

Abstract
Hematopoietic stem cells (HSCs) and mesenchymal stem cells (MSCs) are both adult stem cells residing in the bone marrow. MSCs interact with HSCs, they stimulate and enhance the proliferation of HSCs by secreting regulatory molecules and cytokines, providing a specialized microenvironment for controlling the process of hematopoiesis. In this paper we discuss how MSCs contribute to HSC niche, maintain the stemness and proliferation of HSCs, and support HSC transplantation.
1. Introduction
Hematopoietic stem cells (HSCs) are rare cells residing in the bone marrow (BM; 1 in 104 to 1 in 108 of BM nucleated cells), and they are progenitors that become progressively restricted to several or single lineages. These progenitors yield blood precursors devoted to unilineage differentiation and the production of mature blood cells, including red blood cells, megakaryocytes, myeloid cells (monocyte/macrophage and neutrophil), and lymphocytes [1, 2]. CD34 surface antigen (CD34+) is commonly used as a marker to identify and quantify the population of progenitor cells [3], according to which, sorting HSCs from BM, peripheral blood (PB), and umbilical cord (UC)/placenta blood is relatively simple and practical [2, 4–6]. Human HSCs are known to exhibit CD34+, Thy1+, CD38lo/−, Ckit−/lo, CD105+, and Lin− phenotype. However, there is no general agreement on the association between any combination of these antigenic properties and function of stem cells [3, 6]. HSCs depend on their microenvironment, the niche, for regulating self-renewal and differentiation [7]. For instance, the disruption of BMP pathway can increase the numbers of osteoblasts and HSCs [8, 9], and the chemokine CXCL12 regulates the cyclical release and the migration of HSCs [10, 11]. Activation of β-catenin enforces HSCs enter cell cycle, thus leading to exhaustion of the long-term stem cell pool [12–14]. These findings suggest that signaling pathways and cellular interactions regulate the BM niche for HSCs. Besides, hypoxia regulate hematopoiesis in BM by maintaining important HSC functions and the interplay between HSCs and neighboring cells [15, 16].
Plating studies indicate that mesenchymal stem cells (MSCs) are present as a rare population of cells in the BM. They represent approximately 0.001% to 0.01% of the nucleated cells, about 10-fold less abundant than HSCs, but MSCs can be readily grown in culture [17]. Though predominantly residing in the BM, MSCs also present similar but not identical features in many other tissues such as blood, placenta, dental pulp, and adipose tissue. MSCs have the potential to differentiate into multiple phenotypes such as osteoblasts, chondrocytes, adipocytes, neural cells, and probably other cell lineages [18–21]. International Society for Cellular Therapy (ISCT) has provided the following minimum criteria for defining multipotent mesenchymal stromal cells as follows: plastic-adherent under standard culture conditions; express CD105, CD73, and CD90 and lack expression of CD45, CD34, CD14, or CD11b, CD79 or CD19 and HLA-DR, and must differentiate into osteoblasts, adipocytes, and chondroblasts in vitro [22].
BM has received the most attention because it carries MSCs as well as HSCs. Evidence indicates that MSCs are key component of the HSC niche in the BM where these two distinct stem cell populations arrange closely, ensuring hematopoietic and skeletal homeostasis [18]. MSCs interact with HSCs, secreting chemokines that contribute to HSC niche and support long-term growth of HSCs [23, 24]. MSCs can be cotransplanted with HSCs to improve their engraftment [25–27] (Table 1).
Table 1.
The cytokines secreted by MSCs that regulate HSCs.
| Cytokines | Function | References |
|---|---|---|
| CXCL12 | Regulate the adhesion, expansion, migration, and homing of HSCs | [10, 11, 28–32] |
| (SDF-1) | Reduce the production of inflammatory cytokines and chemokines | [33] |
|
| ||
| FL | Maintain HSC proliferation and self-renewal, regulate hematopoietic growth | [28] |
|
| ||
| IL-6, TPO | Influence HSC proliferation and differentiation | [29, 34] |
|
| ||
| GM-CSF | Regulate HSC engraftment | [35] |
|
| ||
| SCF | Maintain HSC proliferation and self-renewal | |
| Regulate hematopoietic growth | [28, 36] | |
| Regulate HSC engraftment | [35] | |
|
| ||
| VCAM1, E-selectin, collagen I, fibronectin | Regulate HSC homing and adhesion | [35, 37] |
2. Mesenchymal Stem Cells Contribute to Hematopoietic Stem Cell Niche
The term “niche” for the specific HSC BM microenvironment was first coined in 1978, proposing that HSCs are in intimate contact with the bone, which was responsible for the apparently unlimited capacity of HSCs' proliferation and the inhibition of HSCs' maturation [38]. Niches exist within the BM which preserve specific aspects of hematopoiesis, such as HSC survival, self-renewal, and differentiation, supporting the maintenance of the blood system under normal and stressed conditions [39]. Research has made it increasingly clear that the stem cell niches provide a microenvironment which is important in protecting the self-renewing, undifferentiated state of their residents [40]. Three types of HSC niches have been hypothesized, defined according to the HSC uniformity [18, 41]. Two of these proposed niches are provided by cells directly descending from MSCs: the osteoblastic niche, where HSCs reside in close contact with endosteal cells [8], and the reticular stromal niche, where HSCs reside in close contact with stromal cells which are also known as mural cells or pericytes, the smooth muscle cells lining arteriolar side of the sinusoids [42]. The third proposed niche is the vascular/sinusoidal niche, where HSCs reside in direct contact with endothelial cells in the venous side of the sinusoids [43]. It is well known that HSC circulation involves HSCs leaving the BM, entering the vascular system (mobilization), and returning to the BM (homing) [44, 45]. The BM vascular structure provides a barrier between the hematopoietic compartment and the peripheral circulation. Most primitive HSCs remain physiologically quiescent within the BM niche; however, a portion of HSCs leave this resting pool and start the process of mobilization [39, 46–48].
