

Abstract
Naïve T cells continuously traffic to secondary lymphoid organs, including peripheral lymph nodes, to detect rare expressed antigens. The migration of T cells into lymph nodes is a complex process which involves both cellular and chemical factors including chemokines. Recently, the use of two-photon microscopy has permitted to track T cells in intact lymph nodes and to derive some quantitative information on their behavior and their interactions with other cells. While there are obvious advantages to an in vivo system, this approach requires a complex and expensive instrumentation and provides limited access to the tissue. To analyze the behavior of T cells within murine lymph nodes, we have developed a slice assay 1, originally set up by neurobiologists and transposed recently to murine thymus 2. In this technique, fluorescently labeled T cells are plated on top of an acutely prepared lymph node slice. In this video-article, the localization and migration of T cells into the tissue are analyzed in real-time with a widefield and a confocal microscope. The technique which complements in vivo two-photon microscopy offers an effective approach to image T cells in their natural environment and to elucidate mechanisms underlying T cell migration.
Keywords: Immunology, Issue 53, mouse, lymph node, organotypic slices, T cell, migration, fluorescence, microscopy, confocal
Protocol
1. Preparing slices from lymph nodes
Prepare a 4% low-melting point agarose solution for embedding. Dilute agarose in PBS and microwave this solution. When the agarose is completely dissolved, maintain the solution at 37°C until ready for use.
Prepare a 6-well tissue culture plate. Add 1.1 ml RPMI complete culture medium per well, and insert an organotypic filter in each well. Place the 6-well plate at 4°C until ready to transfer slices.
Place stainless steel washers, with inner diameters of 4 mm, into a plastic dish filled with RPMI complete medium. Washers will be further used to concentrate the cells on the vibratome cut slice.
Sacrifice the mouse in accordance with local animal welfare regulations. Carefully remove the peripheral lymph nodes from the surrounding tissue and place them in a plastic dish containing ice-cold PBS. Care must be taken not to damage the lymph node structure, as these organs are very soft.
Pour the 4%-agarose gel into a 35-mm plastic dish and delicately transfer the nodes into the gel. Leave the agarose gel on ice for 5 min to harden.
Remove the block from the dish and trim the agar in order to leave 3-5 mm of gel around each lymph node.
Attach the embedded nodes on the specimen disk of the vibratome with non-toxic tissue glue. Install the specimen disk in the tray filled with ice-cold PBS.
Section the agar-embedded tissue at 320 μm thickness with the vibratome speed set on a slow range (0.3 mm/s) and the vibration frequency set on a medium range (1.5 mm).
Carefully transfer lymph node slices as they are being cut onto the organotypic culture inserts using fine forceps. Place 3 slices on each insert. Use great care as slices can be easily damaged. A typical peripheral lymph node will yield 5 sections when cut at 320 μm. Discard the first and last slices, as they only contain superficial tissue.
Place stainless steel washers on each individual slice. Make sure washers are well positioned on the agarose surrounding the tissue.
Incubate the culture plate at 37°C in a 5% CO2 humidified incubator until ready to plate T cells.
2. Isolating T cells from mouse lymph nodes
Isolate T cells according to the previously published JoVE article 3.
Use T cells immediately and proceed to the next section. Otherwise, lymphocytes might be cultured overnight in complete RPMI-medium supplemented with 10 ng/ml IL-7.
3. Labeling of isolated T cells
Wash cells in HBSS. Resuspend them in the same buffer at 10 x 106 per ml.
Add the same volume of HBSS containing 1 μM Cell Tracker green CMFDA giving a final concentration of 0.5 μM.
Incubate the cell suspension at 37°C during 5 min. Wash cells twice with complete RPMI-medium.
Resuspend cells at a final concentration of 10 x 106 per ml in complete RPMI-medium. These labeled cells will be plated onto the slices.
Other fluorescent dyes than CMFDA might be used too, but keep in mind that these molecules are potentially toxic above certain concentrations.
4. Plating labeled T cells onto lymph node slices
Remove excess medium contained in the inner part of the washer with a pipette. Do not allow the tissue to become dry; work quickly during this step.
Gently dispense 10 to 20 μl labeled T cells that corresponds to 1 to 2 x 105 cells in the inner part of the washer. Care must be taken not to touch the tissue with the tip of the pipette. Be sure that the drop of cell suspension stays in place.
Place the slices with cells in incubator at 37°C and 5% CO2 for at least 30 min to allow lymphocytes to migrate into the tissue.
5. Imaging T cells within lymph node slices
In this video-article, we image fluorescent T cells in lymph node slices with a widefield inverted microscope and a confocal upright microscope.
Set the temperature at 37°C. Our microscopes are equipped with temperature-controlled chambers.
Install a perfusion system enabling continuous perfusion of the lymph node slice with oxygenated (5% CO2, 95 % O2) phenol red-free RPMI medium. In our system, perfused media enters into the imaging chamber from one side by gravity. The medium is aspirated with a pump connected to a waste collection flask. Set the media flow rate to 1 ml/min.
