

Abstract
Defective mRNAs are degraded more rapidly than normal mRNAs in a process called mRNA surveillance. Eukaryotic cells use a variety of mechanisms to detect aberrancies in mRNAs and a variety of enzymes to preferentially degrade them. Recent advances in the field of RNA surveillance have provided new information regarding how cells determine which mRNA species should be subject to destruction and also novel mechanisms by which a cell tags an mRNA once that decision has been reached. In this review we will highlight recent progress in understanding these processes.
Defective mRNAs are preferentially degraded by mRNA surveillance pathways
Although some RNAs such as rRNA and tRNA are extremely stable, mRNAs and regulatory RNAs need to be degraded in a timely manner. The degradation of these RNAs is important to control their proper expression pattern and allow for their rapid up- and downregulation. In addition to degrading normal cellular RNAs at the end of their life span, the RNA decay machinery preferentially degrades aberrant RNAs that fail to function properly. For mRNAs, whose function is to be translated into a protein, this failure to function generally means a failure to be translated properly. The first discovered aspect of mRNA surveillance was the rapid degradation of mRNAs that contain a premature stop codon [1]. In this review we will use a broader definition, namely that mRNA surveillance is the preferential degradation of an mRNA molecule that fails to function properly.
mRNA surveillance pathways are typically discovered by analyzing genetic mutations that introduce a defect into an RNA molecule. However, the mRNAs that are encountered most often under physiologically normal conditions are not mutant mRNAs, but mRNAs that have been improperly processed. For example, if an mRNA fails to be spliced or is spliced incorrectly, it often is subject to mRNA surveillance. In this respect the mRNA surveillance pathways increase the overall fidelity of RNA processing, by degrading the mistakes that are unavoidable in a highly complex process. In this brief review we will focus on some recent studies that illustrate key concepts of mRNA surveillance in eukaryotes. However, other nonfunctional RNAs, including defective tRNAs and rRNAs, are also subject to surveillance and we will refer to those pathways briefly to highlight parallels.
mRNAs with gross abnormalities are rapidly degraded
All RNA polymerase II transcripts are cotranscriptionally modified with a 7mGpppN cap, and most mRNAs are also modified at the 3' end with a poly(A) tail. The absence of these general mRNA features leads to rapid mRNA decay through dedicated surveillance pathways.
Capping of RNA polymerase II transcripts has long been considered a default mechanism that occurs constitutively shortly after transcription initiation. More recently, it has been demonstrated that capping might be regulated by specific transcription factors in mammalian cells [2] and upon nutrient starvation in Saccharomyces cerevisiae (budding yeast) [3]. These findings suggest that eukaryotes must have a pathway to degrade mRNAs that fail to receive a 7mGpppN cap, either because of regulation or errors. Addition of this cap occurs by three sequential enzymatic events (Figure 1): first, the gamma phosphate of the RNA is removed from the 5' triphosphate containing RNA (ppp-RNA) generating a diphosphate RNA intermediate (pp-RNA). Then a guanine nucleotide is attached through a unique 5'-5' linkage resulting in the GpppN capped intermediate (GpppN-RNA). Finally a methyltransferase transfers a methyl group from S-adenosyl methionine (SAM, see Glossary) to the RNA generating the 7mGpppN capped RNA. Until recently, it had been assumed that if capping did not happen, the RNA would be susceptible to 5' exoribonucleases. This assumption, however, creates a conundrum because the known 5' exoribonucleases have a strong preference for 5' monophosphate RNAs and do not readily digest the primary ppp-RNA, or the processing intermediates pp-RNA and Gppp-RNA [4].
Figure 1. Capping and decapping pathways.
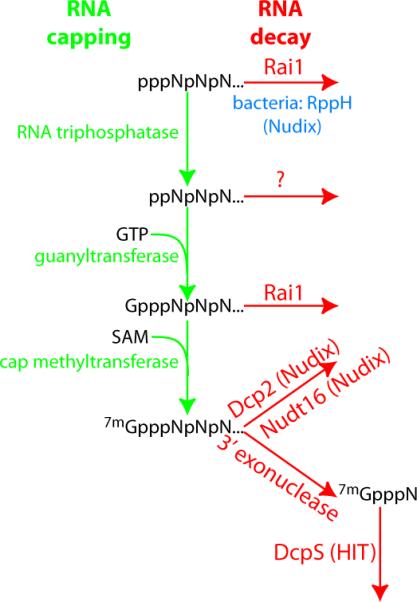
The 5' end of all RNA polymerase II transcripts is modified with a 7mGpppN cap. The required enzymes of the synthesis pathway are depicted in green. Either the fully formed cap or its precursors can be removed by various decapping enzymes (red). Alternatively, after the bulk of the transcript is degraded by an exonuclease, the residual 7mGpppN cap structure can be degraded by the scavenging decapping enzyme (DcpS). For comparison, the bacterial RNA pyrophosphatase RppH (blue) is included.
Initial hints that ppp-RNAs could be targeted for rapid decay came from the X-ray crystal structure of a complex of Rat1 and Rai1 [5]. Rat1 is a nuclear 5' exoribonuclease that is required for a variety of RNA processing pathways, and previous work in S. cerevisiae had identified Rai1 as a protein that binds and activates Rat1 [6]. The crystal structure of the Rat1–Rai1 complex revealed what appeared to be a novel enzymatic active site in Rai1 [5]. Further biochemical characterization showed that Rai1 can remove a pyrophosphate from ppp-RNA, thus converting an uncapped primary transcript into a preferred Rat1 substrate (Figure 1). These data elegantly show that Rai1 has the capacity to initiate the decay of RNA polymerase II transcripts that fail to be capped.
Subsequently Rai1 was demonstrated to not only be active on ppp-RNA, but also on the non-methylated GpppN form [3]. Moreover, on this substrate, Rai1 cleaves between the first and second transcribed nucleotides, releasing the GpppN cap and a pRNA (Figure 1). This is in contrast to the well-documented activity of the canonical decapping enzyme Dcp2, which releases a 7mGpp [7, 8]. Regardless, both enzymes generate the 5'-monophosphate required for either of the Xrn1 or Rat1 exoribonuclease activities [4]. Both the Rai1 ppp-RNA pyrophosphatase and GpppN RNase activity are inactivated by simultaneously mutating the cation binding residues E199 and D201, suggesting that they use identical or overlapping catalytic sites [3].
