

Abstract
Taurine is an abundant amino acid in the brain that shares pharmacological effects and similar potency with ethanol. Recently, taurine-containing beverages have been reported to enhance the euphoric effects of ethanol, though the extent of this effect and the role of taurine remain speculative. The present study was designed to explore interactions between taurine and ethanol on several behaviors including locomotion, ataxia, and loss of righting. Two strains of mice, C57BL/6J and DBA/2J mice, were used to examine potential strain differences. In the first experiment, effects of various doses of taurine (0.3–3.0 g/kg), ethanol (1.0–4.2 g/kg), or taurine in combination with ethanol were assessed in a within-subjects design. Although taurine did not appear to alter effects of ethanol on any measure in either strain, the development of tolerance to locomotor effects and sensitization to ataxic effects of ethanol in DBA/2J mice complicated interpretation of these results. In a second experiment, drug-naïve mice were assigned to one of four treatment groups: saline + saline, saline + ethanol (1.78 g/kg), taurine (1.78 g/kg) + saline, or ethanol + taurine. In this experiment, taurine pretreatment significantly attenuated the locomotor-stimulating effect of ethanol in both strains (but to a greater extent in C57BL/6J mice) and appeared to reduce the ataxic effects of ethanol in C57BL/6J mice. In conclusion, the interaction between taurine and ethanol is subtle. Further, results are inconsistent with the notion that taurine plays a major role in the locomotor, ataxic, or loss of righting effects of ethanol.
Keywords: sleep-time, alcohol, mouse, motor
1. Introduction
Taurine is a sulfonated β-amino acid found in high concentrations in brain and muscle tissue (Olive, 2002). Relatively high concentrations of taurine are found in popular energy drinks and such drinks are reported to enhance the positive effects of ethanol, possibly by reducing the depressant effects of ethanol (Ferreira et al., 2004a). Yet, taurine alone appears to exert an inhibitory effect in the brain (Olive, 2002). Direct injection of taurine into the central nervous system results in decreased locomotion, which peaks at approximately 20 min (Sgaragli and Pavan, 1972; Baskin et al., 1974). Taurine is the most abundant amino acid in brain extracellular fluid and may be effective at low- to mid-micromolar concentrations, similar to ethanol (Olive, 2002). Acute ethanol administration (1–3 g/kg) increases extracellular taurine levels 50–100% in mouse and rat brain (Olive, 2002). Because taurine and ethanol appear to share some pharmacological effects, and acute ethanol administration increases extracellular taurine levels, taurine may play a role in the behavioral effects of ethanol.
That C57BL/6J and DBA/2J strains of mice display strain-dependent responses to ethanol is well-established. Moderate doses of ethanol (1–2 g/kg) increase open-field locomotion persistently in DBA/2J mice, but only transiently in C57BL/6J mice (Crabbe et al., 1982). These doses produce ataxia in both strains. Higher doses of ethanol decrease open-field locomotion and result in loss of the righting reflex in both strains. The stimulant and ataxic effects of ethanol have been related to reinforcing effects, while depressant effects have been related to aversive effects of higher ethanol doses (Wise and Bozarth, 1987).
Low doses of taurine (30 and 45 mg/kg) have been reported to inhibit locomotor stimulation following a 1.0 g/kg dose of ethanol in Swiss-Webster mice (Aragon et al., 1992). However, similar doses of taurine (30, 45, and 60 mg/kg) enhance the stimulant effect of 2.0 g/kg ethanol. Several studies have reported that taurine (30–80 mg/kg) reduces the duration of ethanol-induced loss of righting reflex, however a putative taurine antagonist does not alter this effect (Iida and Hikichi, 1976; Boggan et al., 1978; Ferko, 1987). Currently, there are no reported studies of ataxic effects of ethanol-taurine interactions.
The purpose of the present study was to extend the characterization of ethanol-taurine interactions by assessing locomotion, ataxia, and loss of righting reflex in C57BL/6J and DBA/2J mice. Because of the well-documented differences in ethanol effects between these two strains (Crabbe, et al., 1982), we hypothesized that interactions between ethanol and taurine would show similar strain-differences.
2. Materials and Methods
2.1 General Methods
The behavioral effects of taurine and ethanol on open field locomotion, ataxia on an inverted screen, and loss of righting reflex were assessed. Following the drug treatments described below, mice were placed in a locomotor activity chamber for the time described. Immediately after completing locomotor measurements, mice were removed from the chamber and placed on the benchtop for determination of loss of righting reflex and ataxic effects, in that order.
2.1.1 Locomotor activity
Locomotion was assessed using four 30 cm × 15 cm × 15 cm customized acrylic boxes with floors of a parallel grid of 2.3 mm stainless steel rods mounted 6.4 mm apart (Instrumentation Services, University of Texas Health Science Center at San Antonio). Each box was enclosed in a commercially-available sound- and light-attenuating chamber (Model #ENV-022M; MED Associates Inc., St. Albans, VT). Four infrared light beams were evenly spaced 6 cm apart and located 2 cm from the floor of each box. Disruptions of the light beams were counted using commercially available computer software (Multi-Varimex v1.00, Colombus Instruments, Colombus, OH). The floors and inside of the boxes were wiped with a damp sponge and the litter paper beneath the floors was changed between tests with different animals.
2.1.2 Loss of Righting Reflex
Each subject was placed on his back on a clean paper towel on the benchtop. Subjects failing to place all four feet on the benchtop within 20-sec were scored as having lost the righting reflex.
