

Abstract
Swimming performance tests of fish have been integral to studies of muscle energetics, swimming mechanics, gas exchange, cardiac physiology, disease, pollution, hypoxia and temperature. This paper describes a flexible protocol to assess fish swimming performance using equipment in which water velocity can be controlled. The protocol involves one to several stepped increases in flow speed that are intended to cause fish to fatigue. Step speeds and their duration can be set to capture swimming abilities of different physiological and ecological relevance. Most frequently step size is set to determine critical swimming velocity (Ucrit), which is intended to capture maximum sustained swimming ability. Traditionally this test has consisted of approximately ten steps each of 20 min duration. However, steps of shorter duration (e.g. 1 min) are increasingly being utilized to capture acceleration ability or burst swimming performance. Regardless of step size, swimming tests can be repeated over time to gauge individual variation and recovery ability. Endpoints related to swimming such as measures of metabolic rate, fin use, ventilation rate, and of behavior, such as the distance between schooling fish, are often included before, during and after swimming tests. Given the diversity of fish species, the number of unexplored research questions, and the importance of many species to global ecology and economic health, studies of fish swimming performance will remain popular and invaluable for the foreseeable future.
Keywords: Physiology, Issue 51, fish, swimming, Ucrit, burst, sustained, prolonged, schooling performance
Protocol
1. Capture and Acclimation
Using a knotless net, collect individual fish from their holding tank. Minimize capture time, avoid collecting multiple fish and do not hold fish for more than a few seconds in the net as these factors can increase stress. Stress can affect swimming performance.
Place fish either in a transfer tank with anesthesia or directly to the swim tunnel respirometer ('swim tunnel'). If fish are to be anesthetized, either tricaine methanesulfonate (MS222) or clove oil can be used. Acceptable concentrations will depend on many factors, including species sensitivity and temperature, but typically range from 10-100 mg/L for either anesthetic1,2. The anesthetic concentration should induce stage 3 anesthesia (i.e. unresponsive to external stimuli) and permit rapid recovery (i.e. seconds). When using MS222 in freshwater, sodium bicarbonate should be used at equal parts (by mass) as a buffer. Without buffer, MS222 will greatly reduce pH, and low pH may harm gill tissue. Buffering is not necessary in seawater as it contains sufficient buffering capacity.
Anesthetized fish can be sexed, measured for mass and length, and given a cursory external exam of their condition. Length can be taken as total length (TL), fork length (FL) or body length (BL). These different metrics are typically applied according to standard practice, species morphology (e.g. no forked fin) or fish condition (e.g. caudal fin worn away).
Both anesthetized and non-anesthetized fish should be introduced in the swim tunnel as rapidly as possible and into flowing water as this will facilitate recovery. The flow rate is typically set such that fish will be able to rest on the bottom while barely swimming. An example rate for adult sockeye salmon is 0.3 BL/s.
Fish need to be left in the swim tunnel to acclimate. Acclimation period can be set to traditional periods (e.g. 12 hr), or to those used in most current studies which can be as short as 30 min. To determine an ideal acclimation period for a given fish, the time taken to reach a stable, ideally low, metabolic rate can be used (detailed next section).
2. Measuring Metabolic Rates
Changes in oxygen within the swim tunnel should be recorded over the duration of the swim test(s). Oxygen probes can be sealed directly into the swim tunnel; however, it must first be determined whether water flow over the probe affects the function of the probe. If the probe is affected by water speeds that may occur during the test, the probe will need to be placed in its own chamber outside the tunnel and supplied with tunnel water by a pump.
To replenish oxygen and mitigate waste accumulation, the swim tunnel will likely need to be flushed periodically. Oxygen saturation should not be permitted to fall lower than 70% for most species as swimming performance may be reduced or there may be a shift to anaerobic metabolism. Flushing tunnel water may also help reduce any increases in water temperature; however, water temperature should be held constant by coupling the swim tunnel to a chiller. Changes in temperature will affect metabolic rate.
Routine and maximum metabolic rates, abbreviated 'RMR' and 'MMR', are typically determined (Figure 1A). These rates can be expressed in volume (Vo2) or mass (Mo2) of oxygen per unit mass, per unit time, e.g. mg O2 / kg / hr. The difference between the two rates can be taken to get the 'scope for activity'. The 'oxygen debt' following exercise, referred to as 'excess post-exercise oxygen consumption' (EPOC), can also be determined and is equal to the oxygen required during the post-exercise return to RMR.