Mutations in the S. cerevisiae RAI1 gene do not result in any significant defects in mRNA decay under standard growth conditions, but instead enhance stability of mRNAs when cells are deprived of glucose or amino acids [3]. Furthermore, the combination of starvation and rai1Δ led to the accumulation of mRNAs that failed to be precipitated with an anti-7mGpppN antibody, suggesting that cells grown under these conditions accumulate uncapped or incompletely capped RNAs. The molecular mechanism of how starvation affects cap methylation (or capping) remains to be elucidated. One possibility is that cap methylation is actively downregulated in response to starvation. A second scenario is that starvation reduces the availability of the methyl donor SAM. Interestingly, mutations in the cap methyltransferase can be suppressed by exogenously added SAM [9], suggesting that the SAM concentration can affect cap methylation efficiency. Collectively, these findings demonstrate that capping is not as efficient as previously thought and underscore the importance of mRNA surveillance pathways to respond to dynamic changes in the cellular milieu.
Rai1 does not show any structural similarity to previously identified decapping enzymes of the Nudix and HIT motif families, thus increasing the number of distinct protein families that hydrolyze a cap structure to three. Most strikingly, the pppRNA pyrophosphatase reaction carried out by Rai1 is identical to that carried out by the Escherichia coli Nudix family enzyme RppH [10]. The first decapping enzyme discovered was Dcp2, which similar to RppH is a member of the Nudix family [7, 8, 11–13]. In S. cerevisiae, under nonstarvation conditions, deletion of DCP2 results in a slower decay of most mRNAs [11]. However, mice that express reduced amounts of DCP2 are not only phenotypically normal but embryonic fibroblasts generated from mice lacking any detectable DCP2 display nearly normal mRNA decay [14]. These observations were reconciled through the identification of a second mammalian decapping enzyme called NUDT16, which is also a member of the Nudix family and contributes to mRNA stability [15]. Interestingly, the downregulation of NUDT16 in cells lacking detectable DCP2 results in a stabilization of only a subset of mRNAs suggesting that there might be other decapping enzymes yet to be identified [15]. These findings underscore the complexity of mammalian mRNA decay and will likely fuel future searches for other decapping enzymes. This also raises the intriguing possibility that classes of mRNAs initiate decay through the activity of specific decapping enzymes. Candidates for these other proteins include uncharacterized members of the Nudix, HIT, and Rai1 families.
Shortening of the poly(A) tail to an oligo(A) form is the initial step in normal mRNA decay, and triggers subsequent decapping of the mRNA [16, 17]. However, mRNAs that lack a poly(A) tail are not rapidly decapped, but rather represent a distinct target of the mRNA surveillance machinery [18]. The functional consequence of synthesizing a mRNA lacking a poly(A) tail has been best characterized using artificial reporter mRNAs that contain a self-cleaving ribozyme. In S. cerevisiae, a fraction of cleaved unadenylated mRNA accumulates in nuclear foci [19], but the majority of these species are exported and rapidly degraded by the cytoplasmic exosome [18, 20]. This observation is recapitulated in mammals: the insertion of a ribozyme in the 3' untranslated region (UTR) of an mRNA also leads to reduced mRNA levels [21, 22]. These data suggest that the mRNA surveillance pathway for degrading unadenylated mRNAs is conserved between S. cerevisiae and mammals. These reporter experiments likely represent some native circumstances; indeed, self-cleaving mRNAs have been found in a number of eukaryotes [22, 23]. Thus, eukaryotes might have exploited a quality control pathway for the decay of unadenylated mRNAs for the decay of some endogenous transcripts, as has been found for other mRNA surveillance pathways (e.g. [24–26]).
RNA surveillance can also detect internal single nucleotide defects in mRNA
In addition to lacking 5' and 3' modifications, RNAs can be aberrant because they lack some internal modification or have gained an inappropriate modification. This process has been best characterized in tRNA, where the lack of a single methylation can target specific tRNAs to surveillance pathways (see [27] for an excellent review). mRNAs are generally not targeted for internal modification, but can be inadvertently modified by a variety of damaging agents, including reactive oxygen species (ROS), UV light, and alkylating agents. Emerging studies have demonstrated that ROS can cause damage to mRNA [28], and over 20 different nucleotide modifications have been observed in response to ROS [29]. Interestingly, ROS is a critical causative agent in human neurological disorders such as Alzheimer's and Parkinson's disease, and mRNA damage precedes degeneration [30]. Surprisingly, ROS-induced mRNA damage might be restricted to a subset of mRNAs and does not necessarily target those mRNAs that are most abundant [30].
Although little is known about the surveillance response to these damaged mRNAs, we can envision two general models to explain how these modified mRNAs are targeted for decay. First, some protein might specifically recognize aberrancies in mRNA. For example, the translation regulatory protein, Y-box 1 (YB1) preferentially binds 8-hydroxyguanosine containing mRNA, which is a common ROS-induced nucleotide modification [31]. YB1–mRNA binding could be a signal to the mRNA surveillance, leading to a recruitment of the decay machinery. Alternatively, one general cellular mechanism that monitors mRNA integrity could respond to a variety of damaged nucleotides. The immediate consequence of mRNA damage is that it impacts the translation competence by interfering with codon–anticodon pairing. ROS damaged mRNAs are inappropriately shifted to polyribosome fractions, which is often indicative of translational stalling [32], and stalled ribosomes on damaged mRNAs are a known trigger for mRNA surveillance.
One challenge with studying the surveillance of damaged mRNAs is that it is experimentally difficult to damage a specific mRNA at a specific nucleotide in vivo. This hurdle, however, was recently overcome. [33]. Unlike the nonspecific modifications caused by ROS, the pokeweed antiviral protein (PAP) depurinates RNAs with some specificity towards viral RNAs. When PAP is coexpressed with a target RNA derived from brome mosaic virus (BMV) it will specifically bind BMV RNA and elicit depurination of specific adenosine residues resulting in the degradation of the BMV RNA. Using S. cerevisiae mutant strains defective in various mRNA decay pathways and biochemical tools, the authors discovered that ribosomes stall on depurinated mRNAs, resulting in the degradation of the mRNA by the no-go decay pathway [33].