With fine forceps, delicately take out the slice from the 6-well plate and dip the preparation in warmed oxygenated RPMI medium for a few seconds to get rid of fluorescent cells not infiltrated into the tissue.
Imaging T cells with a wide-field inverted microscope
Place the slice upside down on a custom-made imaging chamber which consists of nylon threads glued onto the inner part of a glass bottom dish. In this chamber, the nylon threads elevate the slice from the glass and enable the oxygenated solution to diffuse into the tissue. This is required as T cell migration in lymph nodes is strongly dependent on oxygen. Secure the slice by adding onto it a stainless steel washer. In these conditions, the slice lies a few microns above the bottom of the dish, which facilitates renewal of the perfusion solution.
To capture a large field of view (800 x 800 μm, approximately one third of the node's slice surface), collect images using a 10x dry objective lens. We found that the 10x Nikon S fluor 0.5 NA was most suitable for our analyses.
A typical time-lapse imaging experiment captures 5 optical planes spanning a total depth of 50 μm in the axial (z) dimension. Fluorescent T cells are usually imaged at intervals ranging 10 to 30 seconds during 10 to 20 min.
Imaging T cells with a confocal upright microscope
The confocal microscope used in this protocol is a Leica SP5 equipped with a 20x water immersion objective (Olympus, 20x/0.95 NA).
Secure the slice to the imaging stage of the microscope. Note: We usually place a washer on the preparation to immobilize it.
Set an imaging session by collecting a series of images along the z axis. To image T cells in an intact tissue structure, the start position is usually set at 5 to 10 μm below the first labeled T cell.
6. Representative results
By following this video-protocol you should expect to visualize a large number of fluorescent T cells accumulated in the T zone of the node, a phenomenon that normally occurs in this particular environment in vivo (Figure 1). In particular, T cells should be excluded from B cell zones, which are usually just beneath the capsule. A successful experiment will also result in T cells recruited into the tissue (Figure 2), displaying a highly motile behavior (Figure 3) consistent with published results obtained in intact lymph nodes 4. On average, the mean velocity of individual T cells within slices should be close to 10 μm/min (Figure 4).
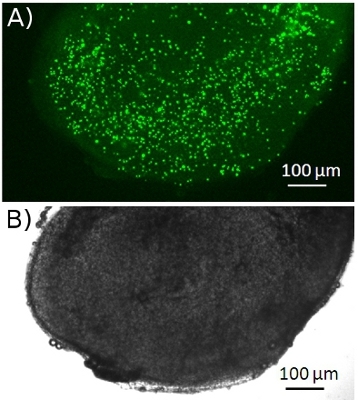 Figure 1. T cells accumulate in a lymph node slice. Fluorescently-labeled T cells (CMFDA, green) were added to a lymph node slice 30 min before image acquisition (top picture). The image is the maximum projection of 5 images spanning 50 μm in the z direction. The bright field image is shown in bottom. Images were captured with a widefield microscope.
Figure 1. T cells accumulate in a lymph node slice. Fluorescently-labeled T cells (CMFDA, green) were added to a lymph node slice 30 min before image acquisition (top picture). The image is the maximum projection of 5 images spanning 50 μm in the z direction. The bright field image is shown in bottom. Images were captured with a widefield microscope.
 Figure 2. T cells are recruited into a lymph node slice. Fluorescently-labeled T cells (CMFDA, green) were added to a lymph node slice 30 min before image acquisition using a confocal microscope. Images were captured at the cut surface (top picture) and 40 μm below (bottom picture).
Figure 2. T cells are recruited into a lymph node slice. Fluorescently-labeled T cells (CMFDA, green) were added to a lymph node slice 30 min before image acquisition using a confocal microscope. Images were captured at the cut surface (top picture) and 40 μm below (bottom picture).
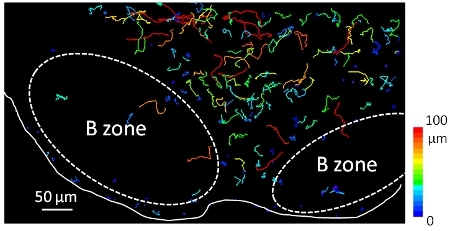 Figure 3. T cells are highly motile within a lymph node slice. Fluorescently-labeled T cells were imaged during 12 min using a widefield microscope. Trajectories of individual T cells are displayed as color-coded tracks to represent increasing displacements from blue (low motile cells) to red (high motile cells). Tracks were calculated using Imaris software. The white line follows the edge of the node, whereas the dashed oval delimits the putative B cell zone.
Figure 3. T cells are highly motile within a lymph node slice. Fluorescently-labeled T cells were imaged during 12 min using a widefield microscope. Trajectories of individual T cells are displayed as color-coded tracks to represent increasing displacements from blue (low motile cells) to red (high motile cells). Tracks were calculated using Imaris software. The white line follows the edge of the node, whereas the dashed oval delimits the putative B cell zone.
