

Abstract
Anti-CD3 immunotoxins, which induce profound but transient T cell depletion in vivo by inhibiting eukaryotic protein synthesis in CD3+ cells, are effective reagents in large animal models of transplantation tolerance and autoimmune disease therapy. A diphtheria toxin based anti-porcine CD3 recombinant immunotoxin was constructed by fusing the truncated diphtheria toxin DT390 with two identical tandem single chain variable fragments (scFv) derived from the anti-porcine CD3 monoclonal antibody 898H2-6-15. The recombinant immunotoxin was expressed in a diphtheria-toxin resistant yeast Pichia pastoris strain under the control of the alcohol oxidase promoter. The secreted recombinant immunotoxin was purified sequentially with hydrophobic interaction chromatography (Butyl 650 M) followed by strong anion exchange (Poros 50 HQ). The purified anti-porcine CD3 immunotoxin was tested in vivo in four animals; peripheral blood CD3+ T cell numbers were reduced by 80% and lymph node T cells decreased from 74% CD3+ cells pretreatment to 24% CD3+ cells remaining in the lymph node following 4 days of immunotoxin treatment. No clinical toxicity was observed in any of the experimental swine. We anticipate that this conjugate will provide an important tool for in vivo depletion of T cells in swine transplantation models.
INTRODUCTION
Previous work has demonstrated that chemically conjugated anti-porcine CD3 immunotoxin is a very effective T cell depletion reagent in pigs 1 and this reagent has played a role in the maintenance of long-term hematopoietic stem cell transplants in the absence of graft versus host disease.2-4 The chemically conjugated immunotoxin was created by cross-linking the monoclonal antibody to a diphtheria toxin (DT) binding site mutant, CRM9, such that the binding site of the immunotoxin was dictated by the antibody moiety.5 However, several problems were associated with the chemical conjugate including linkage heterogeneity, low yield and strict limitations in dosage due to non-specific neurological toxicity in pigs.1,6
Recombinant anti-human and anti-monkey CD3 T cell immunotoxins have been developed.7,8 These immunotoxins contain a diphtheria toxin protein sequence that is truncated at amino acid residue 390. The antibody moiety is placed C-terminal to the truncated toxin DT390 to prevent any interference with the translocation of biologically active diphtheria toxin A chain.9,10 However, it was found that when the scFv of an anti-human CD3 monoclonal antibody UCHT1 was fused to the C-terminus of DT390, its binding activity was dramatically reduced by a factor of ten. Adding a second scFv moiety separated with a (G4S)3 linker resulted in a 10-fold increase in binding activity compared to a monovalent fusion immunotoxin. This fusion immunotoxin, designated A-dmDT390biscFv(UCHT1), displayed an increase in potency of 10 to 30-fold as compared with the corresponding chemically conjugated immunotoxin and depleted 2.4 logs of T cells in the lymph node compartment of transgenic mice expressing human CD3ε.11 The clinical trial for this immunotoxin in five patients with cutaneous T cell lymphoma has shown promising results.12
In this paper we present the development of the anti-porcine CD3 recombinant immunotoxin, A-dmDT390biscFv(2-6-15). The VL (variable light chain) and VH (variable heavy chain) of the anti-porcine CD3 monoclonal antibody 898H2-6-1513 were cloned by PCR and the resulting sequence was used to synthesize codon-optimized anti-porcine CD3 scFv (2-6-15) DNA sequence suitable for the Pichia pastoris expression. The biscFv format anti-porcine CD3 recombinant immunotoxin was expressed in a diphtheria-toxin resistant yeast Pichia pastoris strain and purified in a two step chromatography protocol. The in vivo porcine CD3 T cell depletion profile for this immunotoxin was assessed.