Many mRNA defects are detected by the translation machinery
As previously mentioned, the rapid degradation of mRNAs that have a premature stop codon (nonsense mRNAs) represents the first example of mRNA surveillance [34–36]. Similarly, mRNAs with no in-frame stop codons (nonstop mRNAs) or mRNAs with features that stall a translating ribosome (no-go mRNAs) are also targeted for rapid decay [37–39]. In each of these cases, strong genetic evidence suggests that the aberrancy in the mRNA is recognized indirectly by their effect on translating ribosomes.
In the original description of no-go decay, a stable secondary structure within the coding region was shown to be effective in triggering no-go decay, and this no-go decay was mediated by Hbs1p,Dom34p, and an unknown endonuclease that cleaves the mRNA near the stalled ribosome [39]. Other structural features also trigger mRNA cleavage, including a pseudoknot, rare codons, stop codon, a CGA codon that requires decoding by an A-I wobble basepair to an tRNA, or mRNA depurination [33, 39, 40]. The common link between all of these signals is that they are thought to stall the translating ribosome by interfering with codon-anticodon pairing.
Hbs1p and Dom34p play an important role in no-go decay. These proteins are similar to the translation termination factors eRF3 and eRF1, respectively (Figure 2a). Based on this similarity, it was proposed that Hbs1 and Dom34 bind to the stalled ribosome [39]. Dom34 and Hbs1 indeed bind to stalled ribosomes in a manner that is similar to, but distinct from, translation termination [41–43]. Although Dom34 is homologous to eRF1, it lacks two motifs that are critical for eRF1 function. The GGQ motif of eRF1 is necessary for hydrolyzing the bond between the last tRNA and the translated protein. Instead of hydrolyzing this bond, Dom34 triggers release of the peptide-tRNA complex from the ribosome (Figure 2b) [41, 43]. Dom34 also lacks the NIKS motif from eRF1. Given that the NIKS motif recognizes stop codons, Dom34 is not codon specific, but will act on ribosomes stalled at the sense codon CAA and the stop codon UAA [41]. This work has led to the development of a cohesive model (Figure 2b). When a ribosome is stalled, it is recognized by the Dom34–Hbs1 dimer, with GTP bound to Hbs1. This results in GTP hydrolysis and dissociation of the ribosome into 40S subunits and 60S subunits, followed by release of the mRNA and the nascent peptide covalently bound to the last tRNA from the 40S subunit [39, 41–43].
Figure 2. Recognition of specific ribosomal states by eRF1/DOM34 and eRF3/Hbs1/Ski7.

A. A ribosome with a normal stop codon in the A-site is recognized by eRF1–eRF3-GTP. GTP hydrolysis leads to release of the nascent peptide. B. A ribosome that is stalled at a sense codon in the A-site is recognized by Dom34–Hbs1-GTP. GTP hydrolysis leads to disassembly of the ribosome. Although the Dom34–Hbs1-GDP complex is depicted as being associated with the 60S subunit, this is not known. Instead, it might remain associated with the 40S subunit or be released. C. Genetic evidence suggests that a ribosome that is stalled at the end of an mRNA is recognized by Ski7. D. Termination at premature stop codons is aberrant. eRF1–eRF3-GTP can disassemble ribosomes if their capacity to hydrolyze the tRNA-peptide bond is blocked. This activity might have a role in nonsense-mediated decay. eRF1 and Dom34 are paralogs of each other, as are eRF3, Hbs1 and Ski7.
Although the in vitro data convincingly explain how a stalled ribosome is recognized and resolved, it raises many questions about nonstop mRNA decay, which is triggered when a ribosome is stalled at the 3' end of an mRNA. The recognition of this stalled ribosome requires the Hbs1 paralog Ski7 (Figure 2c) [38]. Nonstop mRNA does not require Dom34, and the Dom34 interacting residues of Hbs1 are not conserved in Ski7 [44, 45]. Furthermore, disassembly of no-go ribosomes requires GTP hydrolysis by Hbs1, but the GTP-binding residues are poorly conserved in Ski7 [38]. Thus, the similarity between Hbs1 and Ski7 suggests that they act similarly, but the differences in requirement for Dom34 and GTP hydrolysis suggest we have more to learn about nonstop mRNA decay.
The in vitro data on the no-go decay pathway also suggest a very speculative model for nonsense-mediated mRNA decay. Nonsense-mediated mRNA decay is triggered at a premature or aberrant stop codon, and requires the termination factors eRF1 and eRF3. Importantly, the process of termination is different at normal and aberrant stop codons [46, 47]. Specifically, in translation extracts, a ribosome terminating at a premature stop codon can readily be detected by the toeprinting assay, whereas a ribosome terminating at a normal stop codon does not give an obvious toeprint [46]. In vivo, ribosomes that have terminated at a premature stop codon can reinitiate translation, while ribosomes that have terminated at a normal stop codon reinitiate poorly [46, 47]. Alhtough this finding suggests that termination at aberrant stop codons is different, the biochemical difference between aberrant termination and normal termination is poorly defined. Perhaps this aberrant termination is similar to the no-go ribosome disassembly in that GTP could be hydrolyzed and the ribosome is split without hydrolysis of the peptide-tRNA bond (Figure 2d). Consistent with this idea, when the peptide-tRNA hydrolyzing GGQ motif of eRF1 undergoes a substitution to AGQ, eRF1 and eRF3 remain capable of recognizing stop codons, but now cause aberrant release of the peptide-tRNA complex at stop codons in vitro [41]. Is it possible that a similar aberrant termination reaction occurs at premature stop codons triggering nonsense-mediated mRNA decay? Although this model lacks supporting data, it predicts features of aberrant termination that should be testable in vivo. In addition, the in vitro systems developed for no-go decay [41–43] might be adaptable to studying nonstop decay and aberrant termination at premature stop codons.
