

Abstract
The paired box transcription factor Pax8 is critical for development of the eye, thyroid gland as well as the urinary and reproductive organs. In adult, Pax8 overexpression is associated with kidney, ovarian and thyroid tumors and has emerged as a specific marker for these cancers. Recently, Pax8 expression was also reported in human pancreatic islets and in neuroendocrine tumors, identifying Pax8 as a novel member of the Pax family expressed in the pancreas. Herein, we sought to provide a comprehensive analysis of Pax8 expression during pancreogenesis and in adult islets. Immunohistochemical analysis using the most employed Pax8 polyclonal antibody revealed strong nuclear staining in the developing mouse pancreas and in mature human and mouse islets. Astonishingly, Pax8 mRNA in mouse islets was undetectable while human islets exhibited low levels. These discrepancies raised the possibility of antibody cross-reactivity. This premise was confirmed by demonstrating that the polyclonal Pax8 antibody also recognized the islet-enriched Pax6 protein both by Western blotting and immunohistochemistry. Thus, in islets polyclonal Pax8 staining corresponds mainly to Pax6. In order to circumvent this caveat, a novel Pax8 monoclonal antibody was used to re-evaluate whether Pax8 was indeed expressed in islets. Surprisingly, Pax8 was not detected in neither the developing pancreas or in mature islets. Reappraisal of pancreatic neuroendocrine tumors using this Pax8 monoclonal antibody exhibited no immunostaining as compared to the Pax8 polyclonal antibody. In conclusion, Pax8 is not expressed in the pancreas and cast doubts on the value of Pax8 as a pancreatic neuroendocrine tumor marker.
Keywords: Pancreatic islet, Transcription factor, Pax genes, Neuroendocrine tumors, Antibody cross-reactivity
Introduction
A complex network of transcription factors controls pancreatic islet development as well as its subsequent mature function responsible for the exquisite maintenance of blood glucose homeostasis (Servitja and Ferrer 2004). Of particular interest is the family of paired box (Pax) genes, which encode key regulators of tissue development and cellular differentiation. The Pax transcription family comprises 9 members divided into 4 groups based on the specific assembly of 3 structural domains; the highly conserved 128 amino acid DNA binding paired domain, the octapeptide and a complete or residual DNA interacting homeodomain. Combinations of individual Pax members are spatially as well as temporally regulated during development and contribute to the formation of diverse organs (Lang et al. 2007).
Two members of this family, Pax6 and Pax4, comprising group IV and typified by the lack of the octapeptide domain, were shown to be essential for pancreatic islet organogenesis and function. Indeed, deletion of Pax4 in mice results in the absence of both β- and δ-cells with a concomitant increase in α-cells (Sosa-Pineda 2004). In contrast, deletion of Pax6 results in the absence of α-cells together with a decrease in the other types of pancreatic endocrine cells (St-Onge et al. 1997). Although β-, δ- and PP-cells are still present, they fail to form distinct islets and remain disorganize throughout the exocrine pancreas, suggesting that Pax6 is not only essential for α-cell differentiation but also for cell migration necessary for islet formation (St-Onge et al. 1997). Animals with both Pax6 and Pax4 ablation completely lack pancreatic islets demonstrating the absolute requirement of both Pax factors for islet generation.
Surprisingly Pax8, which is essential for development of the eye, thyroid gland as well as the urinary and reproductive organs, has recently emerged as a new member of the Pax family involved in islet physiology. Indeed, although previous reports have not detected Pax8 expression in pancreatic islets (Goode and Elgar 2009), more recent studies claim strong Pax8 immunostaining in pancreatic neuroendocrine tumors (Haynes et al. 2011; Long et al. 2010; Sangoi et al. 2011) as well as in normal human islets. Reported Pax8 expression in human pancreas is restricted to the endocrine compartment, as exocrine acinar and ductal cells exhibited no staining (Ozcan et al. 2011; Sangoi et al. 2011). Moreover, transcript expression profiling performed on islets isolated from pregnant mice revealed a robust induction of Pax8 expression levels during gestation (Rieck et al. 2009). In addition, a genome-wide linkage and admixture mapping study has linked Pax8 to Type 2 diabetes in African American families (Elbein et al. 2009). Taken together, these studies prompt us to investigate whether Pax8 could also be expressed during pancreatic islet development pinpointing to a potential new role of this transcription factor in islet physiology. To address this question, we analyzed Pax8 expression during development using a polyclonal Pax8 antibody previously utilized in several clinical studies that clearly demonstrated Pax8 staining in pancreatic neuroendocrine tumors (Haynes et al. 2011; Long et al. 2010; Sangoi et al. 2011). Consistent with these studies, strong Pax8 immunostaining was detected in the developing pancreas as well as in adult mouse and human islets of different ages. In contrast, Pax8 mRNA levels in both human and mouse islets were low to undetectable. In an attempt to confirm and validate the specificity of this Pax8 antibody, we found that it cross-reacted with the more abundant islet enriched Pax6 protein. In order to circumvent this caveat, we used a monoclonal antibody against Pax8 and re-evaluated whether Pax8 was indeed detected in islets. Astonishingly, we were unable to confirm expression of this Pax member neither in the developing pancreas, islets nor in pancreatic neuroendocrine tumors. Therefore, our results cast serious doubts on the value of Pax8 as a pancreatic neuroendocrine tumor marker and call for vigilance in the use of non-validated antibodies for clinical studies.
Materials and methods
Islet isolation and, culture
Pancreatic mouse islets were isolated by collagenase (Sigma-Aldrich Quimica S.A., Madrid, ES) digestion, handpicked and maintained in 11.1 mM glucose/RPMI-1640 (Invitrogen S.A., Barcelona, ES) supplemented with 10% fetal calf serum (Sigma-Aldrich Quimica S.A.), 100 Units/ml penicillin and 100 mg/ml streptomycin (Sigma-Aldrich Quimica S.A.), and 2 mM glutamine (GlutaMAX; Invitrogen S.A.). Isolated human islets obtained from the Cell Isolation and Transplantation Center, Geneva University Hospitals, were maintained in CMRL 1066 medium w/o phenol red (Sigma-Aldrich Quimica S.A.) supplemented with 10% fetal calf serum (Sigma-Aldrich Quimica S.A.), 100 Units/ml penicillin and 100 mg/ml streptomycin (Sigma-Aldrich Quimica S.A.), 100 mg/ml gentamycin (Sigma-Aldrich Quimica S.A.) and 2 mM glutamine (GlutaMAX, Invitrogen S.A.).