2.1.3 Ataxia
The inverted screen apparatus (Instrumentation Services, University of Texas Health Science Center at San Antonio) consisted of a 13 cm × 13 cm wire mesh screens (No. 4 mesh) located 23 cm above the floor of four Plexiglas containers. Screens were connected to a rod and handle, which could be rotated 180° to invert the screen. Subjects failing to remain on the inverted wire mesh for at least 60-sec were scored as having ataxia. Subjects displaying a loss of righting reflex were scored as ataxic, but not tested.
2.1.4 Statistics
Analyses were performed using R for Mac OS X Aqua GUI written by Stefano M. Iacus and Simon Urbanek, The R Foundation for Statistical Computing, http://www.R-project.org. Graphs were produced with the same software.
2.1.5 Drugs
Taurine (Sigma, St. Louis, MO) and ethanol (Aaper, Shelbyville, KY) were dissolved in 0.9% saline. Taurine was dissolved at doses of 0.56, 1, and 1.78 g/ml. Doses were administered at 1 ml/kg, with the exception of 3 g/kg which was administered at 1.68 ml/kg of 1.78 g/ml solution, due to insolubility at 3 g/ml. Ethanol was dissolved at a concentration of 20% v/v and the volume administered was adjusted according to the mouse body weight to provide doses of 1.0, 1.78, 3.2, and 4.2 g/kg.
2.2 Experiment 1: Within-subject effects of taurine and ethanol
The effects of various doses of taurine and ethanol were assessed as were effects of interactions between taurine and ethanol. Dose-effect curves for were determined in each mouse in the order described below.
2.2.1 Methods
2.2.1.1 Subjects
Adult male C57BL/6J (n=12) and DBA/2J (n=12) mice (Jackson Labs, Bar Harbor, ME) served as subjects. Upon arrival, mice were 6 weeks old and weighed 20–25g. Mice were allowed to habituate to vivarium routines for two weeks after arrival. Six mice of each strain were tested on every other weekday, allowing at least one day between injections or behavioral tests.
Experimental design
2.2.1.2 Habituation
Mice were habituated to the behavioral test procedures by injecting each subject with saline (0.1 ml/g) before daily experiments. Habituation was considered to have occurred when group results from two consecutive days did not significantly differ determined with a repeated-measures two-way ANOVA with days and within-session time interval as factors. Habituation took 5 sessions (data not shown).
2.2.1.3 Taurine effects
Taurine (vehicle, 0.56, 1.0, 1.8, 3.0 g/kg, i.p.) was administered to subjects immediately before the start of locomotor testing in randomized order to counterbalance treatment order. Locomotor effects were determined over consecutive 5-min sampling intervals for 60-min, to establish onset and persistence of any observed locomotor effects.
2.2.1.4 Ethanol effects
Ethanol (vehicle, 1.0, 1.8, 3.2, and 4.2 g/kg, i.p.) was also administered to subjects immediately before locomotor testing in a randomized order to counterbalance treatment order. Locomotor effects were determined over 30-min during consecutive 5-min sampling intervals to establish onset and persistence of any observable locomotor effects.
2.2.1.5 Taurine and Ethanol Interactions
Once the time course and magnitude of taurine and ethanol effects were established, effects of interactions between taurine (0.56, 1.0, and 1.8 g/kg) and ethanol (vehicle, 1.0, 1.8, 3.2, and 4.2 g/kg) were assessed. Based on the effects of taurine and ethanol alone, taurine was injected 20-min before ethanol, and locomotor effects were assessed for 20-min immediately following ethanol injection. Although mice received taurine treatments in the same order (1.0, 1.8, 0.56 g/kg) the order of ethanol administration was counterbalanced by randomizing the dose order.
2.2.1.6 Ethanol
Alone Redetermination: To establish if the effects of ethanol alone had changed over the course of the experiment, each subject received a saline injection 20-min prior to various doses of ethanol (0, 1.0, 1.8, 3.2, and 4.2 g/kg, i.p.). Dose order was randomized within each strain. Locomotor effects were determined over consecutive 5-min sampling intervals for 30-min after ethanol injection.
2.2.2 Analysis
2.2.2.1 Locomotor effects
Taurine effects
A three-way ANOVA were performed with strain, taurine dose and time as factors. Because an interaction was present between strain and time, separate two-way ANOVAs were performed for data from each strain, with taurine dose and time as factors. Post-hoc paired t-tests were performed and adjusted for multiple comparisons with the Benjamani-Hochberg method (Benjamani and Hochberg, 1995).
Ethanol effects
Again, a three-way ANOVA were performed with strain, ethanol dose and time as factors. Because interactions was present between strain and time as well as between strain and dose, separate two-way ANOVAs were performed for data from each strain, with ethanol dose and time as factors. Post-hoc comparisons were made using the method of Benjamani-Hochberg.
Taurine/Ethanol interactions
Because the effects of taurine and ethanol alone were clearly strain-dependent, separate three-way repeated measures ANOVAs were performed with ethanol dose, taurine dose, and time as factors for each strain.
2.2.2.2 Ataxia and Loss of Righting Reflex
A multiple logistic regression was performed with taurine dose, and ethanol dose as factors for each strain on ataxia or loss of righting reflex results. These analyses were performed by using the generalized linear method for a binomial distribution (glmmPQL function in the MASS package of R). The results of these regressions were then analyzed using an analysis of variance using a Chi-squared distribution. Because of apparent changes in the response to ethanol over repeated sessions, a second analysis in each strain was performed to compare effects of ethanol alone before and after taurine pretreatment studies.
