

Abstract
The isolation effect (or von Restorff effect) occurs when one item in a to-be-remembered list is distinctive from all remaining items, and memory for that item is enhanced. Four chimpanzees were presented with a serial list of four photographs. In the homogeneous condition, all list items were from the same semantic category (e.g., four fruits). In the isolate condition, three items were from the same category, but the fourth item (the isolate) was from a different category (e.g., three fruits and one toy). Then, two photographs were presented, and the chimpanzees had to select the one that was from the list. Two of four chimpanzees were significantly more likely to select a correct isolate item than an item from the same list position in the homogeneous condition for at least some list positions. This facilitation in performance was for isolate items only, as presenting an isolate item in a list did not facilitate greater recognition of other list items compared to the homogeneous condition. These results indicated that some chimpanzees perceived the semantic categories of the photographs, and categorization of photographs led to the isolation effect. Thus, chimpanzees may share with humans some aspects of memory organization that involve spontaneously categorizing visual stimuli and recognizing categorically unique stimuli.
Keywords: Chimpanzees, Pan troglodytes, von Restorff effect, Isolation effect, Memory
Introduction
The isolation effect, or von Restorff effect, occurs when one item in a list or set is distinctive from all remaining items, and memory for that item is enhanced (von Restorff 1933). Typically, the distinctiveness of an item entails it differing from the prevailing context of the list in general (i.e., it is perceptually or semantically unique; Hunt 1995; Schmidt 1991; Wallace 1965). Although there are numerous demonstrations of the isolation effect in human adults and children (e.g., Howe et al. 2000; Kishiyama and Yonelinas 2003; McLaughlin 1966), there is less evidence of this memory phenomenon in nonhuman animals (here-after, animals) in large part because of the difficulty of establishing categorically related stimulus sets into which distinctive items can be inserted. Often, tests with animals rely on perceptual rather than categorical or semantic distinctiveness for assessing isolation effects.
Reed et al. (1991) reported that rats showed increased recognition of goal boxes that were accompanied by a salient stimulus in a non-matching-to-sample paradigm. The rats were trained to select from two goal boxes the one that was not among the five previously visited goal boxes. When one of the previously visited goal boxes was made distinct by a change in lighting condition (dark goal boxes were illuminated and lighted goal boxes were darkened), the rats performed significantly better in avoiding that goal box during the non-matching-to-sample response. In a spatial memory test (Reed and Richards 1996), rats showed decreased numbers of errors in selecting already visited arms of a radial maze when those arms were made distinct by either having experimenters handle the rats immediately after entry or by having a tone follow immediately after entry into one of the arms as compared to trials without such distinction. Rats also showed decreasing numbers of errors in maze running when vivid stimuli were located within the maze (McLaughlin et al. 1968).
Parker et al. (1998) reported that monkeys and humans showed the isolation effect in a recognition paradigm after serial list presentation. After viewing a list of two-dimensional visual objects (either typographic stimuli or fractal stimuli), subjects were presented with a list stimulus and a novel (i.e., non-list) stimulus. Some lists contained isolate items that appeared in a different color from the other stimuli in the list. The monkeys and humans made fewer recognition errors for these isolate stimuli than for comparable position stimuli in the homogeneous (non-isolate) lists.
In these previous experiments, the isolation effect in nonhuman animals resulted from distinct perceptual experiences (in terms of lighting, color changes to stimuli, or visceral experiences). However, in humans, the isolation effect occurs with regard to semanticity and categorical similarity as when words or pictures that are categorically distinct within otherwise categorically homogeneous lists are recalled or recognized at higher levels than words without this semantic distinction (e.g., Fabiani and Donchin 1995). For example, the word hammer in a list of fruit names would be recalled at higher levels than another fruit name in the same list position. Whether nonhuman animals are susceptible to this type of isolation effect is unknown. Given that nonhuman animals are capable of some levels of categorization of two-dimensional stimuli (e.g., Bovet and Washburn 2003; Brown and Boysen 2000; Tanaka 2001; Wasserman et al. 2001), it is possible to present lists of items in which stimuli differ not in terms of physical and perceptual features but in terms of categorical semantic features.
In this study, four chimpanzees (Pan troglodytes) were presented with lists of four photographs of familiar foods, objects, people, and apes. The chimpanzees then were presented with a single item from the list and a foil stimulus from the same category, and the animals were rewarded for selecting the item that had been in the list. Two types of lists were presented. Homogeneous lists contained four items from the same semantic category followed by a probe item from that list and a foil item from the same category. Isolate lists contained three items from the same category, and a fourth categorically distinct item followed by the presentation of that distinct item and a foil item from the same category as that distinct item. Here, increased performance for the isolate item compared to performance for the same list position with homogeneous lists would indicate an isolation effect. Importantly, however, this increased performance should only be for isolate items. Performance should not improve for all items when an isolate is presented. Phase 2 of the experiment was designed to assess whether any facilitation in performance was for isolate items only.
