

Abstract
Social interactions are a fundamental and adaptive component of the biology of numerous species. Social recognition is critical for the structure and stability of the networks and relationships that define societies. For animals, such as mice, recognition of conspecifics may be important for maintaining social hierarchy and for mate choice 1.
A variety of neuropsychiatric disorders are characterized by disruptions in social behavior and social recognition, including depression, autism spectrum disorders, bipolar disorders, obsessive-compulsive disorders, and schizophrenia. Studies of humans as well as animal models (e.g., Drosophila melanogaster, Caenorhabditis elegans, Mus musculus, Rattus norvegicus) have identified genes involved in the regulation of social behavior 2. To assess sociability in animal models, several behavioral tests have been developed (reviewed in 3). Integrative research using animal models and appropriate tests for social behavior may lead to the development of improved treatments for social psychopathologies.
The three-chamber paradigm test known as Crawley's sociability and preference for social novelty protocol has been successfully employed to study social affiliation and social memory in several inbred and mutant mouse lines (e.g. 4-7). The main principle of this test is based on the free choice by a subject mouse to spend time in any of three box's compartments during two experimental sessions, including indirect contact with one or two mice with which it is unfamiliar. To quantitate social tendencies of the experimental mouse, the main tasks are to measure a) the time spent with a novel conspecific and b) preference for a novel vs. a familiar conspecific. Thus, the experimental design of this test allows evaluation of two critical but distinguishable aspects of social behavior, such as social affiliation/motivation, as well as social memory and novelty. "Sociability" in this case is defined as propensity to spend time with another mouse, as compared to time spent alone in an identical but empty chamber 7. "Preference for social novelty" is defined as propensity to spend time with a previously unencountered mouse rather than with a familiar mouse 7. This test provides robust results, which then must be carefully analyzed, interpreted and supported/confirmed by alternative sociability tests. In addition to specific applications, Crawley's sociability test can be included as an important component of general behavioral screen of mutant mice.
Keywords: Neuroscience, Issue 48, Mice, behavioral test, phenotyping, social interaction
Protocol
1. Equipment and Room Set Up:
The apparatus for Crawley's sociability and preference for social novelty test is comprised a rectangular, three-chamber box. Each chamber is 19 x 45 cm and the dividing walls are made from clear Plexiglas, with an open middle section, which allows free access to each chamber.
Use two identical, wire cup-like containers with removable lids that large enough to hold a single mouse. These are placed vertically inside the apparatus, one in each side chamber, and will contain the naíve/unfamiliar mouse. Each container is comprised of metal wires to allow for air exchange between the interior and exterior of the cylinder but small enough to prevent direct physical interactions between an animal on the inside with one on the outside.
Behavioral testing should be performed between 9:00am and 6:00 pm.
General room lighting is 650 lux.
Employ Observer 6.0 (Noldus) program (or a suitable alternative) to monitor and record the observation and follow up parameters, mentioned in section 4.3.
After each trial, clean all chambers with 70% ethanol (between mice) and then with Clidox 1:5:1 (between the cages) to prevent olfactory cue bias and to ensure proper disinfection, respectively.
The person who makes the observation has to be at least 2 meters away from the apparatus.
2. Animal Preparation:
House three to five mice per cage in a room with a 12 hr light/dark cycle (lights on at 7:00 am) with ad libitum access to food and water, according to standards and requirements of local Animal Care Committee guidance and protocols.
Transfer all the cages containing mice into the behavioral room 30 minutes before the first trial begins.
There are two classes of mouse required for this experiment, one which acts as a control, naíve or "unfamiliarized" animal and one that is the test subject. For the control mouse use a mouse of the same background (usually C57BL6), age (usually 8-12 weeks old), gender and weight, without any prior contact (not littermates) with the subject mouse. Two control mice are required per experiment, one is used for session I and another for session II. The same control mice may be used between trials.
3. Habituation (adaptation):
Isolate the right and left compartments by using the dividing Plexiglass walls.