Aberrant RNAs can be tagged with a covalent modification
Prior to the identification of any RNA as being defective, it must be marked as such to allow for its specific decay. A seminal discovery demonstrating this process was that initiator tRNAs lacking a specific methyl group (m1A58) are targeted for rapid degradation by the exosome [48]. In this case, the lack of a methyl group is thought to lead to a subtle defect in the folded state of initiator tRNA, which is recognized by the TRAMP complex. This complex contains a noncanonical poly(A) polymerase, which adds an oligo(A) tail to the 3' end of the tRNA, thus marking it for degradation by the exosome [48–52]. The TRAMP complex appears to add oligo(A) tails to a variety of RNAs, but not to mRNAs. Instead, mRNAs are marked by related enzymes.
Bioinformatic analyses uncovered what appeared to be more noncanonical poly(A) polymerases, but empirical data demonstrated that many of these enzymes prefer uridine rather than adenosine and thus are TUTases (terminal uridine transferases) instead of poly(A) polymerases [53]. The initial observation of oligo(U) addition to mRNAs was in Arabidopsis thaliana where it was found that mRNAs that are cleaved by RISC (RNA-induced silencing complex) are targeted for decay through the addition of 5–10 nontemplated uridines (Figure 3, left panel) [54]. A second example is provided by the metazoan replication-dependent histone mRNAs. These messages are unique because they lack 3' poly(A) tails, and instead end in a stem loop structure [55]. Inhibition of DNA replication causes the unexpected addition of 8–10 nontemplated uridine residues to the 3' end of histone mRNA (Figure 3, middle panel) [56, 57]. Similarly, Cid1-dependent uridylation is important in the normal decay of the Schizosaccharomyces pombe URG1 mRNA [58]. Most recently, short tails of mixed C and U residues have been detected on Aspergillus nidulans mRNAs [59].
Figure 3. The Role of Uridylation in Eliciting Various Types of mRNA Decay.

(i) In Arabidopsis thaliana, microRNA-mediated cleavage of mRNA targets results in an unadenylated but 5' capped product that is uridylated by a yet-to-be identified TUTase. (ii) The metazoan replication-dependent histone mRNAs are uridylated at the conclusion of S phase or under conditions of DNA replication inhibition. Several candidate TUTases have been shown to be involved in this event. (iii) Bulk mRNA turnover in Schizosaccharomyces pombe utilizes the TUTase Cid1, which adds a small number of uridines to the poly(A) tail to stimulate decay. All three types of mRNAs that attain oligo(U) tails may recruit the Lsm1-7 complex, which in turn recruits decapping enzymes including DCP2 and possibly NUDT16. The decapped message is then degraded by the 5' to 3' exonuclease XRN1.
The addition of uridines to mRNAs by the TUTases is reminiscent to the addition of adenosines to the 3' ends of misprocessed nuclear noncoding RNAs by the TRAMP complex. Although the addition of short A rich tails to noncoding RNAs is mostly specific for misprocessed RNAs, it is not yet clear whether oligo(U) tailing targets aberrant mRNAs, or is a part of a more general mRNA decay pathway. The 5' mRNA cleavage product produced by RISC lacks a poly(A) tail and therefore could be viewed as an aberrant mRNA, similar to the ribozyme cleaved mRNAs discussed above. In fact, both the RISC-cleaved mRNAs in Drosophila cells and the ribozyme-cleaved mRNAs in S. cerevisiae are degraded by the exosome [60, 61]. Replication-dependent histone mRNAs are functional when produced during S-phase, but they are not typical mRNAs and might be recognized as aberrant outside of S-phase when they are oligo-uridylated and rapidly degraded [56, 57]. The addition of oligo(U) tails to apparently normal S. pombe and A. nidulans mRNAs [59, 62], may represent an additional example of how mRNA surveillance pathways get usurped during evolution to degrade some specific normal mRNAs, or may reflect that oligo(U) tailing is a normal part of mRNA decay in these fungi and potentially in other organisms.
The addition of uridines to mRNAs or adenosines to misprocessed noncoding RNAs probably has an ancient origin, given that bacteria also add oligo(A) tails to RNAs to initiate their decay. These observations lead to the inherent question of how might A or U nucleotides added to the 3' end of transcripts increase their degradation rate? One possibility is that the oligo(A) or oligo(U) tails provide an unstructured tail accessible to 3' exoribonucleases. Many 3' exoribonucleases inefficiently act on RNAs with a secondary structure at their 3' ends. This would be particularly relevant for histone mRNAs because the stem loop forms a stable complex with the stem loop binding protein (SLBP). Thus, adding an oligo(U) tail could provide a single stranded region for an exonuclease. Similarly, the exosome is thought to require a 30 nt single stranded region on the 3' end of its substrates and the TRAMP complex could increase exosome activity by adding a oligo(A) tail [63]. Likewise, E. coli poly(A) polymerase has long been thought to aid 3' exoribonucleases by adding an unstructured region [64]. However, two lines of evidence cannot be explained by this model. First, many tails are simply too short to provide a significant unstructured tail: high throughput sequencing of TRAMP products revealed that the median oligo(A) tail added by TRAMP was only 3–5 nts long [65], much shorter than the 30 nts the exosome is thought to need. Similarly, it is hard to imagine that adding one or two uridine residues to mRNAs in A. thaliana or S. pombe [54, 58] helps a 3' exonuclease get started. Second and perhaps even more striking, in some cases oligo(U) tails are known to stimulate decapping: elegant in vitro studies demonstrate that the presence of an oligo(U) tail can stimulate decapping of an mRNA substrate with an optimal tail length of ~5–10 uridines [66]. Thus in many cases, the oligo tails that are added are too short to provide a significant unstructured region for 3' exoribonuclease attack but rather might affect decapping.
The observations above can be resolved if oligo(A) and oligo(U) tails are actively recognized by some factor, rather than providing a passive single-stranded tail. The process of post-transcriptionally tagging an RNA molecule destined for degradation is reminiscent of the ubiquitylation of proteins. In this case, it is very clear that ubiquitin serves as a tag, and not an unfolded or unstructured region where proteases can initiate degradation. This raises the question, what are the factors that recognize oligo(A) or oligo(U) tails? A likely candidate for recognizing the oligo(U) tails is the Lsm1-7 complex. The Lsm1-7 complex is thought to specifically bind short U-rich sequences [66] and stimulate decapping of other mRNAs [67, 68]; downregulation of Lsm1 leads to a stabilization of histone mRNAs [56]. Analogously, the bacterial Lsm homolog Hfq binds the poly(A) tails of mRNAs and influences their further polyadenylation and degradation [69–71]. A candidate factor recognizing the short oligo(A) tails present on misprocessed nuclear noncoding RNA is Mtr4, an RNA helicase that acts with the exosome. This suggestion is based on data that shows Mtr4 binds specifically to oligo(A) [72].