Tissue samples and preparation
Embryonic, pup and adult mouse tissues were dissected and fixed in 4% paraformaldehyde overnight at 4°C, embedded in paraffin and sectioned at 7 μm. Sections from biopsies of human pancreatic neuroendocrine tumor were kindly provided by the Hospital Universitario Virgen del Rocio (Sevilla, Spain). Sections included both neuroendocrine tumor and neighbouring non-neoplastic areas.
Quantitative real time PCR (Q-RT-PCR)
Total RNA from islets was extracted using the RNeasy Micro Kit (Qiagen, Madrid, ES) and Q-RT-PCRs performed as described previously (Hu He et al. 2011). Briefly, 2 μg RNA were used to synthesize first-strand cDNA using Superscript II (Invitrogen S.A.) in a total volume of 20 μL. 3 μL of diluted cDNA samples were used as template for the Q-RT-PCR using SYBR green (Roche Farma S.A., Madrid, ES). Q-RT-PCR data were normalized using the 2–∆Ct method for transcripts levels evaluated in different tissues. In situation in which transcript levels were derived from the same tissue, the 2∆∆Ct method was employed and normalized to Pax6 expression levels. Briefly, the ∆∆Ct is calculated as the Pax6 ∆Ct minus the ∆Ct of the gene of interest. The ∆Ct is calculated as the crossing point of the housekeeping gene (cyclophilin) minus the crossing point of the gene of interest. Primer sequences listed in Table 1 were designed using the Primer3 Software (http://frodo.wi.mit.edu/primer3/).
Table1.
Sequence of the primers used in the study
| Gene | Primers (5′–3′) | |
|---|---|---|
| Human | ||
| Pax4 | Forw. | ACCCCACCTAAAGCCTGTCT |
| Rev. | AGGCAAAGCAGTCCTGAGTC | |
| Pax6 | Forw. | CTCGGGCACCACTTCAACA |
| Rev. | TCCGGGAACTTGAACTGGAA | |
| Pax8 | Forw. | CCCCCTACTCCTCCTACAGC |
| Rev. | GGCCTTGATGTGGAACTGTAA | |
| Cyclophilin | Forw. | TACGGGTCCTGGCATCTTGT |
| Rev. | CCATTTGTGTTGGGTCCAGC | |
| Mouse | ||
| Pax4 | Forw. | TCGAATTGCCCAGCTAAAGG |
| Rev. | TTCAGTGCAAAGCTGGTGTTG | |
| Pax6 | Forw. | CAACCTGGCTAGCGAAAAGC |
| Rev. | TCTGCCCGTTCAACATCCTT | |
| Pax8 | Forw. | GTTTGAGCGGCAGCATTAC |
| Rev. | GTAAGGGCAGTGGGTACAGC | |
| Cyclophilin | Forw. | ATGGCAAATGCTGGACCAA |
| Rev. | GCCATCCAGCCATTCAGTCT | |
Transient transfection and immunoblot analysis
Transient transfection of human Pax4 (Brun et al. 2008), Pax6 and Pax8 (Origene Technologies, Rockville USA) expressing plasmids were performed using Lipofectamine 2000 (Invitrogen S.A.) following the manufacturer’s protocol. Briefly, MCF-7 cells plated on six multiwell plates were transfected with 4 μg DNA. 24 h after transfection cells were harvested and whole-cell extracts were prepared by lysing the cells at 4°C for 1 h in WCB (20 mM Hepes (pH 7.7), 300 mM NaCl, 0.2 mM EDTA, 0.1% Triton X-100, 1.5 mM MgCl2, 0.5 mM DTT) supplemented with protease inhibitors. 15 μg of protein extracts were resolved in SDS-PAGE gels and transferred to PVDF membranes. Membranes were blocked with 5% milk in Tris-buffered saline—0.1% Tween (TBST) and subsequently incubated overnight at 4°C with the primary antibody in TBST containing 3% BSA. The membranes were then incubated with the corresponding secondary antibody in TBST-3% nonfat milk for 1 h at room temperature and the immunoreactive bands were visualized using the enhanced chemiluminescence detection system (ECL) (GE-Healthcare, Barcelona ES). Specific antibodies were used as follow: Polyclonal Pax8 (No 10336-1-AP ProteinTech Group, Chicago, USA) 1:500 dilution; monoclonal Pax8 (No ab53490, Abcam, Cambridge UK) 1:100 dilution; monoclonal Pax6 (DSHB-University of Iowa, USA) 1:1,000 dilution; c-Myc (Sigma-Aldrich Quimica S.A.) 1 μg/mL; α-tubulin (Sigma-Aldrich Quimica S.A.) 1:5,000 dilution.
Immunostaining
Immunohistochemistry analysis was performed as previously described (Rojas et al. 2009). Briefly, subsequent to antigen retrieval by autoclaving in citrate buffer for mouse tissues or in EDTA buffer using PT-Link (DAKO, Barcelona, ES) for human sections; tissue samples were blocked in PBS-3% BSA for 1 h. Overnight incubation with primary antibodies (rabbit polyclonal Pax8 antibody at 1:400 dilution, mouse monoclonal Pax8 at 1:10 dilution and monoclonal Pax6 antibody at 1:800 dilution) was performed at 4°C. Corresponding biotinylated secondary antibodies (dilution 1:300) were used for 1 h at room temperature and DAB (Vector Lab, Barcelona, ES) was used for signal development. Counterstaining with Hematoxilin (Panreac, Barcelona, ES) was performed.
Alternatively, for immunofluorescence, cells grown on coverslips were fixed 10 min at room temperature in 4% paraformaldehyde and methanol for 2 min at −20°C. Cells were then permeabilized with PBS containing 0.1% Tween-20 and 2% BSA and blocked in 5% BSA in TBS-0.1% Triton X-100. Primary antibodies, rabbit polyclonal Pax8 antibody at 1:800 dilution, a mouse monoclonal Pax8 at 1:10 dilution and a monoclonal Pax6 antibody at 1:800 dilution, were used overnight at 4°C. Secondary antibodies used were either rabbit or mouse Alexa Fluor 568 and Alexa Fluor 488. Counterstaining with DAPI was performed in order to reveal nuclei. The slides were analyzed using epifluorescence microscopy (Leica DM6000B, Leica).