EXPERIMENTAL PROCEDURES
Cloning the VL and VH of the anti-porcine CD3 monoclonal antibody 898H2-6-15
To construct the anti-porcine CD3 recombinant immunotoxin, the scFv DNA sequence of hybridoma 898H2-6-15 was obtained by RT-PCR. RNA was isolated from the anti-porcine CD3 hybridoma cell line 898H2-6-15 developed and characterized in our lab 13 with the RNeasy kit (Invitrogen). cDNA was generated from DNAse (Invitrogen) treated RNA using oligo dT primer and the Superscript III kit (Invitrogen). Preliminary VH sequence was isolated using a commercially available Mouse Ig Primer Set (EMD BioSciences), specifically primers MuIgVH5′-A and MuIgGVH3′-2. Gene specific primers were designed and used for generating cDNA (2-6-15 Hr-1) and for 5′ Rapid Amplification of cDNA Ends (RACE) PCR (2-6-15 Hr-3, 2-6-15 Hr-4) (5′ RACE Kit, Invitrogen). The sequence of the 3′ end was determined by 3′ RACE (Invitrogen) using gene specific primers 2-6-15 Hf-3 for generating cDNA as well as 2-6-15 Hf-2 and 2-6-15 Hf-1 for PCR. To circumvent the expression of aberrant light chain by hybridoma 898H2-6-15, saturating levels of a primer specific for the aberrant chain (Ab Vk 5′) along with the VL primers UMIgVK 5′ and MK-3Seq 3′ were used to amplify light chains.14 As previously, 5′ sequence was determined by 5′ RACE using cDNA generated with gene specific primers 2-6-15 Lr-1; 5′RACE PCR was performed with 2-6-15 Lr-2 and 2-6-15 Lr-3 (5′ RACE Kit, Invitrogen). The sequence of the 3′ end was determined by generating cDNA with gene specific primer 2-6-15 Lf-3 and 3′RACE PCR with 2-6-15 Lf-2 and 2-6-15 Lf-1 (3′ RACE Kit, Invitrogen). All of the PCR primers used to amplify the VL and VH are listed in Table 1. The VL and VH DNA and derived amino acid sequences are shown in Fig. 1 (VL GenBank accession: JF918977; VH GenBank accession: JF918978). The borders of 898H2-6-15 VL and VH were determined according to Kabat et al (1987).18
Table 1.
PCR primers used for 2-6-15 VL and VH amplification
| MuIgVH5′-A: | GGGAATTCATGAACTTGTGGTTAAGCTGGGTTT |
| MuIgGVH3′-2: | CCCAAGCTTCCAGGGGCCAGGGGATAGACGGGTGG |
| 2-6-15 Hr-1: | TGTTGTTTTGGCTGAGGAGA |
| 2-6-15 Hr-3: | TCCATATAGT CCCCCCGTCTT |
| 2-6-15 Hr-4: | TGTCCTCAGACCTCAGACTG |
| 2-6-15 Hf-3: | GAGGCTTAGTGAAGCCTGGA |
| 2-6-15 Hf-2: | TATACCATGTCTTGGGTTCGC |
| 2-6-15 Hf-1: | TGGAGTGGGTCGCATACATTA |
| Ab Vk 5′: | ACCTATTACTGTCAGCACATTA |
| UMIgVK 5′: | GACATTCTGATGACCCAGTCT |
| MK-3Seq 3′: | TACAGTTGGTGCAGCATCAGC |
| 2-6-15 Lr-1: | TTTCAGCTCCAGCTTGGTC |
| 2-6-15 Lr-2: | TGAGCGGCAAGTTATCACTT |
| 2-6-15 Lr-3: | AAGGAGCTTAGGAGGTTTCCC |
| 2-6-15 Lf-3: | CAGATGCATAACCAGCACTGA |
| 2-6-15 Lf-2: | GCTCCTTATTTCAGAAGGCAA |
| 2-6-15 Lf-1: | ATTCTCCAGCAGTGGCTATGG |
Fig. 1.

Anti-porcine CD3 monoclonal antibody 898H2-6-15 variable light (VL) and variable heavy (VH) domain DNA and derived amino acid sequence.
Plasmid construction
Codon optimization is necessary to express DT390-based immunotoxins in Pichia pastoris.8 We used the optimized DT390 nucleotide sequence described by Woo et al 2002 for the DT390 domain in the new anti-porcine CD3 immunotoxin. The DT390 has been modified to include an NH2 terminal alanine (A) and double mutations (dm) to prevent glycosylation in the eukaryotic expression system, Pichia pastoris.8,15 The anti-porcine CD3 immunotoxin A-dmDT390biscFv (2-6-15) was constructed using the scFv derived from the anti-porcine CD3 monoclonal antibody 898H2-6-15 following the strategy used to construct A-dm-DT390biscFv (UCHT1) in Pichia pastoris8 except for the addition of an extra G4S linker between A-dm-DT390 and biscFv (2-6-15) (Fig. 2). The codon-optimized scFv (2-6-15) DNA was synthesized using the Pichia pastoris preferred codons.16 Ten primers were designed for both the VL and the VH of the 2-6-15 scFv (Table 2). The primer length was limited to under 70 bases with a 21 base overlap between neighboring primers. Ten pmol of the first and the last primers, and two pmol of the rest of the primers were used in each PCR reaction (Pfu Turbo polymerase, Aglient, dNTP mixture at 2.5 mM each). The PCR program was 95°C for 5 min, 25 cycles of 95°C for 30 sec, 55°C for 30 sec, 72°C for 1 min and then extension for another 10 min. Amplification products were analyzed by electrophoresis in 1% agarose gels and purified using QIAquik Gel Extraction Kit (Qiagen). BamHI digested VL PCR fragment was ligated to Bgl II digested VH fragment using T4 ligase for 6 hr. The first scFv (2-6-15), amplified with primers 15L1a (carrying NcoI site) and 15 H10a (carrying BamHI site), was cloned into pET27b. The second scFv (2-6-15), amplified with 15L1b (carrying BglII site) and 15H10b (carrying stop codon and EcoRI site), was cloned into pET17b. After sequence confirmation, the first and second scFv (2-6-15) were subcloned into pwPICZα-DT390 (NcoI-BamHI/BglII-stop codon-EcoRI)8.
