

Abstract
Candida albicans is the most prevalent human fungal pathogen, with an ability to inhabit diverse host niches and cause disease in both immunocompetent and immunocompromised individuals. C. albicans also readily forms biofilms on indwelling medical devices and mucosal tissues, which serve as an infectious reservoir that is difficult to eradicate, and can lead to lethal systemic infections. Biofilm formation occurs within a complex milieu of host factors and other members of the human microbiota. Polymicrobial interactions will likely dictate the cellular and biochemical composition of the biofilm, as well as influence clinically relevant outcomes such as drug and host resistance and virulence. In this manuscript, we review C. albicans infections in the context of in vivo polymicrobial biofilms and implications for pathogenesis.
Keywords: Candida albicans, Polymicrobial, Biofilm, Infection, Morphogenesis, Infection
Candida albicans monomicrobial and polymicrobial biofilms
Recently there has been a tremendous interest in the role of biofilms on infectious diseases. The NIH estimates that 80% of human infections result from pathogenic biofilms (http://grants.nih.gov/grants/guide/pa-files/PA-99-084.html). Biofilms are heterogeneous communities of microorganisms that attach to abiotic or biotic surfaces and are encased in an extracellular matrix (ECM), forming a complex three-dimensional architecture. Clinically, biofilm infections represent an overwhelming problem due to the highly recalcitrant nature of the embedded microbes, which are resistant to both antimicrobial drugs as well as host defenses. Biofilms can form on medical devices as well as on exposed and mucosal host tissues. As such, biofilms are often polymicrobial in nature, formed from members of the endogenous microbiota as well as nosocomial pathogens. An emerging finding in polymicrobial biofilm research is the presence of both eukaryotic and prokaryotic pathogens. Therefore, these biofilms can be difficult to both diagnose as well as treat, requiring complex multi-drug treatment strategies. Polymicrobial biofilms represent an understudied and clinically relevant health problem, with the potential to serve as an infectious reservoir for a variety of microorganisms, including bacteria and fungi.
The most prevalent fungal biofilm-forming pathogen is Candida albicans, which can cause both superficial and systemic infections. Much of what is known about C. albicans biofilm development has been learned from in vitro monomicrobial biofilm studies. In vitro, biofilm formation can be divided into several growth phases: early, intermediate and mature [1, 2]. During the early phase, yeast cells adhere to an appropriate surface and undergo morphogenesis, which is essential for normal biofilm formation; mutants of C. albicans that are deficient in hyphal growth form less adherent biofilms that lack the normal three-dimensional architecture of wild-type biofilms [3, 4]. It should be noted that in vitro biofilm assays are typically performed using media that promotes morphogenesis, which could skew the importance of the role of morphogenesis in biofilm formation. Indeed, mutants in the transcriptional regulator BCR1, form thin, weakly adherent biofilms but are able to form hyphae [5]. The intermediate phase is characterized by continued hyphal growth and ECM production, which consists of cell wall polysaccharides and protein [6, 7]. Mature biofilms consist of a yeast base, with hyphal elements forming a complex network encased in ECM extending away from the surface. Although it has not been definitively proven in fungal biofilms, it is predicted that a dispersal phase exists [8]. This is supported by evidence that farnesol, a quorum sensing molecule that inhibits germination and biofilm formation, increases in concentration in aging cultures [9]. Farnesol does not inhibit growth of mature biofilms, but does inhibit germination of new blastospores, thus potentially promoting dispersal [10]. In addition, negative regulators of hyphal formation could play a role in biofilm dispersal by promoting yeast growth. Once such regulator, NRG1, was shown to promote dispersal when overexpressed during biofilm growth [11].
Candida biofilms have been studied primarily on abiotic surfaces [8, 12]. Significant attention has been given to Candida biofilm formation of indwelling catheters, which can lead to life-threatening systemic infections [13-15]. More recently, biofilm formation on biotic surfaces has been reported, including both oral and vaginal tissues [16, 17]. The mucosa provides an excellent environment for biofilm development (reviewed in [18]), and biofilm formation phases are similar to that observed in vitro, even in the absence of morphogenesis inducing media [17]. Microbes easily adhere to the mucosal surface and aggregate with other microbes. Because of the heterogeneity of species within polymicrobial biofilms, it has been difficult to assess the relevance and contribution of individual species to pathogenesis and disease. However, with the use of advanced molecular and microscopic techniques to identify and characterize clinically relevant biofilms, it has become clear that interspecies interactions play a role in colonization and infection dynamics, as well as the host response, which will be discussed below. There have been numerous reports concerning the effect of bacterial species on both C. albicans germination and in vitro biofilm formation. Candida-bacterial interactions have been reviewed previously, and both agonistic and antagonistic relationships can occur [19-21]. Therefore, the focus of this review is the clinical significance and in vivo infection models of polymicrobial biofilms with C. albicans.
Systemic infections
An estimated 27-56% of nosocomial C. albicans bloodstream infections are polymicrobial [22, 23]. In a survey of 372 patients with candidemia, the top three most commonly co-isolated bacterial species were Staphylococcus epidermidis, Enterococcus spp., and Staphylococcus aureus [22]. In vitro, using a catheter disc biofilm model, the presence of C. albicans enhanced the growth of S. epidermidis, with extensive association of the bacteria and fungal hyphae [24]. ECM production by S. epidermidis inhibited penetration of fluconazole into the mixed species biofilm; similarly, vancomycin resistance was increased in the presence of C. albicans [24-26]. Whether this increase in drug resistance occurs in vivo in unknown. However, if this is the case, it would help explain why concomitant bacteremia is associated with a poor prognosis despite antimicrobial therapy compared with monomicrobial candidemia [27, 28].
S. aureus does not form biofilms as readily on abiotic surfaces, requiring pre-coating and nutrient supplementation [29]. Although S. aureus forms poor monoculture biofilms in serum, it forms a substantial polymicrobial biofilm in the presence of C. albicans (Figure 1). In terms of architecture, S. aureus is found throughout the biofilm and forms microcolonies on the surface of the biofilm, with C. albicans serving as a scaffold upon which S. aureus adheres [30]. Ability to form hyphae is required for the interaction, as S. aureus preferentially adheres to hyphae as opposed to yeast forms [31, 32]. S. aureus resistance to vancomycin was also enhanced within the polymicrobial biofilm, required viable C. albicans, and was in part mediated by the C. albicans ECM, and required the ability of C. albicans to adhere to the substratum [30, 31]. While no single fungal adhesin was identified as being required for vancomycin resistance, there could be a combination involved as many C. albicans adhesins have overlapping function [33]. In addition, proteomic analysis demonstrated that interspecies interaction influences protein expression, which could help explain modulation of antimicrobial resistance and virulence [32].