Results
Pax8 transcript levels are low to undetectable in adult pancreatic islets
Recent studies revealing Pax8 staining in human pancreatic neuroendocrine tumors as well as in normal adult islets prompted us to evaluate expression levels of this transcription factor in human and mouse islets as compared to Pax6 and Pax4, two Pax family members expressed in the endocrine pancreas. Consistent with previous reports (Brun et al. 2004; Hu He et al. 2011; Kutlu et al. 2009), Pax6 was highly expressed in both human and mouse islets whereas Pax4 levels were much lower, nonetheless detectable (Fig. 1). In contrast, the Pax8 transcript was not detected in mouse islets whereas human islets exhibited low levels as compared to Pax6 and Pax4 (Fig. 1). As several Pax8 splice variants have been reported (Kozmik et al. 1993), various combinations of primers were evaluated to eliminate the possibility that only a subset of these spliced transcripts were expressed in islets. Astonishingly, identical results were obtained independently of the primer pairs used (Data not shown).
Fig. 1.
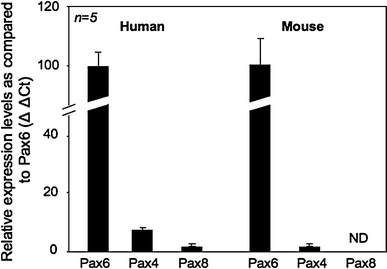
Pax8 transcript levels are low to undetectable in adult human and mouse islets as compared to Pax6 and Pax4. Quantitative RT-PCR using RNA purified from freshly isolated mouse and human islets were performed on Pax8, Pax6, Pax4 and the housekeeping gene cyclophilin. The Pax4 and Pax8 relative mRNA abundance levels were established using the 2∆∆Ct method and compared to the expression levels of Pax6 (arbitrarily indicated as 100). Results represent the average of at least five experiments performed in duplicate ± SE (ND non detected)
A Pax8 polyclonal antibody reveals staining in the developing mouse pancreas and in neonatal islets
Low levels of the Pax8 transcript in adult islets led us to investigate whether Pax8 expression could be temporally expressed during development. To address this question, we performed immunohistochemical analysis on kidney, pancreas and liver of mouse embryos at embryonic stage (E)15.5 and E17.5 as well as newborn pups using the most cited Pax8 polyclonal antibody. We opted for this approach in order to accurately determine whether Pax8 is specifically expressed in the developing pancreas. Consistent with a previous study detecting Pax8 transcript (Plachov et al. 1990), strong Pax8 nuclear immunostaining was observed in the endodermal compartment of both the developing and newborn kidney (Fig. 2A; a, d and g). In the liver, Pax8 staining was only detected in scattered cells (Fig. 2A; c, f and i). Consistent with our hypothesis, the developing pancreas also exhibited Pax8 staining in the endocrine compartment while the surrounding exocrine tissue displayed no expression of the transcription factor (Fig. 2A; b and e). Intriguingly, Pax8 immunostaining also persisted in the majority of islets cells of neonatal animals (Fig. 2A; h). These findings were substantiated by immunofluorescence analysis (Fig. 2B).
Fig. 2.
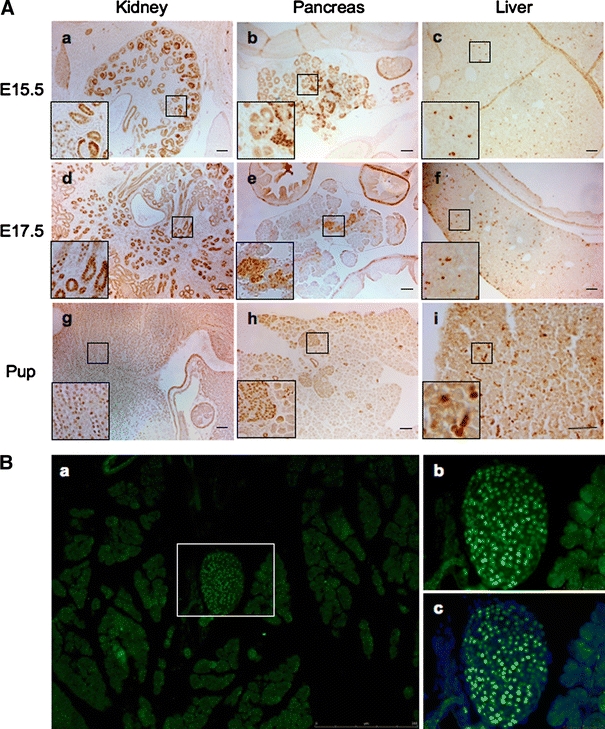
A polyclonal Pax8 antibody reveals expression of the transcription factor in kidney, pancreas and liver during development. A Immunohistochemistry staining using a polyclonal Pax8 antibody shows strong nuclear staining in kidney and pancreas and in scattered cells in liver during embryonic development (a–f) and in new born pups (g–i) (scale bar 50 μm). B Immunofluorescence images of pancreatic tissue from neonatal animal incubated with polyclonal Pax8 (green) exhibiting strong Pax8 nuclear staining in the islets while the surrounding exocrine tissue is negative. b and c are the enlarged detail of the islet indicated in a, showing in c the co-staining with DAPI (blue) to reveal the totality of islet cells
The Pax8 polyclonal antibody reveals immunostaining in the adult pancreas
Detection of Pax8 positive cells in neonatal islets using a polyclonal antibody prompt us to investigate whether the protein could also be detected in adult islets. Consistent with the immunostaining observed in embryos and newborn mice, Pax8 was also detected in most cells of adult mouse and human islets (Fig. 3A). Surprisingly, these results are in sharp contrast to our gene expression analysis in which the Pax8 transcript was not detected in adult mouse islets (Fig. 1). In order to verify these results, immunohistochemistry analysis were repeated using a recently commercialized Pax8 monoclonal antibody generated against the C-terminal region of Pax8. In contrast to the polyclonal antibody, the monoclonal Pax8 antibody revealed no staining in either islets from adult mice or human (Fig. 3B). Discrepancies in the immunohistochemical data combined with undetectable Pax8 transcripts in islets raise the possibility that the Pax8 polyclonal antibody may cross-react with other pancreatic islet-enriched Pax members.