Fig. 2.
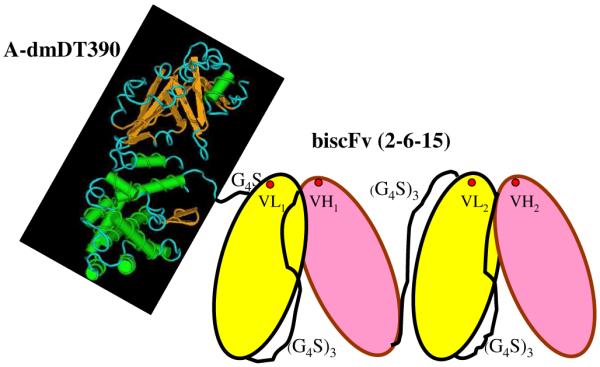
Schematic description of the anti-porcine CD3 recombinant immunotoxin A-dmDT390biscFv(2-6-15). The putative antigen binding sites are shown as red spots.
Table 2.
PCR primers used to synthesize the codon-optimized scFv (2-6-15)
| For 2-6-15 VL domain: |
| 15L1a 5′ C ATG CCA TGG GGT GGT GGT GGT TCT GAG ACT ACT GTT ACT CAA TCT CCA GCT TCT TTG 3′ |
| 15L1b 5′ GGA AGA TCT GGT GGT GGT GGT TCT GAG ACT ACT GTT ACT CAA TCT CCA GCT TCT TTG 3′ |
| 15L2 5′ ACA TCT AAT AGT AAC TTG CTC ACC AAT AGC CAT AGA CAA AGA AGC TGG AGA TTG AGT 3′ |
| 15L3 5′ GAG CAA GTT ACT ATT AGA TGT ATT ACT TCT ACT GAC ATT GAC GAC GAC ATG AAC 3′ |
| 15L4 5′ CTT TGG TGG CTT ACC TGG TCT TTG TTG GTA CCA GTT CAT GTC GTC GTC AAT GTC 3′ |
| 15L5 5′ AGA CCA GGT AAG CCA CCA AAG TTG TTG ATT TCT GAG GGT AAC ACT TTG AGA CCA 3′ |
| 15L6 5′ GTA ACC AGA AGA AGA GAA TCT AGA TGG AAC ACC TGG TCT CAA AGT GTT ACC CTC 3′ |
| 15L7 5′ AGA TTC TCT TCT TCT GGT TAC GGT ACT GAC TTC GTT TTC ACT ATT GAG AAC ATG 3′ |
| 15L8 5′ CAA ACA GTA GTA GTC AGC AAC GTC CTC AGA CAA CAT GTT CTC AAT AGT GAA AAC 3′ |
| 15L9 5′ GTT GCT GAC TAC TAC TGT TTG CAA TCT GAC AAC TTG CCA TTG ACT TTC GGT GCT 3′ |
| 15L10 5′ CGC GGA TCC ACC ACC ACC AGA ACC ACC ACC ACC CTT CAA CTC CAA CTT AGT ACC AGC ACC GAA AGT CAA TGG CAA 3′ |
| For 2-6-15 VH domain: |
| 15H1 5′ GGA AGA TCT GGT GGT GGT GGT TCT GAG GTT AAG TTG GTT GAG TCT GGT GGT GGT TTG GTT AAG CCA GGT GGT TCT 3′ |
| 15H2 5′ AGA GAA AGT GAA ACC AGA AGC AGC ACA AGA CAA CTT CAA AGA ACC ACC TGG CTT AAC CAA 3′ |
| 15H3 5′ GCT TCT GGT TTC ACT TTC TCT TCT TAC ACT ATG TCT TGG GTT AGA CAA ACT CCA GAG AAG 3′ |
| 15H4 5′ AGA ACC ACC AGA AGA AAT GTA AGC AAC CCA CTC CAA TCT CTT CTC TGG AGT TTG TCT AAC 3′ |
| 15H5 5′ TAC ATT TCT TCT GGT GGT TCT AAC TCT TAC TAC TCT GAC TCT GTT AAG GGT AGA TTC 3′ |
| 15H6 5′ GTA CAA AGT GTT TCT AGC GTT GTC TCT AGA AAT AGT GAA TCT ACC CTT AAC AGA GTC 3′ |
| 15H7 5′ AAC GCT AGA AAC ACT TTG TAC TTG CAA ATG TCT TCT TTG AGA TCC GAG GAC ACT GCT 3′ |
| 15H8 5′ GTC CAT GTA GTC ACC TCT TCT AGC ACA GTA GTA CAT AGC AGT GTC CTC AGA TCT CAA 3′ |
| 15H9 5′ AGA AGA GGT GAC TAC ATG GAC TAC TGG GGT CAA GGT ACT TCT GTT ACT GTT TCT TCT 3′ |
| 15H10a 5′ CGC GGA TCC ACC ACC ACC AGA ACC ACC ACC ACC AGA AGA AAC AGT AAC AGA AGT 3′ |
| 15H10b 5′ CCG GAA TTC TTA AGA AGA AAC AGT AAC AGA AGT 3′ |