Studies showed that both mouse and human osteoblast cell lines secreted a large number of cytokines that promote the proliferation of haematopoietic cells in culture, proving that cells involved in bone formation have stem-cell-supporting activity [49, 50]. MSCs reside in the bone cavity and are proposed to give rise to the majority of marrow stromal cell lineages, including chondrocytes, osteoblasts, and adipocytes, as suggested in numerous studies [48–50]. MSCs and HSCs form a structurally unique niche in the BM, which is regulated by local input from the surrounding microenvironment, and long-distance cues from hormones and the autonomic nervous system [51]. MSCs isolated from BM produce several growth factors and chemokines, such as CXCL12 (SDF-1), stem cell factor (SCF), Flt-3 ligand (FL), thrombopoietin (TPO), interleukin (IL)-6, IL-11, leukemia inhibitory factor (LIF), macrophage colony-stimulating factor (M-CSF), tumor necrosis factor- (TNF-) α, and transforming growth factor- (TGF-) β1 [28, 52–54]. HSCs are reduced in the BM after the depletion of MSCs, owing at least in part to mobilization towards extramedullary sites [51]. Loss of SCF from supporting cells or the receptor in HSCs leads to hematopoietic failure, indicating MSCs play an essential role in HSC niche function [36]. SCF and FL are implicated in maintaining HSC proliferation and self-renewal, regulating hematopoietic growth [28]. IL-3 or IL-6 combined with TPO signaling can influence HSC proliferation and differentiation [29, 34]. Besides, as mentioned previously, the chemokine CXCL12 interacts with its receptor CXCR4, regulates the cyclical release of HSCs, the migration of HSCs to the vascular niche from BM, and the homing of HSCs to the BM [10, 11, 29–32], and promotes adhesive interactions between HSCs and stromal cells [55]. In addition, CXCL12 chemokine signaling pathway contribute to the ex vivo expansion of HSCs [28]. Moreover, CXCL12 mediates angiogenic responses, promotes differentiation of CD34+ cells to endothelial progenitor cells, and appears to affect many other factors, including G-CSF, VEGF, and CXCL16 that relate to HSC mobilization and homing [33]. However, only β-catenin-activated MSCs but not naïve MSCs have stimulatory effect on HSC self-renewal in vivo [56].
3. The Effect of Mesenchymal Stem Cell on the Maintenance of Hematopoietic Stem Cells
Coculture of HSCs with MSCs might be an ideal method for maintaining the HSC pluripotency, because the growth or survival signals might be transferred to the HSC via adhesive molecules by modulating the cytokines and growth factor-dependent signals [57]. 5-aza-deoxycytidine (aza-D) and trichostatin A (TSA) have potent activity to maintain the stemness of HSCs, being candidate additives for HSCs ex vivo expansion, but they can also cause serious cell death [58, 59]. Koh et al. examined the effects of MSCs on the maintenance of CD34+ cells driven by aza-D and TSA in culture with the combined cytokines, and found that the total cell number of HSCs cultured with MSCs was higher in aza-D or TSA than in any culture conditions without MSCs, while most of HSCs cultured with cytokine treatment but without MSCs would lose their pluripotency and then differentiate, though they were induced to proliferate effectively [60]. It suggested that the co-culture of CD34+ cells with MSCs might not simply deliver the proliferation signals but also stemness and survival signals, and overlap the action of epigenetic regulators [57, 60].
4. Application of Bone Marrow Mesenchymal Stem Cells in Hematopoietic Stem Cell Transplantation
HSCs were primarily used in the treatment of patients with hematological malignancies. During the course of treatment, patients' cancerous cells are first destroyed by chemo/radiotherapy and then replaced with BM or PB/G-CSF transplant from a human leukocyte antigen- (HLA-) matched donor [61, 62]. In most cases, autologous HSCs are collected prior to the treatment and reinfused into the patients, but the patient's cancerous cells may be inadvertently collected and reinfused back into the patients along with HSCs [63]. Allogeneic marrow transplants have also been used in the treatment of hereditary blood disorders including aplastic anemia, β-thalassemia, Wiskott-Aldrich syndrome, and SCID, as well as inborn errors of metabolism disorders such as Hunter's syndrome and Hurler's syndrome [64–68]. One of the major challenges with HSC transplants is failure to engraft, which is mediated by donor T cells as a result of graft-versus-host disease (GVHD). Graft-versus-tumor effect of allogeneic HSC transplants may be a result of an immune reaction between donor cytotoxic T cells and patient's malignant cells [69]. MSCs are known to interact with HSCs and immune cells, and represent potential cellular therapy to enhance allogeneic hematopoietic engraftment and prevent GVHD [70–72]. Coculture of MSCs and HSCs could cause a significantly increase in CD34+ cells [73]. Aside from BM-derived MSCs, MSCs from adipose tissue can also be applied in hematopoietic engraftment, which would be an innovative supplement for cellular therapies [74, 75].
Cotransplantation studies in animal models as well as in humans showed that primary or culture-expanded MSCs promote the engraftment of HSCs. Cotransplantation of MSCs and cord blood or mobilized peripheral blood CD34+ cells resulted in a significantly higher level of engraftment than transplantation of CD34+ cells only [35, 37, 76–81]. This enhancement was greater after cotransplantation of GM-CSF and SCF-transfected MSCs, indicating that these growth factors relate to engraftment, though the mechanism of the enhancing effect is still unknown [35]. It is likely that the ability to promote engraftment is maintained along lineage differentiation [76]. Several lines of evidence suggest that MSCs produce several essential hematopoietic growth factors, adhesion molecules [28, 52–54], and extracellular matrix (ECM) proteins (such as VCAM1, E-selectin, collagen I, and fibronectin) that are known to play an important role in HSC homing [35, 37]. Selective adhesion of progenitors and cytokines to ECM components or stromal cells then result in the colocalization of progenitors at a specific stage of differentiation with a specific array of cytokines in so-termed niches [77]. This provides a level of growth and differentiation regulation [37]. Although it would mean exposure to allogeneic donor antigens, allogeneic MSCs can provide equal enhancement of engraftment as autologous cells. Cotransplanted MSCs shift the differentiation pattern from a lymphoid to a myeloid predominance and enhance megakaryocytic engraftment [78]. The cotransplantation of HSCs and MSCs enhanced engraftment as the dose of MSCs increased whereas an excessive dose of MSCs might decrease engraftment efficiency [79]. Besides, human allogeneic MSC layers in a serum-free culture system enabled the ex vivo expansion/maintenance of human HSCs [80], which indicates that MSCs may be used as a universal and reproducible stromal feeder layer to efficiently expand and maintain human BM HSCs ex vivo [81].