Fig. 3.
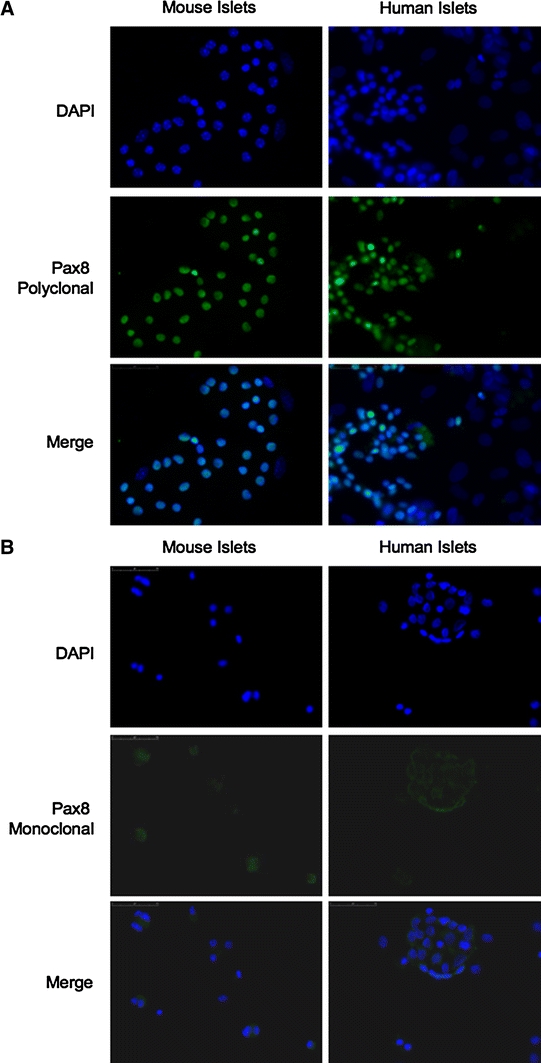
A polyclonal but not a monoclonal antibody against Pax8 immunostains adult mouse or human islets. A Immunofluorescence analysis of disaggregated mouse (left panels) and human (right panels) islet cells (×400) using a polyclonal Pax8 antibody (green) shows, in the majority of islet cells, a strong nuclear Pax8 staining. B No immunostaining was detected either in mouse (left panels) or human (right panels) islets when a monoclonal Pax8 antibody (green) was used (×400). For the visualization of all cells, counterstaining with DAPI (blue) was performed
The polyclonal Pax8 antibody cross-reacts with the islet-enriched Pax6 family member
To authenticate or rebut the specificity of the Pax8 polyclonal antibody, we performed Western blotting analysis on cells transfected with either Pax6 or Pax4 expressing vectors. These two Pax members are expressed in both the developing pancreas as well as in mature islets (Brun et al. 2004, 2008; Sosa-Pineda et al. 1997; St-Onge et al. 1997) (Fig. 1). In order to identify the most suitable experimental cell model and exclude cross reactivity with other potential proteins, we initially screened several cell lines for the expression of Pax8 using Q-RT-PCR. The Pax8 transcript was not detected in the human epithelial carcinoma Hela cell line, the human breast cancer MCF7 cell line or in the rat insulinoma INS-1E cell line (Fig. 4A). However, immunofluorescence and Western blot analysis using the polyclonal Pax8 antibody revealed strong positive staining in both HeLa and INS-1E, further indicating a possible non-specificity of the polyclonal antibody (Fig. 4B, C). In contrast, MCF7 cells were negative for Pax8 staining (Fig. 4B, C) and we therefore selected this cell line to further study the specificity of the polyclonal Pax8 antibody. Protein extracts derived from MCF7 cells transfected with human Pax4, Pax6 or Pax8 expression vectors were blotted with an anti-myc (Pax4) and an anti-Pax6 serum as well as with the polyclonal Pax8 antibodies. The anti-myc and anti-Pax6 sera specifically detected Pax4 and Pax6, respectively. However, although the polyclonal Pax8 antibody revealed a strong signal in Pax8-tranfected cells, it also recognized, albeit with lower intensity, a band of similar molecular weight in Pax6-transfected MCF7 cells (Fig. 5A). In contrast, the Pax8 monoclonal antibody exclusively recognized a band in MCF7 cells transfected with Pax8 (Fig. 5A). Immunofluorescence analysis performed on Pax-transfected MCF7 cells confirmed that the polyclonal Pax8 antibody recognized both Pax6 and Pax8 while the monoclonal antibody was specific to Pax8 (Fig. 5B).
Fig. 4.
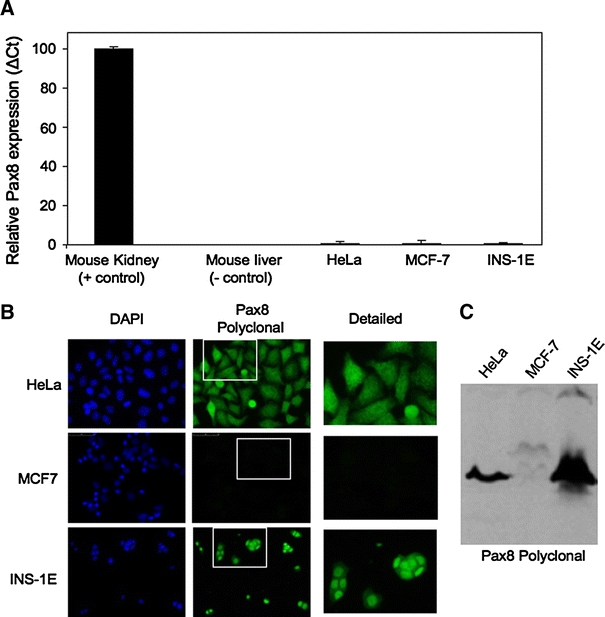
The Pax8 protein but not the transcript is detected in various cell lines. A Q-RT-PCR analysis shows nearly non-detectable expression levels of Pax8 in HeLa, MCF-7 and INS-1E cell lines. The graphic represents the relative abundance of Pax8 mRNA in the different cell lines as compared to the expression levels in kidney (arbitrarily indicated as 100). B Immunofluorescence analysis of the same cell lines shows that only MCF-7 cell line is negative for immunostaining when using the Pax8 polyclonal antibody (green) (middle panel). Counterstaining with DAPI (blue) is shown in the left panels (×400). A detailed image corresponding to an enlargement of the indicated area is showed in the right panels. C Western blot analysis of cell extracts using the polyclonal Pax8 antibody revealed the presence of an immunoreactive band of approximately 48 kD (corresponding to the molecular weight of Pax8) in all cell lines except MCF-7 cells
Fig. 5.