Protein expression and purification in Pichia pastoris
The linearized anti-porcine CD3 immunotoxin construct, A-dmDT390-biscFv (2-6-15) in pwPICZα, was transformed into a diphtheria-toxin resistant yeast Pichia pastoris strain mutEF2JC307-8(2) 17 and the transformants were selected on YPD plates containing zeocin (100 μg/ml). Six colonies were randomly picked and cultivated in small tubes containing 5 mL YPD (1% yeast extract, 2% peptone and 2% dextrose) at 30°C at 250 rpm for 24h as growth phase I, then in YPG (1% yeast extract, 2% peptone, 1% glycerol) at 30°C at 250 rpm for another 24h as growth phase II. The cultures were induced with methanol in 2 mL BMMYC (1% yeast extract, 2% peptone, 100 mM potassium phosphate, pH 7.0, 1.34% yeast nitrogen base without amino acids, 4 × 10−5 % biotin, 0.5% methanol and 1% casamino acids) for 48h at 25°C at 225 rpm. Antifoam 0.02% (Emerald Performance Materials, Cat# KFO673) was added in all of the growth and induction medium. 1mM PMSF (Phenylmethanesulfonyl fluoride, Sigma) was added with methanol to inhibit the protein degradation during the induction phase. The culture supernatants were analyzed on SDS-PAGE under non-reducing condition. One clone (ZW130) was selected and cultivated to purify A-dmDT390-biscFv (2-6-15). Sodium sulfate (Fisher Scientific) was added to 1 L of culture supernatant to a final concentration of 200 mM. The sample was loaded onto 50 mL Butyl 650M (Tosoh) in a 5 cm × 20 cm XK50 column (GE healthcare Cat#18-1000-71). The bound A-dmDT390-biscFv (2-6-15) was eluted with TGE buffer (5% glycerol, 20 mM Tris HCl, pH 8.0, 1 mM EDTA, pH 8.0). Pooled fractions (Fig. 3A, Elution fraction #2 to #8, 50 mL per fraction) containing A-dmDT390-biscFv (2-6-15) were diluted six fold with TE (20 mM Tris-HCl pH 8.0, 1 mM EDTA pH 8.0) buffer and loaded directly onto 10 mL of Poros 50HQ (Applied Biosystems) in a 1.6 cm × 20 cm XK16 column (GE healthcare, Cat#18-8773-01). The bound A-dm-DT390-biscFv (2-6-15) was eluted with 150 mM sodium borate in TGE buffer and 200 mM sodium borate + 50 mM NaCl in TGE buffer. The immunotoxin containing elution fractions were mixed, concentrated down with Centricon Plus-70 (30 kDa cut off, Millipore), dialyzed against PBS pH 7.4 + 5% glycerol, filter sterilized and stored at 80°C.
Fig. 3.