Methods
Participants
Four chimpanzees participated in this study. At the time of testing, Lana was a 33-year-old female, Sherman was a 30-year-old male, Panzee was a 17-year-old female, and Mercury was a 16-year-old male. All four chimpanzees were housed together but were observed separately during test sessions. All chimpanzees worked for preferred food treats and were otherwise maintained on their normal diet of fruit, vegetable, and primate chow (i.e., no food or water deprivation was used). Three of the four animals (excluding Mercury) had learned to associate geometric forms called lexigrams that are used to represent items such as foods, locations, objects, and people (see Rumbaugh and Washburn 2003). These chimpanzees had experience in performing symbolic matching-to-sample tasks in which comparison stimuli differed from samples (including tasks such as selecting lexigrams to match spoken English words or photographs; Beran et al. 1998). All chimpanzees recognized the stimuli represented in the photographs that make up the presentation lists, and they could match photographs to real world objects (see Rumbaugh and Washburn 2003; Savage-Rumbaugh 1986). All chimpanzees previously were trained to perform a serial list recognition memory test like the one described below, but with stimuli that were not photographs of items like those used in this experiment.
Apparatus
Trials were presented on a Compaq DeskPro computer with an attached joystick. The program was written in Visual Basic for Windows. Joysticks were mounted on the chimpanzees' home cages so that they could manipulate the joysticks with their hands to control a cursor on the screen, and all chimpanzees were familiar with the use of this procedure. The stimuli presented during trials were colored photographic stimuli taken of real world items and individuals using a digital camera.
It is important to note that these photographs were sufficiently variable in their color and form that photos within a category were not perceptually distinct from all others from outside that category. The issue here is that perceptual distinctiveness had to be accounted for in order for an assessment of semantic isolation effects to occur. In other words, it was critical to know that an isolate item from one category would not always be perceptually distinctive from the other three items in the list, but would be semantically distinct.
Assessing the perceptual similarity between two or more photographs can be difficult to do in an objective manner. For the purposes of this study, a commercially available software package was used to assist in this assessment. The Visual Similarity Duplicate Image Finder (version 3.8.0.1) available at MindGems.com was used. This program allows the user to compare a digital image to all other digital images within a set, and it flags those groups of images that show perceptual similarity that exceeds the user's preset criterion. This allows one measure of how similar any two images are to each other on the basis of their perceptual features. This analysis showed that with one exception (the category of human faces), all images in any given category were not perceptually distinct from all images from another category (see ESM1 for more details on this analysis and the outcome rankings of photograph similarity). The main point here is that photographs of within category items were not always more physically similar to each other than they were to photographs from other categories. This does not mean that the categories used throughout this paper are accurate reflections of how chimpanzees might categorize the set of photos. The organization of these photos into the different categories is based entirely on human-based assumptions of what those categories are, and the chimpanzees' perspective on how to classify those photos may well differ from that of humans. Although it is possible to assess item similarity behaviorally (e.g., Astley and Wasserman 1992), this approach was not used in the present experiment because the training required to get the chimpanzees to indicate which photos were categorically related might have artificially inflated the likelihood of finding something that looked like an isolation effect, but that was reflective of training rather than a true isolation effect. Thus, the issue about whether chimpanzees really see these categories as clearly defined and as being inclusive of the photos that were objectively assigned to them cannot be confirmed. However, if chimpanzees do not see these categories the same way as they are defined, it should make it less likely that an isolation effect would be found rather than more likely.
Design and procedure
The task
The experimenter initiated the start of each trial by key press. A gray square appeared in the center of the screen, and a red cursor appeared at the bottom center of the screen. A chimpanzee moved the cursor into contact with the gray square to begin the list presentation. List items were presented in the center of the screen for 2 s each. These items were photographs that were 7.5 by 7.5 cm in size. Each photograph was presented immediately after the previous photograph was removed from the screen, and 1 s after the fourth item was presented, the comparison stimuli (5.5 cm by 5.5 cm) appeared on the screen. These comparison stimuli were located at the left center and right center of the screen, and the cursor was located at the bottom center of the screen. The chimpanzee moved the cursor into contact with one of the stimuli, and if it was the correct selection, a melodic tone sounded. If it was the incorrect selection, a buzz tone sounded. Correctly completed trials were rewarded with preferred food items distributed by the experimenter, who was unaware of the list items, the choice items, or the stimulus selected by the chimpanzees. Incorrectly completed trials were not rewarded. Two chimpanzees were tested within the same session (in separate cages and with separate trial lists). The experimenter moved back and forth throughout the session giving trials alternately to each chimpanzee. Thus, the inter-trial interval was not stable but typically ranged from 15 to 30 s.