Figure 1.
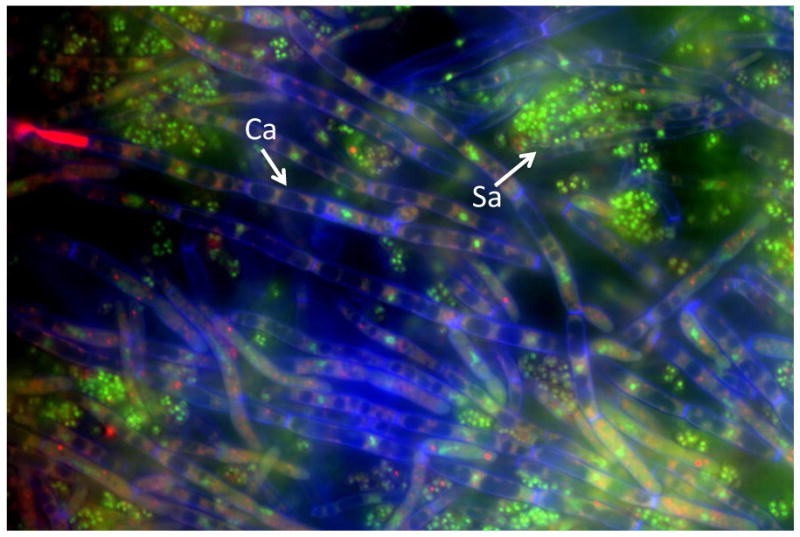
Polymicrobial biofilm of C. albicans and S. aureus. C. albicans SC5314 was grown overnight in Sabouraud dextrose broth (SDB) at 30°C. S. aureus 29523 was grown overnight at 37°C in brain heart infusion (BHI) broth. Both species were washed, counted, and diluted in RPMI-1640 medium + 10% FBS. C. albicans (106 CFU/ml) and S. aureus to (107 CFU/ml) were concurrently added to 96-well tissue culture-treated chamber slides and incubated for 24 h at 37°C. Biofilm formation was monitored by fluorescence microscopy and scanning electron microscopy. To visualize the fungi and bacteria, biofilms were stained with SYTO 9 (green, live bacteria), FUN-1 (intracellular red, live fungal cells), calcofluor white (blue, fungal cell wall). Magnification of 1000×. Abbreviations: Ca, C. albicans hyphae; Sa, S. aureus.
Using an in vivo model of systemic intraperitoneal infection, co-inoculation of C. albicans and S. aureus had a synergistic effect on mortality in mice [34]. Infection with sublethal doses of either species alone resulted in no mortality, but with the same doses, co-infection resulted in 100% mortality; the effect was abrogated when either organism was heat-inactivated. Similar results were observed with non-lethal doses of Serratia marcescens, Streptococcus faecalis, and Escherichia coli during experimental peritonitis [35, 36]. Strains of S. aureus expressing toxic shock syndrome toxin (TSST) or delta toxin had the most dramatic effects on morbitidity and mortality [37, 38]. Culture filtrate of toxin producing strains of S. aureus also mediated this effect, which could be ameliorated with non-steroidal anti-inflammatory drug (NSAID) treatment [38]. Interestingly, when mice were injected with either organism at opposite injection sites (subcutaneous vs. intraperitoneal inoculation), mixed infection was observed at the fungal injection site, but not vice versa [39]. Histological analysis showed that S. aureus was associated throughout areas of fungal growth, interspersed as opposed to at the periphery. This indicates that C. albicans could facilitate S. aureus growth and infection within host tissues and each species could influence gene expression in vivo, resulting in increased virulence and resistance to host defenses as was reported in vitro [32].
Vaginal infections
Vulvovaginal candidiasis (VVC) affects a significant number of women predominantly in their reproductive years [40-42]. An estimated 75% of all women will experience an episode of acute VVC in their lifetime with another 5-10% developing recurrent VVC (RVVC) [40, 41]. Vulvovaginitis involves infections of the vaginal lumen as well as the vulva. Symptoms include burning, itching, soreness, an abnormal discharge, and dyspareunia. Signs include vaginal and vulvar erythema and edema. Acute VVC has several known predisposing factors including antibiotic and oral contraceptive usage, hormone replacement therapy, pregnancy, and uncontrolled diabetes mellitus [40-42]. Antiobiotic use reduces the levels of bacteria that prevent fungal overgrowth. In particular, members of the genus Lactobacillus, which is the most prevalent bacterial group in the vagina, can prevent hyphal growth and exert direct antifungal activity [43, 44]. The antagonistic interactions of vaginal lactobacilli with Candida and potential use in therapeutic applications have been reviewed elsewhere [45, 46].
Vaginal mucosal bacterial biofilms have been previously described for Gardnerella vaginalis, which causes vaginosis [47-49]. However, in VCC and RVVC it was unknown whether C. albicans exist as biofilms on the vaginal mucosa and if so, whether they play a role in the immunopathogenesis. To this end, in vivo and ex vivo murine vaginitis models were recently used to examine mucosal biofilm formation by scanning electron and confocal microscopy [17]. Wild-type C. albicans strains formed biofilms on the vaginal mucosa in vivo and ex vivo as indicated by high fungal burden and microscopic analysis demonstrating typical biofilm architecture and ECM that co-localized with the presence of fungi. In contrast, mutants in a regulator of hyphal formation (efg1/efg1) and biofilm formation (bcr1/bcr1) exhibited weak to no biofilm formation and ECM production in both models despite comparable colonization levels. This raises an interesting question; does the presence of a biofilm determine whether C. albicans behaves as a pathogen and allows the switch from commensalism? In addition, does a biofilm influence the host response? Further, does growth in a biofilm allow other bacterial species to co-colonize and contribute to pathogenesis or resistance? Previous studies have shown that approximately 20-34% of RVVC samples contain vaginal bacterial pathogens such as Streptococcus agalactiae and G. vaginalis [50, 51]. Interestingly, one of the more effective therapies for recurrent and resistant vaginitis is boric acid. While the mechanism of action of this chemical in unknown, it effectively inhibits morphogenesis and biofilm formation in vitro [52].