2.3. Experiment 2: Between-subjects effects of taurine and ethanol
2.3.1 Subjects
Adult male C57BL/6J (n=12) and DBA/2J (n=12) mice (Jackson Labs). Upon arrival at our institution, mice were allowed to habituate to vivarium routines for two weeks. Each week, four flights of four mice (8 C57BL/6J and 8 DBA/2J mice) were tested.
2.3.2 Experimental design
2.3.2.1 Habituation
Mice were habituated to the behavioral test procedures by injecting each with saline (0.1 ml/g) before daily experiments. Mice were placed in the locomotor activity chamber for four days following saline injection (Monday–Thursday), and tested with drug or vehicle on Friday. In each group, locomotor activity on the third and fourth habituation days did not differ as determined with a repeated measures two-way ANOVA for each group (data not shown).
2.3.2.2 Drug doses
In each flight of mice, each subject was randomly assigned to one of four treatment groups (vehicle + vehicle, 1.78 g/kg taurine + vehicle, vehicle + 1.78 g/kg ethanol, or 1.78 g/kg taurine + 1.78 g/kg ethanol). These doses were selected based on results of experiment 1. A dose of 1.78 g/kg taurine produced modest locomotor reductions in both strains, but less than 3 g/kg. The dose of ethanol selected increased locomotor activity in DBA/2J mice. Chamber placement was counterbalanced to prevent a location bias.
2.3.2.3 Analysis
2.3.3 Locomotor effects
A three-way mixed ANOVA were performed with strain and treatment condition as between-subject factors and time as a within-subject factor. Because an interaction was present between strain and treatment condition, separate two-way mixed ANOVAs were performed for data from each strain, with treatment condition and time as factors. Post-hoc paired t-tests were performed and adjusted for multiple comparisons with the Benjamani-Hochberg method.
2.3.4 Ataxic effects
For each strain, a one-tailed Fisher test for binomial, independent samples was performed between selected treatment groups, and p-values were adjusted according to the Benjamani-Hochberg method. A one-tailed test was selected based on our original hypothesis that taurine would reduce the ataxic effects of ethanol.
3. Results
3.1 Experiment 1: Within-subjects effects
3.1.1 Taurine
3.1.1.1 Locomotor activity
Taurine (1.78–3.0 g/kg) modestly reduced locomotion (Fig. 1). In both strains, taurine reduced locomotor behavior within 20–25 min after administration and the effect persisted for at least 20-min. A three-way ANOVA revealed a significant interaction between strain and time (F[11,292] = 6.05, P<0.001) as well as a significant main effect of strain (F[1, 292] = 178.98, P<0.001). To further investigate effects over time within each strain, separate two-way ANOVAs were performed for each strain. In both strains, effects of taurine dose and time were significant; further, an interaction between taurine dose and time was also present (see Table 1). Post-hoc analyses revealed that locomotor activity following doses of 1.78 and 3 g/kg was lower than control in both strains (P<0.05). Further, effects plateaued between 20–25 min post-injection and remained stable until at least 50–55 min post injection. Based on these results, ethanol was administered 20-min after taurine in studies of interactions.
Figure 1.

Effecs of taurine (i.p) on locomotor activity in C57BL/6J (A) and DBA/2J (B) mice. Each point represents the mean of locomotor counts over the preceeding 5-min in all 12 subjects. Points labeled SAL represent effects following saline administration. Error bars representing S.E.M. are presented for SAL points (following saline), and the 3 g/kg taurine dose. Asterisks represent points that significantly differ from the corresponding SAL point. Note that for clarity, the y-axis is broken.
Table 1.
Results of ANOVAs on locomotor effects of taurine, ethanol, or their interaction
| Treatment | Strain | F value | P value | ||
|---|---|---|---|---|---|
| Taurine | C57BL/6 | Dose x Time | F(55, 605) | 1.4 | < 0.03 |
| Dose | F(5, 55) | 16.3 | < 0.001 | ||
| Time | F(11, 121) | 42.9 | < 0.0001 | ||
| Taurine | DBA/2J | Dose x Time | F(55, 605) | 1.6 | < 0.01 |
| Dose | F(5, 55) | 4.5 | < 0.005 | ||
| Time | F(11, 121) | 89.8 | < 0.0001 | ||
| Ethanol | C57BL/6 | Dose x Time | F(20, 200) | 3.2 | < 0.0001 |
| Dose | F(4, 40) | 75.8 | < 0.0001 | ||
| Time | F(5, 50) | 8.8 | < 0.0001 | ||
| Ethanol | DBA/2J | Dose x Time | F(20, 220) | 5.0 | < 0.0001 |
| Dose | F(4, 44) | 25.6 | < 0.0001 | ||
| Time | F(5, 55) | 46.2 | < 0.0001 | ||
| Taurine x Ethanol Interactions | Values when first ethanol-alone determination is excluded | ||||
| C57BL/6 | Taurine x Ethanol x Time | F(60, 660) | 0.9 | > 0.7 | |
| Taurine x Ethanol | F(12, 120) | 1.4 | >0.1 | ||
| Taurine x Time | F(15, 150) | 0.8 | >0.6 | ||
| Ethanol x Time | F(20, 200 | 5.2 | < 0.0001 | ||
| Taurine | F(3, 30) | 2.7 | >0.06 | ||
| Ethanol | F(4, 40) | 88.4 | < 0.0001 | ||
| Time | F(5, 50) | 67.2 | < 0.0001 | ||
| DBA/2J | Taurine x Ethanol x Time | F(60, 660) | 0.9 | >0.7 | |
| Taurine x Ethanol | F(12, 132) | 0.9 | >0.5 | ||
| Taurine x Time | F(15, 165) | 0.9 | >0.5 | ||
| Ethanol x Time | F(20, 220) | 57.1 | < 0.0001 | ||
| Taurine | F(3, 33) | 7.5 | < 0.0001 | ||
| Ethanol | F(4, 44) | 98.9 | < 0.0001 | ||
| Time | F(5, 55) | 144.3 | < 0.0001 | ||
3.1.1.2 Ataxia and loss of righting reflex
As shown in Table 2, taurine did not produce ataxia or loss of righting reflex at any of the doses tested. Although up to one-third of C57BL/6J mice displayed ataxia following 1 g/kg, ataxic effects were not dose-related, and were not significant in either strain (P>0.05).