The lists
The stimuli used in the experiment consisted of 100 photographs from 10 separate categories (see Table 1 for the names of the items in the photographs and ESM2 for the actual images). These categories were fruits, vegetables, liquids, treats, tools, food utensils, toys, husbandry items, human faces, and ape faces. All of the chimpanzees were familiar with all of the items in the photographs. In previous tests, Lana, Sherman, and Panzee were able to use photographs in ways that suggested those photos represented real items for the chimpanzees. For example, they matched real world items to those photographs or to their lexigram symbols, and they sorted photos into different categories such as food and tools (e.g., Beran et al. 1998; Rumbaugh and Washburn 2003; Savage-Rumbaugh et al. 1980). Mercury is not as sophisticated in these abilities, but he has been shown to match items to photographs, suggesting that he also might show an isolation effect in this test.
Table 1.
The photographic stimuli used in the lists
| Vegetables | Fluids | Fruits | Treats | Toys | Utensils | Husbandry | Tools | Apes | Humans |
|---|---|---|---|---|---|---|---|---|---|
| Tomato | Cola | Kiwi | Cereal | Ball | Bowl | Shot | Screw | Panzee | C1 |
| Carrot | Coffee | Strawberry | Cracker | Crayons | Knife | Oil | Dustpan | Austin | C2 |
| Onion | Water | Banana | Peanuts | Mask | Plate | Mirror | Nails | Lana | S |
| Celery | Koolaid® | Pineapple | m&m® | Paper | Pan | Toothbrush | Lever | Sherman | J |
| Lettuce | Lemonade | Apple | Pretzel | Paint | Cup | Thermometer | Hose | Panban | M1 |
| Broccoli | Juice | Pear | Jelly | Bear | Spoon | Stethoscope | Broom | Tamuli | D1 |
| Potato | Org. soda | Peach | Yogurt | Balloon | Fork | Toothpaste | Lighter | Kanzi | D2 |
| Sweet potato | Milk | Grapes | Raisins | Bubbles | Straw | Clippers | Wrench | Matata | M2 |
| Green beans | Org. juice | Orange | Gold fish | Clay | Can opener | Brush | Hammer | Mercury | S |
| Peas | Perrier® | Blackberry | Cheese | Chalk | Box | Medicine | Keys | Nyota | L |
The actual images are shown in the online supplementary materials (ESM2) that accompany this article
Phase 1
Examples of each of the list conditions are shown in Fig. 1. In the homogeneous list condition, four items were selected randomly from one of the 10 categories. The correct choice came from the list, and the foil (incorrect) item came from the same category but was not in the list. In the isolate list condition, three items were selected randomly from one of the 10 categories. The fourth item was selected randomly from one of the other categories with the stipulation that the isolate item could not come from any of the food lists (fruits, vegetables, drinks, treats) if the other three items in the list were from a food category, it could not come from any of the object lists (tools, food utensils, toys, and husbandry items) if the other three items were from an object list, and it could not come from the people or ape categories if the other three items came from either of those categories. This was done to insure the maximum level of categorical distinctiveness. In this condition, this isolate item was always the item that was tested. So, if an isolate was presented, it was the test item. Phase 2 included trials where this was not the case (see below).
Fig. 1.
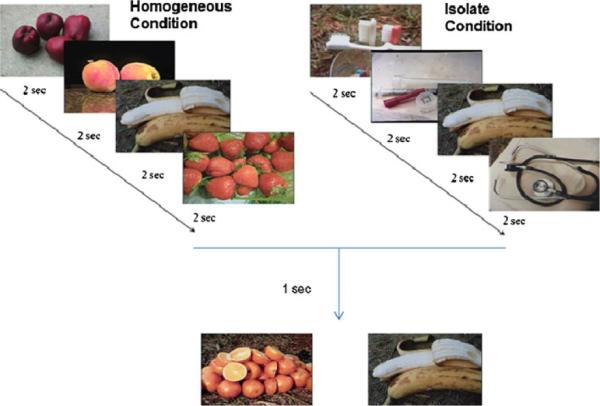
A schematic of trials in each of the two conditions. In the homogeneous list condition, each of the stimuli came from the same category (apple, peach, banana, strawberries). In the isolate list condition, one item differed categorically from the rest (toothbrush, syringe, banana, stethoscope). After each list was presented, two items were available as choice stimuli. Here, the presented items at the bottom were equally viable for each of the two conditions. In both cases, banana is the correct selection
Critically, for isolate lists, the foil stimulus came from the same category as the isolated item. For example, if a face were presented in a list of fruits, two faces (the one from the list and another face) would be presented as choice options. This prevented any possible facilitation in performance on isolate trials being solely the result of novelty of that isolate list item. In other words, on those trials, both choices were semantically and categorically distinct from the majority of the list items that had been presented, and so the chimpanzees could not succeed by simply choosing any categorically distinct item. Rather, they had to remember which specific distinct item was actually in the list.