Place empty wire containment cups in the middle of right and left chamber (one per each site).
Place the subject mouse at the center of the middle chamber for adaptation.
Habituate for 5 minutes.
4. Social Affiliation Aspect of the Test (session I):
Place one of the control mice ("Stranger 1") inside a wire containment cup that is located in one of the side chambers. The placement of Stranger 1 in the left or right side of the chamber is systematically altered between trials.
Remove the walls between the compartments, to allow free access for the subject mouse to explore each of the three chambers.
- Immediately, begin monitoring and recording the following parameters:
- duration and
- number of direct (active) contacts between the subject mouse and the containment cup housing or not housing the Stranger 1 mouse, for each chamber individually. Direct contact between the subject mouse and the containment cup, or stretching of the body of the subject mouse in an area 3-5 cm around the cup is counted as an active contact;
- duration and
- number of other behaviors by the subject mouse in each compartment, including walking, self-grooming, lack of any body movements for more than 5 seconds ("freezing"), as well as unusual behavior, such as jumping, repetitive behavior, etc;
- duration and
- number of entries to each compartment. The mouse is considered to be in the chamber when its head and four paws have entered into the chamber.
The duration of session I is 10 minutes.
5. Social Novelty/preference Session of the Test (Session II):
Place a second control mouse ("Stranger 2") inside an identical wire containment cup in the opposite side chamber (that had been empty during the Session I). Monitor the same parameters described in 4.3, differentiating the behaviors between the subject mouse in the presence of Stranger 1 compared with Stranger 2.
The duration of session II is 10 minutes.
6. Statistical Analysis:
- By using the Origin 6.0 program (or a suitable alternative) analyze the following parameters, using the recorded data:
- the total number of contacts and
- total duration of contacts between experimental mouse and empty containment cup vs or cup housing Stranger 1 (in session I), or between experimental mouse and the cup housing Stranger 1 vs Stranger 2 (in session II);
- mean duration per contact;
- total number and duration of other behaviors (freezing, self-grooming, walking)
- total time spent by the subject mouse in each compartment.
- Analyze the significant differences for each of the aforementioned parameters, by comparing the groups:
- "Empty" containment cup vs "Stranger 1" for a wild type (WT) subject
- "Empty" containment cup vs "Stranger 1" for the experimental mouse (e.g. a knockout, transgenic or drug-treated mouse, etc.).
- "Stranger 1" vs "Stranger 2" for a WT subject
- "Stranger 1" vs "Stranger 2" for the experimental mouse (knockout, transgenic, or drug-treated, etc.).
Present each dataset of the aforementioned parameters as Session I: WT and experimental subjects; Session II: WT and experimental subjects, as shown in Figure 1(A and B) and 2(A and B), respectively.
It is also advisable to take into consideration other parameters, which help in standardizing this test, such as prior experience in recognizing specific mouse behavioral patterns, such as sniffing, grooming, rubbing, fighting, mounting, which can be largely dependent on subjective personal observation. The person acting as the recorder of the behavioral parameters during the test should be blind to the genotype or treatment conditions and this information decoded afterwards.
7. Representative Results:
An example of the two steps of the social interaction test is shown in Figures 1 and 2. The first session of the test allows estimation of social affiliation and motivation of the subject mouse. In this part of the test, the stranger mouse was enclosed in a circular wire cup that allowed nose contact, but prevented fighting between mice. These conditions ensure that social approaches are initiated by the subject mouse and are investigatory only, without direct physical contact. The mouse has the choice to spend more time around the containment cup containing the stranger mouse or to avoid any contact by moving to the compartment with the empty cup. Other parameters, such as number of individual contacts, can be used in additional to duration spent near to the stranger mouse.
Typically, a wild type animal will spend significantly more time in the compartment with Stranger 1 compared to the compartment with empty cup (Figure 1A), indicating normal sociability, social motivation and affiliation. In GSK-3α knockout mice, for example, there are no significant differences between the time spent with empty cup compared to the cup containing stranger 1 (Figure 1B), indicating impaired sociability in these animal model 6.