Table 2.
Field Tests for Taurine and Ethanol
| Strain | Taurine dose (g/kg) | Loss of Righting (percent with) | Ataxia (percent with) |
|---|---|---|---|
| C57 | 0.00 | 0% | 8.3% |
| 0.30 | 0% | 25.0% | |
| 0.56 | 0% | 25.0% | |
| 1.00 | 0% | 33.3% | |
| 1.78 | 0% | 16.7% | |
| 3.00 | 0% | 25.0% | |
| DBA | 0.00 | 0% | 0.0% |
| 0.30 | 0% | 0.0% | |
| 0.56 | 0% | 0.0% | |
| 1.00 | 0% | 8.3% | |
| 1.78 | 0% | 0.0% | |
| 3.00 | 0% | 0.0% | |
| Strain | Ethanol Dose (g/kg) | Loss of Righting (percent with) | Ataxia (percent with) |
| C57 | 0.00 | 0% | 0.0% |
| 1.00 | 0% | 0.0% | |
| 1.78 | 0% | 0.0% | |
| 3.20 | 27.3% | 90.9% | |
| 4.20 | 100.0% | 100.0% | |
| DBA | 0.00 | 0% | 0.0% |
| 1.00 | 0% | 0.0% | |
| 1.78 | 0% | 9.1% | |
| 3.20 | 8.3% | 100.0% | |
| 4.20 | 83.3% | 100.0% |
3.1.2 Ethanol
3.1.2.1 Locomotor activity
Lower doses of ethanol (1–1.78 g/kg) increased motor activity in both strains, while higher doses (3.2–4.2 g/kg) reduced locomotion (Fig. 2). A three-way ANOVA reveald a significant interaction between strain and ethanol dose (F[4, 600] = 11.40, P<0.001) as well as an interaction between strain and time [F(5, 600] = 4.24, P<0.001). Significant main effects of strain, time, and ethanol dose were also present (F[1, 600] = 11.45, F[5, 600] = 6.10, and F[4, 600] = 64.74, P<0.001). To further investigate effects of ethanol doses over time in each strain, separate two-way ANOVAs were performed for each strain. Ethanol and time were significant factors in both strains (see Table 1), as ethanol effects peaked between 5–10 min after administration. Post-hoc analysis revealed that a dose of 1.0 g/kg ethanol transiently increased locomotor activity in C57BL/6J mice, while 1.0 and 1.8 g/kg ethanol persistently increased locomotion in DBA/2J mice. Higher doses of ethanol (3.2 and 4.2 g/kg) reduced locomotor activity in both strains, although the effects of 3.2 g/kg dissipated within the first 20 minutes after injection in DBA/2J mice.
Figure 2.

Effecs of ethanol (i.p) on locomotor activity in C57BL/6J (A) and DBA/2J (B) mice. Each point represents the mean (± S.E.M.) of locomotor counts over the preceeding 5-min in all 12 subjects. Points labeled SAL represent effects following saline administration. Asterisks represent points that significantly differ from the corresponding VEH point. Some points have been offset for clarity.
3.1.2.2 Ataxia and Loss of Righting Reflex
As shown in Table 2, ethanol produced dose-dependent increases in ataxia and loss of righting reflex. Effects of ethanol dose on percent of mice displaying ataxia were significant in both strains (F[4,55]=90.75 for C57BL/6J and 94.25 for DBA/2J, P<0.05). The incidence of the loss of righting reflex was significantly increased following ethanol (F[4,55] = 27.28 for C57BL/6J and 34.06 for DBA/2J, P<0.05). Ataxic effects of ethanol occurred at slightly lower doses than loss of righting reflex in both strains. Strains did not differ in their sensitivity to ataxic or loss of righting reflex effects of ethanol.
3.1.3 Interactions between ethanol and taurine
3.1.3.1 Locomotor Activity
3.1.3.1.1 C57BL/6J mice
Taurine pretreatments did not alter ethanol effects on locomotion (Fig. 3A). There was a significant interaction between ethanol dose and time, as well as main effects of ethanol dose and time, but no interactions between taurine and ethanol or taurine and time, nor was there a main effect of taurine dose (see Table 1). A repeated measures two-way ANOVA revealed that the effect of ethanol alone was not different between the first and second determinations [F(1, 704) = 1.86].
Figure 3.

Effect of taurine and ethanol interactions on locomotor activity in in C57BL/6J (A) and DBA/2J (B) mice. Each point represents the mean (± S.E.M.) of locomotor counts over 30-min in all 12 subjects. Points labeled “SAL” and “SAL redet” represent the effects of ethanol alone in subjects before or after all interaction studies were complete, respectively.