MSCs produce a microenvironment supporting hematopoiesis and may contribute to immune tolerance because of low immunogenicity and the suppressive effect of alloreactivity [75, 82]. MSCs had a potent immunosuppressive effect in vivo after allogeneic stem-cell transplantations [26]. The CXCL12-α secreted by MSCs could reduce the production of a variety of inflammatory cytokines and chemokines, including IL-13, IL-3 Rβ, IL-4, IL-5, IL-9, IL-10, L-selectin, MIP-3α/β, TCA3/CCL1, TNF-a, IL-1β, lymphotactin/CXCL1, L-selectin, leptin receptor, eotaxin-2, CTACK/CCL27, CRG-2/CXCL10, and CD30L [33]. In allogeneic transplantation, the simultaneous infusion of MSCs may promote hematopoietic engraftment across the major histocompatibility complex (MHC) barrier and decrease the incidence of GVHD, even though the exact mechanisms have not been clarified [83–85]. MSCs are lack of MHC class II and most of classical costimulatory molecules [86, 87]. Moreover, MSCs directly inhibit the expansion and activation of alloreactive Tlymphocytes and this T cell-suppressive effect may have important therapeutic implications in preventing or treating acute and chronic GVHD [70]. MSCs can significantly reduced the expression of activation markers CD25 (interleukn-2 receptor), CD38, and CD69 on phytohaemagglutinin- (PHA-) stimulated lymphocytes, making allogeneic HSCs and MSCs escape from recognition by alloreactive T-cells, because the expression of CD25 (IL-2 receptor), CD38 and CD69 was unchanged. Besides, MSC suppressed the proliferation of PHA-stimulated CD3+, CD4+, and CD8+ lymphocytes [87–89]. However, MSCs inhibit naïve and memory T-cell responses to their cognate antigens by the engagement of the inhibitory molecule PD-1 while the expression of MHC molecules and the presence in culture of antigen-presenting cells (APCs) or CD4+/CD25+ regulatory T cells were not required for MSCs to inhibit preferentially [87–91]. MSCs can regulate B-cell functions including migration, proliferation, and immunoglobulin(Ig) synthesis. For example, MSCs inhibit the proliferation of B-cells by arresting them at G0/G1 phase of the cell cycle, and the production of IgM, IgA, and IgG of B-cells [88, 92]. Dendritic cells (DCs) play an important role in supporting antigen-specific CD4+ T-cell proliferation and modulating diverse T-cell responses including GVHD [93]. MSCs can inhibit the differentiation of mature DCs from HSCs by arresting them at the precursor stage, interfere with DC antigen presentation, prevent DC migration ability, and induce DC apoptosis by downregulate TNF-α and TGF-β1 levels and upregulated IL-6 levels [93–95]. IFN-γ, which is produced by donor T-cells in response to antigen recognition, displays natural cytolytic activity against the cells missing markers of self-MHC class I, serves as an initiating stimulus for MSC immunosuppressive activity in vivo [88]. This indicates that the exposure to concentrated amounts of IFN-γ of MSCs can stimulate MSCs to exhibit induction of class II molecule expression, to prevent GVHD and provide the basis for a new potential strategy in prevention of GVHD [87–89, 96]. There is also evidences that MSCs can inhibit the IL-2-induced proliferation of natural killer (NK) cells by producing prostaglandin E2 (PGE2), a product of arachidonic acid metabolism that acts as a powerful immune suppressant, and inhibits T-cell mitogenesis and IL-2 production [88, 97, 98] (Figure 1).
Figure 1.
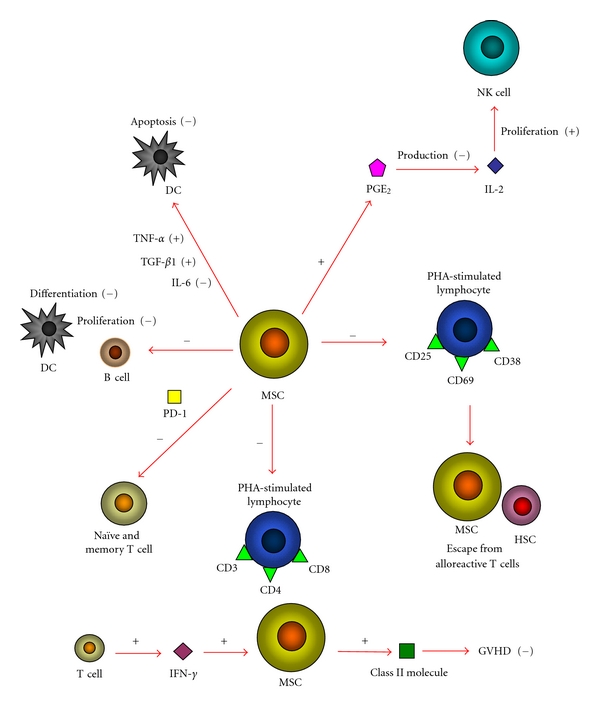
MSCs interact with immune cells, representing potential cellular therapy to enhance allogeneic hematopoietic engraftment and prevent GVHD. MSCs reduced the expression of activation markers CD25, CD38 and CD69 on PHA-stimulated lymphocytes, making allogeneic HSCs and MSCs escape from recognition of alloreactive T-cells. MSCs suppressed the proliferation of PHA-stimulated CD3+, CD4+ and CD8+ lymphocytes. MSCs inhibit naïve and memory T-cell responses to their cognate antigens by the engagement of the inhibitory molecule PD-1. MSCs inhibit the proliferation of B-cells and the differentiation of mature DCs from HSCs. MSCs induce DC apoptosis by downregulate TNF-α and TGF-β1 levels and upregulated IL-6 levels. MSCs inhibit the IL-2-induced proliferation of NK cells by producing PGE2. IFN-γ can stimulate MSCs to exhibit induction of class II molecule expression to prevent GVHD.