Two step purification of the anti-porcine CD3 immunotoxin A-dmDT390biscFv (2-6-15). (A) Capturing step with hydrophobic interaction resin Butyl 650 M. (B) Second step purification with strong anion exchange resin Poros 50HQ.
HPLC analysis
A-dm-DT390biscFv (2-6-15) final protein product was analyzed with Shimadzu HPLC system using Superdex 200 size-exclusion column, 10/300 GL (GE healthcare, Cat#: 17-5175-01). The sample volume was 100 μl using 100 μl loop. The flow rate was 0.35 ml / min. The running time was 120 min and the running buffer was 90 mM NaSO4, 10 mM NaPO4, pH 8.0, 1 mM EDTA.
Porcine CD3 depletion study in vivo
Four miniature swine, aged two months, were brought to our AAALAC accredited animal facility and were allowed to acclimate to their new surroundings for one week. Three days prior to the start of treatment they were bled for baseline blood values: complete blood counts, serum chemistry values and for flow cytometry analysis of CD3+ T cell populations. They were then injected through a 23g butterfly catheter, through a peripheral vessel, with an intravenous (IV) bolus of undiluted A-dmDT390biscFv (2-6-15) at a dose of 50 μg/kg, twice daily for 4 days. Prior to each day’s injections, morning blood samples were taken to monitor the aforementioned blood values. Each day’s values were then compared to the pre-treatment values. The animals were also injected with 4mg/kg diphenhydramine IV bolus immediately prior to immunotoxin infusion to prevent any unexpected anaphylactic reactions, of which none were observed. To ensure the full volume of immunotoxin was administered to the animals, each dose was flushed with 10 mL of phosphate buffered saline (pH of between 6.5 and 7.5). The animals were closely monitored for signs of change in clinical condition such as lethargy and significant weight loss and no signs of toxicity were seen in any animal. The animals were bled again on days 7 and 14 following the start of injections to monitor their T cell recovery and any possible toxicity.
The percentage of CD3+ T cells in the peripheral blood was determined by flow cytometry of heparinized whole blood samples after staining with FITC conjugated swine specific CD3 antibody (898H2-6-15, mouse IgGaK)13 using BD FACS lysing solution whole blood staining procedure according to the manufacturer (BD Biosciences, San Jose, Ca). The absolute T-cell count each day was calculated by multiplying the percentage of CD3+ cells in the peripheral blood by the white blood cell count. All experiments were approved by the Massachusetts General Hospital Institutional Animal Care and Use Committee.
RESULTS AND DISCUSSION
Plasmid construction for the anti-porcine CD3 recombinant immunotoxin
The VL and VH DNA sequence and the deduced amino acid sequences of anti-porcine CD3 mAb 898H2-6-15 are shown in Fig. 1. According to Sreekrishna 1993,16 we synthesized the codon-optimized scFv (2-6-15) and cloned it into the truncated diphtheria toxin DT390 containing yeast Pichia pastoris expression vector pwPICZα-A-dmDT390 between NcoI and EcoRI to replace the biscFv (UCHT1) portion.8 Initially we utilized the single chain fold back diabody format as in the anti-monkey CD3 immunotoxin construct7 but treatment with this recombinant immunotoxin did not result in depletion of porcine CD3+ T cells in vivo (data not shown). We speculate that this could be because the single chain foldback diabody format is not suitable for binding the porcine CD3 epitope. Even though both 898H2-6-15 and FN18 recognize CD3 epsilon, it is likely that they bind different conformational CD3 epitopes. The traditional biscFv format, which was used in the anti-human CD3 recombinant immunotoxin (Fig. 2),8 was constructed. The bivalent immunotoxin is a multi-domain protein containing the catalytic and translocation domains of DT and two VL and two VH of the anti-porcine CD3 mAb 898H2-6-15. The VL and VH are joined by a 15 amino acid (G4S)3 linker (L) in the order of VL-L-VH-L-VL-L-VH.