3.1.3.1.2 DBA/2J mice
Taurine pretreatments did not appear to alter locomotor effects of ethanol, but this analysis was confounded by a change in the locomotor effects of ethanol over successive treatments (Fig. 3B). A significant interaction was present between ethanol dose and taurine dose (see Table 1). Main effects of ethanol dose, taurine dose, and time were also present. Further analysis revealed that this interaction was likely due to the initial determination of ethanol effects differing from subsequent determinations. A second ANOVA excluding the initial determination of ethanol effects alone yielded no effect of taurine dose, nor any interaction between taurine dose and ethanol dose or time. Indeed, the first and second determinations of ethanol effects alone differed, as determined by a repeated-measures ANOVA (F[1, 704] = 37.52, P<0.001). Thus, an order effect was present with the first ethanol determination differing from all subsequent determinations, including the re-determination in the absence of taurine pretreatment.
3.1.3.2 Ataxia
In C57BL/6J mice (Fig. 4A), there was no interaction between ethanol dose and taurine dose (F[16,264]=0.47, P>0.95). Nor was there a significant effect of taurine dose (F[4,264] = 1.08, P>0.36), although ethanol dose was a significant factor (F[4,264]=20.50, P<0.001).
Figure 4.

Ataxia and Loss of Righting Reflex in C57BL/6J (A, C) and DBA/2J (B, D). Points represent the percentage of subjects with ataxia or loss of righting reflex under each condition. Points labeled “SAL” and “SAL redet” represent the effects of ethanol alone in subjects before or after all interaction studies were complete, respectively. Some points have been offset for clarity.
In DBA/2J mice (Fig. 4B), there was no interaction between taurine dose and ethanol dose (F[16,264]=0.60, P>0.88), although significant main effects of taurine dose and ethanol dose were present (F[4,264] = 6.15, and F[4,264]=10.99, P<0.001 respectively). Because the effect of taurine could be due to a change in the ataxic effects of ethanol alone over repeated administration, a logistic regression was performed to compare only the first and last determinations, when ethanol was administered alone. There was an order effect (F[1,99]=9.82, P<0.005), although no order effect was present in C57BL/6J mice when the same analysis was performed (P=1).
3.1.3.3 Loss of Righting Reflex
In C57BL/6J mice (Fig. 4C), there was no interaction between ethanol dose and taurine dose (F[16,259]=0.08, P>0.99), nor was there a main effect of taurine dose (F[4,259]=0.08, P>0.98), although a main effect of ethanol dose was significant (F[4,259]=3.64, P<0.01). Results in DBA/2J mice were similar (Fig. 4D), with no significant interaction (F[2,264]=0.03, P>0.99), nor a main effect of taurine (F[4,264]=1.96, P>0.10), although ethanol dose was significant (F[4,264]=24.64, P<0.0001). These results demonstrate that taurine had no effect on the loss of righting reflex, nor was there any change in the effect of subsequent ethanol exposures in either strain.
3.2 Experiment 2: Between-subject effects
3.2.1 Locomotor effects
A three-way ANOVA revealed an interaction between strain and treatment condition (F[3, 2120] = 41.56, P<0.0001). Main effects of strain and time were also present (F[1, 2120] = 81.69 and F[19, 2120] = 11.14, P<0.0001). To evaluate the effects of taurine pretreatment in each strain, separate analyses were performed.
3.2.1.1 C57BL/6J mice
Taurine treatment altered the locomotor stimulant effects of ethanol in C57BL/6J mice. Considered across the entire 20-min after ethanol (or vehicle) injection, an interaction was present between treatment and time (F[57, 1080]=2.1). Main effects were also present for treatment and time (F[3, 1080]= 20.1 and F[19, 1080] = 12.9, respectively). Benjamani-Hochberg corrected t-tests revealed that the stimulant effect of ethanol waned after 5-min, so a second analysis, limited to data collected in the first 5-min after ethanol injection was performed (Fig. 5A). Though no interaction between treatment and time was present (F[15, 324]=0.8), main effects of treatment and time were significant (F[3, 324]=27.0 and F[6, 324]=5.6, respectively). Benjamani-Hochberg adjusted p-values revealed that locomotor activity was significantly elevated in the treatment group receiving only ethanol (saline + ethanol) compared with each of the other treatment groups (saline alone, or taurine + saline or taurine + ethanol). Taurine treatment alone significantly elevated locomotor activity when compared with saline treatment alone, but not compared with taurine + ethanol treatment.
Figure 5.

Between-subject effects on locomotor activity of interactions between taurine (1.78 g/kg) and ethanol (1.78 g/kg) in C57BL/6J (A) or DBA/2J (B) mice. Each point represents the mean (± S.E.M.) of locomotor counts over the preceeding 1-min in all 12 subjects. Conditions with “Sal” represent effects following saline alone (Sal Sal), taurine alone (Tau Sal), or ethanol alone (Sal Eth).
3.2.1.2 DBA/2J mice
Taurine treatment (1.78 g/kg) did not alter the locomotor stimulant effects of ethanol (1.78 g/kg) in DBA/2J mice. Considered across the 20-min following ethanol (or vehicle) injection, no interaction was present between treatment and time (F[57, 1080] = 0.7, P>.9). However significant effects of treatment and time were present ([F[3, 1080]=82.1 and F[19, 57]=2.1, resepctively). To provide parallel comparisons between strains, a second analysis was limited to the first 5-min following ethanol or vehicle injection (Fig. 5B). Main effects of treatment and time remained significant (F[3, 324]=29.2 and F[5, 324]=3.7, respectively). Benjamani-Hochberg adjusted t-tests revealed that locomotor activity was significantly elevated following ethanol alone compared with each of the other treatments. Taurine + ethanol treatment produced significant elevations in locomotion compared with saline alone or taurine alone, and significant reduction of locomotion compared with ethanol alone (P<0.05). Effects of taurine alone did not differ from effects following saline alone. Thus, in DBA/2J mice, taurine pretreatment significantly attenuated ethanol-stimulated locomotion, though not to ethanol-free levels, while taurine treatment alone had no effect.