5. Conclusion
Lines of evidence have indicated that MSCs are capable of supporting the expansion and differentiation of HSCs and enhancing hematopoietic engraftment in the past two decades, but the exact mechanisms by how MSCs support HSCs are still unclear. MSCs may affect HSCs by producing growth factors and chemokines that take parts in signaling pathways regulating HSCs. Meanwhile, HSCs interact with MSCs though this has been less understood. MSCs can home to injured tissues when coinfused with HSCs [99]. A better understanding of the interaction between MSCs and HSCs will substantially ultimately help develop novel therapies for hematopoietic diseases.
Acknowledgment
This paper was supported by grants from Natural Science Foundation of China (nos. 30871273, 30971496, and U1032003) and from Shenzhen (JC201005280597A) to Y. Wu.
References
- 1.Orkin SH, Zon LI. Hematopoiesis: an evolving paradigm for stem cell biology. Cell. 2008;132(4):631–644. doi: 10.1016/j.cell.2008.01.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Group SCTC. Allogeneic peripheral blood stem-cell compared with bone marrow transplantation in the management of hematologic malignancies: an individual patient data meta-analysis of nine randomized trials. Journal of Clinical Oncology. 2005;23(22):5074–5087. doi: 10.1200/JCO.2005.09.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Murray L, Chen B, Galy A, et al. Enrichment of human hematopoietic stem cell activity in the CD34+Thy- 1+Lin subpopulation from mobilized peripheral blood. Blood. 1995;85(2):368–378. [PubMed] [Google Scholar]
- 4.Flomenberg N, Devine SM, DiPersio JF, et al. The use of AMD3100 plus G-CSF for autologous hematopoietic progenitor cell mobilization is superior to G-CSF alone. Blood. 2005;106(5):1867–1874. doi: 10.1182/blood-2005-02-0468. [DOI] [PubMed] [Google Scholar]
- 5.Gluckman E, Broxmeyer HA, Auerbach AD, et al. Hematopoietic reconstitution in a patient with Fanconi's anemia by means of umbilical-cord blood from an HLA-identical sibling. New England Journal of Medicine. 1989;321(17):1174–1178. doi: 10.1056/NEJM198910263211707. [DOI] [PubMed] [Google Scholar]
- 6.Pierelli L, Scambia G, Bonanno G, et al. CD34+/CD105+ cells are enriched in primitive circulating progenitors residing in the G0 phase of the cell cycle and contain all bone marrow and cord blood CD34+/CD38low/- precursors. British Journal of Haematology. 2000;108(3):610–620. doi: 10.1046/j.1365-2141.2000.01869.x. [DOI] [PubMed] [Google Scholar]
- 7.Morrison SJ, Spradling AC. Stem cells and niches: mechanisms that promote stem cell maintenance throughout life. Cell. 2008;132(4):598–611. doi: 10.1016/j.cell.2008.01.038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Calvi LM, Adams GB, Weibrecht KW, et al. Osteoblastic cells regulate the haematopoietic stem cell niche. Nature. 2003;425(6960):841–846. doi: 10.1038/nature02040. [DOI] [PubMed] [Google Scholar]
- 9.Zhang JW, Niu C, Ye L, et al. Identification of the haematopoietic stem cell niche and control of the niche size. Nature. 2003;425(6960):836–841. doi: 10.1038/nature02041. [DOI] [PubMed] [Google Scholar]
- 10.Méndez-Ferrer S, Lucas D, Battista M, Frenette PS. Haematopoietic stem cell release is regulated by circadian oscillations. Nature. 2008;452(7186):442–447. doi: 10.1038/nature06685. [DOI] [PubMed] [Google Scholar]
- 11.Sugiyama T, Kohara H, Noda M, Nagasawa T. Maintenance of the hematopoietic stem cell pool by CXCL12-CXCR4 chemokine signaling in bone marrow stromal cell niches. Immunity. 2006;25(6):977–988. doi: 10.1016/j.immuni.2006.10.016. [DOI] [PubMed] [Google Scholar]
- 12.Scheller M, Huelsken J, Rosenbauer F, et al. Hematopoietic stem cell and multilineage defects generated by constitutive β-catenin activation. Nature Immunology. 2006;7(10):1021–1023. doi: 10.1038/ni1387. [DOI] [PubMed] [Google Scholar]
- 13.Suda T, Arai F. Wnt signaling in the niche. Cell. 2008;132(5):729–730. doi: 10.1016/j.cell.2008.02.017. [DOI] [PubMed] [Google Scholar]
- 14.Fleming HE, Janzen V, Celso CL, et al. Wnt signaling in the niche enforces hematopoietic stem cell quiescence and is necessary to preserve self-renewal in vivo. Cell Stem Cell. 2008;2(3):274–283. doi: 10.1016/j.stem.2008.01.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Eliasson P, Jönsson J. The hematopoietic stem cell niche: low in oxygen but a nice place to be. Journal of Cellular Physiology. 2010;222(1):17–22. doi: 10.1002/jcp.21908. [DOI] [PubMed] [Google Scholar]
- 16.Hosokawa K, Arai F, Yoshihara H, et al. Function of oxidative stress in the regulation of hematopoietic stem cell-niche interaction. Biochemical and Biophysical Research Communications. 2007;363(3):578–583. doi: 10.1016/j.bbrc.2007.09.014. [DOI] [PubMed] [Google Scholar]
- 17.Pittenger MF, Martin BJ. Mesenchymal stem cells and their potential as cardiac therapeutics. Circulation Research. 2004;95(1):9–20. doi: 10.1161/01.RES.0000135902.99383.6f. [DOI] [PubMed] [Google Scholar]
- 18.Valtieri M, Sorrentino A. The mesenchymal stromal cell contribution to homeostasis. Journal of Cellular Physiology. 2008;217(2):296–300. doi: 10.1002/jcp.21521. [DOI] [PubMed] [Google Scholar]