Concluding remarks
A sign of healthy scientific progress is that in addition to uncovering answers, more questions are generated. The recent advances in mRNA surveillance we highlighted here exemplify this idea, as they raise many new questions (Box 1). We can envision two ways to recognize aberrant RNA molecules for degradation. In many of the cases of mRNA decay highlighted here, the signal recognized by the surveillance machinery is a stalled ribosome resulting from improper mRNA function. Alternatively, aberrancies could be recognized based solely on structure. The example best demonstrating this is the targeting of incorrectly or incompletely processed mRNA caps in which the overall function does not instigate a signal for decay but rather the improper cap structure. In either case, the notion of mRNA surveillance as a nonspecific cellular `garbage-disposal' is an antiquated oversimplification. Instead, the process of mRNA surveillance is highly regulated, carefully specific, and above all an active process. We expect that advances described here will act as intellectual primers for future studies that will provide us more ammunition for the next brief survey of mRNA surveillance.
Box 1. Outstanding Questions
What is the full complement of enzymes that act on normal or aberrant mRNA cap structures?
How can Rai1 catalyze both pyrophosphates and nuclease reactions using a sing active site?
What is the physiological role of the pyrophosphate activity of Rai1?
How is the disassembly of stalled ribosomes by Dom34 and Hbs1 related to the endonuclease cleavage of the mRNA detected in vivo?
What is the relationship between Dom34/Hbs1 mediated disassembly of stalled ribosomes to nonstop mRNA decay and aberrant termination in nonsense-mediated decay?
Is oligo-uridylation a general mRNA decay pathway or mostly used for aberrant mRNAs?
Do tails provide an unstructured region for 3' exonucleases or a recognition site for a specific factor ?
How does the surveillance machinery can distinguish which target mRNAs receive covalent modifications?
Why do some RNAs receive oligo(A) and others oligo(U)?
How does the decay machinery discern the difference between oligo(A) and poly(A) tails, and those long poly(A) tails that contain additional uridines?
ACKNOWLEDGEMENTS
We thank Allan Jacobson, Aaron Goldstrohm, Phillip Carpenter, the van Hoof lab, and the Wagner lab for insightful comments. Work in the van Hoof lab is supported by grants from NIH (RO1GM069900), NSF (1020739), and the Welch Foundation (AU-1773). Work in the Wagner lab is supported by grants from the NIH (1R21NS0676601 and 5R00GM080447).
Glossary
- Dcp2
a Nudix hydrolase that is the primary decapping enzyme and is found in all eukaryotes.
- Dom34
a protein, related to eRF1, involved in no-go decay. Together with Hbs1 it recognizes ribosomes that have stalled during translation elongation.
- eRF1 and eRF3
eukaryotic translation termination factors.
- Exosome
a protein complex that has both 3' to 5' exoribonuclease and endonuclease activities and is required for a variety of RNA processing and degradation functions.
- Hbs1
a yeast GTPase, related to Ski7 and eRF3, that is involved in no-go decay. Together with Dom34 it recognizes ribosomes that have stalled during translation elongation. The budding yeast, S. cerevisiae has a paralog, Ski7, required for nonstop mRNA degradation. The duplication of Hbs1 and Ski7 is unique to very close relatives of this budding yeast. Other eukaryotes, including some other Saccharomyces species, have only one Hbs1/Ski7 ortholog (often referred to as Hbs1) that presumably performs both functions.
- HIT motif
Histidine triad (HϕHϕHϕϕ; ϕ is a hydrophobic amino acid), found in proteins that contain nucleotide binding capability. This domain is also present in proteins that are capable of hydrolyzing nucleotides. The scavenging decapping enzyme is a member of this family and cleaves 7mGMP from eukaryotic mRNAs.
- Lsm1-7
a complex of seven proteins that are like the Sm proteins found in the spliceosome. The Lsm1-7 complex recognizes short A- or U-rich stretches and stimulates mRNA decapping.
- No-go decay
an mRNA surveillance pathway that rapidly degrades mRNA that contain a stalled ribosome within the coding region. Several mRNA features are known to cause stalling of a ribosome, including stable secondary structure and mRNA damage. In at least some cases of no-go decay, the mRNA is cleaved near the stalled ribosome releasing the message.
- Nonsense-mediated mRNA decay
an mRNA surveillance pathway that rapidly degrades mRNA that contain a premature termination codon (i.e. nonsense codon). This surveillance pathway has been well-characterized in a variety of eukaryotes. In some organisms the mRNA is degraded by decapping and 5' to 3' decay, but in others endonucleolytic cleavage near the site of the terminating ribosome appears important.
- Nonstop mRNA decay
an mRNA surveillance pathway that rapidly degrades mRNA that lack an in-frame stop codon. A ribosome that reaches the end of a nonstop mRNA is thought to stall and recruit Ski7 and the exosome.
- Nudix hydrolases
enzymes that hydrolyze nucleoside diphosphate linked to some other moiety X. Also called MutT-motif proteins after the E. coli enzyme. In the case of the Dcp2, Nud16, and RppH enzymes, the other moiety is as large as a complete RNA. By contrast, MutT hydrolyzes 8-oxo-dGTP and thus, the other moiety is as small as a phosphate.
- NUD16
a Nudix hydrolase that was originally identified as a mammalian U8 small nucleolar RNA (snoRNA) decapping enzyme present in the nucleolus. NUD16 was subsequently found to exist in the cytoplasm and is involved in decapping mRNA in a manner identical to DCP2.
- Rai1
an enzyme capable of releasing pyrophosphate from the 5' end of a primary transcript, or GpppN from a transcript that has completed the first two steps of the capping pathway, but not the methylation step.