3.2.2 Ataxic effects
3.2.2.1 C57BL/6J
Taurine pretreatment (1.78 g/kg) reduced the probability that ethanol (1.78 g/kg) treatment resulted in ataxia (Fig. 6A), an effect that approached significance at α=0.95, as determined with a Benjamani -Hochberg-adjusted Fisher test (P<0.054). The difference between ataxia following treatment with saline alone and ethanol alone approached significance at α=10% (P<0.108). Though inconclusive, these results are consistent with taurine pretreatment reducing the ataxic effect of ethanol in C57BL/6J mice.
Figure 6.
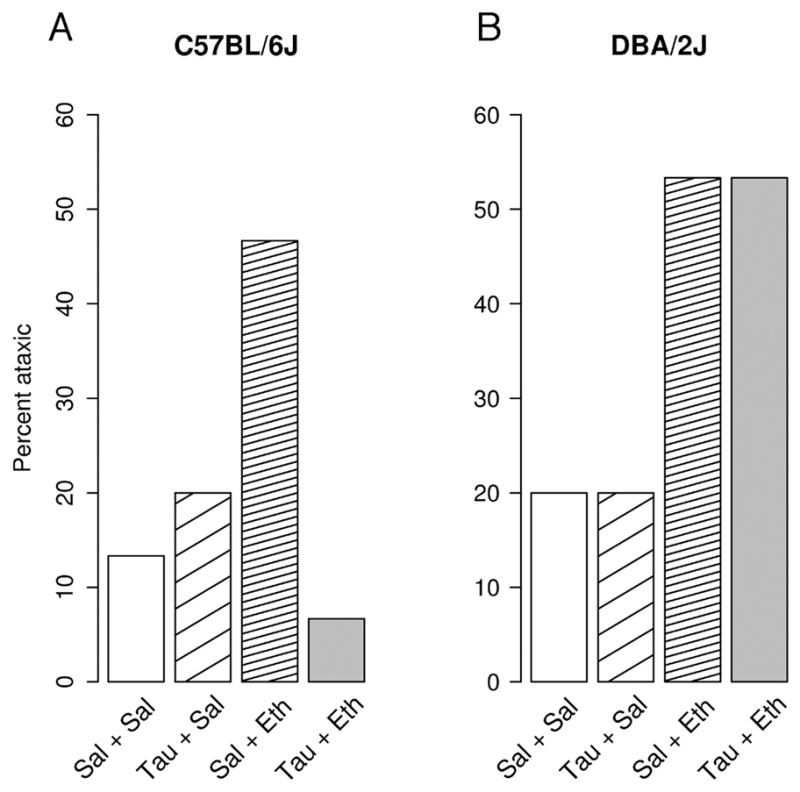
Ataxic effects of ethanol (1.78 g/kg) in combination with taurine (1.78 g/kg), as well as the effects of matched controls in C57BL/6J (A) or DBA/2J (B) mice. Subjects were the same as those represented in Fig. 5. Bars represent the proportion of subjects displaying ataxia under each treatment condition.
3.2.2.2 DBA/2J mice
Ataxia following ethanol administration alone tended to increase when compared with saline treatment alone as determined with Benjamani-Hochberg-adjusted Fisher tests (Fig. 6B), however these results were not significant (P<0.192). Taurine pretreatment had no effect on ethanol-induced ataxia P<0.642), suggesting that taurine does not alter the effect in DBA/2J mice.
4. Discussion
The major finding of the present study was that there is no interaction between taurine and ethanol under most of the conditions we studied. However, a dose of taurine (1.78 g/kg) with modest locomotor depressant activity significantly inhibited the locomotor stimulant and ataxic effects of ethanol (1.78 g/kg) in ethanol-naïve C57BL/6J and DBA/2J mice; however the reduction was greater in C57BL/6J mice. Various doses of taurine did not alter ethanol effects in subjects of either strain with previous ethanol experience. Thus, the present study demonstrates strain-specific effects of taurine on behavioral effects of ethanol, however this effect is modest and limited.
The effects of taurine alone did not differ between strains, although C57BL/6J mice appeared to be slightly less sensitive to the modest locomotor depressant effect of taurine. Further, taurine administration did not result in ataxia or loss of righting reflex at any dose tested. In light of the relatively large doses administered, this is surprising given the findings of others (Iida and Hikichi, 1976; Barbeau et al., 1975). It remains possible that peripherally administered taurine did not cross into the central nervous system, however taurine from the periphery does cross the blood-brain barrier via an active uptake mechanism (Lahdesmaki and Oja, 1973; Tamai et al., 1995). Further, the time course of taurine locomotor depressant effects is similar to that reported by Barbeau et al., (1975) although in that study taurine was administered centrally. Thus, the effects of taurine observed in the present study are consistent with effects in the CNS.
In C57BL/6J mice, ethanol administration can produce fleeting, modest increases in locomotor activity (in the first few minutes after administration) and also results in ataxia (Crabbe et al., 1982). Higher doses suppress locomotor activity and result in a loss of the righting reflex. Interestingly, Risinger et al., (1998) report that ethanol reinforces self-administration behavior in C57BL/6J mice, but not in DBA/2J mice (which display a much more robust locomotor-stimulant response to ethanol). In the present study, the effects of ethanol alone were as expected in both strains. Few others have reported the locomotor-enhancing effect of ethanol in C57BL/6J mice, likely due to longer sampling periods in other studies compared with the present study.
