

Abstract
4-Hydroxyphenylpyruvate dioxygenase (4HPPD) catalyzes the formation of homogentisate (2,5-dihydroxyphenylacetate) from p-hydroxyphenylpyruvate and molecular oxygen. In plants this enzyme activity is involved in two distinct metabolic processes, the biosynthesis of prenylquinones and the catabolism of tyrosine. We report here the molecular and biochemical characterization of an Arabidopsis 4HPPD and the compartmentation of the recombinant protein in chlorophyllous tissues. We isolated a 1508-bp cDNA with one large open reading frame of 1338 bp. Southern analysis strongly suggested that this Arabidopsis 4HPPD is encoded by a single-copy gene. We investigated the biochemical characteristics of this 4HPPD by overproducing the recombinant protein in Escherichia coli JM105. The subcellular localization of the recombinant 4HPPD in chlorophyllous tissues was examined by overexpressing its complete coding sequence in transgenic tobacco (Nicotiana tabacum), using Agrobacterium tumefaciens transformation. We performed western analyses for the immunodetection of protein extracts from purified chloroplasts and total leaf extracts and for the immunocytochemistry on tissue sections. These analyses clearly revealed that 4HPPD was confined to the cytosol compartment, not targeted to the chloroplast. Western analyses confirmed the presence of a cytosolic form of 4HPPD in cultured green Arabidopsis cells.
4HPPD (EC 1.13.11.27) catalyzes the formation of homogentisate (2,5-dihydroxyphenylacetate) from 4HPP and molecular oxygen. This reaction proceeds through an oxidative decarboxylation of the 2-oxoacid side chain of the substrate, which is accompanied by hydroxylation of the aromatic ring and a 1,2-migration of the carboxymethyl group (Jefford and Cadby, 1981). The purified enzyme was shown to contain nonheme-reduced iron, which is essential for catalytic activity (Wada et al., 1975; Lindblad et al., 1977; Roche et al., 1982; Endo et al., 1992; Rûetschi et al., 1993). This enzyme belongs to the extradiol α-ketoacid-dependent group of dioxygenases. However, in contrast to the other members of the group, the α-ketoacid is not a cofactor but forms part of the substrate 4HPP. In most organisms this enzyme activity is involved in the catabolism of the aromatic amino acid Tyr (Goodwin, 1972). All mammalian 4HPPDs purified so far behave as homodimers, with subunits of 43 to 49 kD (Wada et al., 1975; Lindblad et al., 1977; Roche et al., 1982; Endo et al., 1992; Rûetschi et al., 1993). In contrast, the Pseudomonas sp. strain P.S. 874 enzyme behaves as a homotetramer with subunits of 41 kD (Lindstedt et al., 1987).
In photosynthetic organisms, however, this enzyme plays a specific and crucial role because the product of the reaction, homogentisate, is the aromatic precursor of all plastoquinones and tocopherols, essential elements of the photosynthetic electron transport chain and of the antioxidative systems, respectively. Presumably, as in other organisms, plant 4HPPD is also involved in the degradation of Tyr (Durand and Zenk, 1974; Fernandez-Canon and Penalva, 1995). Interest in the plant enzyme was raised by the recent demonstration that 4HPPD is the target enzyme of the new bleaching herbicide families, sulcotrione (Prisbylla et al., 1993; Schulz et al., 1993; Secor, 1994) and isoxasol (Luscombe et al., 1995). The bleaching effect is associated with an accumulation of the carotenoid precursor, phytoene. It is believed that the effect results from an indirect inhibition of phytoene desaturase activity as a consequence of the depletion of the plastoquinone-cofactor pool (Schulz et al., 1993; Norris et al., 1995).
To improve our knowledge of this herbicide target, we recently purified to near homogeneity the 4HPPD of cultured carrot cells (Lenne et al., 1995) and isolated the corresponding cDNA (Garcia et al., 1997). Carrot 4HPPD resembles the mammalian enzyme because it behaves as a homodimer with 48-kD subunits (Lenne et al., 1995; Garcia et al., 1997). In contrast, it was recently reported that partially purified maize 4HPPD coelutes with a monomeric polypeptide of 43 kD (Barta and Böger, 1996). The involvement of plant 4HPPD activity in metabolic processes as different as Tyr degradation and prenylquinone biosynthesis suggests the existence of different isoforms of 4HPPD associated with different subcellular compartments. In our previous study of carrot 4HPPD, we demonstrated, for the first time to our knowledge, that this enzyme activity in plants was associated with the cytosol (Lenne et al., 1995; Garcia et al., 1997). Such a subcellular localization is in apparent contradiction with the situation previously described in spinach by Fiedler et al. (1982), who reported the existence of two pools of 4HPPD activity, one associated with the chloroplast and the other with peroxisomes.
We report here the molecular and biochemical characterization of an Arabidopsis 4HPPD. We also examined the cellular compartmentation of the recombinant Arabidopsis 4HPPD by overexpressing the complete coding sequence in transgenic tobacco.
MATERIALS AND METHODS
Isolation of a Full-Length 4HPPD cDNA
A keyword search of the database identified an Arabidopsis expressed sequence tag clone that contained an open reading frame similar to human and rat 4HPPD. We sequenced this clone, 96B13T7 (accession no. T20952), obtained from the Arabidopsis Stock Center (The Ohio State University, Columbus). Its open reading frame coded for a polypeptide of 75 amino acids, exhibiting high homology with the C-terminal sequence of mammalian 4HPPD. To obtain a full-length Arabidopsis 4HPPD clone, the insert 96B13T7 was radioactively labeled and used as a probe to screen a cDNA library of young Arabidopsis leaves constructed in λZAPII (Stratagene). Plaque screening was performed according to the manufacturer's instructions. Approximately 250,000 clones were screened, yielding six positive cDNA clones. We analyzed the clone containing the longest insert and carried out DNA sequence analysis on both strands, using a kit (PRISM, Applied Biosystems) with fluorescent dideoxynucleotides, Taq DNA polymerase, and T3 and T7 universal primers. Specific oligonucleotide primers were used for further sequencing. The programs, Gene Works 5.2 (Oxford Molecular Group, Oxford, UK) and PCGENE (Intelligenetics, Oxford Molecular Group), performed the sequence analyses.