In each session, 20 trials were completed—10 with homogeneous lists and 10 with isolate lists, presented in random order. For each condition, two trials each were presented in which the probe item came from list positions 1 and 4, and three trials each were presented in which the probe item came from list positions 2 and 3. Thus, when an isolated item was presented during a trial, it was presented in all four possible serial positions across trials. More trials were sampled from the middle list positions because it was assumed that performance might already be quite high in the first and fourth positions because of primacy and recency effects, and thus an effect of an isolate item would be difficult to demonstrate because of possible ceiling effects. Session duration was approximately 15 min. Each chimpanzee completed only one session per day (with an average of four sessions per week), and a total of 15 sessions (300 trials).
Phase 2
At the end of the first phase of the experiment, each chimpanzee completed an additional 200 trials (20 trials per day for 10 sessions). These trials were all identical in their presentation to those of the first phase. Half of the trials in each session were of homogeneous lists. For those trials, 20 each involved a test on items from list position 1 and list position 4; whereas 30 trials each involved a test on items from list position 2 and list position 3. The other 100 trials involved the presentation of an isolated item within an otherwise homogeneous list (as in the first phase). However, the test item was one of those homogeneous items compared to another item from the same category (but not from the list). For example, if the isolate item came from position 3 on a given trial, the chimpanzees were given a memory test for either the first or third item in the list. On those trials, 40 involved a test on items from list position 1, whereas 30 trials each involved a test on items from list position 2 and list position 3. List position 4 was not tested because of the ceiling effect seen in Phase 1.
Phase 2 acted as a control to determine whether the insertion of a distinctive item into an otherwise homogeneous list would facilitate recognition of only that item, or of any item on that list. The isolation effect (with higher performance for isolate lists compared to homogeneous lists) should only occur when chimpanzees are asked to recognize the single distinctive item in an isolate list, not any item from that list. Thus, if chimpanzees show higher performance in Phase 1 for isolate lists compared to homogeneous lists but that advantage disappears in Phase 2, this provides stronger evidence for the isolation effect in chimpanzee memory.
Results
The mean performances of the chimpanzees on homogeneous and isolate lists in Phase 1 at each list position are presented in Fig. 2. A within subjects ANOVA with list position (1, 2, 3, or 4) and list type (homogeneous and isolate) as independent variables indicated that there was a significant effect of list position, F (3, 9) = 12.40, P = 0.002, = 0.81, and the best fitting relation for that effect was linear. The effect of list type approached but did not exceed levels of statistical significance, F (1, 9) = 7.74, P = 0.069, = 0.72. The interaction between list position and list type was not significant, F (3, 9) = 1.87, P = 0.20, = 0.38. Power analyses indicated that although power was adequate for the list position variable (0.98), it was low (0.48) for list type. This was unavoidable because only four chimpanzees were available for the experiment. Given the near statistical significance level for list type, the lack of power for that analysis and the clear variability in performance across chimpanzees (as shown by the confidence bars in Fig. 2), performance was evaluated on an individual basis.
Fig. 2.
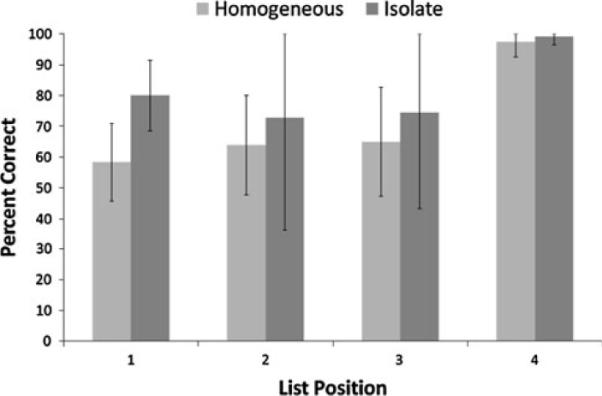
The mean performance of the chimpanzees in Phase 1 at each list position for each list type. Error bars show 95% confidence intervals
The performance of each chimpanzee in Phase 1 is shown in Fig. 3 and was assessed using a 4 × 2 × 2 log linear analysis with list position, list type, and outcome (correct versus incorrect). Because four such tests were conducted, the Bonferroni correction was applied, and the corrected alpha level was set at 0.0125. As indicated in the group analysis, all four chimpanzees showed a significant effect of list position on performance, all G2 (df = 3) > 15.20, P < 0.002. In addition, Lana and Panzee demonstrated the isolation effect. Lana performed significantly better in the isolate condition than in the homogeneous condition, G2 (df = 1) = 9.22, P = 0.002. Panzee also performed significantly better in the isolate condition than in the homogeneous condition, G2 (df = 1) = 27.26, P < 0.0001. Neither Mercury nor Sherman performed differently between the isolate list and the homogeneous list conditions overall, both G2 (df = 1) < 1.30, P > 0.25. However, a visual examination showed a clear facilitation in performance for the isolate list over the homogeneous list for position 1. A post hoc assessment of only this list position indicated the both chimpanzees were significantly better in the isolate condition than the homogeneous condition, Sherman X2 (1, N = 60) = 4.59, Mercury X2 (1, N = 60) = 4.16, both P < 0.05.