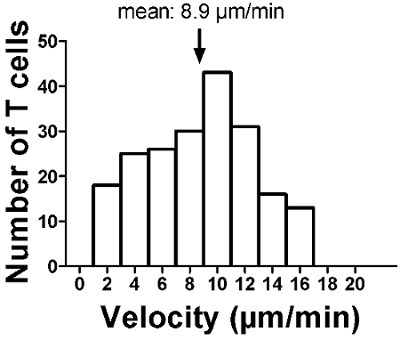 Figure 4. Within a lymph node slice, the T cell velocity is close to 10 μm/min. The speeds were calculated using Imaris software from tracks represented in Figure 3.
Figure 4. Within a lymph node slice, the T cell velocity is close to 10 μm/min. The speeds were calculated using Imaris software from tracks represented in Figure 3.
Discussion
We have described a straightforward, quick and robust technique for generating lymph node slices, which are used to investigate the behavior of introduced T cells. In recent years, this method has been applied successfully using thymic and lymph node slices to identify the extracellular factors controlling T cell positioning and motility 1,5. It has also been used to measure Ca2+ responses in thymocytes during positive selection and in T cells upon antigen recognition 2,6. The overlay slice assay presents advantages and limitations that deserve to be discussed. Of note, this system permits access to the tissue, useful if one needs to manipulate slices pharmacologically in order to interfere with the molecular control of cell migration. Subsequent to imaging, the slices can be processed for immunohistochemistry to collect further information about the structures that have been imaged. Moreover, observation can be made with a traditional widefield fluorescence microscope. Although the resolution is not as good as with a confocal or a two-photon microscope and that phototoxicity is in principle more severe with one-photon than with two-photon microscopy, it has the advantages of simplicity, a lower cost, and larger choice of excitation wavelengths.
The imaging of T cells in an intact lymph node requires intravenous injection of labeled lymphocytes that subsequently home to lymphoid organs. As we have shown previously 1, the slice assay is perfectly compatible with adoptive transfer experiment. However, the length of time needed for T cells to home into lymph nodes can complicate the use of fluorescent dyes that have a tendency to leak out of the cells over time. This is the case of the Ca2+ dye fura-2. With the fast recruitment (< 30 min) of T cells into the tissue, the slice assay provides an opportunity to use such dyes. Finally, this method has the great advantage to analyze T cell functions in several human tissues kept alive.
This experimental system presents also limitations that need to be kept in mind. Damage associated with the slicing may affect T cell functioning, especially in the superficial region of the tissue near the cut surface. In order to assess cell death within the slice, we have used the fluorescent dye SYTOX green that penetrates cells with compromised plasma membrane. Our experiments reveal that about 20% of total nodal cells, mostly localized in the superficial region of the tissue, were fluorescently labeled with this nuclear dye. Another potential concern with the assay is the ability of the slices to retain important soluble factors including chemokines. Although, we have no indication that our data were affected by such problems since T cells display a good motility within the slice, we would like to stress the importance of imaging T cells in healthy regions located at several tens of microns from the cut surface. Whereas, this could be done with traditional microscopes (widefield or confocal) like in this protocol, it is likely that the lymph node slice preparation combined with two-photon imaging will increase the spatial resolution in depth and reduce the phototoxicity.
Disclosures
No conflicts of interest declared.
Acknowledgments
The authors would like to thank Dr. Alain Trautmann who encouraged us to perform lymph node slices. This work was supported in part by grants from the Ligue Nationale Contre le Cancer, the Fondation pour la Recherche Médicale en France and the Association pour la recherche sur le Cancer.
References
- Asperti-Boursin F. CCR7 ligands control basal T cell motility within lymph node slices in a phosphoinositide 3-kinase-independent manner. J Exp Med. 2007;204:1167–1167. doi: 10.1084/jem.20062079. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bhakta NR, Oh DY, Lewis RS. Calcium oscillations regulate thymocyte motility during positive selection in the three-dimensional thymic environment. Nat Immunol. 2005;6:143–143. doi: 10.1038/ni1161. [DOI] [PubMed] [Google Scholar]
- Matheu MP, Cahalan MD. Isolation of CD4+ T cells from mouse lymph nodes using Miltenyi MACS purification. J Vis Exp. 2007. [DOI] [PMC free article] [PubMed]
- Miller MJ, Wei SH, Parker I, Cahalan MD. Two-photon imaging of lymphocyte motility and antigen response in intact lymph node. Science. 2002;296:1869–1869. doi: 10.1126/science.1070051. [DOI] [PubMed] [Google Scholar]
- Ehrlich LI, Oh DY, Weissman IL, Lewis RS. Differential contribution of chemotaxis and substrate restriction to segregation of immature and mature thymocytes. Immunity. 2009;31:986–986. doi: 10.1016/j.immuni.2009.09.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gollmer K. CCL21 mediates CD4+ T-cell costimulation via a DOCK2/Rac-dependent pathway. Blood. 2009;114:580–580. doi: 10.1182/blood-2009-01-200923. [DOI] [PMC free article] [PubMed] [Google Scholar]