Cloning of the Arabidopsis 4HPPD Sequence into the Expression Vector pTrc 99A
The pTrc 99A-AT4-4HPPD plasmid coding for an Arabidopsis 4HPPD protein was constructed via site-directed mutagenesis using PCR amplification of the entire Arabidopsis 4HPPD cDNA. The following oligonucleotides were used: P1 (5′-GTTGGTGAAATCCATGGGCCACCAAAACGCCG-3′), which introduces a NcoI restriction site containing the ATG translation-initiation codon (underlined), and P2 (5′-GTTGGTTCTTCGTCGACTTCATCCCAC-3′), which is complementary to the 3′ end of the cDNA-coding region and introduces a SalI restriction site (underlined) 1 bp after the TGA stop codon. PCR was performed for 15 cycles, including 1 min of denaturation at 95°C, 1 min of annealing at 58°C, and 2 min of DNA elongation at 72°C, driven by the Pwo DNA polymerase (Boehringer Mannheim). The PCR DNA fragment was subcloned into the pTrc 99A vector (Pharmacia), which was digested by NcoI-SalI restriction enzymes. This oriented cloning put the Arabidopsis 4HPPD cDNA under the control of the lacUV5 promoter. The DNA insert was sequenced on both strands to ensure that no mutation had been introduced during the course of PCR amplification. The plasmid was termed pTrc 99A-AT4-4HPPD.
Genomic Southern Analysis
Total DNA was isolated from young Arabidopsis plants, and 4 to 8 μg was digested overnight with the appropriate restriction enzymes (New England Biolabs). Fragments were then separated by 0.8% (w/v) agarose gel electrophoresis for 6 to 8 h. After alkaline blotting to a Hybond-N+ membrane (Amersham), hybridization was carried out with the full-length cDNA or with the 5′ or 3′ end of the full-length cDNA (BamHI-PvuI, 568 bp for the 5′ end and PvuI-XhoI, 959 bp for the 3′ end) that had been 32P-labeled with a random-priming kit (Ready-to-Go, Pharmacia).
Northern Analysis
mRNAs isolated from Arabidopsis seedlings using an mRNA-isolation system (Straight A, Novagen, Madison, WI) were denatured (2–4 μg, 1 h at 50°C in 10 mm NaPi, pH 7.0, 1.08 m glyoxal, 2% [v/v] DMSO), separated by 1% (w/v) agarose gel electrophoresis, and transferred to a nylon membrane (Nytran, Schleicher & Schuell) according to the method of Thomas (1980). The resulting blots were subjected to hybridization with the desired 32P-labeled cDNA probe.
Overproduction of Recombinant Arabidopsis 4HPPD
Escherichia coli JM105 cells harboring the pTrc 99A-AT4-4HPPD plasmid were grown at 37°C in 1 L of Luria-Bertani broth supplemented with 100 μg mL−1 carbenicillin and 100 μg mL−1 streptomycin (Maniatis et al., 1982). Isopropyl-β-d-thiogalactoside was added to a final concentration of 1 mm when bacterial growth was equivalent to an A600 of 0.6. The cells were grown for another 16 h at 30°C. The cells were harvested by centrifugation and the pellet was resuspended in 10 mL of buffer A (20 mm potassium phosphate, pH 6.8, 1 mm EDTA, 1 mm DTT, 5 mm 6-aminohexanoic acid, and 1 mm benzamidine) and sonicated using a cell disrupter (Vibra-Cell, Sonics & Materials, Danbury, CT) (100 pulses every 3 s on a power setting of 5). The crude extract was centrifuged at 35,000g for 30 min to yield a cell-free supernatant.
Electrophoretic Analyses of Proteins
Proteins were separated by SDS-PAGE containing 12% (w/v) acrylamide. The experimental conditions for gel preparation, sample solubilization, electrophoresis, and gel staining were as detailed by Chua (1980). PAGE under nondenaturing conditions was carried out at equilibrium in the absence of any denaturing agent (SDS or DTT), as described by Lasky (1978), on a linear acrylamide gradient (3.5%–27%) with a 3.5% acrylamide stacking gel.
Immunoblotting Analysis
After separation by PAGE, the proteins were electrophoretically transferred onto nitrocellulose membranes (Bio-Rad) according to the method of Towbin et al. (1979). Membranes were incubated for 30 min in TBS (10 mm Tris-HCl, pH 7.6, and 150 mm NaCl) containing 2% (v/v) Tween 20. They were incubated for 2 h with the specific antibodies, in TBS plus 0.05% (v/v) Tween 20, and 1 h with goat anti-rabbit IgG horseradish peroxidase conjugate (Bio-Rad). Membranes were stained for peroxidase activity using chloronaphtol and hydrogen peroxide as substrates. Western analyses of the subcellular fractions of Arabidopsis cells were revealed by chemiluminescence according to the manufacturer's protocol (Boehringer Mannheim).
Assay for 4HPPD
HPLC Determination of Homogentisate
Activity was measured in a 200-μL volume containing 100 mm Tris-HCl, pH 7.5, 50 mm ascorbate, and 200 μm 4HPP. The reaction medium was incubated for 15 min at 30°C, except when otherwise stated. The reaction was stopped by the addition of 70 μL of 20% (w/v) perchloric acid. The precipitated protein was removed by centrifugation at 15,000g for 5 min. The amount of homogentisate formed was determined by HPLC. An aliquot fraction (50–100 μL) of the perchloric acid supernatant was injected onto a pico-Tag C18 column (3.9 mm × 15 cm, 10-μm particle size; Millipore) connected to an HPLC system. The buffers used for elution were: buffer A, 0.1% (v/v) trifluoroacetic acid in distilled water, and buffer B, 0.07% (v/v) trifluoroacetic acid in 80% (v/v) CH3CN. The following linear gradients were used: 0% (100% buffer A) to 70% buffer B, 0 to 17 min; 70% to 100% buffer B, 17 to 20 min; 100% buffer B, 20 to 24 min; 100% to 0% buffer B, 24 to 28 min, flow rate 1 mL min−1. We detected the homogentisate by measuring the UV A288. We quantitated the homogentisate by measuring peak areas with a chromatograph data system (450-MT2 software, Kontron Instruments, Eching, Germany). Peak areas were converted to nanomoles of homogentisate by comparison with a standard curve.
Oxygen Consumption
The activity of the recombinant Arabidopsis 4HPPD was followed by measuring the oxygen consumption with a Clark-type electrode (Hansatech, King's Lynn, Norfolk, UK). We measured the activity in 500 μL, containing 100 mm Tris-HCl, pH 7.5, 50 mm ascorbate, and 200 μm 4HPP, at 30°C.
