status and prospects. Osteoarthritis and Cartilage. 2002;10(6):432–463. doi: 10.1053/joca.2002.0801. [DOI] [PubMed] [Google Scholar]
- 35.Freed LE, Marquis JC, Nohria A, Emmanual J, Mikos AG, Langer R. Neocartilage formation in vitro and in vivo using cells cultured on synthetic biodegradable polymers. Journal of Biomedical Materials Research. 1993;27(1):11–23. doi: 10.1002/jbm.820270104. [DOI] [PubMed] [Google Scholar]
- 36.Freed LE, Vunjak-Novakovic G, Biron RJ, et al. Biodegradable polymer scaffolds for tissue engineering. Bio/Technology. 1994;12(7):689–693. doi: 10.1038/nbt0794-689. [DOI] [PubMed] [Google Scholar]
- 37.Na K, Kim S, Park K, et al. Heparin/poly(l-lysine) nanoparticle-coated polymeric microspheres for stem-cell therapy. Journal of the American Chemical Society. 2007;129(18):5788–5789. doi: 10.1021/ja067707r. [DOI] [PubMed] [Google Scholar]
- 38.Park JS, Na K, Woo DG, Yang HN, Park KH. Determination of dual delivery for stem cell differentiation using dexamethasone and TGF-β3 in/on polymeric microspheres. Biomaterials. 2009;30(27):4796–4805. doi: 10.1016/j.biomaterials.2009.05.054. [DOI] [PubMed] [Google Scholar]
- 39.Cho CS, Seo SJ, Park IK, et al. Galactose-carrying polymers as extracellular matrices for liver tissue engineering. Biomaterials. 2006;27(4):576–585. doi: 10.1016/j.biomaterials.2005.06.008. [DOI] [PubMed] [Google Scholar]
- 40.Nuttelman CR, Benoit DSW, Tripodi MC, Anseth KS. The effect of ethylene glycol methacrylate phosphate in PEG hydrogels on mineralization and viability of encapsulated hMSCs. Biomaterials. 2006;27(8):1377–1386. doi: 10.1016/j.biomaterials.2005.08.014. [DOI] [PubMed] [Google Scholar]
- 41.Li WJ, Danielson KG, Alexander PG, Tuan RS. Biological response of chondrocytes cultured in three-dimensional nanofibrous poly(ε-caprolactone) scaffolds. Journal of Biomedical Materials Research—Part A. 2003;67(4):1105–1114. doi: 10.1002/jbm.a.10101. [DOI] [PubMed] [Google Scholar]
- 42.Hwang NS, Varghese S, Zhang Z, Elisseeff J. Chondrogenic differentiation of human embryonic stem cell-derived cells in arginine-glycine-aspartate-modified hydrogels. Tissue Engineering. 2006;12(9):2695–2706. doi: 10.1089/ten.2006.12.2695. [DOI] [PubMed] [Google Scholar]
- 43.Park JS, Woo DG, Sun BK, et al. In vitro and in vivo test of PEG/PCL-based hydrogel scaffold for cell delivery application. Journal of Controlled Release. 2007;124(1-2):51–59. doi: 10.1016/j.jconrel.2007.08.030. [DOI] [PubMed] [Google Scholar]
- 44.Engler AJ, Sen S, Sweeney HL, Discher DE. Matrix elasticity directs stem cell lineage specification. Cell. 2006;126(4):677–689. doi: 10.1016/j.cell.2006.06.044. [DOI] [PubMed] [Google Scholar]
- 45.Park JS, Yang HJ, Woo DG, Yang HN, Na K, Park KH. Chondrogenic differentiation of mesenchymal stem cells embedded in a scaffold by long-term release of TGF-β3 complexed with chondroitin sulfate. Journal of Biomedical Materials Research—Part A. 2010;92(2):806–816. doi: 10.1002/jbm.a.32388. [DOI] [PubMed] [Google Scholar]
- 46.Huang Q, Goh JCH, Hutmacher DW, Lee EH. In vivo mesenchymal cell recruitment by a scaffold loaded with transforming growth factor β1 and the potential for in situ chondrogenesis. Tissue Engineering. 2002;8(3):469–482. doi: 10.1089/107632702760184727. [DOI] [PubMed] [Google Scholar]
- 47.Fisher JP, Jo S, Mikos AG, Reddi AH. Thermoreversible hydrogel scaffolds for articular cartilage engineering. Journal of Biomedical Materials Research—Part A. 2004;71(2):268–274. doi: 10.1002/jbm.a.30148. [DOI] [PubMed] [Google Scholar]