While C. albicans biofilm formation on the mucosal surface represents biotic biofilm formation, there is the possibility of fungal or polymicrobial biofilm formation on vaginal or intrauterine devices (IUD) serving as a reservoir for various types of infections. A survey of biofilms formed on IUDs confirmed the presence of multiple bacterial pathogens as well as C. albicans [53]. The longer the IUD was in place, the greater the total microbial burden and heterogeneity of species isolated from the device. There is also a significantly higher prevalence of infections such as bacterial vaginosis in IUD users in comparison with users of other contraceptive methods [54, 55]. In addition, there is a higher risk of pelvic inflammatory disease (PID) immediately following IUD insertion, which is caused by several bacterial pathogens including Chlamydia trachomatis and Neisseria gonorrhoeae [56, 57]. Another type of contraception that has recently been introduced is the combined contraceptive vaginal ring (CCVR), which releases ethinyl estradiol and etonogenstrel hormones locally. C. albicans readily adheres to the ring in vitro, and estrogen is known to promote hyphal formation [58, 59]. Clinically vaginitis was reported more often in women using the ring as opposed to oral hormonal contraceptives [58]. Therefore monomicrobial or polymicrobial biofilm formation on these devices might contribute to incidence and recurrence of genital infections.
Skin infection and wound healing
Biofilms contribute to chronicity and delayed healing of most, if not all, chronic wounds [60]. Recently, the contribution of fungal pathogens to mixed species biofilms has been assessed due to the extremely recalcitrant nature of many wounds to antibacterial agents. Using molecular diagnostics, a survey of 915 clinical specimens taken from chronic wounds revealed 23% were positive for fungal species [61]. The most abundant fungi were yeasts in the genus Candida; however, Curvularia, Malessezia, Aureobasidium, Cladosporium, Ulocladium, Engodontium and Trichtophyton were also found to be prevalent components of these polymicrobial infections. Quantification of bacteria versus fungi in these chronic wounds demonstrated that fungi contributed to >50% of the microbial burden in the majority of the wounds. In several cases, identification of fungal pathogens in the chronic wound biofilm led to the addition of an antifungal drug to the treatment regimen and gradual healing of the wound site. This clinical survey demonstrates that the incidence of fungal pathogens in wound biofilm infection is more significant than previously reported. However, to date, there are no reports of in vivo models to investigate fungal or bacterial-fungal biofilms in wounds. Nevertheless, new cost-effective diagnostics have been developed [62], which can aid in identification of underreported fungal pathogens in chronic wounds, thereby promoting targeted therapies and improving healing trajectories.
Oral infections
C. albicans is responsible for several types of oral infections, including oropharyngeal candidiasis (OPC) or thrush, and denture stomatitis (DS). OPC encompasses infections of the hard and soft palate, tongue, buccal mucosa, and floor of the mouth, and can present as reddened patches (erythematous) or white curd-like lesions (pseudomembranous). Although OPC occurs with several immunocompromising conditions, it appears to be much more common in HIV-infected persons than under any other condition [63-66]. The pseudomembranous lesions on oral mucosa consist of blastospores, pseudohyphae, and hyphae attached to underlying epithelium [67]. It has been hypothesized that these plaques are biofilms, however, this concept had not been studied in vivo. Using an immunosuppressed mouse model, researchers were able to induce development of characteristic white lesions on the surface of the tongue after inoculation with C. albicans [16]. Three-dimensional confocal imaging revealed a complex architecture similar to in vitro biofilms with considerable β-glucan being exposed during invasion of the epithelium. This masking and unmasking of β-glucan in vivo is hypothesized to play a role in immunomodulation, because it serves as a pathogen recognition receptor ligand [68]. In particular, β-glucan signals via dectin-1 and TLR2 to promote Th17 cytokine production, which has been proposed to play a protective role against mucosal fungal infections [69-72]. Another interesting observation was the contribution of host components to the biofilm and ECM. Both keratin and polymorphonuclear leukocytes (PMN) aggregates were observed within the ECM, suggesting that the ECM might inhibit migration of effector cells. PMNs exert potent antifungal defenses, and if they become trapped in the ECM, this would limit the ability of these cells to control the fungal infection. The contribution of bacterial species to the fungal biofilm was also investigated. Using fluorescent in situ hybridization (FISH), several groups of bacteria were identified on the basis of 16S RNA using probes that recognize Lactobacillus/Enterococcus spp. and Staphylococcus spp. They noted that mice were generally positive for only one bacterial group and were most often associated with the apical surface of the biofilm [16]. How these bacterial species play a role in disease remains to be determined.
DS is an inflammatory fungal infection affecting approximately 50-75% of otherwise healthy denture wearers and the most common form of oral candidiasis [73-75]. Recently, a new rat model of oral device associated infection was developed to study oral biofilm formed in vivo on dental material [76]. In this model, non-custom fitted oral devices were installed in mice, which were constructed with a 1 mm gap between the palate and device, to allow inoculation. This design does not permit contact between the device and the oral tissue. C. albicans formed biofilms on the device in vivo, and bacteria were found to be associated with the fungi. However, there was no evaluation of palatal biofilm formation and no palatal inflammation [76]. Therefore, the model does not represent a clinically relevant model of denture stomatitis. To investigate the role of biofilms in DS, a novel contemporary rat model was recently developed using a custom fitted denture system composed of both fixed and magnetic removable plates [77]. The denture system was installed against the hard palate of rats without alteration of the dental architecture. The novel design of this denture system (removable portion) allows for longitudinal studies to evaluate the progression of the disease. Biofilm formation was analyzed on the denture and palate via scanning electron and confocal microscopy. Biofilm formation on the denture occurred by week 4 post-inoculation, characterized by the presence of yeast and hyphae coated with ECM. However, on the palate tissue, only blastospore colonization was observed and no clinical evidence of disease was observed. By week 6 post-inoculation, biofilm formation was observed on both the denture and the palate tissue and palatal erythema was evident [77]. This suggests that during DS, C. albicans biofilm formation occurs initially on the denture plate, which in turn seeds the palatal tissue, resulting in mucosal biofilm formation and signs of disease. Microscopic analysis of the infected denture and tissues revealed co-association of bacteria (Figure 2). Therefore, further investigation at the species level is warranted to determine whether these species play a role in disease.