Transformation of Tobacco Plants
Plasmid Constructions
We used two different constructs: Ara5, which contained the complete coding sequence of the 4HPPD cDNA of Arabidopsis, and Ara9, which was a fusion between an OTP and the complete coding sequence of the 4HPPD cDNA of Arabidopsis. The coding sequence for the OTP was constructed from the coding sequence for the transit peptide (171 bp) of the Rubisco small subunit from sunflower (Waksman and Freyssinet, 1987), followed by the first 66 nucleotides coding for the mature part of the Rubisco small subunit from maize. The latter sequence was followed by the 150 nucleotides coding for the transit peptide of the Rubisco small subunit from maize (Lebrun et al., 1987). This yielded a NcoI-SphI fragment of 387 bp. The presence of the first 66 nucleotides of the mature Rubisco improved the targeting, and the second transit peptide prevented the production of a fusion protein. Details of this OTP fragment have been published (Lebrun et al., 1992).
The fusion OTP complete coding sequence of the 4HPPD cDNA of Arabidopsis was obtained by digesting the plasmid pRPA-RD-7 (Sailland et al., 1996), which contained the OTP fragment, with SphI. The SphI end was then treated with T4 polymerase, and the plasmid was digested with SalI. The complete coding sequence of the 4HPPD cDNA of Arabidopsis cloned in the plasmid pTrc 99A-AT4-HPPD was digested with NcoI; its NcoI end was treated with the Klenow polymerase, and the fragment was digested with SalI. This fragment was cloned into digested pRPA-RD-7, yielding the pRPA-Fus8 plasmid. The NcoI-SalI OTP-4HPPD fragment digested from pRPA-Fus8 and the NcoI-SalI 4HPPD fragment digested from pTrc 99A-AT4-HPPD were cloned into the pRPA S plasmid. This plasmid contained the duplicated promoter from the histone gene H4748 of Arabidopsis and the transcription activator from the tobacco etch virus, yielding, respectively, pRPA-S-ARA9 and pRPA-S-ARA5. A PmeI-SalI fragment containing the duplicated promoter of the histone gene, the transcription activator from tobacco etch virus, and either the fusion OTP-4HPPD or the 4HPPD was digested from pRPA-S-ARA9 and pRPA-S-ARA5, respectively, and cloned into the binary vector, pRPA 150 Aα2–2005, which was digested by SmaI and SalI to introduce the Nos terminator at the end of the construct, yielding the plasmids pRPA 150 Aα2-ARA5 and pRPA 150 Aα2-ARA9.
Plant Transformation
Plasmids pRPA 150 Aα2-ARA5 and pRPA 150 Aα2-ARA9 were transferred by the helper plasmid JC2073 (Lebrun et al., 1992) to the Agrobacterium tumefaciens EHA 101 (Sailland et al., 1996) by triparental mating (Lebrun et al., 1992) and used for transformation of tobacco (Nicotiana tabacum cv PBD6) by leaf-disc transformation, according to the method of Horsh et al. (1985). Transformed cells were selected on regenerating medium containing 0.3 mg L−1 isoxafutole, a potent inhibitor of 4HPPD (Luscombe et al., 1995), for at least 5 weeks. In the presence of this 4HPPD inhibitor, untransformed cells developed white callus, whereas transformed cells developed green callus as a consequence of the overexpression of Arabidopsis 4HPPD (Sailland et al., 1996). The presence of the transgene was confirmed by PCR and western analyses.
Immunocytochemical Localization of 4HPPD in Transformed Tobacco Plants
Samples were dissected from the median part of young, subapical tobacco leaves in a drop of fixative solution and then immersion fixed for 3 h at room temperature in 2.5% paraformaldehyde and 0.1% glutaraldehyde in 0.05 m citrate phosphate buffer, pH 7.0. Leaf samples were subsequently rinsed in 0.1 m buffer, dehydrated in ethanol up to 70%, and then embedded in London White resin in gelatin capsules. Polymerization was carried out for 22 h at 56°C. Cross-sections, 80 nm thick, were cut from the blocks with a diamond knife and harvested on 300-mesh, Formvar-coated nickel grids. The grids were successively floated for 1 h on a blocking solution of 3% BSA and 0.01% Tween 20 in 0.05 m TBS and then twice for 5 min on a dilution buffer composed of 0.3% BSA and 0.01% Tween 20 in 0.05 m TBS. The primary antibody reaction for 4HPPD detection was performed in a 1:5 dilution of affinity-purified rabbit anti-4HPPD serum in a dilution buffer. This step was omitted in the controls. Secondary antibody reaction was made by floating the grids on drops of a 1:50 dilution of the commercial gold-conjugated goat anti-rabbit serum (British Biocell International, Cardiff, UK). Two gold particle sizes were used: 15 nm, which was directly viewed, and 10 nm, which was subsequently amplified with a silver-enhancing kit (British Biocell International). In both cases, grids were floated for 20 min on a 3% (w/v) aqueous solution of uranyl acetate for additional contrast. Control and labeled sections were viewed at 80 kV on an electron microscope (model CM10, Philips, Eindhoven, The Netherlands) at the Electron Microscopy Center for Biology and Geology at the University of Lyon (France).
Preparation and Fractionation of Arabidopsis Cell Protoplasts
Preparation of protoplasts by enzymic digestion of 7-d-old cell cultures of Arabidopsis cell T87 (Axelos et al., 1992) and measurement of marker enzyme activities were carried out as described previously for carrot cell protoplasts (Garcia et al., 1997).
RESULTS
Isolation of cDNA Clones Encoding Arabidopsis 4HPPD
To characterize the Arabidopsis 4HPPD at the molecular level, we isolated a full-length 4HPPD cDNA by screening a cDNA library of young Arabidopsis leaves with an Arabidopsis expressed sequence tag as a nucleotide probe (accession no. T20952). The predicted polypeptide encoded by this 4HPPD clone is presented in Figure 1 and matches closely that of the carrot 4HPPD (Garcia et al., 1997).
Figure 1.

Amino acid sequence comparison between Arabidopsis and carrot 4HPPD. Conserved regions are boxed. Dashes indicate gaps introduced to maximize alignment.
Southern and Northern Analyses
Southern analysis was used to examine the number of copies of the gene encoding the Arabidopsis 4HPPD cDNA. Total DNA was digested with restriction enzymes that cut once (PvuI) or did not cut (BamHI, EcoRV, and HindIII) within the cDNA; the DNA was then double digested with the following enzyme combinations: PvuI-BamHI, PvuI-HindIII, and EcoRV-HindIII. As shown in Figure 2, hybridization of the DNA digested with BamHI, EcoRV, or HindIII with the full-length probe produced one band of 23.0, 9.4, and 3.9 kb, respectively. Digestion with PvuI resulted in two bands (3.0 and 25 kb). When the DNA was double digested by PvuI-BamHI, two bands of 3.0 and 12.6 kb were revealed. The double digestion by PvuI-HindIII revealed three bands of 3.9, 2.5, and 1.6 kb. These results, along with those obtained by hybridization of the 5′ and 3′ ends of the probe, and the fact that Southern hybridization carried out at the lower temperature of 50°C did not change the pattern of hybridization, strongly suggest the existence of a single copy of the gene encoding this 4HPPD in Arabidopsis.