Note: many oxygen probes and their output will automatically provide temperature-corrected changes in dissolved oxygen. Those that do not will need to be corrected. Additionally, some values will need to be corrected if measurements were conducted in saline conditions.
Swimming performance measurement
A variety of swimming performance abilities can be assessed in a swim tunnel. The most common way to characterize swimming performance is by the duration for which it can be maintained3. The slower the speed, the longer it can be maintained and vice versa. Speeds are generally referred to as 'burst', 'prolonged' or 'sustained', which correspond to durations of seconds, minutes to hours (max = 200 min), and hours and beyond (>200 min), respectively. To determine these speeds, fish are subjected to user defined 'steps', consisting of set 'height', i.e. water flow speed increase, and 'length', i.e. step duration. Maximizing step height and minimizing step length will capture the greatest burst speed, while doing the reverse will capture the greatest sustained speed. In step tests, the traditional way to report the maximum speed achieved is equal to the speed of the last fully completed step plus the temporal portion of the final, unfinished step (Figure 1A).
Burst swimming performance assessment
To capture burst swimming or sprint ability, following acclimation, bring the flow speed to an estimated maximum (e.g. twice Ucrit; see below), and motivate the fish to swim. Fish should be resting at the rear of the chamber as the flow speed is rapidly increased. Fish may need to be motivated to swim. Motivation can be mechanical (e.g. touch) or electrical (e.g. a small charge applied to a shock grid at the back of the swim tunnel).Although not specifically a test of burst ability, fish can also be given a 'constant acceleration test' (CAT or UΔV) consisting of several steps of short duration (e.g. 1 min)4,5.
Record the duration of swimming. The number of individual burst and their distance can also be recorded6. Speed can be given as the maximum achieved or calculated according to the equation in Figure 1A.Prolonged and sustained swimming performance assessmentThe most frequent measure used to estimate prolonged or sustained swimming ability is referred to as 'critical swimming performance' (Ucrit). Step length for Ucrit has traditionally been set at 20 min, although shorter times such as 10 min have also been used. A general rule is that at least ten steps should be taken prior to fatigue, which can result in lengthy tests, e.g. >3 hrs/fish. To shorten the Ucrit test, the initial seven steps can be abbreviated to 5 min (the test is then termed 'ramp- Ucrit').
Following acclimation and/or a practice swim, increase flow speed according to step size.
To set the appropriate step height for Ucrit, it will likely be necessary to estimate Ucrit for a sample of the fish to be tested. This can be accomplished by using literature values. Alternately, fish can be given a 'practice swim' after introduction to the tunnel. For this, give fish short, user-defined steps, until fatigue. Use the final speed reached as an early estimate of Ucrit. Adjust step size (height and length) for each fish.
To test recovery ability or determine individual variation in performance, fish can be given a second swimming test following a recovery period7-9.
Fish can also be tested in schools, although fish tend to fatigue at different times during the tests which can be problematic for removing them from the tunnel and eliminate the ability to measure oxygen uptake.
Once fatigued, fish should be allowed to recover or removed immediately for tissue sampling.Several performance metrics related to swimming can be collected during the tests or after using video analysis (Figure 1 C,D).
To measure ventilation rate and tail beat frequency, the opercular pumping rate and number of tail beats should be sampled over periods of several seconds (Figure 1B)10,11.
To measure tail beat amplitude, the maximum deflection of the caudal fin can be measured (Figure 1B)12.
Following testing, fish can be euthanized, given a post-mortem exam and tissues collected to determine hematocrit (packed red cell volume), stress hormone concentrations, muscle energy stores (e.g. phosphocreatine, glycogen), enzyme activities (e.g. lactate dehydrogenase, acetylcholine esterase), or other physiological parameters.