Figure 2.
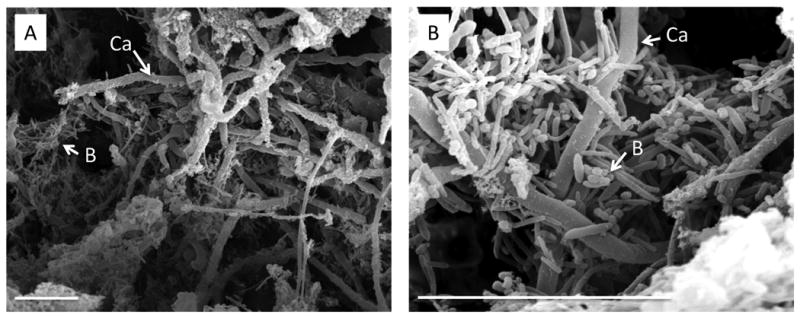
Polymicrobial biofilms formed on dentures from a rat model of C. albicans denture stomatitis. A denture system was installed in Wistar rats that consists of a custom-fitted removable plate that fits over the rat palate and a fixed denture that is anchored to the rear molars. C. albicans 3153A was grown overnight in SDB at 30°C. Rats were inoculated with 25 mg of pelleted yeast on the palate tissue underlying the removable denture. Dentures were removed at 6 weeks post-inoculation and biofilm formation was analyzed by scanning electron microscopy. (a) 1000×, (b) 4000× magnification. Scale bar indicates 50 μM. Abbreviations: Ca, C. albicans hyphae; B, bacteria.
There have been several reports investigating the interaction between C. albicans and oral streptococci. Streptococcus gordonii is a non-pathogenic oral commesal bacterium, but plays an integral role of formation of dental plaque on tooth surfaces, creating an adherent surface amenable to colonization by other oral pathogens. During early polymicrobial biofilm formation in vitro on saliva-coated surfaces, S. gordonii enhances hyphal development, binding to C. albicans hyphae via cell wall proteins SspA and SspB [78]. C. albicans hyphal specific adhesin Als3 is also required for optimal interaction with S. gordonii during biofilm formation [79]. A mutant lacking ALS3 was unable to form a biofilm on a salivary pellicle or on a layer of adherent S. gordonii nor were the bacteria able to adhere to mutant hyphae. Heterologous expression of C. albicans adhesins in Saccharomyces cerevisiae demonstrated a similar requirement for Als3 and Ssp protein in mediating the interaction with S. gordonii. Screening of several fungal adhesins heterologously expressed in S. cereivisiae revealed that specific proteins mediate binding to S. gordonii including: Als3 and Eap1, and to a lesser degree Hwp1 [80]. Therefore, multiple adhesins with overlapping functions work to promote polymicrobial interactions. Repression of C. albicans hyphae and biofilm production by the fungal quorum sensing molecule farnesol was also relieved by S. gordonii, which was mediated in part by the bacterial quorum sensing molecule, autoinducer 2 (AI-2). These results suggest that interactions between C. albicans and S. gordonii involve physical (adherence) and chemical (diffusible) signals that influence the development of biofilms. However, it remains to be investigated in vivo whether S. gordonii can promote C. albicans colonization of the oral cavity.
Lung Infections
While C. albicans is not often associated with pulmonary colonization or infection, there are several reports documenting isolation from clinical samples in the context of bacterial infections, including ventilator associated infection, pneumonia, and cystic fibrosis related infections with Pseudomonas aeruginosa [81, 82]. Interestingly, in vitro studies have pointed to an antagonistic interaction between bacteria and fungi. P. aeruginosa kills yeast, hyphae, and biofilms of C. albicans [83, 84]. This is accomplished in part through production of bacterial phenazine derivatives, which affect cell wall integrity and allows P. aeruginosa to form biofilms on killed hyphae [83, 85, 86]. While the clinical relevance of this relationship is unclear, both organisms have been isolated from lung samples of patients with cystic fibrosis [81, 87]. Although there appears to be an antagonistic relationship in vitro, killing of highly adherent hyphae on a mucosal surface could produce a fungal scaffold upon which P. aeruginosa can form a biofilm, thereby parasitizing C. albicans. In addition, in vivo rat studies have demonstrated that pre-colonization of lung tissue by C. albicans increases rates of P. aeruginosa pneumonia [88]. These results are corroborated by a clinical study of 804 ventilator patients, which showed that pulmonary Candida colonization (which occurred in 26% of patients) is an independent risk factor for Pseudomonas pneumonia [82]. Therefore, this could represent a novel mechanism whereby prior C. albicans mucosal biofilm formation promotes secondary biofilm formation by a bacterial pathogen, leading to disease.
Gastrointestinal Infections
Candida species are normal inhabitants of the gastrointestinal (GI) tract, with colonization rates ranging from 30-70% among healthy adults [89, 90]. Persistence in the GI tract can lead to gastrointestinal candidiasis (defined as infection of the stomach and small and large intestines). More importantly, GI colonization and infection predispose patients to systemic candidiasis due to outgrowth from the GI tract, which is also known as candidiasis of endogenous origin. GI candidiasis is not easily recognized clinically due to the lack of specific symptoms and the lack of the ability to differentiate between pathogenic and commensal C. albicans. However, it is clear from many human studies that two major predisposing factors are immune deficiency and antibiotic therapy [91, 92]. Therefore, both the host immune response and a normal bacterial microbiota are involved in controlling commensalism vs. disease. While it could be assumed that Candida exists as a biofilm in the GI tract, this has not been demonstrated. Gastrointestinal colonization with C. albicans has been associated with alterations in mucosal immunity, promoting atopic responses at other mucosal sites [93, 94]. This was also associated with increases in enteric bacteria, which might have contributed to the alterations in mucosal immune responses. Further studies examining GI tract biofilms with C. albicans could provide insights as to the role of polymicrobial biofilms in disease and/or dissemination from the GI tract.