Figure 2.

Southern hybridization of total Arabidopsis DNA with the complete 4HPPD cDNA as a probe. Total leaf DNA from Arabidopsis was digested (5 μg per reaction) with: lane 1, BamHI; lane 2, EcoRV; lane 3, HindIII; lane 4, PvuI; lane 5, PvuI and BamHI; lane 6, PvuI and HindIII; and lane 7, EcoRV and HindIII. DNA restriction fragments were separated on a 0.8% agarose gel, transferred to a nylon membrane, and then hybridized with the 32P-labeled probe corresponding to the complete coding region of the 4HPPD cDNA.
The steady-state concentration of 4HPPD mRNA was examined in both etiolated and green young Arabidopsis seedlings by northern analyses and compared with the corresponding mRNA steady-state concentration of a constitutively expressed gene (i.e. the translation initiation factor NelF-4A10; Mandel et al., 1995). Figure 3 shows that with both probes, the pattern of expression is similar. The transcripts were found to be more abundant in mRNA samples from etiolated tissue. This was certainly due to the presence of light-dependent mRNAs in the green tissues, which led to an underrepresentation of light-independent mRNAs. mRNA of 4HPPD therefore seemed to be similarly expressed in etiolated and greening tissues.
Figure 3.

Northern analysis of Arabidopsis mRNA of etiolated and green seedlings. mRNA (2 μg) from 15-d-old etiolated seedlings and 10-d-old green seedlings were submitted to northern analyses as described in Methods. The blot was then subjected to hybridization with a 32P-labeled cDNA probe coding for 4HPPD, and submitted to autoradiography for 1 week (A). The northern blot was then dehybridized, and rehybridized with a 32P-labeled cDNA probe of the constitutively expressed gene of the translation initiation factor NelF-4A10 (Mandel et al., 1995), and submitted to autoradiography for 24 h (B).
Functional Characterization of the Recombinant 4HPPD
As shown in Figure 4, the recombinant Arabidopsis 4HPPD was overproduced to approximately 5% to 10% of the total soluble protein of E. coli. (Fig. 4A). The recombinant 4HPPD was specifically detected by a polyclonal antibody raised against the purified carrot 4HPPD (Fig. 4B). Western analysis revealed a major band that corresponded to the recombinant 4HPPD and a minor proteolytic degradation. E. coli JM105 harboring the plasmid PTrc 99A-AT4-HPPD produced a characteristic brownish pigment that we had previously observed with E. coli when overproducing the recombinant carrot 4HPPD (Garcia et al., 1997); the brownish pigment had also been noted by other groups cloning 4HPPD from various origins. This pigment is an oxidation product of homogentisate (Denoya et al., 1994; Wyckoff et al., 1995; Lee et al., 1996). Activity of recombinant 4HPPD in crude extracts was characterized using either an HPLC assay to follow the formation of homogentisate (Fig. 5A) or the continuous measurement of oxygen consumption during catalysis (Fig. 5B). Both methods yielded similar values for the apparent Km for 4HPP (5–8 μm). However, the continuous measurement of oxygen consumption allowed a more accurate monitoring of the initial rate of the reaction. By this method, the apparent Vmax was slightly higher (100–120 nmol homogentisate formed min−1 mg−1 protein extract) than that measured by the HPLC assay (70–90 nmol). The enzyme had a rather broad pH optimum between 6.5 and 7.5 in our standard conditions (Fig. 5C) and the activity was found to be strictly dependent on the presence of ascorbate (results not shown). In agreement with previous data on plant 4HPPD (Prisbylla et al., 1993; Schulz et al., 1993; Secor, 1994; Lenne et al., 1995; Barta and Böger, 1996; Garcia et al., 1997), this recombinant 4HPPD activity was inhibited by low amounts of sulcotrione (Fig. 5A).
Figure 4.

Overproduction of Arabidopsis 4HPPD in E. coli JM105 cells. Soluble protein extracts of JM105 cells harboring the plasmid pTrc 99A-AT4-4HPPD harvested after 16 h of isopropyl-β-d-thiogalactoside induction were subjected to SDS-PAGE (12%) and stained with Coomassie blue (A), or analyzed by western blotting using immunopurified polyclonal rabbit antibody raised against the recombinant carrot 4HPPD (B) (Garcia et al., 1997).
Figure 5.

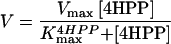 |
Subcellular Localization of the Recombinant 4HPPD
The program PSORT failed to predict conclusively the presence of an N-terminal transit peptide from the complete primary peptide sequence of this Arabidopsis 4HPPD. To clarify this point, we overexpressed its complete coding sequence in tobacco and examined the subcellular localization of the recombinant protein. Two different constructs were used for the transformation of tobacco: Ara5, which contained the complete coding sequence of the protein, and Ara9, which contained a fusion between an OTP and the complete coding region of 4HPPD. The latter construct served as a positive control for chloroplastic targeting of a heterologous recombinant protein. Both constructs were under the control of a duplicated histone promoter of Arabidopsis and the Nos terminator of the nopaline synthase gene of the T-DNA (for more details, see Methods).
Selection of transformed material was carried out in the presence of isoxafutole. After 15 d of selection, only the cells overexpressing 4HPPD were able to regenerate green calli (Sailland et al., 1996). Indeed, calli from nontransformed cells became white, presumably due to the depletion of their plastoquinone pool, which depressed their phytoene desaturase activity and thus prevented the accumulation of carotenoids. The presence of the coding sequence of Arabidopsis 4HPPD in the genome of transformed tobacco was verified by PCR amplification using specific primers corresponding to the 5′ and 3′ ends of the coding sequence. In all cases a DNA fragment of approximately 1.3 kb was amplified. The length of this fragment matched the length of the coding sequence of Arabidopsis 4HPPD. No amplification was observed when using genomic DNA from wild-type tobacco as the template for the PCR amplification. Thus, the presence of this 1.3-kb fragment confirmed that tobacco plants selected on an isoxafutole-containing medium were effectively transformed.
Overproduction of recombinant Arabidopsis 4HPPD in transformed tobacco plants was confirmed by immunodetection, using a polyclonal antibody raised against carrot 4HPPD (Garcia et al., 1997). As previously observed with tobacco plants that overexpress the 4HPPD of P. fluorescence (Sailland et al., 1996), tobacco plants that overexpress the Arabidopsis 4HPPD became resistant to high doses of isoxafutole (up to 400 g ha−1 for the best events of transformation).