Fig. 3.

The chimpanzees' individual performances in Phase 1 for homogeneous and isolate lists at each list position
The mean performances of the chimpanzees on homogeneous and isolate lists in Phase 2 at list positions 1, 2, and 3 are presented in Fig. 4. A within subjects ANOVA with list position (1, 2, or 3) and list type (homogeneous and isolate) as independent variables indicated that there was a significant effect of list position, F (2, 6) = 6.02, P = 0.037, = 0.69. There was no effect of list type, F (1, 6) = 1.49, P = 0.31, = 0.33. The interaction between list position and list type was not significant, F (2, 6) = 4.42, P = 0.07, = 0.59. Given that the interaction approached statistical levels of significance, indicating that perhaps there was some difference between performance on each list type as a function of position, the performance of each chimpanzee was also assessed, post hoc, for list position 1 (this was the only possible candidate for a difference, as illustrated in Fig. 4). However, none of the chimpanzees showed a significant difference in performance at list position 1 between the isolate condition and the homogeneous condition, all P > 0.10.
Fig. 4.
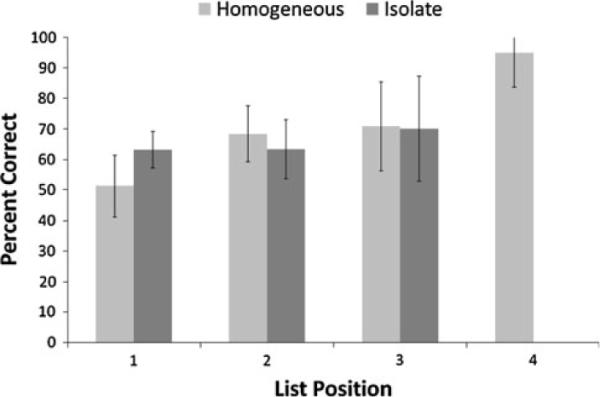
The mean performance of the chimpanzees in Phase 2 at each list position for each list type. Error bars show 95% confidence intervals
Discussion
There was evidence of an isolation effect in chimpanzee memory. Two of the four chimpanzees demonstrated facilitated performance for isolate list items compared to homogeneous list items. Panzee and Lana made fewer errors when the probe and foil stimuli were categorically distinct from the majority of the list items that had been presented. Sherman and Mercury showed this facilitation only for list position 1. None of the chimpanzees showed significantly improved performance for homogeneous lists over isolate lists at any list position, and the presentation of an isolate item did not simply improve performance for any item in the presented list but only operated to improve performance in remembering that isolate item (as shown by the results of Phase 2).
These results indicate that some chimpanzees were not responding solely on the basis of color and form information but also were using the categorical information that accrued during list presentation. Panzee's performance was particularly impressive as she maintained a very high level of responding across all list positions for the isolate items compared to the homogeneous condition. It is worth noting that she is generally regarded as the most sophisticated of these chimpanzees in terms of her semantic/symbolic competence using lexigrams and responding to human speech (e.g., Rumbaugh and Washburn 2003). However, Sherman and Lana also show differences in semantic sophistication that do not generally line up with performance on this task (Sherman being somewhat more sophisticated, but not more likely to show the isolation effect in the present experiment). Thus, individual differences in performance here might be related to rearing as it affected symbol competence and semantic sophistication, but the relation is not a strong one. It is likely not related to some inability to recognize the photographs represent real world items, given the past experiences of most of these chimpanzees, although this remains a possibility for Mercury given that he has not shown the same sophistication in using photographs flexibly compared to the other chimpanzees (e.g., Beran et al. 1999).