- 19.Kögler G, Sensken S, Airey JA, et al. A new human somatic stem cell from placental cord blood with intrinsic pluripotent differentiation potential. Journal of Experimental Medicine. 2004;200(2):123–135. doi: 10.1084/jem.20040440. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Tondreau T, Lagneaux L, Dejenefle M, et al. Bone marrow-derived mesenchymal stem cells already express specific neural proteins before any differentiation. Differentiation. 2004;72(7):319–326. doi: 10.1111/j.1432-0436.2004.07207003.x. [DOI] [PubMed] [Google Scholar]
- 21.Dennis JE, Charbord P. Origin and differentiation of human and murine stroma. Stem Cells. 2002;20(3):205–214. doi: 10.1634/stemcells.20-3-205. [DOI] [PubMed] [Google Scholar]
- 22.Dominici M, Le Blanc K, Mueller I, et al. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy. 2006;8(4):315–317. doi: 10.1080/14653240600855905. [DOI] [PubMed] [Google Scholar]
- 23.Imai E, Ito T. Can bone marrow differentiate into renal cells? Pediatric Nephrology. 2002;17(10):790–794. doi: 10.1007/s00467-002-0949-4. [DOI] [PubMed] [Google Scholar]
- 24.Mohanty ST, Kottam L, Gambardella A, et al. Alterations in the self-renewal and differentiation ability of bone marrow mesenchymal stem cells in a mouse model of rheumatoid arthritis. Arthritis Research and Therapy. 2010;12(4):p. R149. doi: 10.1186/ar3098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Koç ON, Gerson SL, Cooper BW, et al. Rapid hematopoietic recovery after coinfusion of autologous-blood stem cells and culture-expanded marrow mesenchymal stem cells in advanced breast cancer patients receiving high-dose chemotherapy. Journal of Clinical Oncology. 2000;18(2):307–316. doi: 10.1200/JCO.2000.18.2.307. [DOI] [PubMed] [Google Scholar]
- 26.Blanc KL, Rasmusson I, Sundberg B, et al. Treatment of severe acute graft-versus-host disease with third party haploidentical mesenchymal stem cells. Lancet. 2004;363(9419):1439–1441. doi: 10.1016/S0140-6736(04)16104-7. [DOI] [PubMed] [Google Scholar]
- 27.Aggarwal S, Pittenger MF. Human mesenchymal stem cells modulate allogeneic immune cell responses. Blood. 2005;105(4):1815–1822. doi: 10.1182/blood-2004-04-1559. [DOI] [PubMed] [Google Scholar]
- 28.Mishima S, Nagai A, Abdullah S, et al. Effective ex vivo expansion of hematopoietic stem cells using osteoblast-differentiated mesenchymal stem cells is CXCL12 dependent. European Journal of Haematology. 2010;84(6):538–546. doi: 10.1111/j.1600-0609.2010.01419.x. [DOI] [PubMed] [Google Scholar]
- 29.Zhang J, Li L. Stem cell niche: microenvironment and beyond. Journal of Biological Chemistry. 2008;283(15):9499–9503. doi: 10.1074/jbc.R700043200. [DOI] [PubMed] [Google Scholar]
- 30.Burness ML, Sipkins DA. The stem cell niche in health and malignancy. Seminars in Cancer Biology. 2010;20(2):107–115. doi: 10.1016/j.semcancer.2010.05.006. [DOI] [PubMed] [Google Scholar]
- 31.Lam BS, Adams GB. Hematopoietic stem cell lodgment in the adult bone marrow stem cell niche. International Journal of Laboratory Hematology. 2010;32(6):551–558. doi: 10.1111/j.1751-553X.2010.01250.x. [DOI] [PubMed] [Google Scholar]
- 32.Burns JM, Summers BC, Wang Y, et al. A novel chemokine receptor for SDF-1 and I-TAC involved in cell survival, cell adhesion, and tumor development. Journal of Experimental Medicine. 2006;203(9):2201–2213. doi: 10.1084/jem.20052144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Thevenot PT, Nair AM, Shen J, Lotfi P, Ko CY, Tang L. The effect of incorporation of SDF-1α into PLGA scaffolds on stem cell recruitment and the inflammatory response. Biomaterials. 2010;31(14):3997–4008. doi: 10.1016/j.biomaterials.2010.01.144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Rappold I, Watt SM, Kusadasi N, Rose-John S, Hatzfeld J, Ploemacher RE. Gp130-signaling synergizes with FL and TPO for the long-term expansion of cord blood progenitors. Leukemia. 1999;13(12):2036–2048. doi: 10.1038/sj.leu.2401597. [DOI] [PubMed] [Google Scholar]
- 35.Han JY, Goh RY, Seo SY, et al. Cotransplantation of cord blood hematopoietic stem cells and culture-expanded and GM-CSF-/SCF-transfected mesenchymal stem cells in SCID mice. Journal of Korean Medical Science. 2007;22(2):242–247. doi: 10.3346/jkms.2007.22.2.242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Bernstein A, Forrester L, Reith AD, Dubreuil P, Rottapel R. The murine W/c-kit and steel loci and the control of hematopoiesis. Seminars in Hematology. 1991;28(2):138–142. [PubMed] [Google Scholar]
- 37.Verfaillie M. Adhesion receptors as regulators of the hematopoietic process. Blood. 1998;92(8):2609–2612. [PubMed] [Google Scholar]
- 38.Schofield R. The relationship between the spleen colony-forming cell and the haemopoietic stem cell. Blood Cells. 1978;4(1-2):7–25. [PubMed] [Google Scholar]
- 39.Garrett RW, Emerson SG. Bone and blood vessels: the hard and the soft of hematopoietic stem cell niches. Cell Stem Cell. 2009;105(4):503–506. doi: 10.1016/j.stem.2009.05.011. [DOI] [PubMed] [Google Scholar]
- 40.Fuchs E, Tumbar T, Guasch G. Socializing with the neighbors: stem cells and their niche. Cell. 2004;116(6):769–778. doi: 10.1016/s0092-8674(04)00255-7. [DOI] [PubMed] [Google Scholar]