Expression and purification of the anti-porcine CD3 recombinant immunotoxin
The anti-porcine CD3 recombinant immunotoxin A-dmDT390biscFv (2-6-15) was expressed using shaker flasks by scaling-up from small tube expression as described in the Experimental Procedures. The scaled up expression level is about 20 mg / L. The secreted anti-porcine CD3 immunotoxin in the supernatant was captured directly by hydrophobic interaction chromatography resin Butyl 650M (Fig. 3A). For second step purification a strong anion exchange resin Poros 50 HQ was used. Sodium chloride alone did not work well for eluting the immunotoxin as the glycosylated yeast host protein was co-eluted with the immunotoxin19. Sodium borate was chosen as it would separate the immunotoxin from the glycosylated yeast host protein and the aggregates. However the maximal solubility for sodium borate is restricted to 200 mM, a concentration which is not strong enough to elute the majority of the immunotoxin. To overcome this problem, we added 50 mM NaCl to the 200 mM sodium borate elution buffer to increase the salt concentration. The majority of the immunotoxin was eluted with 200 mM sodium borate and 50 mM NaCl (Fig. 3B). As shown in Fig. 4, after two step purification we obtained the pure 95 kDa anti-porcine CD3 recombinant immunotoxin. The purity reached around 95%. The final purification yield was ~13 mg per liter of the original harvested supernatant. As shown in Fig. 4A, A-dmDT390biscFv (2-6-15) showed a second weak band migrating around 70 kDa. Western blots identified it as breakdown product.
Fig. 4.

Anti-porcine CD3 immunotoxin A-dmDT390biscFv (2-6-15) analysis. (A) NuPAGE 4-12% Bis-Tris gel analysis. (B) HPLC analysis.
Porcine CD3 T cell depletion in vivo using the anti-porcine CD3 recombinant immunotoxin
The CD3 T cell depletion function of this recombinant immunotoxin was assessed in four MGH miniature swine, aged two months. Results are shown in Fig. 5. This recombinant immunotoxin was given through IV bolus at a dose of 50 g/kg, twice daily for four days. Complete blood counts, serum chemistry values and CD3+ T cell numbers were measured. The immunotoxin treatment decreased the number of CD3+ T cells in the peripheral blood to approximately 20% of the pretreatment level (Fig. 5A). There was no significant effect on the number of peripheral B cells (Fig. 5B). The percent CD3+ T cells in the lymph node decreased from 74% pretreatment to 24% following 4 days of immunotoxin treatment reversing the ratio of T:B cells in the lymph node (Fig. 5C).
Fig. 5.

Porcine CD3 T cell depletion profile in vivo for four animals using anti-porcine CD3 immunotoxin, A-dmDT390biscFv (2-6-15). A) Absolute number of CD3+ T cells in the peripheral blood. B) Absolute number of B cells in the peripheral blood. C) Lymph node percent CD3+ T cells.
Clinically, no toxicities were observed in any of the treated animals. No anaphylactic reactions were observed. Kidney toxicity was not observed and renal values (blood urea nitrogen and creatinine) remained within normal limits. Two animals 18316 and 19586 developed mild increases of liver enzymes from baseline pre-immunotoxin. Lactate dehydrogenase levels for animal 18316 increased from 1166 u/L to 1960u/L and for animal 19586 from 1722 u/L to 2017 u/L. Alkaline phosphatase levels for animal 18316 increased from 102 u/L to 204 u/L. No changes were observed in any of the other animals. Despite slight increases from baseline lactate dehydrogenase and alkaline phosphatase levels, these enzymes remained well within the normal limits in swine (575-3200 u/L for lactate dehydrogenase and 92-295 u/L for alkaline phosphatase). The transaminase remained normal and histopathology was unremarkable. In summary, we did not observe any side effect after the imunotoxin treatment. At the dose tested, the anti-porcine CD3 recombinant immunotoxin did not deplete CD3 T cells as well as its counterpart anti-human or anti-monkey CD3 recombinant immunotoxins.7,12 The dose used in swine was at least two times higher than the dose of its counterpart anti-human or anti-monkey CD3 recombinant immunotoxins.7,12 It is possible that miniature swine are more tolerant to the recombinant immunotoxin thereby allowing even further increased dosing for improved depletion effect. Given the absence of any signs of toxicity, we speculate that it is possible to increase the current daily dose or extend the administration days. A dosing study will need to be done to find the maximal safe dose and best dosing strategy for improved depletion. Since the half-life of recombinant anti-human CD3 immunotoxin is only 40 min12, it may not be necessary to spread 8 doses in 4 days. It is also possible that increasing the affinity of the scFv (2-6-15) similar to the affinity maturation approach used for the anti-monkey CD3 recombinant immunotoxin may improve depletion function. Affinity maturation of the scFv (2-6-15) by yeast display20 is currently in progress to isolate a higher affinity scFv (2-6-15) for improved porcine T cell depletion in vivo.
Although CD3+ T cell depletion following anti-porcine CD3 recombinant immunotoxin treatment was not complete, in combination with other treatments, this reagent may provide sufficient T cell depletion for certain applications.