The fact that the effect of list position was best fit with a linear relation has implications for certain aspects of the chimpanzees' list memory. The chimpanzees showed high performance for the last list position (a recency effect). Although often reported in other studies with nonhuman animals (e.g., Reed et al. 1991; Reed and Richards 1996; Sands and Wright 1980; Wright et al. 1985), there was no evidence of increased performance at the early list position (a primacy effect). However, primacy effects are not always found in memory tests with nonhuman animals (e.g., MacPhail 1980; Thompson and Herman 1977). Wright et al. (1985) reported that primacy effects were dependent on the use of longer retention periods. With short retention periods, primacy effects were not found, and this is consistent with the data from the chimpanzees in the present experiment given the short retention interval that was used.
A number of processes have been proposed to account for the isolation effect in humans (for reviews, see Hunt 1995; Hunt and Lamb 2001; Waddill and McDaniel 1998). One account suggests that the isolate item is surprising, highly salient, and differentially attended to at presentation as a function of its surprise or salience (e.g., Green 1956; Jenkins and Postman 1948; Schmidt 1991). In this account, the individual vividness of the isolate or the surprise of its different properties relative to the other list items at presentation leads to differential attention, processing, and encoding. Another account suggests that the isolate item is remembered better because it is not subject to the same interference effects that occur for the other list items (Deutsch and Sternlicht 1967; Gibson 1940; McLaughlin 1968; McLaughlin et al. 1968; but also see McLaughlin 1966). Thus, it is not the perception of the isolate as unusual or salient that improves memory for that item but the fact that the isolate is not subject to the stimulus generalization that occurs for the other list items (Hunt 1995; von Restorff 1933; also see Hunt and McDaniel 1993).
More recently, the isolation effect has been proposed to occur because of the representational aspects of memory (Hunt 1995; Waddill and McDaniel 1998). Isolate items are categorized differently in relation to the remaining list items because of their perceptual, semantic, or categorical distinction. In this view, distinctiveness is not an independent variable of each item but rather occurs as an outcome of the processing of the entire list (Hunt 1995). Evidence in favor of this view comes from enhanced memory for isolate items presented even at the beginning of lists (von Restorff 1933). This early item distinctiveness with enhanced memory occurs before the establishment of context, and there cannot be surprise, enhanced salience, or differential attention and encoding as a result of that surprise (Hunt 1995; Kelley and Nairne 2001). Thus, with no established context salience, distinctiveness is not necessary for the isolation effect. As noted by Hunt and Lamb (2000), “Distinctiveness is a psychological resultant, not a property of actual objects and events…Differences are only distinct in the context of perceived similarity” (p. 1365).
An examination of Fig. 3 shows that three of four chimpanzees showed enhanced recognition of list position 1 stimuli in the isolate condition compared to the homogeneous condition. This suggests that perhaps the chimpanzees were not encoding the isolate stimuli any differently but were recognizing the categorical dissimilarity of those stimuli prior to selection. At present, there is no indication as to when this perception of categorical dissimilarity occurs. It may occur at some point during list presentation or may not occur until presentation of the probe and the foil. In any case, some chimpanzees were assessing the categorical information contained within the lists because they were able to recognize the isolate at higher levels than the same-position stimuli in the homogeneous lists, and this was done using other than the perceptual features of the stimuli (as shown in the supplementary materials).
It is important to note that although there was some evidence of the isolation effect in chimpanzees, it was not shown by all animals, and so replication will be important. Furthermore, as noted, some of these chimpanzees have very unique rearing histories that included emphasis on training semantic and symbolic competence. So, it will be important to test other chimpanzees without this background. It might also be productive to take advantage of stimuli that likely provide the clearest categorical distinctiveness for chimpanzees, such as testing food isolate versus food homogeneous and object isolate versus object homogeneous lists.
If these results can be replicated, future research using this methodology then could begin to distinguish between the different mechanisms that might underlie chimpanzee isolation effects. For example, a comparison of performance in a condition in which subjects are presented with a sequence of four photographs of objects from four different categories with that of the current homogeneous condition would provide some insight into the mechanism responsible for the isolation effect. This method also could be used to investigate whether chimpanzees might show the Deese–Roediger—McDermott memory illusion in which items related to those presented in a homogeneous list are erroneously reported as having been in the list (Roediger and McDermott 1995). The present demonstration of the isolation or von Restorff effect, along with demonstrations of very-long-term memory (Beran et al. 2000), recall memory (Menzel 1999), working memory span (Kawai and Matsuzawa 2000), and serial position effects (e.g., Buchanan et al. 1981) indicates another close similarity in memory phenomena between humans and at least some chimpanzees.
Supplementary Material
Acknowledgments
Michael J. Beran, Language Research Center, Georgia State University. This research was supported by Grant HD-38051 and Grant HD-060563 from the National Institute of Child Health and Human Development and by Grant BCS-0924811 from the National Science Foundation. The author thanks Ted Evans and David Washburn for their comments on an earlier version of this paper. All applicable institutional rules and regulations regarding animal care and use were followed in the care and testing of the chimpanzees, and the experiment complied with all laws of the United States of America.