The second session of the test is designed to estimate social novelty and social memory. In this part, the subject mouse has a free choice between the first, already-investigated, unfamiliar mouse (Stranger 1), and a novel unfamiliar mouse (Stranger 2). Usually, a WT animal recalls its previous contact with Stranger 1, and, in this session, tends to spend more time with the newly encountered mouse (Stranger 2), indicating intact social memory and predilection for novel experiences (Figure 2A).
Unlike wild-type mice, GSK-3α mutant animals do not show preference for the chamber containing a newly introduced mouse (Stranger 2) over a chamber containing a now familiar mouse (Stranger 1) in session II (Figure 2B). Indifferent behavior of KO mice in this test is indicative of decreased social motivation and novelty.
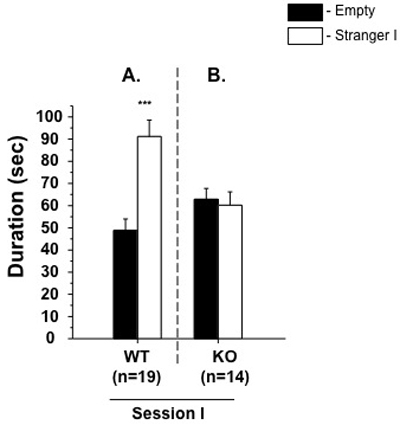 Figure 1. Session I: Social affiliation and sociability. Mean length of time (±SEM) in the chamber with the stranger ("stranger side") compared to the opposite chamber ("empty side"). Unlike WT males (A), KO (B) animals failed to demonstrate a preference for social proximity by spending same time in both chambers. ***p ≤ 0.0001 in WT group.
Figure 1. Session I: Social affiliation and sociability. Mean length of time (±SEM) in the chamber with the stranger ("stranger side") compared to the opposite chamber ("empty side"). Unlike WT males (A), KO (B) animals failed to demonstrate a preference for social proximity by spending same time in both chambers. ***p ≤ 0.0001 in WT group.
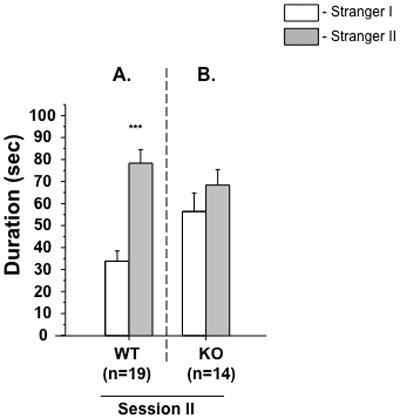 Figure 2. Session II: Social memory and novelty. Mean duration of time (±SEM) in the chamber with the unfamiliar mouse from the sociability phase ("Stranger 1") and in the opposite chamber with a new unfamiliar mouse ("Stranger 2"). Unlike WT males (A), KO (B) animals failed to demonstrate a preference for social novelty by spending same time in both chambers. ***p ≤ 0.0001 in WT group.
Figure 2. Session II: Social memory and novelty. Mean duration of time (±SEM) in the chamber with the unfamiliar mouse from the sociability phase ("Stranger 1") and in the opposite chamber with a new unfamiliar mouse ("Stranger 2"). Unlike WT males (A), KO (B) animals failed to demonstrate a preference for social novelty by spending same time in both chambers. ***p ≤ 0.0001 in WT group.