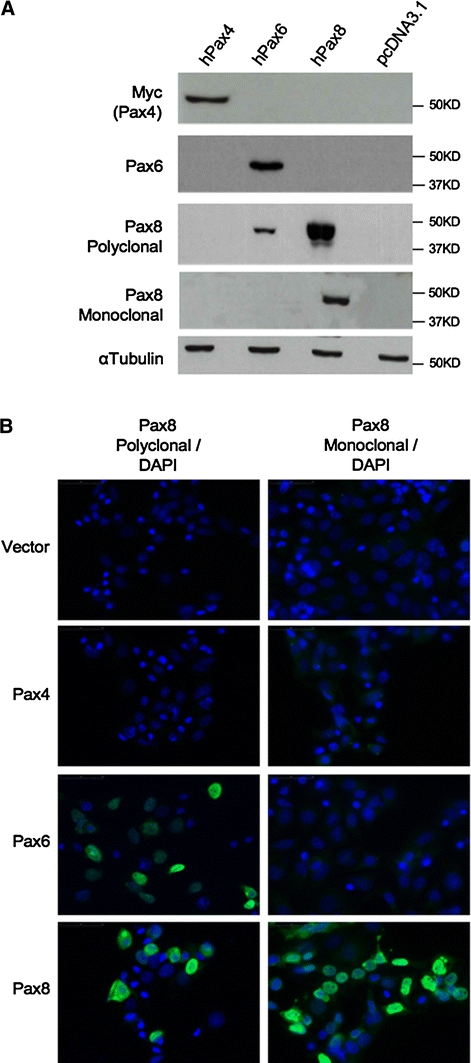
The polyclonal Pax8 antibody cross-reacts with Pax6. A Western blot analysis of extracts from MCF-7 cells transfected with Pax4, Pax6 or Pax8 expression vectors using c-Myc (for the detection of Pax4), Pax6, polyclonal Pax8 and monoclonal Pax8 antibodies. Pax8 polyclonal antibody detects not only Pax8 (lane 3) but also Pax6 (lane 2), indicating a cross-reactivity of this antibody with Pax6. The other three antibodies used, including a new monoclonal Pax8 antibody, are specific. α-tubulin staining was used for loading control. B Immunofluorescence analysis of transfected MCF-7 cells from A, using either the polyclonal (left panels) or a new monoclonal (right panels) Pax8 antibodies (green). No immunostaining was detected in Pax4-transfected MCF-7 cells with any of the Pax8 antibodies. However, polyclonal Pax8 antibody is able to detect exogenous Pax6, indicating the existence of cross-reactivity of polyclonal Pax8 antibody with Pax6 protein. Counterstaining with DAPI (blue) is shown to visualize also the non-transfected cells
The polyclonal Pax8 antibody detects Pax6 in islets as well as in the developing pancreas
Our findings indicating cross-reactivity of Pax8 polyclonal antibody with Pax6 raise the possibility that the signal detected in islet is not Pax8 but Pax6. To verify this possibility, co-immunofluorescence studies were performed on both human and mouse disaggregated islets using the polyclonal Pax8 antibody together with the anti-Pax6 sera. Consistent with our hypothesis, all Pax8-positive cells were also Pax6-positive and single staining for either Pax8 or Pax6 was never observed in either human or mouse islets (Fig. 6). These results suggest that Pax8, as initially reported (Plachov et al. 1990), is most likely not expressed in the mature pancreas. The latter conclusion, prompt us to re-appraise whether Pax8 was expressed during pancreas development. Consecutive sections from E17.5 embryos as well as from newborn pups and adult pancreas were analyzed by immunohistochemistry using the polyclonal and monoclonal Pax8 antibodies or the Pax6 monoclonal antibody. Strikingly, overlapping expression patterns within the developing pancreas and islets were again observed with the polyclonal Pax8 and Pax6 antibodies (Fig. 7, compare images from a, d and g to b, e and h). In contrast, the Pax8 monoclonal antibody revealed no signal either in the developing pancreas or in adult islets (Fig. 7c, f, i). These results clearly demonstrate that the Pax8 polyclonal antibody cross-reacts with Pax6.
Fig. 6.
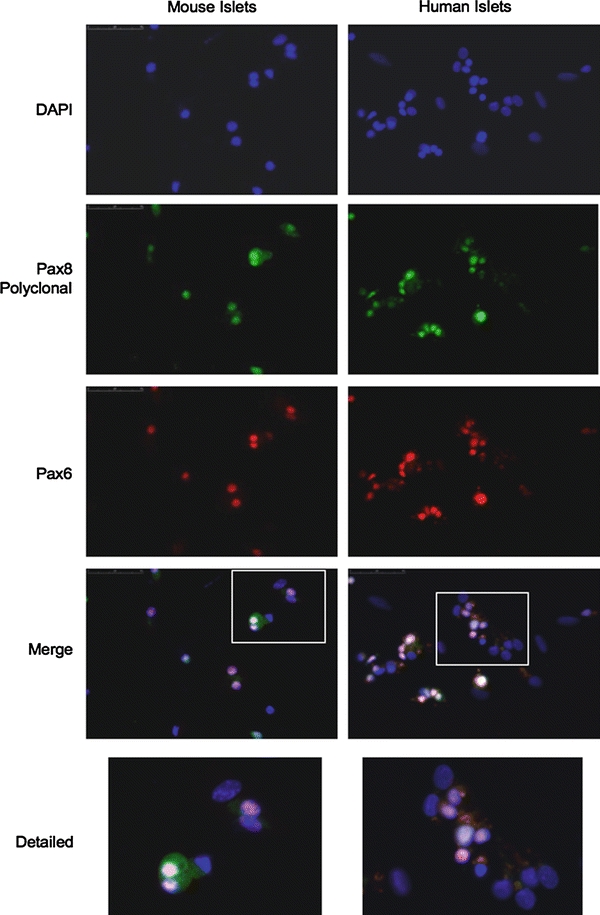
The Pax8 polyclonal antibody immunostains Pax6 in both human and mouse adult islets. Immunofluorescence analysis of disaggregated mouse (left panels) and human (right panels) pancreatic islet cells co-immunostained with polyclonal Pax8 (green) and Pax6 (red) antibodies, shows an overlap in the staining both in human and in mouse islets (×400). The enlargement of the indicated area (lower panels) shows this overlap. Counterstaining with DAPI (blue) is also shown
Fig. 7.