- 41.Fierro F, Illmer T, Jing D, et al. Inhibition of platelet-derived growth factor receptor-β by imatinib mesylate suppresses proliferation and alters differentiation of human mesenchymal stem cells in vitro. Cell Proliferation. 2007;40(3):355–366. doi: 10.1111/j.1365-2184.2007.00438.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Sacchetti B, Funari A, Michienzi S, et al. Self-renewing osteoprogenitors in bone marrow sinusoids can organize a hematopoietic microenvironment. Cell. 2007;131(2):324–336. doi: 10.1016/j.cell.2007.08.025. [DOI] [PubMed] [Google Scholar]
- 43.Kiel MJ, Yilmaz OH, Iwashita T, Yilmaz OH, Terhorst C, Morrison SJ. SLAM family receptors distinguish hematopoietic stem and progenitor cells and reveal endothelial niches for stem cells. Cell. 2005;121(7):1109–1121. doi: 10.1016/j.cell.2005.05.026. [DOI] [PubMed] [Google Scholar]
- 44.Yin T, Li L. The stem cell niches in bone. Journal of Clinical Investigation. 2006;116(5):1195–1201. doi: 10.1172/JCI28568. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Raaijmakers MH. Regulating traffic in the hematopoietic stem cell niche. Haematologica. 2010;95(9):1439–1441. doi: 10.3324/haematol.2010.027342. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Heissig B, Hattori K, Dias S, et al. Recruitment of stem and progenitor cells from the bone marrow niche requires MMP-9 mediated release of Kit-ligand. Cell. 2002;109(5):625–637. doi: 10.1016/s0092-8674(02)00754-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Arai F, Hirao A, Ohmura M, et al. Tie2/angiopoietin-1 signaling regulates hematopoietic stem cell quiescence in the bone marrow niche. Cell. 2004;118(2):149–161. doi: 10.1016/j.cell.2004.07.004. [DOI] [PubMed] [Google Scholar]
- 48.Xie Y, Yin T, Wiegraebe W, et al. Detection of functional haematopoietic stem cell niche using real-time imaging. Nature. 2009;457(7225):97–101. doi: 10.1038/nature07639. [DOI] [PubMed] [Google Scholar]
- 49.Taichman RS, Emerson SG. The role of osteoblasts in the hematopoietic microenvironment. Stem Cells. 1998;16(1):7–15. doi: 10.1002/stem.160007. [DOI] [PubMed] [Google Scholar]
- 50.Muguruma Y, Yahata T, Miyatake H, et al. Reconstitution of the functional human hematopoietic microenvironment derived from human mesenchymal stem cells in the murine bone marrow compartment. Blood. 2006;107(5):1878–1887. doi: 10.1182/blood-2005-06-2211. [DOI] [PubMed] [Google Scholar]
- 51.Méndez-Ferrer S, Michurina TV, Ferraro F, et al. Mesenchymal and haematopoietic stem cells form a unique bone marrow niche. Nature. 2010;466(12):829–834. doi: 10.1038/nature09262. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Majumdar MK, Thiede MA, Haynesworth SE, Bruder SP, Gerson SL. Human marrow-derived mesenchymal stem cells (MSCs) express hematopoietic cytokines and support long-term hematopoiesis when differentiated toward stromal and osteogenic lineages. Journal of Hematotherapy and Stem Cell Research. 2000;9(6):841–848. doi: 10.1089/152581600750062264. [DOI] [PubMed] [Google Scholar]
- 53.Shi L, Hu LH. The normal flora may contribute to the quantitative preponderance of myeloid cells under physiological conditions. Medical Hypotheses. 2011;21(12):141–143. doi: 10.1016/j.mehy.2010.09.004. [DOI] [PubMed] [Google Scholar]
- 54.Jeltsch KS, Radke TF, Laufs S, et al. Unrestricted somatic stem cells: interaction with CD34+ cells in vitro and in vivo, expression of homing genes and exclusion of tumorigenic potential. Cytotherapy. 2011;13(3):357–365. doi: 10.3109/14653249.2010.523076. [DOI] [PubMed] [Google Scholar]
- 55.Ratliff BB, Singh N, Yasuda K, et al. Mesenchymal stem cells, used as bait, disclose tissue binding sites. American Journal of Pathology. 2010;177(2):873–883. doi: 10.2353/ajpath.2010.090984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Ahn JY, Park G, Shim JS, Lee JW, Oh IH. Intramarrow injection of β-catenin-activated, but not naïve mesenchymal stromal cells stimulates self-renewal of hematopoietic stem cells in bone marrow. Experimental and Molecular Medicine. 2010;42(2):122–131. doi: 10.3858/emm.2010.42.2.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Tavassoli M, Friedenstein A. Hemopoietic stromal microenvironment. American Journal of Hematology. 1983;15(2):195–203. doi: 10.1002/ajh.2830150211. [DOI] [PubMed] [Google Scholar]
- 58.Marks PA, Richon VM, Rifkind RA. Histone deacetylase inhibitors: inducers of differentiation or apoptosis of transformed cells. Journal of the National Cancer Institute. 2000;92(15):1210–1216. doi: 10.1093/jnci/92.15.1210. [DOI] [PubMed] [Google Scholar]
- 59.Wolffe AP, Jones PL, Wade PA. DNA demethylation. Proceedings of the National Academy of Sciences of the United States of America. 1999;96(11):5894–5896. doi: 10.1073/pnas.96.11.5894. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Koh SH, Choi HS, Park ES, Kang HJ, Ahn HS, Shin HY. Co-culture of human CD34+ cells with mesenchymal stem cells increases the survival of CD34+ cells against the 5-aza-deoxycytidine- or trichostatin A-induced cell death. Biochemical and Biophysical Research Communications. 2005;329(3):1039–1045. doi: 10.1016/j.bbrc.2005.02.077. [DOI] [PubMed] [Google Scholar]
- 61.Tabata M, Satake A, Okura N, et al. Long-term outcome after allogeneic bone marrow transplantation for hematological malignancies with non-remission status. Results of a single-center study of 24 patients. Annals of Hematology. 2002;81(10):582–587. doi: 10.1007/s00277-002-0506-z. [DOI] [PubMed] [Google Scholar]