Discussion
Mus musculus is a social species that engages in high levels of reciprocal social interactions, communal nesting, sexual and parenting behaviors, territorial scent marking and aggressive behaviors 8-10. Several tests have been developed to examine social behavioral in mice (reviewed in 3). Here, we present a widely employed test to evaluate sociability in mice. Crawley's sociability and preference for social novelty test was originally developed by Crawley and colleagues 7, 11. There are several advantages to the three-chamber paradigm, which improve data reliability for examination of sociability, social affiliation, social memory and preference, using similar procedures. The main specificity of this test is the design in which a mouse, previously unencountered by the subject animal, is placed under a wire cup and unable to move freely. This prevents direct physical contact, eliminating fighting and/or aggressive behavior but allows sensory interactions (smell, sight, sound, taste). Under these conditions, the experimental mouse initiates and terminates any interactions, always having a choice between unfamiliar mouse vs empty container (in session 1), or between familiar mouse (Stranger 1) vs the novel mouse (Stranger 2). For example, in one-chamber social interaction tests, the subject mouse is exposed to a stranger in the same chamber, and since both mice are able to move freely the aforementioned active physical contact tends to be dominant and cannot be eliminated. The three-chamber apparatus is much larger than a typical single chamber device, which allows the subject mouse to remain in proximity to or move far away from the Stranger mouse. Thus, the stress level of Crawley's sociability test is low and mostly caused by placing the experimental mouse into a novel environment, which allows repetition of this test over time, for example at different ages of the same mouse cohort: juvenile vs adult. By comparing a young group of animals with the same but older cohort, it is possible to follow up and discriminate between impairment in social recognition vs social memory and/or cognitive impairments, which can be observed in Session 2, and be related to age-dependent physiology.
This test is relatively time-consuming (~40 minutes per mouse), which is a disadvantage. However, multiple repertoires of specific parameters can be monitored and recoded (manually and/or video), permitting robust analysis. For example, some of additional parameters, such as freezing and/or self-grooming behavior, may indicate increased anxiety and decreased motivation for social behaviour. By using the same protocol, mice can be retested in the next day with the same Stranger 1 mouse to rule out an effect of social anxiety. Additional parameters (such as tail rattling, biting of the wire cup where Stranger 1 is located) can be helpful in distinguishing aggressive versus unusual friendliness behavior towards the novel animal. Observed anxiety can be due to inappropriate adaptation of the mice to the experimental room, excessive room lighting, poor mouse health, or as a result of genetic modification and/or neophobia (in some KO models). Moreover, impaired habituation may be interpreted as a cognitive impairment, and may affect the results of second session. Before subjecting mice to this test, it is important to subject animals to general health and neurological screening, including body temperature and weight, coat examination, and the series of following reflex examinations: Whisker, Righting, Ear-Twitch, Reaching and Key Jangling (described in 12-15). Additional behavioral tests can be helpful in discriminating specific responses. An increased number of contacts and/or an increase in the total duration of active contacts maybe associated with hyperactive behavior of specific mouse stains, which can be confirmed by using an open field test (to measure locomotor activity). Decreased duration and/or number of contacts may be associated with depressive- and/or anxiety-like behavior, which can be evaluated by using FST, TST or EPM, zero-maze, light/dark tests, respectively (described in 6, 13, 15).
Once appropriate conditions are applied and significant differences observed, there is a requirement to perform follow-up tests, in order to confirm the findings. Olfaction plays a key role in social recognition in rodents, since either chemically induced anosmia or removal of the vomeronasal organ blocks individual recognition 16, 17. Mice tend to sniff a novel odour and then quickly habituate to its novelty. The olfactory bulb test can be used for this purpose (described in 6, 7).
There are also modifications and alternative usage of three-compartment chamber sociability test, such as estimation of maternal behavior, by examining the time spent by a subject female proximal to a cup containing her pups (reflecting maternal motivation), as well as sexual motivation, by assessing the response of a tested male/female to a female/male under cup instead of Stranger 1 in session 1 7, 11.
Further tests can be used to examine sociability in mice by using alternative approaches/paradigms, such as social interaction in the home cage and/or in a novel environment (one-chamber social interaction test), resident intruder, partition test, social approach test, reciprocal social interaction, etc (described in 3, 18). Vocalizations in the ultrasonic and sonic ranges, visual cues, gustatory and tactile modalities may also contribute to communication of information and to social bonding (reviewed in 19).