Footnotes
Electronic supplementary material The online version of this article (doi:10.1007/s10071-011-0398-1) contains supplementary material, which is available to authorized users.
References
- Astley SL, Wasserman EA. Categorical discrimination and generalization in pigeons: all negative stimuli are not created equal. J Exp Psychol Anim Behav Proc. 1992;18:193–207. [Google Scholar]
- Beran MJ, Savage-Rumbaugh ES, Brakke KE, Kelley JW, Rumbaugh DM. Symbol comprehension and learning: a “vocabulary” test of three chimpanzees (Pan troglodytes) Evol Communication. 1998;2:171–188. [Google Scholar]
- Beran MJ, Savage-Rumbaugh ES, Pate JL, Rumbaugh DM. Delay of gratification in chimpanzees (Pan troglodytes) Dev Psychobiol. 1999;34:119–127. doi: 10.1002/(sici)1098-2302(199903)34:2<119::aid-dev5>3.0.co;2-p. [DOI] [PubMed] [Google Scholar]
- Beran MJ, Pate JL, Richardson WK, Rumbaugh DM. A chimpanzee's (Pan troglodytes) long-term retention of lexigrams. Anim Learn Behav. 2000;28:201–207. [Google Scholar]
- Bovet D, Washburn DA. Rhesus macaques (Macaca mulatta) categorize unknown conspecifics according to their dominance relations. J Comp Psychol. 2003;117:400–405. doi: 10.1037/0735-7036.117.4.400. [DOI] [PubMed] [Google Scholar]
- Brown DA, Boysen ST. Spontaneous discrimination of natural stimuli by chimpanzees (Pan troglodytes) J Comp Psychol. 2000;114:392–400. doi: 10.1037/0735-7036.114.4.392. [DOI] [PubMed] [Google Scholar]
- Buchanan JP, Gill TV, Braggio JT. Serial position and clustering effects in a chimpanzee's “free recall”. Memory Cogn. 1981;9:651–660. doi: 10.3758/bf03202360. [DOI] [PubMed] [Google Scholar]
- Deutsch MR, Sternlicht M. The role of “surprise” in the von Restorff effect. J Gen Psychol. 1967;76:151–159. doi: 10.1080/00221309.1967.9710384. [DOI] [PubMed] [Google Scholar]
- Fabiani M, Donchin E. Encoding processes and memory organization: a model of the von Restorff effect. J Exp Psychol Learn Mem Cogn. 1995;21:224–240. doi: 10.1037//0278-7393.21.1.224. [DOI] [PubMed] [Google Scholar]
- Gibson EJ. A systematic application of the concepts of generalization and differentiation to verbal learning. J Exp Psychol. 1940;66:111–119. [Google Scholar]
- Green RT. Surprise as a factor in the von Restorff effect. J Exp Psychol. 1956;52:340–344. doi: 10.1037/h0047496. [DOI] [PubMed] [Google Scholar]
- Howe ML, Courage ML, Vernescu R, Hunt M. Distinctiveness effects in children's long-term retention. Dev Psychol. 2000;36:778–792. doi: 10.1037//0012-1649.36.6.778. [DOI] [PubMed] [Google Scholar]
- Hunt RR. The subtlety of distinctiveness: what von Restorff really did. Psychon Bull Rev. 1995;2:105–112. doi: 10.3758/BF03214414. [DOI] [PubMed] [Google Scholar]
- Hunt RR, Lamb CA. What causes the isolation effect? J Exp Psychol Learn Mem Cogn. 2001;27:1359–1366. [PubMed] [Google Scholar]
- Hunt RR, McDaniel MA. The enigma of organization and distinctiveness. J Memory and Lang. 1993;32:421–445. [Google Scholar]
- Jenkins WO, Postman L. Isolation and the spread of effect in serial learning. Am J Psychol. 1948;61:214–221. [PubMed] [Google Scholar]
- Kawai N, Matsuzawa T. Numerical memory span in a chimpanzee. Nature. 2000;403:39–40. doi: 10.1038/47405. [DOI] [PubMed] [Google Scholar]
- Kelley MR, Nairne JS. Von Restorff revisited: Isolation, generation, and memory for order. J Exp Psychol Learn Mem Cogn. 2001;27:54–66. [PubMed] [Google Scholar]
- Kishiyama MM, Yonelinas AP. Novelty effect on recollection and familiarity in recognition memory. Mem Cogn. 2003;31:1045–1051. doi: 10.3758/bf03196125. [DOI] [PubMed] [Google Scholar]
- MacPhail EM. Short-term visual recognition memory in pigeons. Q J Exp Psychol. 1980;32:173–175. doi: 10.1080/14640748008401843. [DOI] [PubMed] [Google Scholar]