Compared with the corresponding chemical conjugate anti-porcine CD3 immunotoxin (pCD3-CRM9), this recombinant anti-porcine CD3 immunotoxin [A-dmDT390biscFv(2-6-15)] has the following advantages: 1) high production level suitable for scale-up; 2) high purity and linkage homogeneity; 3) no neurological toxicity. In contrast, the chemical conjugate anti-porcine CD3 immunotoxin caused significant dose-dependent neurotoxicity.6 These finding suggest that the mutated DT binding domain of the chemical conjugate anti-porcine CD3 immunotoxin can still bind to the porcine DT receptor. The recombinant anti-porcine CD3 immunotoxin does not contain the DT binding domain (Fig. 2). As expected we did not observe any neurotoxicity in any of the animals treated with recombinant anti-porcine CD3 immunotoxin. A-dmDT390biscFv (2-6-15) is a unique recombinant protein reagent that can be utilized in experimental porcine models of transplantation tolerance, autoimmune disease therapy, T-cell leukemia treatment as well as graft versus host disease studies.
ACKNOWLEDGEMENTS
The work was supported by National Institutes of Health (R01AI084657-02 to CAH) and Dana Farber/Harvard Cancer Center Core development grant. We thank Christina Hermanrud, Prashanth Vallabhajosyula and Lauren L. Springett for their excellent technical assistance; Bob Hawley and Sharon Germana for intensive manuscript review.
REFERENCES
- (1).Huang CA, Yamada K, Murphy MC, Shimizu A, Colvin RB, Neville DM, Jr., Sachs DH. In vivo T cell depletion in miniature swine using the swine CD3 immunotoxin, pCD3-CRM9. Transplantation. 1999;68:855–860. doi: 10.1097/00007890-199909270-00019. [DOI] [PubMed] [Google Scholar]
- (2).Fuchimoto Y, Huang CA, Yamada K, Shimizu A, Kitamura H, Colvin RB, Ferrara V, Murphy MC, Sykes M, White-Scharf M, Neville DM, Jr., Sachs DH. Mixed chimerism and tolerance without whole body irradiation in a large animal model. J. Clin. Invest. 2000;105:1779–1789. doi: 10.1172/JCI8721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (3).Huang CA, Fuchimoto Y, Scheier-Dolberg R, Murphy MC, Neville DM, Jr., Sachs DH. Stable mixed chimerism and tolerance using a nonmyeloablative preparative regimen in a large-animal model. J. Clin. Invest. 2000;105:173–181. doi: 10.1172/JCI7913. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (4).Cina RA, Wikiel KJ, Lee PW, Cameron AM, Hettiarachy S, Rowland H, Goodrich J, Colby C, Spitzer TR, Neville DM, Jr., Huang CA. Stable Multilineage Chimerism without Graft versus Host Disease Following Nonmyeloablative Haploidentical Hematopoietic Cell Transplantation. Transplantation. 2006;81:1677–1685. doi: 10.1097/01.tp.0000226061.59196.84. [DOI] [PubMed] [Google Scholar]
- (5).Neville DM, Jr., Scharff J, Hu HZ, Rigaut K, Shiloach J, Slingerland W, Jonker M. A new reagent for the induction of T-cell depletion, anti-CD3-CRM9. J. Immunother. Emphasis. Tumor Immunol. 1996;19:85–92. doi: 10.1097/00002371-199603000-00001. [DOI] [PubMed] [Google Scholar]
- (6).Gargollo P, Yamada K, Esnaola N, Fuchimoto Y, Newell KL, Sachs DH, Huang CA. Neuropathy in miniature swine following administration of the mutant diphtheria toxin based immunotoxin, pCD3-CRM9. Transplantation. 2001;72:818–822. doi: 10.1097/00007890-200109150-00013. [DOI] [PubMed] [Google Scholar]
- (7).Kim GB, Wang Z, Liu YY, Stavrou S, Mathias A, Goodwin KJ, Thomas JM, Neville DM., Jr. A fold-back single-chain diabody format enhances the bioactivity of an anti-monkey CD3 recombinant diphtheria toxin-based immunotoxin. Protein Eng. Des. Sel. 2007;20:425–432. doi: 10.1093/protein/gzm040. [DOI] [PubMed] [Google Scholar]