- 62.Ball LM, Bernardo ME, Roelofs H, et al. Cotransplantation of ex vivo-expanded mesenchymal stem cells accelerates lymphocyte recovery and may reduce the risk of graft failure in haploidentical hematopoietic stem-cell transplantation. Blood. 2007;110(7):2764–2767. doi: 10.1182/blood-2007-04-087056. [DOI] [PubMed] [Google Scholar]
- 63.Hombach-Klonisch S, Panigrahi S, Rashedi I, et al. Adult stem cells and their trans-differentiation potential-perspectives and therapeutic applications. Journal of Molecular Medicine. 2008;86(12):1301–1314. doi: 10.1007/s00109-008-0383-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Iannone R, Casella JF, Fuchs EJ, et al. Results of minimally toxic nonmyeloablative transplantation in patients with sickle cell anemia and β-thalassemia. Biology of Blood and Marrow Transplantation. 2003;9(8):519–528. doi: 10.1016/s1083-8791(03)00192-7. [DOI] [PubMed] [Google Scholar]
- 65.Pai SY, DeMartiis D, Forino C, et al. Stem cell transplantation for the Wiskott-Aldrich syndrome: a single-center experience confirms efficacy of matched unrelated donor transplantation. Biology of Blood and Marrow Transplantation. 2006;38(10):671–679. doi: 10.1038/sj.bmt.1705512. [DOI] [PubMed] [Google Scholar]
- 66.Peters C, Krivit W. Hematopoietic cell transplantation for mucopolysaccharidosis IIB (Hunter syndrome) Bone Marrow Transplantation. 2000;25(10):1097–1099. doi: 10.1038/sj.bmt.1702398. [DOI] [PubMed] [Google Scholar]
- 67.Salerno M, Busiello R, Esposito V, et al. Allogeneic bone marrow transplantation restores IGF-I production and linear growth in a γ-SCID patient with abnormal growth hormone receptor signaling. Bone Marrow Transplantation. 2004;33(7):773–775. doi: 10.1038/sj.bmt.1704421. [DOI] [PubMed] [Google Scholar]
- 68.Staba SL, Escolar ML, Poe M, et al. Cord-blood transplants from unrelated donors in patients with hurler's syndrome. New England Journal of Medicine. 2004;350(19):1960–1969. doi: 10.1056/NEJMoa032613. [DOI] [PubMed] [Google Scholar]
- 69.Kurokawa T, Fischer K, Bertz H, Hoegerle S, Finke J, Mackensen A. In vitro and in vivo characterization of graft-versus-tumor responses in melanoma patients after allogeneic peripheral blood stem cell transplantation. International Journal of Cancer. 2002;101(1):52–60. doi: 10.1002/ijc.10555. [DOI] [PubMed] [Google Scholar]
- 70.Maitra B, Szekely E, Gjini K, et al. Human mesenchymal stem cells support unrelated donor hematopoietic stem cells and suppress T-cell activation. Bone Marrow Transplantation. 2004;33(6):597–604. doi: 10.1038/sj.bmt.1704400. [DOI] [PubMed] [Google Scholar]
- 71.Resnick I, Stepensky P, Elkin G, et al. MSC for the improvement of hematopoietic engraftment. Bone Marrow Transplantation. 2010;45(3):605–606. doi: 10.1038/bmt.2009.199. [DOI] [PubMed] [Google Scholar]
- 72.El Backly RM, Cancedda R. Bone marrow stem cells in clinical application: harnessing paracrine roles and niche mechanisms. Advances in Biochemical Engineering/Biotechnology. 2010;123:265–292. doi: 10.1007/10_2010_78. [DOI] [PubMed] [Google Scholar]
- 73.McNiece I, Harrington J, Turney J, Kellner J, Shpall EJ. Ex vivo expansion of cord blood mononuclear cells on mesenchymal stem cells. Cytotherapy. 2004;6(4):311–317. doi: 10.1080/14653240410004871. [DOI] [PubMed] [Google Scholar]
- 74.Nakao N, Nakayama T, Yahata T, et al. Adipose tissue-derived mesenchymal stem cells facilitate hematopoiesis in vitro and in vivo. American Journal of Pathology. 2010;177(2):547–554. doi: 10.2353/ajpath.2010.091042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Vanikar AV, Trivedi HL, Feroze A, Kanodia KV, Dave SD, Shah PR. Effect of co-transplantation of mesenchymal stem cells and hematopoietic stem cells as compared to hematopoietic stem cell transplantation alone in renal transplantation to achieve donor hypo-responsiveness. International Urology and Nephrology. 2010;43(1):225–232. doi: 10.1007/s11255-009-9659-1. [DOI] [PubMed] [Google Scholar]
- 76.Noort WA, Kruisselbrink AB, In’t Anker PS, et al. Mesenchymal stem cells promote engraftment of human umbilical cord blood-derived CD34+ cells in NOD/SCID mice. Experimental Hematology. 2002;30(8):870–878. doi: 10.1016/s0301-472x(02)00820-2. [DOI] [PubMed] [Google Scholar]
- 77.Lemischka IR. Microenvironmental regulation of hematopoietic stem cells. Stem Cells. 1997;15(1):63–68. doi: 10.1002/stem.5530150810. [DOI] [PubMed] [Google Scholar]
- 78.Angelopoulou M, Novelli E, Grove JE, et al. Cotransplantation of human mesenchymal stem cells enhances human myelopoiesis and megakaryocytopoiesis in NOD/SCID mice. Experimental Hematology. 2003;31(5):413–420. doi: 10.1016/s0301-472x(03)00042-0. [DOI] [PubMed] [Google Scholar]
- 79.Kim DH, Yoo KH, Yim YS, et al. Cotransplanted bone marrow derived mesenchymal stem cells (MSC) enhanced engraftment of hematopoietic stem cells in a MSC-dose dependent manner in NOD/SCID mice. Journal of Korean Medical Science. 2006;21(6):1000–1004. doi: 10.3346/jkms.2006.21.6.1000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.da Silva CL, Gonçalves R, Crapnell KB, Cabral JM, Zanjani ED, Almeida-Porada G. A human stromal-based serum-free culture system supports the ex vivo expansion/maintenance of bone marrow and cord blood hematopoietic stem/progenitor cells. Experimental Hematology. 2005;33(7):828–835. doi: 10.1016/j.exphem.2005.03.017. [DOI] [PubMed] [Google Scholar]