- McLaughlin JP. The von Restorff effect in serial learning: serial position of the isolate and length of list. J Exp Psychol. 1966;72:603–609. doi: 10.1037/h0023755. [DOI] [PubMed] [Google Scholar]
- McLaughlin JP. Recall and recognition measures of the von Restorff effect in serial learning. J Exp Psychol. 1968;78:99–102. doi: 10.1037/h0026156. [DOI] [PubMed] [Google Scholar]
- McLaughlin JP, Cicala GA, Pierson DK. Von Restorff effect in rat maze learning. J Comp Physiol Psychol. 1968;56:427–431. doi: 10.1037/h0026358. [DOI] [PubMed] [Google Scholar]
- Menzel CR. Unprompted recall and reporting of hidden objects by a chimpanzee (Pan troglodytes) after extended delays. J Comp Psychol. 1999;113:426–434. doi: 10.1037/0735-7036.113.4.426. [DOI] [PubMed] [Google Scholar]
- Parker A, Wilding E, Akerman C. The von Restorff effect in visual object recognition memory in humans and monkeys: the role of frontal/perirhinal interaction. J Cog Neuro. 1998;10:691–703. doi: 10.1162/089892998563103. [DOI] [PubMed] [Google Scholar]
- Reed P, Richards A. The von Restorff effect in rats (Rattus norvegicus) J Comp Psychol. 1996;110:193–198. doi: 10.1037/0735-7036.110.2.193. [DOI] [PubMed] [Google Scholar]
- Reed P, Chih-Ta T, Aggleton JP, Rawlins JNP. Primacy, recency, and the von Restorff effect in rats' nonspatial recognition memory. J Exp Psychol Anim Behav Proc. 1991;17:36–44. [Google Scholar]
- Roediger HL, III, McDermott KB. Creating false memories: remembering words not presented in lists. J Exp Psychol Learn Mem Cogn. 1995;21:803–814. [Google Scholar]
- Rumbaugh DM, Washburn DA. Intelligence of apes and other rational beings. Yale University Press; New Haven: 2003. [Google Scholar]
- Sands SF, Wright AA. Primate recognition: recognition of serial list items by a rhesus monkey. Science. 1980;209:938–939. doi: 10.1126/science.6773143. [DOI] [PubMed] [Google Scholar]
- Savage-Rumbaugh ES. Ape language: from conditioned response to symbol. Columbia University Press; New York: 1986. [Google Scholar]
- Savage-Rumbaugh ES, Rumbaugh DM, Smith ST, Lawson J. Reference: the linguistic essential. Science. 1980;210:922–925. doi: 10.1126/science.7434008. [DOI] [PubMed] [Google Scholar]
- Schmidt SR. Can we have a distinctive theory of memory? Mem Cogn. 1991;19:523–542. doi: 10.3758/bf03197149. [DOI] [PubMed] [Google Scholar]
- Tanaka M. Discrimination and categorization of photographs of natural objects by a chimpanzee (Pan troglodytes) Anim Cogn. 2001;4:201–211. doi: 10.1007/s100710100106. [DOI] [PubMed] [Google Scholar]
- Thompson RKR, Herman LM. Memory for lists of sounds by the bottlenosed dolphin: convergence of memory processes with humans? Science. 1977;191:501–503. doi: 10.1126/science.835012. [DOI] [PubMed] [Google Scholar]
- von Restorff H. Uber die Wirkung von Bereichsbildungen im Spurenfeld. Psychologische Forschung. 1933;18:299–342. [Google Scholar]
- Waddill PJ, McDaniel MA. Distinctiveness effects in recall: differential processing or privileged retrieval? Mem Cogn. 1998;26:108–120. doi: 10.3758/bf03211374. [DOI] [PubMed] [Google Scholar]
- Wallace WP. Review of the historical, empirical, and theoretical status of the von Restorff phenomenon. Psychol Bull. 1965;63:410–424. doi: 10.1037/h0022001. [DOI] [PubMed] [Google Scholar]
- Wasserman EA, Fagot J, Young ME. Same-different conceptualization by baboons (Papio anubis): the role of entropy. J Comp Psychol. 2001;115:42–52. doi: 10.1037/0735-7036.115.1.42. [DOI] [PubMed] [Google Scholar]
- Wright AA, Santiago HC, Sands SF, Kendrick FF, Cook RG. Memory processing of serial lists by pigeons, monkeys, and people. Science. 1985;229:287–289. doi: 10.1126/science.9304205. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.