3. Representative Results
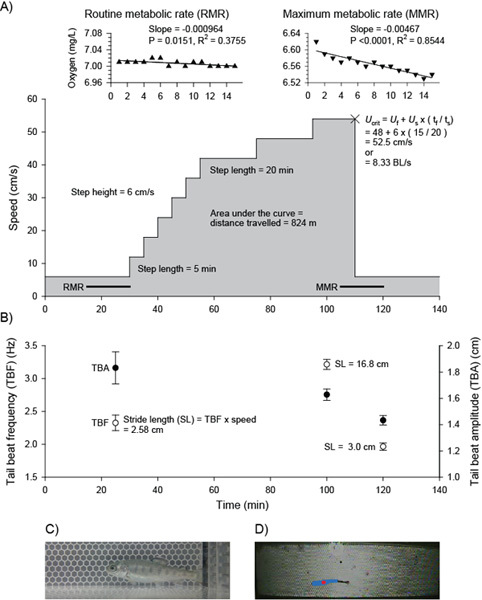 Figure 1. A) An example ramp-Ucrit test given to a juvenile salmonid, a hatchery raised rainbow trout (Oncorhynchus mykiss), 6.3 cm in body length (BL) and 3.5 g in mass. The test was carried out in a 12 L modified Brett-type respirometer (Loligo Systems, Denmark; www.loligosystems.com). Routine and maximum metabolic rates were calculated as slope (mg/hr) × volume (L) / fish mass (kg), and were 198 and 961 mg O2/kg/hr, respectively (scope for activity = 4.84). These rates are in keeping with those reported by Brett for juvenile sockeye salmon (O. nerka)13. Note: for oxygen uptake calculations, the volume of water is equal to the respirometer volume less fish volume. The cross indicates the point where the fish refused to swim further and where Ucrit was calculated. B) Tail beat frequency values were recorded over 10 s six times; tail beat amplitude values were recorded three times; values shown are mean ± SEM. C) An enlarged side view and D) top view of the fish swimming within the swim chamber. D) Shows the fish tracked by EthoVision software (Noldus, Netherlands; www.noldus.com). Abbreviations: Ucrit = critical swimming velocity; Uf = speed of the last fully completed step, Us = step speed, tf = time spent on last step, ts = step time.
Figure 1. A) An example ramp-Ucrit test given to a juvenile salmonid, a hatchery raised rainbow trout (Oncorhynchus mykiss), 6.3 cm in body length (BL) and 3.5 g in mass. The test was carried out in a 12 L modified Brett-type respirometer (Loligo Systems, Denmark; www.loligosystems.com). Routine and maximum metabolic rates were calculated as slope (mg/hr) × volume (L) / fish mass (kg), and were 198 and 961 mg O2/kg/hr, respectively (scope for activity = 4.84). These rates are in keeping with those reported by Brett for juvenile sockeye salmon (O. nerka)13. Note: for oxygen uptake calculations, the volume of water is equal to the respirometer volume less fish volume. The cross indicates the point where the fish refused to swim further and where Ucrit was calculated. B) Tail beat frequency values were recorded over 10 s six times; tail beat amplitude values were recorded three times; values shown are mean ± SEM. C) An enlarged side view and D) top view of the fish swimming within the swim chamber. D) Shows the fish tracked by EthoVision software (Noldus, Netherlands; www.noldus.com). Abbreviations: Ucrit = critical swimming velocity; Uf = speed of the last fully completed step, Us = step speed, tf = time spent on last step, ts = step time.
Discussion
In a natural setting, behaviors such as predator escape and migration depend on being able to swim at specific intensities for different times. In a lab setting, swim tunnel respirometers and stepped increases in speed can be used to estimate the ability of fish to perform numerous behaviours. Measures of metabolic rate and other tissue-level assessments (such as lactate production) can be included with swim tests to help determine physiological mechanisms underlying different swimming modes.
Swim tunnel design can greatly influence swimming performance. There are several types of swim tunnels used to determine swimming performance, most are characterized as either Brett13- or Blazka14-types. For either design, major considerations include the 'evenness' of the water flow (how rectilinear it is), the maximum flow speed and the length of the swim chamber. Evenness is important because fish will instinctively find and swim in low flow areas to minimize effort. High potential flow rate may be necessary to test burst swimming ability or fast swimming species (e.g. tunas). Swim chamber length can be very meaningful as in chambers of small length, i.e. fish length > 10 % of chamber length, fish may be unable to utilize all swimming modes. Specifically, small tubes may restrict the ability of fish to transition from a steady to unsteady swimming gate, i.e. to burst and coast swimming. Therefore, fish may stop swimming at approximately the same speed regardless of the step length and Ucrit may be analogous to 'gate transition speed'15.