- 48.Na K, Park JH, Kim SW, et al. Delivery of dexamethasone, ascorbate, and growth factor (TGF β-3) in thermo-reversible hydrogel constructs embedded with rabbit chondrocytes. Biomaterials. 2006;27(35):5951–5957. doi: 10.1016/j.biomaterials.2006.08.012. [DOI] [PubMed] [Google Scholar]
- 49.Park KH, Na K, Sung WK, Sung YJ, Kyu HP, Chung HM. Phenotype of hepatocyte spheroids behavior within thermo-sensitive poly(NiPAAm-co-PEG-g-GRGDS) hydrogel as a cell delivery vehicle. Biotechnology Letters. 2005;27(15):1081–1086. doi: 10.1007/s10529-005-8453-0. [DOI] [PubMed] [Google Scholar]
- 50.Park JS, Woo DG, Yang HN, et al. Chondrogenesis of human mesenchymal stem cells encapsulated in a hydrogel construct: neocartilage formation in animal models as both mice and rabbits. Journal of Biomedical Materials Research—Part A. 2010;92(3):988–996. doi: 10.1002/jbm.a.32341. [DOI] [PubMed] [Google Scholar]
- 51.Koch TG, Berg LC, Betts DH. Current and future regenerative medicine-Principles, concepts, and therapeutic use of stem cell therapy and tissue engineering in equine medicine. Canadian Veterinary Journal. 2009;50(2):155–165. [PMC free article] [PubMed] [Google Scholar]
- 52.Tan H, Wu J, Lao L, Gao C. Gelatin/chitosan/hyaluronan scaffold integrated with PLGA microspheres for cartilage tissue engineering. Acta Biomaterialia. 2009;5(1):328–337. doi: 10.1016/j.actbio.2008.07.030. [DOI] [PubMed] [Google Scholar]
- 53.Vasilev K, Poh Z, Kant K, Chan J, Michelmore A, Losic D. Tailoring the surface functionalities of titania nanotube arrays. Biomaterials. 2010;31(3):532–540. doi: 10.1016/j.biomaterials.2009.09.074. [DOI] [PubMed] [Google Scholar]
- 54.Yoo HS, Kim TG, Park TG. Surface-functionalized electrospun nanofibers for tissue engineering and drug delivery. Advanced Drug Delivery Reviews. 2009;61(12):1033–1042. doi: 10.1016/j.addr.2009.07.007. [DOI] [PubMed] [Google Scholar]
- 55.Huang G, Zhou Z, Srinivasan R, et al. Affinity manipulation of surface-conjugated RGD peptide to modulate binding of liposomes to activated platelets. Biomaterials. 2008;29(11):1676–1685. doi: 10.1016/j.biomaterials.2007.12.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Mardilovich A, Craig JA, McCammon MQ, Garg A, Kokkoli E. Design of a novel fibronectin-mimetic peptide-amphiphile for functionalized biomaterials. Langmuir. 2006;22(7):3259–3264. doi: 10.1021/la052756n. [DOI] [PubMed] [Google Scholar]
- 57.Cressman S, Dobson I, Lee JB, Tam YYC, Cullis PR. Synthesis of a labeled RGD-lipid, its incorporation into liposomal nanoparticles, and their trafficking in cultured endothelial cells. Bioconjugate Chemistry. 2009;20(7):1404–1411. doi: 10.1021/bc900041f. [DOI] [PubMed] [Google Scholar]
- 58.Morgan AW, Roskov KE, Lin-Gibson S, Kaplan DL, Becker ML, Simon CG. Characterization and optimization of RGD-containing silk blends to support osteoblastic differentiation. Biomaterials. 2008;29(16):2556–2563. doi: 10.1016/j.biomaterials.2008.02.007. [DOI] [PubMed] [Google Scholar]
- 59.Chua PH, Neoh KG, Kang ET, Wang W. Surface functionalization of titanium with hyaluronic acid/chitosan polyelectrolyte multilayers and RGD for promoting osteoblast functions and inhibiting bacterial adhesion. Biomaterials. 2008;29(10):1412–1421. doi: 10.1016/j.biomaterials.2007.12.019. [DOI] [PubMed] [Google Scholar]