- Rat1
the main nuclear 5' to 3' RNA exoribonuclease that degrades or processes a variety of RNAs; also known as Xrn2. Rat 1 is related to Xrn1, and both enzymes are very specific for substrates with a 5' monophosphate.
- ROS (reactive oxygen species)
reactive chemicals that contain oxygen and damage many biomolecules including DNA and RNA.
- RppH
an E. coli Nudix hydrolase that initiates mRNA decay by removing pyrophosphate from the 5' end of a primary transcript.
- SAM (S-adenosyl methionine)
a methyl donor for many reactions within the cell, including synthesis of the cap structure.
- Ski7
a yeast GTPase, related to Hbs1 and eRF3, that is involved in nonstop mRNA degradation. It recognizes ribosomes that have stalled at the end of an mRNA. The budding yeast S. cerevisiae has a paralog, Hbs1, required for no-go decay. The duplication of Hbs1 and Ski7 is unique to very close relatives of this budding yeast. Other eukaryotes, including some other Saccharomyces species, have only one Hbs1/Ski7 ortholog (often referred to as Hbs1) that presumably performs both functions.
- TRAMP complex (Trf4 or 5/Air 1 or2/Mtr4p Polyadenylation complex)
a nuclear complex containing a noncanonical poly(A)polymerase (Trf4 or Trf5 in yeast), the Mtr4 RNA helicase, and a putative RNA binding protein (Air1 or Air2 in yeast). TRAMP adds a short oligo(A) tail to misprocessed noncoding RNAs, facilitating their decay by the exosome.
- TUTase
terminal uridine transferases (poly(U) polymerases), these enzymes are members of the noncanonical poly(A) polymerase family, but use UTP instead of ATP and add short oligo(U) tails to RNA molecules, including mRNAs.
- Xrn1
the main cytoplasmic 5' to 3' RNA exoribonuclease that processively degrades RNA following decapping. It is related to Rat1, and both enzymes are very specific for substrates with a 5' monophosphate.
- YB1
an RNA binding protein that binds mRNAs that contain nucleotides modified by ROS. YB1 might repress their translation or facilitate their degradation.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
REFERENCES CITED
- 1.Pulak R, Anderson P. mRNA surveillance by the Caenorhabditis elegans smg genes. Genes Dev. 1993;7:1885–1897. doi: 10.1101/gad.7.10.1885. [DOI] [PubMed] [Google Scholar]
- 2.Cole MD, Cowling VH. Specific regulation of mRNA cap methylation by the c-Myc and E2F1 transcription factors. Oncogene. 2009;28:1169–1175. doi: 10.1038/onc.2008.463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Jiao X, et al. Identification of a quality-control mechanism for mRNA 5'-end capping. Nature. 2010;467:608–611. doi: 10.1038/nature09338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Stevens A, Maupin MK. A 5' to 3' exoribonuclease of Saccharomyces cerevisiae: size and novel substrate specificity. Arch Biochem Biophys. 1987;252:339–347. doi: 10.1016/0003-9861(87)90040-3. [DOI] [PubMed] [Google Scholar]
- 5.Xiang S, et al. Structure and function of the 5'-->3' exoribonuclease Rat1 and its activating partner Rai1. Nature. 2009;458:784–788. doi: 10.1038/nature07731. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Xue Y, et al. Saccharomyces cerevisiae RAI1 (YGL246c) is homologous to human DOM3Z and encodes a protein that binds the nuclear exoribonuclease Rat1p. Mol Cell Biol. 2000;20:4006–4015. doi: 10.1128/mcb.20.11.4006-4015.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Steiger M, et al. Analysis of recombinant yeast decapping enzyme. RNA. 2003;9:231–237. doi: 10.1261/rna.2151403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Wang Z, et al. The hDcp2 protein is a mammalian mRNA decapping enzyme. Proc Natl Acad Sci U S A. 2002;99:12663–12668. doi: 10.1073/pnas.192445599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Schwer B, et al. Structure-function analysis of yeast mRNA cap methyltransferase and high-copy suppression of conditional mutants by AdoMet synthase and the ubiquitin conjugating enzyme Cdc34p. Genetics. 2000;155:1561–1576. doi: 10.1093/genetics/155.4.1561. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Deana A, et al. The bacterial enzyme RppH triggers messenger RNA degradation by 5' pyrophosphate removal. Nature. 2008;451:355–358. doi: 10.1038/nature06475. [DOI] [PubMed] [Google Scholar]
- 11.Dunckley T, Parker R. The DCP2 protein is required for mRNA decapping in Saccharomyces cerevisiae and contains a functional MutT motif. EMBO J. 1999;18:5411–5422. doi: 10.1093/emboj/18.19.5411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.van Dijk E, et al. Human Dcp2: a catalytically active mRNA decapping enzyme located in specific cytoplasmic structures. EMBO J. 2002;21:6915–6924. doi: 10.1093/emboj/cdf678. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Lykke-Andersen J. Identification of a human decapping complex associated with hUpf proteins in nonsense-mediated decay. Mol Cell Biol. 2002;22:8114–8121. doi: 10.1128/MCB.22.23.8114-8121.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Song MG, et al. Multiple mRNA decapping enzymes in mammalian cells. Molecular cell. 2010;40:423–432. doi: 10.1016/j.molcel.2010.10.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Li Y, et al. Differential utilization of decapping enzymes in mammalian mRNA decay pathways. RNA. 2011;17:419–428. doi: 10.1261/rna.2439811. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Muhlrad D, et al. Deadenylation of the unstable mRNA encoded by the yeast MFA2 gene leads to decapping followed by 5'-->3' digestion of the transcript. Genes Dev. 1994;8:855–866. doi: 10.1101/gad.8.7.855. [DOI] [PubMed] [Google Scholar]
- 17.Decker CJ, Parker R. A turnover pathway for both stable and unstable mRNAs in yeast: evidence for a requirement for deadenylation. Genes Dev. 1993;7:1632–1643. doi: 10.1101/gad.7.8.1632. [DOI] [PubMed] [Google Scholar]