Conclusions
The development of animal models has been essential in demonstrating the ability of Candida to form polymicrobial biofilms on mucosal tissues in vivo. Advances in microscopic techniques will aid in the assessment of biofilm architecture in situ, as well as identify host and bacterial species that co-associate within the biofilm. It will be especially important to monitor expression profiles of in vivo mucosal biofilms, which could differ significantly from in vivo abiotic biofilms or in vitro biofilms. It has been shown that C. albicans alternatively regulates its gene expression in the oral cavity compared with the GI tract [95]. Therefore, virulence needs to be evaluated in terms of site-specific roles and in terms of colonization vs. biofilm growth in monomicrobial vs. polymicrobial biofilms. It is likely that site specific immunity plays a large role in determining how C. albicans responds within each microenvironment and tailors gene expression to maintain commensal status and/or biofilm growth. In addition, these fungal and bacterial biofilms could serve as a source of systemic infections that exert synergistic effects on morbidity and mortality. The role of polymicrobial biofilms in immune evasion and manipulation of immune responses is of utmost importance for future studies and will likely direct novel therapies that target the multiplicity of species comprising this immune and drug resistant mode of growth.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Bibliography
- 1.Chandra J, et al. Biofilm formation by the fungal pathogen Candida albicans: development, architecture, and drug resistance. J Bacteriol. 2001;183(18):5385–94. doi: 10.1128/JB.183.18.5385-5394.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Ramage G, et al. Characteristics of biofilm formation by Candida albicans. Rev Iberoam Micol. 2001;18(4):163–70. [PubMed] [Google Scholar]
- 3.Nobile CJ, et al. Function of Candida albicans adhesin Hwp1 in biofilm formation. Eukaryot Cell. 2006;5(10):1604–10. doi: 10.1128/EC.00194-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Ramage G, et al. The filamentation pathway controlled by the Efg1 regulator protein is required for normal biofilm formation and development in Candida albicans. FEMS Microbiol Lett. 2002;214(1):95–100. doi: 10.1111/j.1574-6968.2002.tb11330.x. [DOI] [PubMed] [Google Scholar]
- 5.Nobile CJ, et al. Critical role of Bcr1-dependent adhesins in C. albicans biofilm formation in vitro and in vivo. PLoS Pathog. 2006;2(7):e63. doi: 10.1371/journal.ppat.0020063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Baillie GS, Douglas LJ. Matrix polymers of Candida biofilms and their possible role in biofilm resistance to antifungal agents. J Antimicrob Chemother. 2000;46(3):397–403. doi: 10.1093/jac/46.3.397. [DOI] [PubMed] [Google Scholar]
- 7.Nett J, et al. Beta -1,3 glucan as a test for central venous catheter biofilm infection. J Infect Dis. 2007;195(11):1705–12. doi: 10.1086/517522. [DOI] [PubMed] [Google Scholar]
- 8.Blankenship JR, Mitchell AP. How to build a biofilm: a fungal perspective. Curr Opin Microbiol. 2006;9(6):588–94. doi: 10.1016/j.mib.2006.10.003. [DOI] [PubMed] [Google Scholar]
- 9.Ramage G, et al. Inhibition of Candida albicans biofilm formation by farnesol, a quorum-sensing molecule. Appl Environ Microbiol. 2002;68(11):5459–63. doi: 10.1128/AEM.68.11.5459-5463.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Jabra-Rizk MA, et al. Effect of farnesol on Candida dubliniensis biofilm formation and fluconazole resistance. FEMS Yeast Res. 2006;6(7):1063–73. doi: 10.1111/j.1567-1364.2006.00121.x. [DOI] [PubMed] [Google Scholar]
- 11.Uppuluri P, et al. The transcriptional regulator Nrg1p controls Candida albicans biofilm formation and dispersion. Eukaryot Cell. 2010;9(10):1531–7. doi: 10.1128/EC.00111-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Ramage G, Martinez JP, Lopez-Ribot JL. Candida biofilms on implanted biomaterials: a clinically significant problem. FEMS Yeast Res. 2006;6(7):979–86. doi: 10.1111/j.1567-1364.2006.00117.x. [DOI] [PubMed] [Google Scholar]
- 13.Kojic EM, Darouiche RO. Candida infections of medical devices. Clin Microbiol Rev. 2004;17(2):255–67. doi: 10.1128/CMR.17.2.255-267.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Crump JA, Collignon PJ. Intravascular catheter-associated infections. Eur J Clin Microbiol Infect Dis. 2000;19(1):1–8. doi: 10.1007/s100960050001. [DOI] [PubMed] [Google Scholar]
- 15.Dominic RM, Shenoy S, Baliga S. Candida biofilms in medical devices: Evolving trends. Kathmandu Univ Med J (KUMJ) 2007;5(3):431–6. [PubMed] [Google Scholar]
- 16.Dongari-Bagtzoglou A, et al. Characterization of mucosal Candida albicans biofilms. PLoS One. 2009;4(11):e7967. doi: 10.1371/journal.pone.0007967. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Harriott MM, et al. Candida albicans forms biofilms on the vaginal mucosa. Microbiology. 2010;156(Pt 12):3635–44. doi: 10.1099/mic.0.039354-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Natero J, et al., editors. Colonization of Mucosal Surfaces. ASM Press; Chicago: 2005. [Google Scholar]
- 19.Shirtliff ME, Peters BM, Jabra-Rizk MA. Cross-kingdom interactions: Candida albicans and bacteria. FEMS Microbiol Lett. 2009 doi: 10.1111/j.1574-6968.2009.01668.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Wargo MJ, Hogan DA. Fungal--bacterial interactions: a mixed bag of mingling microbes. Curr Opin Microbiol. 2006;9(4):359–64. doi: 10.1016/j.mib.2006.06.001. [DOI] [PubMed] [Google Scholar]