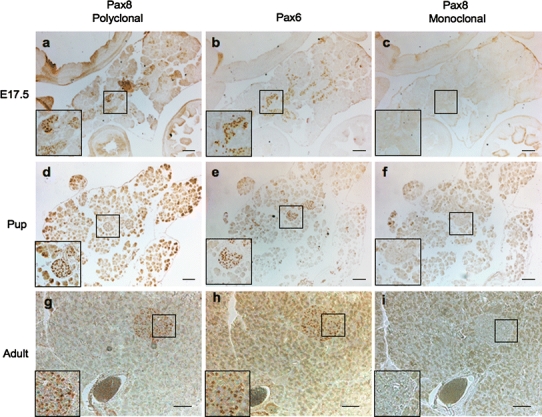
Pax8 is not expressed in the developing mouse pancreas or in mature islets. Immunostaining of pancreatic consecutive sections using either the polyclonal (left panels) or monoclonal (right panels) Pax8 antibody, or the Pax6 antibody (middle panels) reveals an overlapping signal between Pax6 and the polyclonal Pax8 antibodies. The monoclonal Pax8 antibody does not stain mouse islets at any studied age. a–c E17.5, d–f Pup and g–i adult mice. Scale bar 50 μm
Pax8 is not expressed in pancreatic neuroendocrine tumors
A crucial question that stems from our study is whether Pax8 is truly expressed in pancreatic neuroendocrine tumors as claimed by several independent studies, all using the same Pax8 polyclonal antibody (Haynes et al. 2011; Long et al. 2010; Sangoi et al. 2011). To address this important issue, serial sections of pancreatic neuroendocrine tumors obtained from 9 independent donors were immunostained with Pax8 antibodies (polyclonal and monoclonal) as well as with the anti-Pax6 sera. Consistent with previous reports, the Pax8 polyclonal antibody revealed positive staining in some but not all tumors. Indeed, seven out of nine tumors were positive using this antibody (two representative tumors are shown in Fig. 8A). However, this staining was not confirmed using the more specific monoclonal Pax8 antibody (Fig. 8A, compare a and d, to c and f). In parallel, human kidney and liver samples were used as positive and negative controls for Pax8 expression. Both Pax8 antibodies exhibited strong nuclear staining in kidney sections while no signal was detected in liver sections (Fig. 8B, C). Interestingly, 3 out of 7 samples displayed strong positive signal for Pax8 polyclonal antibody as well as for Pax6 antibody suggesting that the signal given by the Pax8 polyclonal antibody is due, at least in part, to the recognition of the endogenous Pax6 protein (Fig. 8A, upper panels). However, in the four remaining tumors that reacted with the Pax8 polyclonal antibody, no staining was detected for Pax6 (Fig. 8A, compare d and e).
Fig. 8.
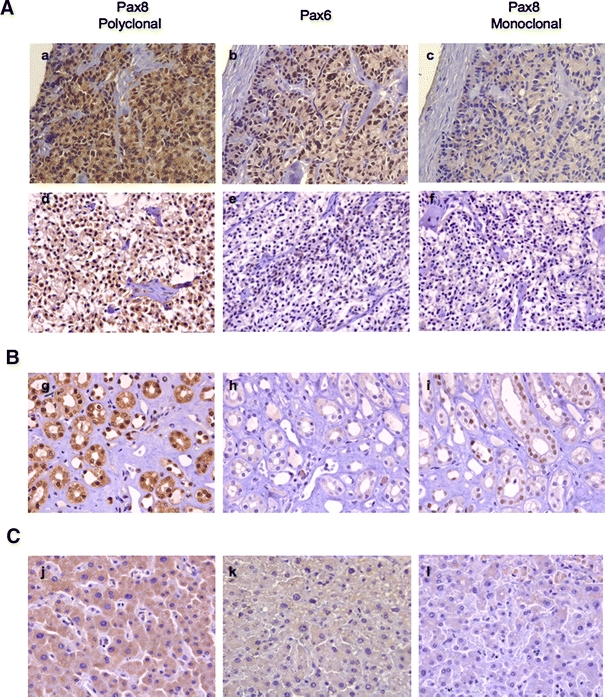
Pax8 is not expressed in human pancreatic neuroendocrine tumors. A Pax8 or Pax6 immunostaining in two different pancreatic neuroendocrine tumors using polyclonal (a, d) or monoclonal (c, f) Pax8 antibody, or Pax6 antibody (b, e) reveals the lack of Pax8 staining when Pax8-specific monoclonal antibody is used. Of note, the lower panel tumor is also negative for Pax6. B Pax8 or Pax6 immunostaining in human kidney and C liver using polyclonal (g, j) or monoclonal (i, l) Pax8 antibody, or Pax6 antibody (h, k). An overlap in the immunostaining signal between the two Pax8 antibodies in kidney tubule epithelia, indicates the expression of Pax8 in this tissue. No nuclear immunostaining for Pax6 was detected in kidney. Pax8 or Pax6 antibodies revealed no signals in the liver
Discussion
The aim of the current study was to establish whether Pax8 was expressed during pancreas development. The rational was to complement recent clinical studies demonstrating the presence of this transcription factor in normal human islets (Ozcan et al. 2011; Sangoi et al. 2011) as well as in pancreatic neuroendocrine tumors (Haynes et al. 2011; Long et al. 2010; Sangoi et al. 2011). However, our Q-RT-PCR data does not substantiate these studies as Pax8 transcript was not detected in mouse islets and was barely detectable in human islets. Furthermore, our results reveal that the most commonly used Pax8 polyclonal antibody cross-reacts with Pax6 in normal islets and pancreatic neuroendocrine tumors. The lack of Pax8 immunodetection in the developing pancreas, islets or pancreatic neuroendocrine tumors using a more specific Pax8 monoclonal antibody suggest that this Pax family member may not be expressed in the pancreas under normal physiological or pathophysiological conditions. More importantly, our results highlight the essentiality of validating the specificity of antibodies before embarking into large immunohistochemical studies such as the ones performed for Pax8 (Haynes et al. 2011; Long et al. 2010; Sangoi et al. 2011). Our results also cast serious concerns on the reliability of using Pax8 as a marker to identify pancreatic neuroendocrine tumors as proposed by Sangoi et al. (2011).