Immunolocalization of the Recombinant 4HPPD in Transformed Tobacco Leaves
4HPPD was immunodetected by western analyses of protein extracts from purified chloroplasts and from total soluble leaf extract. In denaturating conditions the large subunit of Rubisco, the most abundant protein of green tissue, had a molecular mass close to that of 4HPPD (52 and 49 kD, respectively), and its migration during SDS-PAGE interfered with that of 4HPPD, thereby preventing its accurate immunodetection. Under native conditions, the recombinant Arabidopsis 4HPPD migrated as a 100-kD polypeptide (Fig. 6), confirming that, as with the carrot enzyme (Garcia et al., 1997), it behaved as an homodimer. In these conditions the recombinant 4HPPD was well separated from Rubisco, which had a native molecular mass in excess of 500 kD. Protein extracts were separated by nondenaturating electrophoresis prior to western analysis using immunopurified anti-carrot 4HPPD antibody.
Figure 6.

Immunodetection of Arabidopsis 4HPPD overproduced in transgenic tobacco. Proteins from total extracts (lanes 1) and Percoll- purified chloroplasts (lanes 2) obtained from young leaves of wild-type tobacco (a) (120 μg), transgenic tobacco transformed with ARA9 construct (b) (40 μg), and transgenic tobacco transformed with ARA5 construct (c) (120 μg) were separated on a linear gradient of 3% to 27% (w/v) acrylamide in nondenaturing conditions and analyzed by western blotting using immunopurified polyclonal rabbit antibodies (1:2000 dilution) raised against the purified recombinant carrot 4HPPD (Garcia et al., 1997).
No 4HPPD protein was immunodetected in the protein extracts of wild-type tobacco leaves (Fig. 6a). Anti-carrot 4HPPD antibody was nevertheless able to immunodetect 4HPPD in maize and maple cells (results not shown), suggesting that the endogenous tobacco 4HPPD was of low abundance. In tobacco harboring the Ara9 construct (fusion OTP-4HPPD), a peptide of 98 to 100 kD, which corresponds to the homodimer of the recombinant Arabidopsis 4HPPD, was immunodetected in both the total cellular extract and the chloroplastic extract (Fig. 6b). In contrast, plants transformed with the Ara5 construct (4HPPD alone) did not exhibit any signal in the chloroplastic extract (Fig. 6c); the recombinant 4HPPD was immunodetected only in the total cellular extract, indicating that the recombinant protein was not targeted to the chloroplast.
The subcellular localization of the recombinant 4HPPD in transformed tobacco plants was further examined by immunocytochemistry. No specific reaction was observed in sections of wild-type tobacco leaves (results not shown), consistent with the finding that this enzyme was at low abundance. Sections obtained from tobacco transformed with the Ara9 construct (OTP-4HPPD) revealed a strong specific reaction exclusively inside the chloroplasts (Fig. 7, A and C). In contrast, sections from tobacco leaves transformed with the Ara5 construct (4HPPD) showed no reaction over the level of the background inside chloroplasts, mitochondria, or peroxisomes. The specific reaction occurred exclusively in the cytosolic compartment, indicating that the recombinant 4HPPD remains in the cytosol (Fig. 7, B and D).
Figure 7.

Subcellular localization of the recombinant Arabidopsis 4HPPD overproduced in parenchyma cells of transgenic tobacco leaves by immunocytochemistry. A, Transgenic tobacco transformed by the construct ARA9 (×5,700); palisade layer (AB1 1/25, AB2-gold Φ 10 nm, 1/50, amplification 2 min). A very intense signal was specifically revealed within the chloroplasts. B, Transgenic tobacco transformed by the construct ARA5 (×5,700); spongy mesophyll (AB1 1/5, AB2-gold Φ 10 nm, 1/50, amplification 2 min). The recombinant 4HPPD was specifically revealed in the cytosol, the other subcellular compartments were at background level. C, Transgenic tobacco transformed by the construct ARA9 (×21,000); spongy mesophyll (AB1 = 1/25; AB2-gold Φ 10 nm, 1/50). Detail of chloroplasts containing the recombinant 4HPPD. D, Transgenic tobacco transformed by the construct ARA5 (×21,000); spongy mesophyll (AB1 = 1/5; AB2-gold Φ 10 nm, 1/50). High magnification showing the cytosolic signal of 4HPPD. The few signals within the chloroplast were at backgrown level. ch, Chloroplast; cy, cytosolic compartment; i, intercellular space; m, mitochondrion; v, vacuole; w, cell wall. Immunopurified rabbit IgG raised against carrot 4HPPD (AB1) was the primary antibody. Goat anti-rabbit conjugated with colloidal gold particles of 10 or 15 nm (AB2) was the secondary antibody. Scale bar = 1 μm.
Immunolocalization of 4HPPD by Western Analyses of Cultured Arabidopsis Cells
To confirm the existence of a cytosolic form of 4HPPD in Arabidopsis, we carried out a large-scale purification of enriched cytosolic and chloroplastic fractions. The starting material was green cultured cell suspensions, because it was not possible to obtain subcellular fractions directly from Arabidopsis plantlets. Intact protoplasts obtained from Arabidopsis cultured cells (T87) (Axelos et al., 1992) were fractionated by gentle rupture through a nylon mesh, and fractions enriched in cytosol and chloroplasts were prepared by differential centrifugation, as described previously (Garcia et al., 1997). Protein extracts were then separated by nondenaturing electrophoresis prior to western analysis, using immunopurified anti-carrot antibodies. To overcome the problem of the weak immunological detection of the enzyme, we incubated the nitrocellulose membrane with anti-4HPPD antibodies for 24 h at 4°C, and peroxidase was revealed by chemiluminescence (Boehringer Mannheim). Under these conditions anti-carrot 4HPPD antibodies reacted only with a polypeptide of 100 kD in a crude protein extract of protoplasts (Fig. 8, lane 1). This polypeptide was specifically detected in the cytosolic fraction (Fig. 8, lane 3). Conversely, no polypeptide was revealed in the chloroplastic fraction (Fig. 8, lane 2), confirming the presence of a cytosolic form of 4HPPD in Arabidopsis cells.
Figure 8.