- 21.Thein ZM, et al. Community lifestyle of Candida in mixed biofilms: a mini review. Mycoses. 2009;52(6):467–75. doi: 10.1111/j.1439-0507.2009.01719.x. [DOI] [PubMed] [Google Scholar]
- 22.Klotz SA, et al. Polymicrobial bloodstream infections involving Candida species: analysis of patients and review of the literature. Diagn Microbiol Infect Dis. 2007;59(4):401–6. doi: 10.1016/j.diagmicrobio.2007.07.001. [DOI] [PubMed] [Google Scholar]
- 23.Pulimood S, et al. Polymicrobial candidemia. Diagn Microbiol Infect Dis. 2002;44(4):353–7. doi: 10.1016/s0732-8893(02)00460-1. [DOI] [PubMed] [Google Scholar]
- 24.Adam B, Baillie GS, Douglas LJ. Mixed species biofilms of Candida albicans and Staphylococcus epidermidis. J Med Microbiol. 2002;51(4):344–9. doi: 10.1099/0022-1317-51-4-344. [DOI] [PubMed] [Google Scholar]
- 25.Al-Fattani MA, Douglas LJ. Penetration of Candida biofilms by antifungal agents. Antimicrob Agents Chemother. 2004;48(9):3291–7. doi: 10.1128/AAC.48.9.3291-3297.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Al-Fattani MA, Douglas LJ. Biofilm matrix of Candida albicans and Candida tropicalis: chemical composition and role in drug resistance. J Med Microbiol. 2006;55(Pt 8):999–1008. doi: 10.1099/jmm.0.46569-0. [DOI] [PubMed] [Google Scholar]
- 27.Guerra-Romero L, et al. Polymicrobial fungemia: microbiology, clinical features, and significance. Rev Infect Dis. 1989;11(2):208–12. doi: 10.1093/clinids/11.2.208. [DOI] [PubMed] [Google Scholar]
- 28.Dyess DL, Garrison RN, Fry DE. Candida sepsis. Implications of polymicrobial blood-borne infection. Arch Surg. 1985;120(3):345–8. doi: 10.1001/archsurg.1985.01390270083014. [DOI] [PubMed] [Google Scholar]
- 29.Cassat JE, Lee CY, Smeltzer MS. Investigation of biofilm formation in clinical isolates of Staphylococcus aureus. Methods Mol Biol. 2007;391:127–44. doi: 10.1007/978-1-59745-468-1_10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Harriott MM, Noverr MC. Candida albicans and Staphylococcus aureus form polymicrobial biofilms: effects on antimicrobial resistance. Antimicrob Agents Chemother. 2009;53(9):3914–22. doi: 10.1128/AAC.00657-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Harriott MM, Noverr MC. Ability of Candida albicans mutants to induce Staphylococcus aureus vancomycin resistance during polymicrobial biofilm formation. Antimicrob Agents Chemother. 2010;54(9):3746–55. doi: 10.1128/AAC.00573-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Peters BM, et al. Microbial interactions and differential protein expression in Staphylococcus aureus-Candida albicans dual-species biofilms. FEMS Immunol Med Microbiol. 2010;59(3):493–503. doi: 10.1111/j.1574-695X.2010.00710.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Nobile CJ, et al. Complementary adhesin function in C. albicans biofilm formation. Curr Biol. 2008;18(14):1017–24. doi: 10.1016/j.cub.2008.06.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Carlson E. Synergistic effect of Candida albicans and Staphylococcus aureus on mouse mortality. Infect Immun. 1982;38(3):921–4. doi: 10.1128/iai.38.3.921-924.1982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Carlson E. Enhancement by Candida albicans of Staphylococcus aureus, Serratia marcescens, and Streptococcus faecalis in the establishment of infection in mice. Infect Immun. 1983;39(1):193–7. doi: 10.1128/iai.39.1.193-197.1983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Klaerner HG, et al. Candida albicans and Escherichia coli are synergistic pathogens during experimental microbial peritonitis. J Surg Res. 1997;70(2):161–5. doi: 10.1006/jsre.1997.5110. [DOI] [PubMed] [Google Scholar]
- 37.Carlson E. Effect of strain of Staphylococcus aureus on synergism with Candida albicans resulting in mouse mortality and morbidity. Infect Immun. 1983;42(1):285–92. doi: 10.1128/iai.42.1.285-292.1983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Carlson EC. Synergism of Candida albicans and delta toxin producing Staphylococcus aureus on mouse mortality and morbidity: protection by indomethacin. Zentralbl Bakteriol Mikrobiol Hyg [A] 1988;269(3):377–86. doi: 10.1016/s0176-6724(88)80181-0. [DOI] [PubMed] [Google Scholar]
- 39.Carlson E, Johnson G. Protection by Candida albicans of Staphylococcus aureus in the establishment of dual infection in mice. Infect Immun. 1985;50(3):655–9. doi: 10.1128/iai.50.3.655-659.1985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Sobel JD. Pathogenesis and epidemiology of vulvovaginal candidiasis. Ann NY Acad Sci. 1988;544:547–557. doi: 10.1111/j.1749-6632.1988.tb40450.x. [DOI] [PubMed] [Google Scholar]
- 41.Sobel JD. Pathogenesis and treatment of recurrent vulvovaginal candidiasis. Clin Infect Dis. 1992;14(suppl 1):S148–S153. doi: 10.1093/clinids/14.supplement_1.s148. [DOI] [PubMed] [Google Scholar]
- 42.Kent HL. Epidemiology of vaginitis. Am J Obstet Gynecol. 1991;165:1168–1175. doi: 10.1016/s0002-9378(12)90722-x. [DOI] [PubMed] [Google Scholar]
- 43.Noverr MC, Huffnagle GB. Regulation of Candida albicans morphogenesis by fatty acid metabolites. Infect Immun. 2004;72(11):6206–10. doi: 10.1128/IAI.72.11.6206-6210.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Strus M, et al. The in vitro activity of vaginal Lactobacillus with probiotic properties against Candida. Infect Dis Obstet Gynecol. 2005;13(2):69–75. doi: 10.1080/10647440400028136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Boris S, Barbes C. Role played by lactobacilli in controlling the population of vaginal pathogens. Microbes Infect. 2000;2(5):543–6. doi: 10.1016/s1286-4579(00)00313-0. [DOI] [PubMed] [Google Scholar]