Taurine and ethanol interactions were apparent in the between-subjects experiment, but not in the within-subjects experiment. From these data, it appears that interactions between taurine and ethanol are only evident in ethanol-naïve subjects. This could be due to adaptation following repeated ethanol exposure. While there was evidence for the development of tolerance to the locomotor effects of ethanol in DBA2/J mice, no change in the effects of ethanol was observed in C57BL/6J mice. Even in drug-naïve mice, the effects of taurine on ethanol-induced locomotor activity were short-lived. Thus, taurine impacts ethanol effects quite subtly and transiently.
The behavioral effects of the interaction between taurine and ethanol were strain-dependent. Taurine pretreatment significantly attenuated locomotor stimulating effects of a 1.78 g/kg dose of ethanol in both strains. However, in C57BL/6J mice, locomotor activity was attenuated to a level observed after taurine treatment alone, while in DBA/2J mice taurine pretreatment attenuated ethanol-induced locomotor activity, but not to the degree seen in the absence of ethanol. This difference could be due to different effectiveness of taurine and ethanol in the two strains. Taurine produced greater reductions, and ethanol produced greater increases in locomotor activity in DBA/2J mice compared with C57BL/6J mice. Yet, taurine was more effective at reducing ethanol-induced locomotion in C57BL/6J mice. These results suggest that attenuated ethanol-induced locomotion by taurine is unlikely to be due to general depression of motor output.
Taurine had modest effects on locomotor activity when combined with ethanol. This result was in contrast with a previous study conducted in Swiss-Webster mice (Aragon et al., 1992). In this earlier study, taurine pretreatment at far lower doses than those used in the present study attenuated ethanol-stimulated locomotion at a dose on the ascending limb of the dose-effect curve and increased locomotor activity following the peak stimulant dose of ethanol. Although this discrepancy could be due to the different mouse strains used, Swiss-Webster mice appear to respond to ethanol administration similarly to DBA/2J mice. In the present study, DBA/2J mice were less sensitive to taurine effects than C57BL/6J mice, and no dose combination resulted in supermaximal locomotor activity in either strain. Thus a strain difference is not a completely satisfying rationalization. Differences in the doses of taurine used in each study might account for the different results reported, but it is difficult to argue that effects seen at lower taurine doses would disappear at higher doses, especially when the relatively low potency of taurine is considered (Olive, 2002). Finally, different temporal parameters could account for effects observed by Aragon et al. (1992). In their study, the authors administered taurine and ethanol simultaneously, then placed mice into the activity chamber, and began recording locomotor activity 10-min later. Thus, procedural differences might account for the contrasting results reported by Aragon et al., (1992). Additional studies would be required to bring clarity to this disparity.
Taurine appears to antagonize ethanol-induced ataxia in C57BL/6J, but not DBA/2J mice. Although this effect in C57BL/6J mice failed to reach significance at the 95% confidence level, the trend apparent in the data is consistent with a greater impact of taurine pretreatment in C57BL/6J mice compared with DBA/2J mice. The failure to detect significant differences between conditions in this experiment was likely an issue of power and might be easier to resolve with a larger sample size. Still this difference, in conjunction with the demonstrated strain-dependent locomotor effects, may reveal underlying differences in ethanol-taurine interactions between the two strains.
Others have reported that taurine pretreatment reduces the duration of ethanol-induced sleep-time or loss of righting reflex (Iida and Hikichi, 1976; Boggan et al., 1978). While the present study did not address this question, the results of this work demonstrate that taurine pretreatment does not alter the incidence of ethanol-induced loss of righting reflex. Thus, while it remains possible that the duration of the loss of righting in the present subjects could have been attenuated (but was not measured), subjects lost the righting reflex in the same proportion and at the same ethanol doses regardless of the presence or absence of taurine pretreatment.
Recent evidence suggests that energy drinks that contain taurine have a little to no impact on the behavioral effects of ethanol. However in each of these studies, the energy drink contained several other psychoactive ingredients, including caffeine. Thus, although results of these reports suggest taurine-containing energy drinks do not appear to modify ethanol behavioral effects, interpretation is difficult in the context of taurine-ethanol interactions. In one study in Swiss mice, 10.71 ml/kg of an energy drink blunted the attenuated locomotor activity due to administration of 2.5 g/kg ethanol (Ferreira et al., 2004a). However, the same dose of energy drink failed to alter locomotor activity enhancements of lower ethanol doses (Ferreira et al., 2004a). In humans, a taurine-containing energy drink failed to alter performance on cycle ergometer following ethanol (Ferreira et al., 2004b). The energy drink also had no effect on ethanol-induced changes in other physiological indicators such as oxygen uptake, respiratory exchange rate or blood pressure. Nor was any effect of energy drinks apparent on biochemical indices following ethanol such as blood levels of glucose, lactose, insulin, cortisol, ACTH, or monoamines (Ferreira et al., 2004b). A recent study by the same group found that a taurine-containing energy drink did not improve impairment of reaction time or coordination due to ethanol (Ferreira et al., 2006). Subjects did report increased pleasure and a reduction in sleepiness.