- 60.Shi Z, Neoh KG, Kang ET, Poh C, Wang W. Bacterial adhesion and osteoblast function on titanium with surface-grafted chitosan and immobilized RGD peptide. Journal of Biomedical Materials Research—Part A. 2008;86(4):865–872. doi: 10.1002/jbm.a.31648. [DOI] [PubMed] [Google Scholar]
- 61.Seo SJ, Kim IY, Choi YJ, Akaike T, Cho CS. Enhanced liver functions of hepatocytes cocultured with NIH 3T3 in the alginate/galactosylated chitosan scaffold. Biomaterials. 2006;27(8):1487–1495. doi: 10.1016/j.biomaterials.2005.09.018. [DOI] [PubMed] [Google Scholar]
- 62.Seo SJ, Choi YJ, Akaike T, Higuchi A, Cho CS. Alginate/galactosylated chitosan/heparin scaffold as a new synthetic extracellular matrix for hepatocytes. Tissue Engineering. 2006;12(1):33–44. doi: 10.1089/ten.2006.12.33. [DOI] [PubMed] [Google Scholar]
- 63.Seo SJ, Akaike T, Choi YJ, Shirakawa M, Kang IK, Cho CS. Alginate microcapsules prepared with xyloglucan as a synthetic extracellular matrix for hepatocyte attachment. Biomaterials. 2005;26(17):3607–3615. doi: 10.1016/j.biomaterials.2004.09.025. [DOI] [PubMed] [Google Scholar]
- 64.Seo SJ, Park IK, Yoo MK, Shirakawa M, Akaike T, Cho CS. Xyloglucan as a synthetic extracellular matrix for hepatocyte attachment. Journal of Biomaterials Science, Polymer Edition. 2004;15(11):1375–1387. doi: 10.1163/1568562042368059. [DOI] [PubMed] [Google Scholar]
- 65.Kim IY, Seo SJ, Moon HS, et al. Chitosan and its derivatives for tissue engineering applications. Biotechnology Advances. 2008;26(1):1–21. doi: 10.1016/j.biotechadv.2007.07.009. [DOI] [PubMed] [Google Scholar]
- 66.Park JS, Yang HN, Jeon SY, Woo DG, Na K, Park KH. Osteogenic differentiation of human mesenchymal stem cells using RGD-modified BMP-2 coated microspheres. Biomaterials. 2010;31(24):6239–6248. doi: 10.1016/j.biomaterials.2010.05.002. [DOI] [PubMed] [Google Scholar]
- 67.Tsuchiya K, Chen G, Ushida T, Matsuno T, Tateishi T. The effect of coculture of chondrocytes with mesenchymal stem cells on their cartilaginous phenotype in vitro. Materials Science and Engineering C. 2004;24(3):391–396. [Google Scholar]
- 68.Yang HN, Park JS, Na K, Woo DG, Kwon YD, Park KH. The use of green fluorescence gene (GFP)-modified rabbit mesenchymal stem cells (rMSCs) co-cultured with chondrocytes in hydrogel constructs to reveal the chondrogenesis of MSCs. Biomaterials. 2009;30(31):6374–6385. doi: 10.1016/j.biomaterials.2009.07.062. [DOI] [PubMed] [Google Scholar]
- 69.Chen J, Wang C, Lü S, et al. In vivo chondrogenesis of adult bone-marrow-derived autologous mesenchymal stem cells. Cell and Tissue Research. 2005;319(3):429–438. doi: 10.1007/s00441-004-1025-0. [DOI] [PubMed] [Google Scholar]
- 70.Worster AA, Brower-Toland BD, Fortier LA, Bent SJ, Williams J, Nixon AJ. Chondrocytic differentiation of mesenchymal stem cells sequentially exposed to transforming growth factor-β1 in monolayer and insulin-like growth factor-I in a three-dimensional matrix. Journal of Orthopaedic Research. 2001;19(4):738–749. doi: 10.1016/S0736-0266(00)00054-1. [DOI] [PubMed] [Google Scholar]
- 71.Bian L, Zhai DY, Mauck RL, Burdick JA. Coculture of human mesenchymal stem cells and articular chondrocytes reduces hypertrophy and enhances functional properties of engineered cartilage. Tissue Engineering—Part A. 2011;17(7-8):1137–1145. doi: 10.1089/ten.tea.2010.0531. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Varshney RR, Zhou R, Hao J, et al. Chondrogenesis of synovium-derived mesenchymal stem cells in gene-transferred co-culture system. Biomaterials. 2010;31(26):6876–6891. doi: 10.1016/j.biomaterials.2010.05.038. [DOI] [PubMed] [Google Scholar]