- 18.Meaux S, van Hoof A. Yeast transcripts cleaved by an internal ribozyme provide new insight into the role of the cap and poly(A) tail in translation and mRNA decay. RNA. 2006;12:1323–1337. doi: 10.1261/rna.46306. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Dower K, et al. A synthetic A tail rescues yeast nuclear accumulation of a ribozyme-terminated transcript. RNA. 2004;10:1888–1899. doi: 10.1261/rna.7166704. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Meaux S, et al. Reporter mRNAs cleaved by Rnt1p are exported and degraded in the cytoplasm. Nucleic Acids Res. 2011 doi: 10.1093/nar/gkr627. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Yen L, et al. Exogenous control of mammalian gene expression through modulation of RNA self-cleavage. Nature. 2004;431:471–476. doi: 10.1038/nature02844. [DOI] [PubMed] [Google Scholar]
- 22.Martick M, et al. A discontinuous hammerhead ribozyme embedded in a mammalian messenger RNA. Nature. 2008;454:899–902. doi: 10.1038/nature07117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.de la Pena M, Garcia-Robles I. Ubiquitous presence of the hammerhead ribozyme motif along the tree of life. RNA. 2010;16:1943–1950. doi: 10.1261/rna.2130310. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.He F, et al. Genome-wide analysis of mRNAs regulated by the nonsense-mediated and 5' to 3' mRNA decay pathways in yeast. Mol Cell. 2003;12:1439–1452. doi: 10.1016/s1097-2765(03)00446-5. [DOI] [PubMed] [Google Scholar]
- 25.Rehwinkel J, et al. Nonsense-mediated mRNA decay factors act in concert to regulate common mRNA targets. RNA. 2005;11:1530–1544. doi: 10.1261/rna.2160905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Mendell JT, et al. Nonsense surveillance regulates expression of diverse classes of mammalian transcripts and mutes genomic noise. Nat Genet. 2004;36:1073–1078. doi: 10.1038/ng1429. [DOI] [PubMed] [Google Scholar]
- 27.Phizicky EM, Alfonzo JD. Do all modifications benefit all tRNAs? FEBS letters. 2010;584:265–271. doi: 10.1016/j.febslet.2009.11.049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Kong Q, Lin CL. Oxidative damage to RNA: mechanisms, consequences, and diseases. Cell Mol Life Sci. 2010;67:1817–1829. doi: 10.1007/s00018-010-0277-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Barciszewski J, et al. Some unusual nucleic acid bases are products of hydroxyl radical oxidation of DNA and RNA. Mol Biol Rep. 1999;26:231–238. doi: 10.1023/a:1007058602594. [DOI] [PubMed] [Google Scholar]
- 30.Chang Y, et al. Messenger RNA oxidation occurs early in disease pathogenesis and promotes motor neuron degeneration in ALS. PLoS One. 2008;3:e2849. doi: 10.1371/journal.pone.0002849. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Hayakawa H, et al. Binding capacity of human YB-1 protein for RNA containing 8-oxoguanine. Biochemistry. 2002;41:12739–12744. doi: 10.1021/bi0201872. [DOI] [PubMed] [Google Scholar]
- 32.Shan X, et al. Messenger RNA oxidation is an early event preceding cell death and causes reduced protein expression. FASEB J. 2007;21:2753–2764. doi: 10.1096/fj.07-8200com. [DOI] [PubMed] [Google Scholar]
- 33.Gandhi R, et al. Depurination of Brome mosaic virus RNA3 in vivo results in translation-dependent accelerated degradation of the viral RNA. The Journal of biological chemistry. 2008;283:32218–32228. doi: 10.1074/jbc.M803785200. [DOI] [PubMed] [Google Scholar]
- 34.Losson R, Lacroute F. Interference of nonsense mutations with eukaryotic messenger RNA stability. Proc Natl Acad Sci U S A. 1979;76:5134–5137. doi: 10.1073/pnas.76.10.5134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Maquat LE, et al. Unstable beta-globin mRNA in mRNA-deficient beta o thalassemia. Cell. 1981;27:543–553. doi: 10.1016/0092-8674(81)90396-2. [DOI] [PubMed] [Google Scholar]
- 36.Leeds P, et al. The product of the yeast UPF1 gene is required for rapid turnover of mRNAs containing a premature translational termination codon. Genes Dev. 1991;5:2303–2314. doi: 10.1101/gad.5.12a.2303. [DOI] [PubMed] [Google Scholar]
- 37.Frischmeyer PA, et al. An mRNA surveillance mechanism that eliminates transcripts lacking termination codons. Science. 2002;295:2258–2261. doi: 10.1126/science.1067338. [DOI] [PubMed] [Google Scholar]
- 38.van Hoof A, et al. Exosome-mediated recognition and degradation of mRNAs lacking a termination codon. Science. 2002;295:2262–2264. doi: 10.1126/science.1067272. [DOI] [PubMed] [Google Scholar]
- 39.Doma MK, Parker R. Endonucleolytic cleavage of eukaryotic mRNAs with stalls in translation elongation. Nature. 2006;440:561–564. doi: 10.1038/nature04530. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Letzring DP, et al. Control of translation efficiency in yeast by codonanticodon interactions. RNA. 2010;16:2516–2528. doi: 10.1261/rna.2411710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Shoemaker CJ, et al. Dom34:Hbs1 promotes subunit dissociation and peptidyl-tRNA drop-off to initiate no-go decay. Science. 2010;330:369–372. doi: 10.1126/science.1192430. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Becker T, et al. Structure of the no-go mRNA decay complex Dom34-Hbs1 bound to a stalled 80S ribosome. Nature structural & molecular biology. 2011;18:715–720. doi: 10.1038/nsmb.2057. [DOI] [PubMed] [Google Scholar]
- 43.Pisareva VP, et al. Dissociation by Pelota, Hbs1 and ABCE1 of mammalian vacant 80S ribosomes and stalled elongation complexes. The EMBO journal. 2011;30:1804–1817. doi: 10.1038/emboj.2011.93. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Chen L, et al. Structure of the Dom34-Hbs1 complex and implications for no-go decay. Nature structural & molecular biology. 2010;17:1233–1240. doi: 10.1038/nsmb.1922. [DOI] [PubMed] [Google Scholar]
- 45.Wilson MA, et al. A genomic screen in yeast reveals novel aspects of nonstop mRNA metabolism. Genetics. 2007;177:773–784. doi: 10.1534/genetics.107.073205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Amrani N, et al. A faux 3'-UTR promotes aberrant termination and triggers nonsense-mediated mRNA decay. Nature. 2004;432:112–118. doi: 10.1038/nature03060. [DOI] [PubMed] [Google Scholar]