Immunodetection of 4HPPD in subcellular fractions of green cultured cells of Arabidopsis. Lane 1, Purified protoplasts (100 μg); lane 2, enriched chloroplastic fraction (100 μg); and lane 3, enriched cytosolic fraction (100 μg). Total extracts were separated on a linear gradient of 3% to 27% (w/v) acrylamide in nondenaturing conditions and analyzed by western blotting using a 1:2000 dilution of an immunopurified polyclonal rabbit anti-carrot 4HPPD antibodies (Garcia et al., 1997) revealed by chemiluminescence.
DISCUSSION
Plant 4HPPD is an intriguing enzyme because it is involved in two distinct processes: the catabolism of Tyr and the biosynthesis of prenylquinones. It was first assumed that the substrate of this enzyme activity, 4HPP, was a chloroplastic intermediate in the shikimate biosynthetic pathway leading to Tyr. However, it is now well documented that arogenate appears to be the exclusive precursor of l-Tyr in most plant species, including tobacco and spinach (Jung et al., 1986). Therefore, 4HPP and, consequently, homogentisate derive exclusively from the degradation of Tyr. Homogentisate may either enter the prenylquinone biosynthesis pathway or be metabolized by homogentisate dioxygenase (Tyr catabolism). To regulate the flux of homogentisate between these two processes, one might postulate the involvement of distinct isoforms of 4HPPD. Studies using spinach (Fiedler et al., 1982) suggested the existence of a chloroplastic form of 4HPPD, responsible for the biosynthesis of prenylquinones, and a peroxisomal form, implicated in the degradation of Tyr. However, in our recent reports on carrot cells (Lenne et al., 1995; Garcia et al., 1997), we demonstrated that 4HPPD activity was confined to the cytosol. Furthermore, in these studies no other 4HPPD could be detected either biochemically or immunologically in the other cellular compartments. Because the studies involving carrot 4HPPD were carried out using nongreen cultured cells, we decided to investigate the situation in green plants.
We isolated and cloned an Arabidopsis 4HPPD cDNA and compared the molecular and biochemical characteristics and cellular localization of the recombinant protein with those from carrot cells. The predicted protein encoded by this cDNA was 70% identical to the carrot 4HPPD. The presence of an in-frame TGA stop codon 90 bp upstream from the first ATG in the carrot cDNA confirmed that it was full length (Garcia et al., 1997), and although such a stop codon was not found in the longest Arabidopsis 4HPPD clone presently obtained, the similarity between the two predicted proteins strongly suggested that the coding sequence of this Arabidopsis clone cDNA was also full length.
The biochemical characterization of this Arabidopsis 4HPPD, which was investigated by overproducing the recombinant protein in E. coli JM105, indicated that it behaves very similarly to the native plant 4HPPDs examined so far (Lenne et al., 1995; Secor, 1995; Barta and Böger, 1996; Garcia et al., 1997).
The identification of 4HPPD as a very potent target enzyme for new herbicide molecules, together with the involvement of 4HPPD in processes as different as Tyr degradation and prenylquinone biosynthesis, makes the determination of putative isoforms of crucial importance. Southern analysis strongly suggested that the Arabidopsis 4HPPD is encoded by a single-copy gene. No other bands were revealed in Southern blots carried out at lower stringency, suggesting that, if a second 4HPPD gene was present in Arabidopsis, its sequence was markedly different from that presently reported. The regulation of message levels might give some hint as to the pathway (catabolism or anabolism) in which the presently isolated 4HPPD is involved. For example, one might expect an 4HPPD involved in the biosynthesis of plastoquinones to be more abundant in greening seedlings than in etiolated ones, because the buildup of the photosynthesis apparatus requires a large amount of plastoquinones. In that context, it was recently reported that the mRNA of the barley 4HPPD shows enhanced expression during senescence (Kleber-Janke and Krupinska, 1997); these authors suggested that this is the consequence of an increased synthesis of tocopherols.
Comparison of the 4HPPD mRNA steady-state concentration with that of the mRNA of the constitutively expressed gene of the translation initiation factor NelF-4A10 (Mandel et al., 1995) (as a control) indicated that mRNA of 4HPPD is not more abundant in greening than in etiolated tissues. These preliminary results do not, however, allow us to determine if the cloned 4HPPD is involved only in Tyr degradation or in prenylquinone biosynthesis, or, if it is involved in both processes. Information could also be obtained by the determination of the subcellular localization of this 4HPPD. Although the primary peptide sequence deduced from the Arabidopsis 4HPPD cDNA is longer than that of nonplant 4HPPDs, its analysis did not indicate the presence of a transit peptide. We decided to address this question by overexpressing the complete coding sequence of Arabidopsis 4HPPD in transgenic tobacco using A. tumefaciens transformation.
It is now well documented that transit peptides are able to correctly address the corresponding protein when overexpressed in heterologous systems (Clin and Henry, 1996; Fuks and Schnell, 1997). Thus, if the complete coding sequence possesses an amino- or carboxy-terminal targeting signal, tobacco cells will target the recombinant 4HPPD to the corresponding organelle; if not, the recombinant 4HPPD will remain in the cytosol. The functionality of a heterologous transit peptide was illustrated by overexpressing in tobacco a fusion between an OTP, derived from the coding sequence of the sunflower and maize Rubisco small subunit, and the coding sequence of 4HPPD. Immunodetection carried out by western analysis of chloroplast and total cellular extracts, and by immunocytochemistry on tissue sections, clearly revealed that 4HPPD is not targeted to the chloroplast but remains within the cytosolic compartment. In contrast, when the OTP was fused to the complete sequence, the recombinant 4HPPD targeted the chloroplast, as expected, confirming the functionality of a heterologous transit peptide. This result demonstrated that the coding sequence of the recombinant Arabidopsis 4HPPD did not contain any targeting signal, and therefore it encoded a cytosolic 4HPPD. The presence of a cytosolic form of 4HPPD in Arabidopsis was confirmed by western analyses of subcellular fractions obtained from green Arabidopsis cultured cells.
This cytosolic localization is consistent with our previous results concerning the carrot cell 4HPPD (Lenne et al., 1995; Garcia et al., 1997) and thus seems to be a common feature of plant 4HPPD. This localization is also in agreement with the situation encountered in mammalian liver cells. It has been known for many years that α-ketoisocaproate dioxygenase activity is associated with the cytosolic compartment of liver cells (Sabourin and Bieber, 1981). Recently, it has been demonstrated that this enzyme activity is in fact catalyzed by 4HPPD (Baldwin et al., 1995). By analogy with mammalian liver cells, it could be assumed that plant cytosolic 4HPPD activity is engaged in Tyr degradation.