In summary, taurine pretreatment has a modest inhibitory effect on the locomotor-stimulating and ataxic effects of ethanol in ethanol-naïve mice. This effect appears to be more pronounced in C57BL/6J mice compared with DBA/2J mice. Taurine pretreament had no effect on ethanol-induced loss of righting reflex. However, in subjects with previous ethanol experience, taurine had no effect on any measure. Thus, taken together, the interaction between taurine and ethanol is subtle. The present data are inconsistent with the notion that taurine plays a major role in the neuropharmacology of ethanol. Further, in agreement with human studies exploring the impact of energy-drinks on ethanol neuropharmacology, the present results suggest that taurine does not attenuate the impairing effects of ethanol to any great extent, nor that taurine can stave off the incidence of ethanol-induced sedation.
Acknowledgments
The authors with to thank Roslyn Martinez and Jason Persyn for their technical assistance. Jason Persyn was supported by a summer research grant from the Cajal Institute, for which the authors are also grateful. Supported by PHS grant AA012337 All experiments described herein have been approved by the Institutioanl Animal Use Committee at the University of Texas Heath Science Center at San Antonio, and conform to the guidelines established in the Guide for the Care and Use of Laboratory Animals as adopted by the U.S. National Institutes of Health.
References
- Aragon CM, Trudeau LE, Amit Z. Effect of taurine on ethanol-induced changes in open-field locomotor activity. Psychopharmacology. 1992;107:337–340. doi: 10.1007/BF02245158. [DOI] [PubMed] [Google Scholar]
- Barbeau A, Inoue N, Tsukada Y, Butterworth RF. The neuropharmacology of taurine. Life Sci. 1975;17:669–677. doi: 10.1016/0024-3205(75)90520-2. [DOI] [PubMed] [Google Scholar]
- Baskin SI, Hinkamp DL, Marquis WJ, Tilson HA. Effects of taurine on psychomotor activity in the rat. Neuropharmacology. 1974;13:591–594. doi: 10.1016/0028-3908(74)90047-1. [DOI] [PubMed] [Google Scholar]
- Benjamani Y, Hochberg Y. Controlling the false discovery rate: a practical and powerful approach to multiple testing. J Roy Statist Soc Ser B. 1995;57:289–300. [Google Scholar]
- Boggan WO, Medberry C, Hopkins DH. Effect of taurine on some pharmacological properties of ethanol. Pharmacol Biochem Behav. 1978;9:469–472. doi: 10.1016/0091-3057(78)90043-6. [DOI] [PubMed] [Google Scholar]
- Crabbe JCJ, Johnson NA, Gray DK, Kosobud A, Young ER. Biphasic effects of ethanol on open-field activity: sensitivity and tolerance in C57BL/6n and DBA/2n mice. J Comp Physiol Psychol. 1982;96:440–451. doi: 10.1037/h0077898. [DOI] [PubMed] [Google Scholar]
- Ferko AP. Ethanol-induced sleep time: interaction with taurine and a taurine antagonist. Pharmacol Biochem Behav. 1987;27:235–238. doi: 10.1016/0091-3057(87)90564-8. [DOI] [PubMed] [Google Scholar]
- Ferreira SE, Hartmann Quadros IM, Trindade AA, Takahashi S, Koyama RG, Souza-Formigoni MLO. Can energy drinks reduce the depressor effect of ethanol? an experimental study in mice. Physiol Behav. 2004;82:841–847. doi: 10.1016/j.physbeh.2004.06.017. [DOI] [PubMed] [Google Scholar]
- Ferreira SE, de Mello MT, Pompéíia S, de Souza-Formigoni MLO. Effects of energy drink ingestion on alcohol intoxication. Alcohol Clin Exp Res. 2006;30:598–605. doi: 10.1111/j.1530-0277.2006.00070.x. [DOI] [PubMed] [Google Scholar]
- Ferreira SE, de Mello MT, Rossi MV, Souza-Formigoni MLO. Does an energy drink modify the effects of alcohol in a maximal effort test? Alcohol Clin Exp Res. 2004;28:1408–1412. doi: 10.1097/01.alc.0000139822.74414.ec. [DOI] [PubMed] [Google Scholar]
- Iida S, Hikichi M. Effect of taurine on ethanol-induced sleeping time in mice. J Stud Alcohol. 1976;37:19–26. doi: 10.15288/jsa.1976.37.19. [DOI] [PubMed] [Google Scholar]
- Lähdesmäki P, Oja SS. On the mechanism of taurine transport at brain cell membranes. J Neurochem. 1973;20:1411–1417. doi: 10.1111/j.1471-4159.1973.tb00253.x. [DOI] [PubMed] [Google Scholar]
- Olive MF. Interactions between taurine and ethanol in the central nervous system. Amino Acids. 2002;23:345–357. doi: 10.1007/s00726-002-0203-1. [DOI] [PubMed] [Google Scholar]
- Sgaragli G, Pavan F. Effects of amino acid compounds injected into cerebrospinal fluid spaces, on colonic temperature, arterial blood pressure and behaviour of the rat. Neuropharmacology. 1972;11:45–56. doi: 10.1016/0028-3908(72)90056-1. [DOI] [PubMed] [Google Scholar]
- Tamai I, Senmaru M, Terasaki T, Tsuji A. Na(+)- and cl(−)-dependent transport of taurine at the blood-brain barrier. Biochem Pharmacol. 1995;50:1783–1793. doi: 10.1016/0006-2952(95)02046-2. [DOI] [PubMed] [Google Scholar]
- Wise RA, Bozarth MA. A psychomotor stimulant theory of addiction. Psychol Rev. 1987;94:469–492. [PubMed] [Google Scholar]