- 46.Falagas ME, Betsi GI, Athanasiou S. Probiotics for prevention of recurrent vulvovaginal candidiasis: a review. J Antimicrob Chemother. 2006;58(2):266–72. doi: 10.1093/jac/dkl246. [DOI] [PubMed] [Google Scholar]
- 47.Swidsinski A, et al. An adherent Gardnerella vaginalis biofilm persists on the vaginal epithelium after standard therapy with oral metronidazole. Am J Obstet Gynecol. 2008;198(1):97, e1–6. doi: 10.1016/j.ajog.2007.06.039. [DOI] [PubMed] [Google Scholar]
- 48.van der Meijden WI, et al. Descriptive light and electron microscopy of normal and clue-cell-positive discharge. Gynecol Obstet Invest. 1988;25(1):47–57. doi: 10.1159/000293745. [DOI] [PubMed] [Google Scholar]
- 49.Scott TG, Curran B, Smyth CJ. Electron microscopy of adhesive interactions between Gardnerella vaginalis and vaginal epithelial cells, McCoy cells and human red blood cells. J Gen Microbiol. 1989;135(3):475–80. doi: 10.1099/00221287-135-3-475. [DOI] [PubMed] [Google Scholar]
- 50.Buchta V, Spacek J. Microbiological findings in patients with recurrent vulvovaginal candidiasis in the Hradec Kralove Faculty Hospital 1995-2002. Ceska Gynekol. 2004;69(1):7–14. [PubMed] [Google Scholar]
- 51.Esim Buyukbayrak E, et al. Diagnosis of vulvovaginitis: comparison of clinical and microbiological diagnosis. Arch Gynecol Obstet. 2010;282(5):515–9. doi: 10.1007/s00404-010-1498-x. [DOI] [PubMed] [Google Scholar]
- 52.De Seta F, et al. Antifungal mechanisms supporting boric acid therapy of Candida vaginitis. J Antimicrob Chemother. 2009;63(2):325–36. doi: 10.1093/jac/dkn486. [DOI] [PubMed] [Google Scholar]
- 53.Pal Z, et al. Biofilm formation on intrauterine devices in relation to duration of use. J Med Microbiol. 2005;54(Pt 12):1199–203. doi: 10.1099/jmm.0.46197-0. [DOI] [PubMed] [Google Scholar]
- 54.Guerreiro D, Gigante MA, Teles LC. Sexually transmitted diseases and reproductive tract infections among contraceptive users. Int J Gynaecol Obstet. 1998;63 1:S167–73. doi: 10.1016/s0020-7292(98)00201-x. [DOI] [PubMed] [Google Scholar]
- 55.Ferraz do Lago R, et al. Follow-up of users of intrauterine device with and without bacterial vaginosis and other cervicovaginal infections. Contraception. 2003;68(2):105–9. doi: 10.1016/s0010-7824(03)00109-4. [DOI] [PubMed] [Google Scholar]
- 56.Grimes DA. Intrauterine devices and pelvic inflammatory disease: recent developments. Contraception. 1987;36(1):97–109. doi: 10.1016/0010-7824(87)90063-1. [DOI] [PubMed] [Google Scholar]
- 57.Farley TM, et al. Intrauterine devices and pelvic inflammatory disease: an international perspective. Lancet. 1992;339(8796):785–8. doi: 10.1016/0140-6736(92)91904-m. [DOI] [PubMed] [Google Scholar]
- 58.Oddsson K, et al. Efficacy and safety of a contraceptive vaginal ring (NuvaRing) compared with a combined oral contraceptive: a 1-year randomized trial. Contraception. 2005;71(3):176–82. doi: 10.1016/j.contraception.2004.09.001. [DOI] [PubMed] [Google Scholar]
- 59.White S, Larsen B. Candida albicans morphogenesis is influenced by estrogen. Cell Mol Life Sci. 1997;53(9):744–9. doi: 10.1007/s000180050094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.James GA, et al. Biofilms in chronic wounds. Wound Repair Regen. 2008;16(1):37–44. doi: 10.1111/j.1524-475X.2007.00321.x. [DOI] [PubMed] [Google Scholar]
- 61.Dowd SE, et al. Survey of fungi and yeast in polymicrobial infections in chronic wounds. J Wound Care. 2011;20(1):40–7. doi: 10.12968/jowc.2011.20.1.40. [DOI] [PubMed] [Google Scholar]
- 62.Leake JL, et al. Identification of yeast in chronic wounds using new pathogen-detection technologies. J Wound Care. 2009;18(3):103–4. 106, 108. doi: 10.12968/jowc.2009.18.3.39810. [DOI] [PubMed] [Google Scholar]
- 63.Greenspan D, et al. Oral mucosal lesions and HIV viral load in the Women's interagency HIV study (WIHS) JAIDS - J Acq Imm Def. 2000;25:44–50. doi: 10.1097/00042560-200009010-00006. [DOI] [PubMed] [Google Scholar]
- 64.Nielsen H, et al. Oral candidiasis and immune status of HIV-infected patients. J Oral Pathol Med. 1994;23:140–143. doi: 10.1111/j.1600-0714.1994.tb01102.x. [DOI] [PubMed] [Google Scholar]
- 65.Sangeorzan JA, et al. Epidemiology of oral candidiasis in HIV infected patients: colonization, infection, treatment, and emergence of fluconazole resistance. Am J Med. 1994;97:339–346. doi: 10.1016/0002-9343(94)90300-x. [DOI] [PubMed] [Google Scholar]
- 66.Schuman P, et al. Oral lesions among women living with or at risk for HIV infection. Am J Med. 1998;104:559–563. doi: 10.1016/s0002-9343(98)00110-7. [DOI] [PubMed] [Google Scholar]
- 67.Allen CM, Beck FM. Differences in mucosal reaction related to Candida albicans isolates. J Oral Pathol. 1987;16(2):89–93. doi: 10.1111/j.1600-0714.1987.tb00693.x. [DOI] [PubMed] [Google Scholar]
- 68.Wheeler RT, et al. Dynamic, morphotype-specific Candida albicans beta-glucan exposure during infection and drug treatment. PLoS Pathog. 2008;4(12):e1000227. doi: 10.1371/journal.ppat.1000227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Netea MG, et al. Immune sensing of Candida albicans requires cooperative recognition of mannans and glucans by lectin and Toll-like receptors. J Clin Invest. 2006;116(6):1642–50. doi: 10.1172/JCI27114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Cheng SC, et al. The dectin-1/inflammasome pathway is responsible for the induction of protective T-helper 17 responses that discriminate between yeasts and hyphae of Candida albicans. J Leukoc Biol. 2011 doi: 10.1189/jlb.1210702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Ferwerda B, et al. Human dectin-1 deficiency and mucocutaneous fungal infections. N Engl J Med. 2009;361(18):1760–7. doi: 10.1056/NEJMoa0901053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Conti HR, et al. Th17 cells and IL-17 receptor signaling are essential for mucosal host defense against oral candidiasis. J Exp Med. 2009;206(2):299–311. doi: 10.1084/jem.20081463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Budtz-Jorgensen E. The role of Candida albicans in the development of stomatitis in denture wearers. Med Hyg (Geneve) 1975;33(1164):1434–5. [PubMed] [Google Scholar]