Impaired sociability and dysregulated social behavior has been implicated as a hallmark for many psychiatric disorders. For example, autism spectrum disorder and schizophrenia are both characterized by marked deficits in social cognition and social behavior 20, and disruptions of social relationships result in an increased risk of depression 21. Thus, the described here test can be used in animal models of the aforementioned conditions. Moreover, this test is applicable for assessing potential effects of pharmacological compounds on sociability (reviewed in 3, 22-24).
Disclosures
No conflicts of interest declared.
Acknowledgments
OKB and JRW wrote the manuscript. This work was supported by CIHR grant MOP 74711 (to JRW).
References
- Berry RJ, Bronson FH. Life history and bioeconomy of the house mouse. Biological reviews of the Cambridge Philosophical Society. 1992;67:519–550. doi: 10.1111/j.1469-185x.1992.tb01192.x. [DOI] [PubMed] [Google Scholar]
- Robinson GE, Grozinger CM, Whitfield CW. Sociogenomics: social life in molecular terms. Nature reviews. 2005;6:257–270. doi: 10.1038/nrg1575. [DOI] [PubMed] [Google Scholar]
- Silverman JL, Yang M, Lord C, Crawley JN. Behavioural phenotyping assays for mouse models of autism. Nat Rev Neurosci. 2010;11:490–502. doi: 10.1038/nrn2851. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clapcote SJ, Lipina TV, Millar JK, Mackie S, Christie S, Ogawa F, Lerch JP, Trimble K, Uchiyama M, Sakuraba Y, Kaneda H. Behavioral phenotypes of Disc1 missense mutations in mice. Neuron. 2007;54:387–402. doi: 10.1016/j.neuron.2007.04.015. [DOI] [PubMed] [Google Scholar]
- Labrie V, Lipina T, Roder JC. Mice with reduced NMDA receptor glycine affinity model some of the negative and cognitive symptoms of schizophrenia. Psychopharmacology. 2008;200:217–230. doi: 10.1007/s00213-008-1196-6. [DOI] [PubMed] [Google Scholar]
- Kaidanovich-Beilin O, Lipina TV, Takao K, Eede Mvan, Hattori S, Laliberte C, Khan M, Okamoto K, Chambers JW, Fletcher PJ, Macaulay K, Doble BW, Henkelman M, Miyakawa T, Roder J, Woodgett JR. Abnormalities in brain structure and behavior in GSK-3alpha mutant mice. Molecular brain. 2009;2:35–35. doi: 10.1186/1756-6606-2-35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moy SS, Nadler JJ, Perez A, Barbaro RP, Johns JM, Magnuson TR, Piven J, Crawley JN. Sociability and preference for social novelty in five inbred strains: an approach to assess autistic-like behavior in mice. Genes, brain, and behavior. 2004;3:287–302. doi: 10.1111/j.1601-1848.2004.00076.x. [DOI] [PubMed] [Google Scholar]
- Carter CS, Williams JR, Witt DM, Insel TR. Oxytocin and social bonding. Annals of the New York Academy of Sciences. 1992. pp. 652–6204. [DOI] [PubMed]
- Miczek KA, Maxson SC, Fish EW, Faccidomo S. Aggressive behavioral phenotypes in mice. Behavioural brain research. 2001;125:167–181. doi: 10.1016/s0166-4328(01)00298-4. [DOI] [PubMed] [Google Scholar]
- Arakawa H, Arakawa K, Blanchard DC, Blanchard RJ. A new test paradigm for social recognition evidenced by urinary scent marking behavior in C57BL/6J mice. Behavioural brain research. 2008;190:97–104. doi: 10.1016/j.bbr.2008.02.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crawley JN. Designing mouse behavioral tasks relevant to autistic-like behaviors. Mental retardation and developmental disabilities research reviews. 2004;10:248–258. doi: 10.1002/mrdd.20039. [DOI] [PubMed] [Google Scholar]
