

Abstract
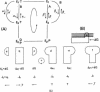
The basic kinetic and bioenergetic theory is outlined for two kinds of translocation on DNA: (i) helicases that use ATP to move along single-stranded DNA or to move on and invade double-stranded DNA at a replication fork; and (ii) DNA-binding proteins (not ATPases) that form bound aggregates on single-stranded DNA and facilitate replication by steady-state treadmilling of molecules between the ends of the aggregate. The respective resemblances to myosin--actin in muscle and to steady-state treadmilling in solution of actin or tubulin are pointed out.
Full text
PDF





Images in this article

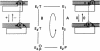
Selected References
These references are in PubMed. This may not be the complete list of references from this article.
- Bergen L. G., Borisy G. G. Head-to-tail polymerization of microtubules in vitro. Electron microscope analysis of seeded assembly. J Cell Biol. 1980 Jan;84(1):141–150. doi: 10.1083/jcb.84.1.141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Das R. H., Yarranton G. T., Gefter M. L. Enzyme-catalyzed DNA unwinding. The role of ATP in helicase III activity. J Biol Chem. 1980 Sep 10;255(17):8069–8073. [PubMed] [Google Scholar]
- Eisenberg E., Hill T. L., Chen Y. Cross-bridge model of muscle contraction. Quantitative analysis. Biophys J. 1980 Feb;29(2):195–227. doi: 10.1016/S0006-3495(80)85126-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hill T. L. Bioenergetic aspects and polymer length distribution in steady-state head-to-tail polymerization of actin or microtubules. Proc Natl Acad Sci U S A. 1980 Aug;77(8):4803–4807. doi: 10.1073/pnas.77.8.4803. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hill T. L. Steady-state head-to-tail polymerization of actin or microtubules. II. Two-state and three-state kinetic cycles. Biophys J. 1981 Mar;33(3):353–371. doi: 10.1016/S0006-3495(81)84900-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huxley A. F., Simmons R. M. Proposed mechanism of force generation in striated muscle. Nature. 1971 Oct 22;233(5321):533–538. doi: 10.1038/233533a0. [DOI] [PubMed] [Google Scholar]
- Kirschner M. W. Implications of treadmilling for the stability and polarity of actin and tubulin polymers in vivo. J Cell Biol. 1980 Jul;86(1):330–334. doi: 10.1083/jcb.86.1.330. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Margolis R. L., Wilson L. Addition of colchicine--tubulin complex to microtubule ends: the mechanism of substoichiometric colchicine poisoning. Proc Natl Acad Sci U S A. 1977 Aug;74(8):3466–3470. doi: 10.1073/pnas.74.8.3466. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wegner A. Head to tail polymerization of actin. J Mol Biol. 1976 Nov;108(1):139–150. doi: 10.1016/s0022-2836(76)80100-3. [DOI] [PubMed] [Google Scholar]
- Yarranton G. T., Gefter M. L. Enzyme-catalyzed DNA unwinding: studies on Escherichia coli rep protein. Proc Natl Acad Sci U S A. 1979 Apr;76(4):1658–1662. doi: 10.1073/pnas.76.4.1658. [DOI] [PMC free article] [PubMed] [Google Scholar]