- 47.Ghosh S, et al. Translational competence of ribosomes released from a premature termination codon is modulated by NMD factors. RNA. 2010;16:1832–1847. doi: 10.1261/rna.1987710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Anderson J, et al. The Gcd10p/Gcd14p complex is the essential two-subunit tRNA(1-methyladenosine) methyltransferase of Saccharomyces cerevisiae. Proceedings of the National Academy of Sciences of the United States of America. 2000;97:5173–5178. doi: 10.1073/pnas.090102597. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Kadaba S, et al. Nuclear surveillance and degradation of hypomodified initiator tRNAMet in S. cerevisiae. Genes Dev. 2004;18:1227–1240. doi: 10.1101/gad.1183804. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Wang X, et al. Degradation of hypomodified tRNA(iMet) in vivo involves RNA-dependent ATPase activity of the DExH helicase Mtr4p. RNA. 2008;14:107–116. doi: 10.1261/rna.808608. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Etheridge RD, et al. Identification and characterization of nuclear non-canonical poly(A) polymerases from Trypanosoma brucei. Mol Biochem Parasitol. 2009;164:66–73. doi: 10.1016/j.molbiopara.2008.11.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.LaCava J, et al. RNA degradation by the exosome is promoted by a nuclear polyadenylation complex. Cell. 2005;121:713–724. doi: 10.1016/j.cell.2005.04.029. [DOI] [PubMed] [Google Scholar]
- 53.Kwak JE, Wickens M. A family of poly(U) polymerases. RNA. 2007;13:860–867. doi: 10.1261/rna.514007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Shen B, Goodman HM. Uridine addition after microRNA-directed cleavage. Science. 2004;306:997. doi: 10.1126/science.1103521. [DOI] [PubMed] [Google Scholar]
- 55.Marzluff WF, et al. Metabolism and regulation of canonical histone mRNAs: life without a poly(A) tail. Nat Rev Genet. 2008;9:843–854. doi: 10.1038/nrg2438. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Mullen TE, Marzluff WF. Degradation of histone mRNA requires oligouridylation followed by decapping and simultaneous degradation of the mRNA both 5' to 3' and 3' to 5'. Genes & development. 2008;22:50–65. doi: 10.1101/gad.1622708. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Schmidt MJ, et al. The human cytoplasmic RNA terminal U-transferase ZCCHC11 targets histone mRNAs for degradation. RNA. 2011;17:39–44. doi: 10.1261/rna.2252511. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Rissland OS, Norbury CJ. Decapping is preceded by 3' uridylation in a novel pathway of bulk mRNA turnover. Nat Struct Mol Biol. 2009;16:616–623. doi: 10.1038/nsmb.1601. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Morozov IY, et al. CUCU modification of mRNA promotes decapping and transcript degradation in Aspergillus nidulans. Molecular and cellular biology. 2010;30:460–469. doi: 10.1128/MCB.00997-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Orban TI, Izaurralde E. Decay of mRNAs targeted by RISC requires XRN1, the Ski complex, and the exosome. RNA. 2005;11:459–469. doi: 10.1261/rna.7231505. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Meaux S, et al. Nonsense-mediated mRNA decay in yeast does not require PAB1 or a poly(A) tail. Mol Cell. 2008;29:134–140. doi: 10.1016/j.molcel.2007.10.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Rissland OS, Norbury CJ. Decapping is preceded by 3' uridylation in a novel pathway of bulk mRNA turnover. Nature structural & molecular biology. 2009;16:616–623. doi: 10.1038/nsmb.1601. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Bonneau F, et al. The yeast exosome functions as a macromolecular cage to channel RNA substrates for degradation. Cell. 2009;139:547–559. doi: 10.1016/j.cell.2009.08.042. [DOI] [PubMed] [Google Scholar]
- 64.Cohen SN. Surprises at the 3' end of prokaryotic RNA. Cell. 1995;80:829–832. doi: 10.1016/0092-8674(95)90284-8. [DOI] [PubMed] [Google Scholar]
- 65.Wlotzka W, et al. The nuclear RNA polymerase II surveillance system targets polymerase III transcripts. The EMBO journal. 2011;30:1790–1803. doi: 10.1038/emboj.2011.97. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Song MG, Kiledjian M. 3' Terminal oligo U-tract-mediated stimulation of decapping. RNA. 2007;13:2356–2365. doi: 10.1261/rna.765807. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Tharun S, et al. Mutations in the Saccharomyces cerevisiae LSM1 gene that affect mRNA decapping and 3' end protection. Genetics. 2005;170:33–46. doi: 10.1534/genetics.104.034322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Boeck R, et al. Capped mRNA degradation intermediates accumulate in the yeast spb8-2 mutant. Mol Cell Biol. 1998;18:5062–5072. doi: 10.1128/mcb.18.9.5062. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Folichon M, et al. The poly(A) binding protein Hfq protects RNA from RNase E and exoribonucleolytic degradation. Nucleic acids research. 2003;31:7302–7310. doi: 10.1093/nar/gkg915. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Hajnsdorf E, Regnier P. E. coli RpsO mRNA decay: RNase E processing at the beginning of the coding sequence stimulates poly(A)-dependent degradation of the mRNA. Journal of molecular biology. 1999;286:1033–1043. doi: 10.1006/jmbi.1999.2547. [DOI] [PubMed] [Google Scholar]
- 71.Mohanty BK, et al. The Sm-like protein Hfq regulates polyadenylation dependent mRNA decay in Escherichia coli. Molecular microbiology. 2004;54:905–920. doi: 10.1111/j.1365-2958.2004.04337.x. [DOI] [PubMed] [Google Scholar]
- 72.Bernstein J, et al. Unique properties of the Mtr4p-poly(A) complex suggest a role in substrate targeting. Biochemistry. 2010;49:10357–10370. doi: 10.1021/bi101518x. [DOI] [PMC free article] [PubMed] [Google Scholar]