All swim tunnels need to be calibrated using devices such as flow probes or particle imaging velocimetry (PIV) equipment. Ideally swim tunnels will have rectilinear flow, but if fish are found to favor an area, their swim speed can be adjusted based on local differences in flow.
Disclosures
No conflicts of interest declared.
Acknowledgments
The author thanks Barbara Berli for providing example data and the gracious funding of the Canadian Wildlife Federation.
References
- Anderson WG, McKinley RS, Colavecchia M. The use of clove oil as an anesthetic for rainbow trout and its effects on swimming performance. North Am. J. Fish Manage. 1997;17:301–307. [Google Scholar]
- Cho GK, Heath DD. Comparison of tricaine methanesulphonate (MS222) and clove oil anaesthesia effects on the physiology of juvenile chinook salmon Oncorhynchus tshawytscha (Walbaum) Aquacult. Res. 2000;31:537–546. [Google Scholar]
- Beamish FWH. In: Swimming capacity in Fish Physiology. Hoar WS, Randall DJ, editors. New York: Academic Press Inc; 1978. pp. 101–187. Vol. 7. [Google Scholar]
- Farrell AP. Comparisons of swimming performance in rainbow trout using constant acceleration and critical swimming speed tests. J. Fish Biol. 2008;72:693–710. [Google Scholar]
- Tierney KB, Casselman M, Takeda S, Farrell AP, Kennedy CJ. The relationship between cholinesterase inhibition and two types of swimming performance in chlorpyrifos-exposed coho salmon (Oncorhynchus kisutch) Environ. Toxicol. Chem. 2007;26:998–1004. doi: 10.1897/06-459r.1. [DOI] [PubMed] [Google Scholar]
- Martinez M, Bedard M, Dutil J-D, Guderley H. Does condition of Atlantic cod (Gadus morhua) have a greater impact upon swimming performance at Ucrit or sprint speeds. J. Exp. Biol. 2004;207:2979–2990. doi: 10.1242/jeb.01142. [DOI] [PubMed] [Google Scholar]
- Marras S, Claireaux G, McKenzie DJ, Nelson JA. Individual variation and repeatability in aerobic and anaerobic swimming performance of European sea bass, Dicentrarchus labrax. J. Exp. Biol. 2010;213:26–32. doi: 10.1242/jeb.032136. [DOI] [PubMed] [Google Scholar]
- Kolok AS, Plaisance EP, Abdelghani A. Individual variation in the swimming performance of fishes: an overlooked source of variation in toxicity studies. Environ. Toxicol. Chem. 1998;17:282–285. [Google Scholar]
- Tierney KB, Farrell AP. The relationships between fish health, metabolic rate, swimming performance and recovery in return-run sockeye salmon, Oncorhynchus nerka (Walbaum) J. Fish Dis. 2004;27:663–671. doi: 10.1111/j.1365-2761.2004.00590.x. [DOI] [PubMed] [Google Scholar]
- Webber JD. Upstream swimming performance of adult white sturgeon: Effects of partial baffles and a ramp. Trans. Am. Fish. Soc. 2007;136:402–408. [Google Scholar]
- Farrell AP, Gamperl AK, Birtwell IK. Prolonged swimming, recovery and repeat swimming performance of mature sockeye salmon Oncorhynchus nerka exposed to moderate hypoxia and pentachlorophenol. J. Exp. Biol. 1998;201:2183–2193. doi: 10.1242/jeb.201.14.2183. [DOI] [PubMed] [Google Scholar]
- Flammang BE, Lauder GV. Speed-dependent intrinsic caudal fin muscle recruitment during steady swimming in bluegill sunfish, Lepomis macrochirus. J. Exp. Biol. 2008;211:587–598. doi: 10.1242/jeb.012096. [DOI] [PubMed] [Google Scholar]
- Brett JR. The respiratory metabolism and swimming performance of young sockeye. J. Fish. Res. Board Can. 1964;21:1183–1226. [Google Scholar]
- Blažka P, Volf M, Čepela M. A new type of respirometer for the determination of the metabolism of fish in an active state. Physiol. Bohemoslov. 1960;9:553–558. [Google Scholar]
- Peake SJ, Farrell AP. Fatigue is a behavioural response in respirometer confined smallmouth bass. J. Fish Biol. 2006;68:1742–1755. [Google Scholar]