What could be the cause of such cross-reactivity? A plausible explanation may be linked to the peptide antigen used to generate the Pax8 polyclonal antibody. Indeed this antibody was raised against a 212 amino acids (aa) polypeptide encompassing the N-terminal region of Pax8. This region includes the notoriously conserved 128 aa paired DNA binding domain found in all members of the pax gene family (Lang et al. 2007; Robson et al. 2006). Thus, immunization using this region will likely produce a multitude of antibodies that will recognize several Pax members, including Pax6. Further substantiating the promiscuity of the Pax8 polyclonal antibody, we also found that this antibody but not the Pax6 or Pax8 monoclonal antibodies immunostained four tumor samples suggesting cross reactivity with yet another potential Pax member. As Pax8 along with Pax5 and Pax2 define group II of the Pax family with over 95% protein homology in the common paired DNA binding domain, it is tempting to speculate that Pax5 or Pax2 are potential culprits. Consistent with this premise, Sangoi and colleagues recently reported Pax8 immunoreactivity in hematopoietic neoplasms, which they attributed to cross-reactivity with Pax5, a well-defined marker of B-lineage hematologic malignancies (Sangoi et al. 2010). Interestingly, we detected Pax8 staining in the developing mouse liver using the polyclonal antibody (Fig. 2c, f, i). This organ is the major site of hematopoiesis during murine embryonic development (Johnson and Moore 1975) and therefore expresses high levels of Pax5. The latter may therefore offer an explanation for false Pax8 staining in the liver. A similar case of promiscuity in Pax family immunoreactivity was recently described for a Pax5 monoclonal antibody that was found to cross-react with its close relative Pax2 in nonhematopoietic tissues (Morgenstern et al. 2010). Interestingly, we have previously demonstrated that Pax2 is expressed in the endocrine pancreas (Ritz-Laser et al. 2000) raising the possibility that the Pax8 polyclonal antibody may also recognize this Pax member. Thus, therefore, a panel of antibodies against different Pax members should be screened to determine which Pax proteins are aberrantly expressed in pancreatic neuroendocrine tumors. Moreover, validation of the specificity with at least two antibodies raised against different epitopes would be highly recommended prior to launching large-scale clinical studies.
In contrast to the Pax8 polyclonal antibody, the immunogen that was used to create the monoclonal antibody was taken from aa 318–426 of the transcription factor. This region, located at the carboxy-terminal end of the protein, is highly divergent among Pax members and should therefore exhibit target-specific recognition. Nonetheless, despite the lack of Pax8 immunodetection in normal pancreatic islets using this monoclonal antibody, Pax8 transcripts were detected, albeit at low levels, in human islets suggesting a possible involvement of this transcription factor in islet physiology. Similar to Pax4, Pax8 expression may be induced in response to physiological situations that requires β-cell mass expansion (Brun and Gauthier 2008; Hu He et al. 2011). Consistent with this idea, expression levels of Pax8 were shown to increase during pregnancy in mice (Rieck et al. 2009).
In summary, this study highlights pitfalls of using ill-defined antibodies as markers to either characterize or classify human tumors. It is of utmost importance to validate the specificity of antibodies generated against proteins, which have high homology with other members of the same family. In this regard, caution should be taken on whether Pax8 is a reliable marker for pancreatic neuroendocrine tumors.
Acknowledgments
We acknowledge the technical assistance of J. Vallejo Ortega. This work was supported by grants from the Fundacion Progreso y Salud (to B.R.G.), the Spanish Ministry of Science and Innovation, Instituto de Salud Carlos III (PI10/00871 to B.R.G) and the Juvenile Diabetes Research Foundation (9-2004-384 to E.C.I.T Plan Nacional de I+D+I 2008-2011, ISCIII (PI08/0018) co-funded by Fondos FEDER (to A.R.) and Consejería de Salud, Junta de Andalucía (PI0008 to A.R.).