The situation in plant cells is complicated, however, by the implication of 4HPPD in prenylquinone biosynthesis. This raised the question of whether it is involved only in Tyr degradation or in both Tyr catabolism and prenylquinone biosynthesis. In the present study and in our previous work on carrot 4HPPD (Garcia et al., 1997), we found evidence of only the cytosolic form of 4HPPD. However, all of the succeeding enzymatic steps involved in prenylquinone biosynthesis have been localized on the chloroplast envelope (Soll et al., 1985) so that one would expect the presence of a chloroplastic 4HPPD involved in prenylquinone biosynthesis. In a recent work published during the revision of the present manuscript, Norris et al. (1998) reported that a 17-bp deletion in the structural gene of the Arabidopsis 4HPPD led to an albino phenotype. The corresponding 4HPPD thus seems to be required for prenylquinone biosynthesis. Their data and those of the present study suggest that cytosolic 4HPPD plays a role in both Tyr catabolism and prenylquinone biosynthesis.
The accession number for the nucleotide sequence of the Arabidopsis 4HPPD cDNA is AF047834.
ACKNOWLEDGMENTS
We are grateful to Roland Douce and Dominique Job for invaluable scientific discussions and critical reading of the manuscript, to Anne Roland for help in the buildup of the fusion OTP-4HPPD, to Martine Freyssinet for assistance in tobacco transformation, and to Claudette Job for her help in photograph reproduction.
Abbreviations:
- 4HPP
4-hydroxyphenylpyruvate
- 4HPPD
4HPP dioxygenase
- OTP
optimized transit peptide
Footnotes
This study was conducted under the BIO Avenir program financed by Rhône-Poulenc with a contribution from the Ministère de la Recherche et de l'Enseignement Supérieur.
LITERATURE CITED
- Axelos M, Curie C, Mazzolini L, Bardet C, Lescure B. A protocol for transient gene expression in Arabidopsis thaliana protoplasts isolated from cell suspension cultures. Plant Physiol Biochem. 1992;30:123–128. [Google Scholar]
- Baldwin JE, Crouch NP, Fujishima Y, Lee MH, MacKinnon CH, Pitt JPN, Willis AA. 4-Hydroxyphenylpyruvate dioxygenase appears to display α-ketoisocaproate dioxygenase activity in rat liver. Bioorg Med Chem Lett. 1995;5:1255–1260. [Google Scholar]
- Barta IS, Böger P. Purification and characterization of 4-hydroxyphenylpyruvate dioxygenase from maize. Pestic Sci. 1996;48:109–116. [Google Scholar]
- Chua NH. Electrophoresis analysis of chloroplast proteins. Methods Enzymol. 1980;69:434–436. [Google Scholar]
- Clin K, Henry R. Import and routine of nucleus-encoded chloroplast proteins. Annu Rev Cell Dev Biol. 1996;12:1–26. doi: 10.1146/annurev.cellbio.12.1.1. [DOI] [PubMed] [Google Scholar]
- Denoya CD, Skinner DD, Morgenstern MR. A Streptomyces avermitilis gene encoding a 4-hydroxyphenylpyruvic acid dioxygenase-like protein that directs the production of homogentisic acid and an ochronotic pigment in Escherichia coli. J Bacteriol. 1994;176:5312–5319. doi: 10.1128/jb.176.17.5312-5319.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Durand R, Zenk MH. The homogentisate ring-cleavage pathway in the biosynthesis of acetate-derived naphthoquinones of Droseraceae. Phytochemistry. 1974;13:1483–1492. [Google Scholar]
- Endo F, Awata H, Tanoue A, Ishiguro M, Eda Y, Titani K, Matsuda I. Primary structure deduced from complementary DNA sequence and expression in cultured cells of mammalian 4-hydroxyphenylpyruvic acid dioxygenase: evidence that the enzyme is a homodimer of identical subunit homologous to rat liver-specific alloantigen F. J Biol Chem. 1992;267:24235–24240. [PubMed] [Google Scholar]
- Fernandez-Canon JM, Penalva MA. Molecular characterization of a gene encoding a homogentisate dioxygenase from Aspergillus nidulans and identification of its human and plant homologues. J Biol Chem. 1995;270:21199–21205. doi: 10.1074/jbc.270.36.21199. [DOI] [PubMed] [Google Scholar]
- Fiedler E, Soll J, Schultz G. The formation of homogentisate in the biosynthesis of tocopherol and plastoquinone in spinach chloroplasts. Planta. 1982;155:511–515. doi: 10.1007/BF01607575. [DOI] [PubMed] [Google Scholar]
- Fuks B, Schnell D. Mechanism of protein transport across the chloroplast envelope. Plant Physiol. 1997;114:405–410. doi: 10.1104/pp.114.2.405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garcia I, Rodgers M, Lenne C, Roland A, Saillant A, Matringe M. Subcellular localization and purification of a p-hydroxyphenylpyruvate dioxygenase from cultured carrot cells and characterization of the corresponding cDNA. Biochem J. 1997;325:761–769. doi: 10.1042/bj3250761. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goodwin BL. The biochemical, physiological, and clinical significance of p-hydroxyphenylpyruvate oxidase. In: Goodwin BL, editor. Tyrosine Catabolism. Oxford, UK: Oxford University Press; 1972. pp. 1–94. [Google Scholar]
- Horsh RB, Fry JE, Hoffman NL, Eichholtz D, Rodgers SG, Fraley RT. A simple and general method for transferring genes into plants. Science. 1985;227:1229–1231. doi: 10.1126/science.227.4691.1229. [DOI] [PubMed] [Google Scholar]
- Jefford CW, Cadby PA. Evaluation of models for the mechanism of 4-hydroxyphenylpyruvate dioxygenase. Experimentia. 1981;37:1134–1137. doi: 10.1007/BF01989880. [DOI] [PubMed] [Google Scholar]
- Jensen RA. The shikimate/arogenate pathway: link between carbohydrate metabolism and secondary metabolism. Physiol Plant. 1985;66:164–168. [Google Scholar]
- Kleber-Janke T, Krupinska K. Isolation of cDNA clones for genes showing enhanced expression in barley leaves during dark-induced senescence as well as during senescence under field conditions. Planta. 1997;203:332–340. doi: 10.1007/s004250050199. [DOI] [PubMed] [Google Scholar]
- Lasky M (1978) Protein molecular weight determination using polyacrylamide gradient gels in the presence and absence of sodium dodecyl sulfate. In G Catsimpoolis, ed, Electrophoresis 1978. Elsevier, New York, pp 195–205