- Miyakawa T, Leiter LM, Gerber DJ, Gainetdinov RR, Sotnikova TD, Zeng H, Caron MG, Tonegawa S. Conditional calcineurin knockout mice exhibit multiple abnormal behaviors related to schizophrenia. Proceedings of the National Academy of Sciences of the United States of America. 2003;100:8987–8992. doi: 10.1073/pnas.1432926100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yamasaki N, Maekawa M, Kobayashi K, Kajii Y, Maeda J, Soma M, Takao K, Tanda K, Ohira K. Alpha-CaMKII deficiency causes immature dentate gyrus, a novel candidate endophenotype of psychiatric disorders. Molecular brain. 2008;1:6–6. doi: 10.1186/1756-6606-1-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crawley J. What's Wrong with My Mouse? Behavioral Phenotyping of Transgenic and Knockout Mice. New York: John Wiley & Sons, Inc; 2000. [Google Scholar]
- Holmes A, Kinney JW, Wrenn CC, Li Q, Yang RJ, Ma L, Vishwanath J, Saavedra Galanin GAL-R1 receptor null mutant mice display increased anxiety-like behavior specific to the elevated plus-maze. Neuropsychopharmacology. 2003;28:1031–1044. doi: 10.1038/sj.npp.1300164. [DOI] [PubMed] [Google Scholar]
- Matochik JA. Role of the main olfactory system in recognition between individual spiny mice. Physiology & behavior. 1988;42:217–222. doi: 10.1016/0031-9384(88)90073-x. [DOI] [PubMed] [Google Scholar]
- Bluthe RM, Dantzer R. Role of the vomeronasal system in vasopressinergic modulation of social recognition in rats. Brain research. 1993;604:1–2. doi: 10.1016/0006-8993(93)90370-3. [DOI] [PubMed] [Google Scholar]
- Tanda K, Nishi A, Matsuo N, Nakanishi K, Yamasaki N, Sugimoto T, Toyama K, Takao K, Miyakawa T. Abnormal social behavior, hyperactivity, impaired remote spatial memory, and increased D1-mediated dopaminergic signaling in neuronal nitric oxide synthase knockout mice. Molecular brain. 2009;2:19–19. doi: 10.1186/1756-6606-2-19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scattoni ML, Crawley J, Ricceri L. Ultrasonic vocalizations: a tool for behavioural phenotyping of mouse models of neurodevelopmental disorders. Neuroscience and biobehavioral reviews. 2009;33:508–515. doi: 10.1016/j.neubiorev.2008.08.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Couture SM, Penn DL, Losh M, Adolphs R, Hurley R, Piven J. Comparison of social cognitive functioning in schizophrenia and high functioning autism: more convergence than divergence. Psychological medicine. 2010;40:569–579. doi: 10.1017/S003329170999078X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zisook S, Shuchter SR. Depression through the first year after the death of a spouse. The American journal of psychiatry. 1991;148:1346–1352. doi: 10.1176/ajp.148.10.1346. [DOI] [PubMed] [Google Scholar]
- Trainor BC, Workman JL, Jessen R, Nelson RJ. Impaired nitric oxide synthase signaling dissociates social investigation and aggression. Behavioral neuroscience. 2007;121:362–369. doi: 10.1037/0735-7044.121.2.362. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Volke V, Soosaar A, Koks S, Bourin M, Mannisto PT, Vasar E. 7-Nitroindazole, a nitric oxide synthase inhibitor, has anxiolytic-like properties in exploratory models of anxiety. Psychopharmacology. 1997;131:399–405. doi: 10.1007/s002130050309. [DOI] [PubMed] [Google Scholar]
- Griffin MG, Taylor GT. Norepinephrine modulation of social memory: evidence for a time-dependent functional recovery of behavior. Behavioral. 1995;109:466–473. doi: 10.1037//0735-7044.109.3.466. [DOI] [PubMed] [Google Scholar]