Open Access
This article is distributed under the terms of the Creative Commons Attribution Noncommercial License which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
References
- Brun T, Franklin I, St-Onge L, Biason-Lauber A, Schoenle E, Wollheim CB, Gauthier BR. The diabetes-linked transcription factor Pax4 promotes beta-cell proliferation and survival in rat and human islets. J Cell Biol. 2004;167:1123–1135. doi: 10.1083/jcb.200405148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brun T, Gauthier BR. A focus on the role of Pax4 in mature pancreatic islet beta-cell expansion and survival in health and disease. J Mol Endocrinol. 2008;40:37–45. doi: 10.1677/JME-07-0134. [DOI] [PubMed] [Google Scholar]
- Brun T, He KH, Lupi R, Boehm B, Wojtusciszyn A, Sauter N, Donath M, Marchetti P, Maedler K, Gauthier BR. The diabetes-linked transcription factor Pax4 is expressed in human pancreatic islets and is activated by mitogens and GLP-1. Hum Mol Genet. 2008;17:478–489. doi: 10.1093/hmg/ddm325. [DOI] [PubMed] [Google Scholar]
- Elbein SC, Das SK, Hallman DM, Hanis CL, Hasstedt SJ. Genome-wide linkage and admixture mapping of type 2 diabetes in African American families from the American Diabetes Association GENNID (Genetics of NIDDM) Study Cohort. Diabetes. 2009;58:268–274. doi: 10.2337/db08-0931. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goode DK, Elgar G. The PAX258 gene subfamily: a comparative perspective. Dev Dyn. 2009;238:2951–2974. doi: 10.1002/dvdy.22146. [DOI] [PubMed] [Google Scholar]
- Haynes CM, Sangoi AR, Pai RK. PAX8 is expressed in pancreatic well-differentiated neuroendocrine tumors and in extrapancreatic poorly differentiated neuroendocrine carcinomas in fine-needle aspiration biopsy specimens. Cancer Cytopathol. 2011;119:193–201. doi: 10.1002/cncy.20136. [DOI] [PubMed] [Google Scholar]
- Hu He KH, Lorenzo PI, Brun T, Jimenez Moreno CM, Aeberhard D, Ortega JV, Cornu M, Thorel F, Gjinovci A, Thorens B, Herrera PL, Meda P, Wollheim CB, Gauthier BR. In Vivo Conditional Pax4 Overexpression in mature islet {beta}-cells prevents stress-induced hyperglycemia in mice. Diabetes. 2011;60:1705–1715. doi: 10.2337/db10-1102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnson GR, Moore MA. Role of stem cell migration in initiation of mouse foetal liver haemopoiesis. Nature. 1975;258:726–728. doi: 10.1038/258726a0. [DOI] [PubMed] [Google Scholar]
- Kozmik Z, Kurzbauer R, Dorfler P, Busslinger M. Alternative splicing of Pax-8 gene transcripts is developmentally regulated and generates isoforms with different transactivation properties. Mol Cell Biol. 1993;13:6024–6035. doi: 10.1128/mcb.13.10.6024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kutlu B, Burdick D, Baxter D, Rasschaert J, Flamez D, Eizirik DL, Welsh N, Goodman N, Hood L. Detailed transcriptome atlas of the pancreatic beta cell. BMC Med Genom. 2009;2:3. doi: 10.1186/1755-8794-2-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lang D, Powell SK, Plummer RS, Young KP, Ruggeri BA. PAX genes: roles in development, pathophysiology, and cancer. Biochem Pharmacol. 2007;73:1–14. doi: 10.1016/j.bcp.2006.06.024. [DOI] [PubMed] [Google Scholar]
- Long KB, Srivastava A, Hirsch MS, Hornick JL. PAX8 Expression in well-differentiated pancreatic endocrine tumors: correlation with clinicopathologic features and comparison with gastrointestinal and pulmonary carcinoid tumors. Am J Surg Pathol. 2010;34:723–729. doi: 10.1097/PAS.0b013e3181d9f18f. [DOI] [PubMed] [Google Scholar]
- Morgenstern DA, Hasan F, Gibson S, Winyard P, Sebire NJ, Anderson J. PAX5 expression in nonhematopoietic tissues. Reappraisal of previous studies. Am J Clin Pathol. 2010;133:407–415. doi: 10.1309/AJCPZPQN0LUGKMME. [DOI] [PubMed] [Google Scholar]
- Ozcan A, Shen SS, Hamilton C, Anjana K, Coffey D, Krishnan B, Truong LD. PAX 8 expression in non-neoplastic tissues, primary tumors, and metastatic tumors: a comprehensive immunohistochemical study. Mod Pathol. 2011;24:751–764. doi: 10.1038/modpathol.2011.3. [DOI] [PubMed] [Google Scholar]
- Plachov D, Chowdhury K, Walther C, Simon D, Guenet JL, Gruss P. Pax8, a murine paired box gene expressed in the developing excretory system and thyroid gland. Development. 1990;110:643–651. doi: 10.1242/dev.110.2.643. [DOI] [PubMed] [Google Scholar]
- Rieck S, White P, Schug J, Fox AJ, Smirnova O, Gao N, Gupta RK, Wang ZV, Scherer PE, Keller MP, Attie AD, Kaestner KH. The transcriptional response of the islet to pregnancy in mice. Mol Endocrinol. 2009;23:1702–1712. doi: 10.1210/me.2009-0144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ritz-Laser B, Estreicher A, Gauthier B, Philippe J. The paired-homeodomain transcription factor Pax-2 is expressed in the endocrine pancreas and transactivates the glucagon gene promoter. J Biol Chem. 2000;275:32708–32715. doi: 10.1074/jbc.M005704200. [DOI] [PubMed] [Google Scholar]
- Robson EJ, He SJ, Eccles MR. A PANorama of PAX genes in cancer and development. Nat Rev Cancer. 2006;6:52–62. doi: 10.1038/nrc1778. [DOI] [PubMed] [Google Scholar]
- Rojas A, Schachterle W, Xu SM, Black BL. An endoderm-specific transcriptional enhancer from the mouse Gata4 gene requires GATA and homeodomain protein-binding sites for function in vivo. Dev Dyn. 2009;238:2588–2598. doi: 10.1002/dvdy.22091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sangoi AR, Ohgami RS, Pai RK, Beck AH, McKenney JK. PAX8 expression reliably distinguishes pancreatic well-differentiated neuroendocrine tumors from ileal and pulmonary well-differentiated neuroendocrine tumors and pancreatic acinar cell carcinoma. Mod Pathol. 2011;24:412–424. doi: 10.1038/modpathol.2010.176. [DOI] [PubMed] [Google Scholar]
- Sangoi AR, West RB, Bonventre JV, Montgomery KD, McKenney JK. Exploring the specificity of putative renal cell carcinoma (RCC) markers in non-renal tissues and neoplasms from various organ systems: a tissue microarray (TMA) study of 501 cases. Mod Pathol. 2010;23:216A. [Google Scholar]
- Servitja JM, Ferrer J. Transcriptional networks controlling pancreatic development and beta cell function. Diabetologia. 2004;47:597–613. doi: 10.1007/s00125-004-1368-9. [DOI] [PubMed] [Google Scholar]
- Sosa-Pineda B. The gene Pax4 is an essential regulator of pancreatic beta-cell development. Mol Cells. 2004;18:289–294. [PubMed] [Google Scholar]
- Sosa-Pineda B, Chowdhury K, Torres M, Oliver G, Gruss P. The Pax4 gene is essential for differentiation of insulin-producing beta cells in the mammalian pancreas. Nature. 1997;386:399–402. doi: 10.1038/386399a0. [DOI] [PubMed] [Google Scholar]
- St-Onge L, Sosa-Pineda B, Chowdhury K, Mansouri A, Gruss P. Pax6 is required for differentiation of glucagon-producing alpha-cells in mouse pancreas. Nature. 1997;387:406–409. doi: 10.1038/387406a0. [DOI] [PubMed] [Google Scholar]