- (8).Woo JH, Liu YY, Mathias A, Stavrou S, Wang Z, Thompson J, Neville DM., Jr. Gene optimization is necessary to express a bivalent anti-human anti-T cell immunotoxin in Pichia pastoris. Protein Expr. Purif. 2002;25:270–282. doi: 10.1016/s1046-5928(02)00009-8. [DOI] [PubMed] [Google Scholar]
- (9).Madshus IH, Olsnes S, Stenmark H. Membrane translocation of diphtheria toxin carrying passenger protein domains. Infect. Immun. 1992;60:3296–3302. doi: 10.1128/iai.60.8.3296-3302.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (10).Hexham JM, King V, Dudas D, Graff P, Mahnke M, Wang YK, Goetschy JF, Plattner D, Zurini M, Bitsch F, Lake P, Digan ME. Optimization of the anti-(human CD3) immunotoxin DT389-scFv(UCHT1) N-terminal sequence to yield a homogeneous protein. Biotechnol. Appl.Biochem. 2001;34:183–187. doi: 10.1042/ba20010073. [DOI] [PubMed] [Google Scholar]
- (11).Thompson J, Stavrou S, Weetall M, Hexham JM, Digan ME, Wang Z, Woo JH, Yu Y, Mathias A, Liu YY, Ma S, Gordienko I, Lake P, Neville DM., Jr. Improved binding of a bivalent single-chain immunotoxin results in increased efficacy for in vivo T-cell depletion. Protein Eng. 2001;14:1035–1041. doi: 10.1093/protein/14.12.1035. [DOI] [PubMed] [Google Scholar]
- (12).Frankel AE, Zuckero SL, Mankin AA, Grable M, Mitchell K, Lee YJ, Neville DM, Jr., Woo JH. Anti-CD3 recombinant diphtheria immunotoxin therapy of cutaneous T cell lymphoma. Curr. Drug. Targets. 2009;10:104–109. doi: 10.2174/138945009787354539. [DOI] [PubMed] [Google Scholar]
- (13).Huang CA, Lorf T, Arn JS, Koo GC, Blake T, Sachs DH. Characterization of a monoclonal anti-porcine CD3 antibody. Xenotransplant. 1999;5:201–212. doi: 10.1034/j.1399-3089.1999.00022.x. [DOI] [PubMed] [Google Scholar]
- (14).Yuan X, Gubbins MJ, Berry JD. A simple and rapid protocol for the sequence determination of functional kappa light chain cDNAs from aberrant-chain-positive murine hybridomas. J. Immunol. Methods. 2004;294:199–207. doi: 10.1016/j.jim.2004.09.001. [DOI] [PubMed] [Google Scholar]
- (15).Liu YY, Gordienko I, Mathias A, Ma A, Thompson J, Woo JH, Neville DM., Jr. Expression of an anti-CD3 single-chain immunotoxin with a truncated diphtheria toxin in a mutant CHO cell line. Protein Expr. Purif. 2000;19:304–311. doi: 10.1006/prep.2000.1255. [DOI] [PubMed] [Google Scholar]
- (16).Sreekrishna K. Strategies for optimizing protein expression and secretion in the methylotrophic yeast Pichia pastoris. In: Baltz RH, Hegeman GD, Skatrud PL, editors. Industrial Microorganism: Basic and Applied Molecular Genetics. American Society of Microbiology; Washington, DC: 1993. pp. 119–126. [Google Scholar]
- (17).Liu YY, Woo JH, Neville DM., Jr. Targeted introduction of a diphtheria toxin resistant mutation into the chromosomal EF-2 locus of Pichia pastoris and expression of immunotoxin in the EF-2 mutants. Protein Expr. Purif. 2003;30:262–274. doi: 10.1016/s1046-5928(03)00129-3. [DOI] [PubMed] [Google Scholar]
- (18).Kabat EA, Wu TT, Reid-Miller M, Gottesman KS. US Department of Health and Human Services. US Governmental Printing Office; Washington, DC: 1987. Sequences of proteins of immunological interest. [Google Scholar]
- (19).Woo JH, Neville DM., Jr. Separation of bivalent anti-T cell immunotoxin from Pichia pastoris glycoproteins by borate anion exchange. BioTechniques. 2003;35:392–398. doi: 10.2144/03352pt04. [DOI] [PubMed] [Google Scholar]
- (20).Wang Z, Kim GB, Woo JH, Liu YY, Mathias A, Stavrou S, Neville DM., Jr. Improvement of a recombinant anti-monkey anti-CD3 diphtheria toxin based immunotoxin by yeast display affinity maturation of the scFv. Bioconjug.Chem. 2007;18:947–955. doi: 10.1021/bc0603438. [DOI] [PubMed] [Google Scholar]