- Lebrun M, Leroux B, Sailland A, inventors (1992) Gène chimère pour la transformation des plantes. European patent application, EP 508909, March 4
- Lebrun M, Waksman G, Freyssinet G. Nucleotide sequence of a gene encoding corn ribulose-1,5-bisphosphate carboxylase/oxygenase small subunit (rbcs) Nucleic Acids Res. 1987;15:4360. doi: 10.1093/nar/15.10.4360. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee M-H, Zhang Z-H, MacKinnon CH, Baldwin JE, Crouch NP. The C-terminal of rat 4-hydroxyphenylpyruvate dioxygenase is indispensable for enzyme activity. FEBS Lett. 1996;393:269–272. doi: 10.1016/0014-5793(96)00902-7. [DOI] [PubMed] [Google Scholar]
- Lenne C, Matringe M, Roland A, Sailland A, Pallett KE, Douce R. Partial purification and localization of p-hydroxyphenylpyruvate dioxygenase activity from cultured carrot cells. In: Mathis P, editor. Photosynthesis: From Light to Biosphere, Vol V. Dordrect, The Netherlands: Kluwer Academic Publishers; 1995. pp. 285–288. [Google Scholar]
- Lindblad B, Lindstedt G, Lindstedt S, Rundgren M. Purification and some properties of human 4-hydroxyphenylpyruvate dioxygenase. J Biol Chem. 1977;252:5073–5084. [PubMed] [Google Scholar]
- Lindstedt S, Odelhög B. 4-Hydroxyphenylpyruvate dioxygenase from Pseudomonas. Methods Ezymol. 1987;142:143–148. doi: 10.1016/s0076-6879(87)42022-3. [DOI] [PubMed] [Google Scholar]
- Luscombe BM, Pallett KE, Loubierre P, Millet JC, Melgarejo J, Vrabel TE (1995) RPA 201772, a novel herbicide for broad leaf and grass weeds control in maize and sugar cane. Proceedings of the Brighton Crop Protection Conference on Weeds, Brighton, UK, pp 35–42
- Mandel T, Fleming AJ, Krähenbühl R, Kuhlemeier C. Definition of constitutive gene expression in plants: the translation initiation factor 4A gene as a model. Plant Mol Biol. 1995;29:995–1004. doi: 10.1007/BF00014972. [DOI] [PubMed] [Google Scholar]
- Maniatis T, Fritsch EF, Sambrook J. Molecular Cloning: A Laboratory Manual. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press; 1982. [Google Scholar]
- Norris SR, Barrette TR, DellaPenna D. Genetic dissection of carotenoid synthesis in Arabidopsis defines plastoquinone as an essential component of phytoene desaturation. Plant Cell. 1995;7:2139–2149. doi: 10.1105/tpc.7.12.2139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Norris SR, Xiaohua S, DellaPenna D. Complementation of the Arabidopsis pds1 mutation with the gene encoding p-hydroxyphenylpyruvate dioxygenase. Plant Physiol. 1998;117:1317–1323. doi: 10.1104/pp.117.4.1317. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prisbylla MP, Onisko BC, Seribbs JM, Adams DO, Liu Y, Ellis MK, Hawkes TR, Mutter LC (1993) The novel mechanism of action of the herbicidal triketones. Proceedings of the Brighton Crop Protection Conference on Weeds, Brighton, UK, pp 731–738
- Roche PA, Moorehead TJ, Hamilton GH. Purification and properties of hog liver 4-hydroxyphenylpyruvate dioxygenase. Arch Biochem Biophys. 1982;216:62–73. doi: 10.1016/0003-9861(82)90188-6. [DOI] [PubMed] [Google Scholar]
- Rûetschi U, Dellsen A, Sahlin P, Stenman G, Rymo L, Lindstedt S. Human 4-hydroxyphenylpyruvate dioxygenase, primary structure and chromosomal localization of the gene. Eur J Biochem. 1993;213:1081–1089. doi: 10.1111/j.1432-1033.1993.tb17857.x. [DOI] [PubMed] [Google Scholar]
- Sabourin PJ, Bieber LL. Subcellular distribution and partial characterization of an α-ketoisocaproate oxidase of rat-liver: formation of β-hydroxyisovaleric acid. Arch Biochem Biophys. 1981;206:132–144. doi: 10.1016/0003-9861(81)90074-6. [DOI] [PubMed] [Google Scholar]
- Sailland A, Rolland A, Pallett KE, Matringe M, inventors (1996) Gène de l'hydroxy-phényl pyruvate dioxygénase et obtention de plantes contenant ce gène résistantes aux herbicides. European patent application, WO 9638567, June 2
- Schulz A, Oswald O, Beyer P, Kleinig H. SC-005, a 2-benzoyl-cyclohexane-1,3-dione bleaching herbicide, is a potent inhibitor of the enzyme p-hydroxyphenylpyruvate dioxygenase. FEBS Lett. 1993;318:162–166. doi: 10.1016/0014-5793(93)80013-k. [DOI] [PubMed] [Google Scholar]
- Secor J. Inhibition of barnyardgrass 4-hydroxyphenylpyruvate dioxygenase by sulcotrione. Plant Physiol. 1994;106:1429–1433. doi: 10.1104/pp.106.4.1429. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soll J, Schultz G, Joyard J, Douce R, Block MA. Localization and synthesis of prenylquinones in isolated outer and inner envelope membranes from spinach chloroplasts. Arch Biochem Biophys. 1985;238:290–299. doi: 10.1016/0003-9861(85)90167-5. [DOI] [PubMed] [Google Scholar]
- Thomas PS. Hybridization of denatured RNA and small DNA fragments transferred to nitrocellulose. Proc Natl Acad Sci USA. 1980;77:5201–5205. doi: 10.1073/pnas.77.9.5201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Towbin H, Staehelin T, Gordon J. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets. Procedure and some applications. Proc Natl Acad Sci USA. 1979;76:4350–4354. doi: 10.1073/pnas.76.9.4350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wada GH, Fellman JH, Fujita FS, Roth E. Purification and properties of avian liver p-hydroxyphenylpyruvate hydroxylase. J Biol Chem. 1975;250:6720–6726. [PubMed] [Google Scholar]
- Waksman G, Freyssinet G. Nucleotide sequence of a cDNA encoding the ribulose-1,5-bisphosphate carboxylase/oxygenase from sunflower (Helianthus annuus) Nucleic Acids Res. 1987;15:1328. doi: 10.1093/nar/15.3.1328. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wyckoff EE, Pishko EJ, Kirkland TN, Cole GT. Cloning and expression of a gene encoding a T-cell reactive protein from Coccidioides immitis: homology to 4-hydroxyphenylpyruvate dioxygenase and the mammalian F antigen. Gene. 1995;161:107–111. doi: 10.1016/0378-1119(95)00250-a. [DOI] [PubMed] [Google Scholar]