- 81.Gonçalves R, da Silva CL, Cabral JMS, Zanjani ED, Almeida-Porada G. A Stro-1+ human universal stromal feeder layer to expand/maintain human bone marrow hematopoietic stem/progenitor cells in a serum-free culture system. Experimental Hematology. 2006;34(10):1353–1359. doi: 10.1016/j.exphem.2006.05.024. [DOI] [PubMed] [Google Scholar]
- 82.Chung NG, Jeong DC, Park SJ, et al. Cotransplantation of marrow stromal cells may prevent lethal graft-versus-host disease in major histocompatibility complex mismatched murine hematopoietic stem cell transplantation. International Journal of Hematology. 2004;80(4):370–376. doi: 10.1532/ijh97.a30409. [DOI] [PubMed] [Google Scholar]
- 83.Gurevitch O, Prigozhina TB, Pugatsch T, Slavin S. Transplantation of allogeneic or xenogeneic bone marrow within the donor stromal microenvironment. Transplantation. 1999;68(9):1362–1368. doi: 10.1097/00007890-199911150-00024. [DOI] [PubMed] [Google Scholar]
- 84.Stanevsky A, Shimoni A, Yerushalmi R, Nagler A. Cord blood stem cells for hematopoietic transplantation. Stem Cell Reviews and Reports. 2011;7:425–433. doi: 10.1007/s12015-010-9183-9. [DOI] [PubMed] [Google Scholar]
- 85.Battiwalla M, Hematti P. Mesenchymal stem cells in hematopoietic stem cell transplantation. Cytotherapy. 2009;11(5):503–515. doi: 10.1080/14653240903193806. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Sugiura K, Hisha H, Ishikawa J, et al. Major histocompatibility complex restriction between hematopoietic stem cells and stromal cells in vitro. Stem Cells. 2001;19(1):46–58. doi: 10.1634/stemcells.19-1-46. [DOI] [PubMed] [Google Scholar]
- 87.Abdi R, Fiorina P, Adra CN, Atkinson M, Sayegh MH. Immunomodulation by mesenchymal stem cells: a potential therapeutic strategy for type 1 diabetes. Diabetes. 2008;57(7):1759–1767. doi: 10.2337/db08-0180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Newman RE, Yoo D, LeRoux MA, Danilkovitch-Miagkova A. Treatment of inflammatory diseases with mesenchymal stem cells. Inflammation and Allergy-Drug Targets. 2009;8(2):110–123. doi: 10.2174/187152809788462635. [DOI] [PubMed] [Google Scholar]
- 89.Krampera M, Glennie S, Dyson J, et al. Bone marrow mesenchymal stem cells inhibit the response of naive and memory antigen-specific T cells to their cognate peptide. Blood. 2003;101(9):3722–3729. doi: 10.1182/blood-2002-07-2104. [DOI] [PubMed] [Google Scholar]
- 90.Dazzi F, Marelli-Berg FM. Mesenchymal stem cells for graft-versus-host disease: close encounters with T cells. European Journal of Immunology. 2008;38(6):1479–1482. doi: 10.1002/eji.200838433. [DOI] [PubMed] [Google Scholar]
- 91.Le Blanc K, Rasmusson I, Gőtherstrőm C, et al. Mesenchymal stem cells inhibit the expression of CD25 (interleukin-2 receptor) and CD38 on phytohaemagglutinin-activated lymphocytes. Scandinavian Journal of Immunology. 2004;60(3):307–315. doi: 10.1111/j.0300-9475.2004.01483.x. [DOI] [PubMed] [Google Scholar]
- 92.Corcione A, Benvenuto F, Ferretti E, et al. Human mesenchymal stem cells modulate B-cell functions. Blood. 2006;107(1):367–372. doi: 10.1182/blood-2005-07-2657. [DOI] [PubMed] [Google Scholar]
- 93.English K, Barry FP, Mahon BP. Murine mesenchymal stem cells suppress dendritic cell migration, maturation and antigen presentation. Immunology Letters. 2008;115(1):50–58. doi: 10.1016/j.imlet.2007.10.002. [DOI] [PubMed] [Google Scholar]
- 94.Lai HY, Yang MJ, Wen KC, Chao KC, Shih CC, Lee OK. Mesenchymal stem cells negatively regulate dendritic lineage commitment of umbilical-cord-blood-derived hematopoietic stem cells: an unappreciated mechanism as immunomodulators. Tissue Engineering. 2010;16(9):2987–2997. doi: 10.1089/ten.tea.2009.0731. [DOI] [PubMed] [Google Scholar]
- 95.Pulavendran S, Vignesh J, Rose C. Differential anti-inflammatory and anti-fibrotic activity of transplanted mesenchymal vs. hematopoietic stem cells in carbon tetrachloride-induced liver injury in mice. International Immunopharmacology. 2010;10(4):513–519. doi: 10.1016/j.intimp.2010.01.014. [DOI] [PubMed] [Google Scholar]
- 96.Polchert D, Sobinsky J, Douglas G, et al. IFN-γ activation of mesenchymal stem cells for treatment and prevention of graft versus host disease. European Journal of Immunology. 2008;38(6):1745–1755. doi: 10.1002/eji.200738129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Spaggiari GM, Capobianco A, Becchetti S, Mingari MC, Moretta L. Mesenchymal stem cell-natural killer cell interactions: evidence that activated NK cells are capable of killing MSCs, whereas MSCs can inhibit IL-2-induced NK-cell proliferation. Blood. 2006;107(4):1484–1490. doi: 10.1182/blood-2005-07-2775. [DOI] [PubMed] [Google Scholar]
- 98.Ghannam S, Bouffi C, Djouad F, Jorgensen C, Noël D. Immunosuppression by mesenchymal stem cells: mechanisms and clinical applications. Stem Cell Research and Therapy. 2010;1(1):2–8. doi: 10.1186/scrt2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Chapel A, Bertho JM, Bensidhoum M, et al. Mesenchymal stem cells home to injured tissues when co-infused with hematopoietic cells to treat a radiation-induced multi-organ failure syndrome. Journal of Gene Medicine. 2003;5(12):1028–1038. doi: 10.1002/jgm.452. [DOI] [PubMed] [Google Scholar]