- 74.Arendorf TM, Walker DM. Denture stomatitis: a review. J Oral Rehabil. 1987;14(3):217–27. doi: 10.1111/j.1365-2842.1987.tb00713.x. [DOI] [PubMed] [Google Scholar]
- 75.Cumming CG, et al. Denture stomatitis in the elderly. Oral Microbiol Immunol. 1990;5(2):82–5. doi: 10.1111/j.1399-302x.1990.tb00232.x. [DOI] [PubMed] [Google Scholar]
- 76.Nett JE, et al. Development and validation of an in vivo Candida albicans biofilm denture model. Infect Immun. 2010;78(9):3650–9. doi: 10.1128/IAI.00480-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Lee H, et al. Fabrication of a multi-applicable removable intraoral denture system for rodent research. J Oral Rehabil. 2011 doi: 10.1111/j.1365-2842.2011.02206.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Bamford CV, et al. Streptococcus gordonii modulates Candida albicans biofilm formation through intergeneric communication. Infect Immun. 2009;77(9):3696–704. doi: 10.1128/IAI.00438-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Silverman RJ, et al. Interaction of Candida albicans cell wall Als3 protein with Streptococcus gordonii SspB adhesin promotes development of mixed-species communities. Infect Immun. 2010;78(11):4644–52. doi: 10.1128/IAI.00685-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Nobbs AH, Vickerman MM, Jenkinson HF. Heterologous expression of Candida albicans cell wall-associated adhesins in Saccharomyces cerevisiae reveals differential specificities in adherence and biofilm formation and in binding oral Streptococcus gordonii. Eukaryot Cell. 2010;9(10):1622–34. doi: 10.1128/EC.00103-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Williamson DR, et al. The relationship between Candida species cultured from the respiratory tract and systemic inflammation in critically ill patients with ventilator-associated pneumonia. Can J Anaesth. 2011;58(3):275–84. doi: 10.1007/s12630-010-9439-5. [DOI] [PubMed] [Google Scholar]
- 82.Azoulay E, et al. Candida colonization of the respiratory tract and subsequent pseudomonas ventilator-associated pneumonia. Chest. 2006;129(1):110–7. doi: 10.1378/chest.129.1.110. [DOI] [PubMed] [Google Scholar]
- 83.Hogan DA, Kolter R. Pseudomonas-Candida interactions: an ecological role for virulence factors. Science. 2002;296(5576):2229–32. doi: 10.1126/science.1070784. [DOI] [PubMed] [Google Scholar]
- 84.Morales DK, et al. Antifungal mechanisms by which a novel Pseudomonas aeruginosa phenazine toxin kills Candida albicans in biofilms. Mol Microbiol. 2010;78(6):1379–92. doi: 10.1111/j.1365-2958.2010.07414.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Kerr JR, et al. Pseudomonas aeruginosa pyocyanin and 1-hydroxyphenazine inhibit fungal growth. J Clin Pathol. 1999;52(5):385–7. doi: 10.1136/jcp.52.5.385. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Brand A, et al. Cell wall glycans and soluble factors determine the interactions between the hyphae of Candida albicans and Pseudomonas aeruginosa. FEMS Microbiol Lett. 2008;287(1):48–55. doi: 10.1111/j.1574-6968.2008.01301.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Leclair LW, Hogan DA. Mixed bacterial-fungal infections in the CF respiratory tract. Med Mycol. 2010;48 1:S125–32. doi: 10.3109/13693786.2010.521522. [DOI] [PubMed] [Google Scholar]
- 88.Roux D, et al. Candida albicans impairs macrophage function and facilitates Pseudomonas aeruginosa pneumonia in rat. Crit Care Med. 2009;37(3):1062–7. doi: 10.1097/CCM.0b013e31819629d2. [DOI] [PubMed] [Google Scholar]
- 89.Kleinegger CL, et al. Frequency, intensity, species, and strains of oral Candida vary as a function of host age. J Clin Microbiol. 1996;34(9):2246–54. doi: 10.1128/jcm.34.9.2246-2254.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Soll DR, et al. Genetic dissimilarity of commensal strains of Candida spp. carried in different anatomical locations of the same healthy women. Journal of Clinical Microbiology. 1991;29:1702–1710. doi: 10.1128/jcm.29.8.1702-1710.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Samonis G, et al. Prospective study of the impact of broad-spectrum antibiotics on the yeast flora of the human gut. Eur J Clin Microbiol. 1994;13(8):665–667. doi: 10.1007/BF01973996. [DOI] [PubMed] [Google Scholar]
- 92.Anaissie EJ, Bodey GP. Fungal infections in patients with cancer. Pharmacotherapy. 1990;10(6 (Pt 3)):164S–169S. [PubMed] [Google Scholar]
- 93.Noverr MC, et al. Role of antibiotics and fungal microbiota in driving pulmonary allergic responses. Infect Immun. 2004;72(9):4996–5003. doi: 10.1128/IAI.72.9.4996-5003.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Noverr MC, et al. Development of allergic airway disease in mice following antibiotic therapy and fungal microbiota increase: role of host genetics, antigen, and interleukin-13. Infect Immun. 2005;73(1):30–8. doi: 10.1128/IAI.73.1.30-38.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.White SJ, et al. Self-regulation of Candida albicans population size during GI colonization. PLoS Pathog. 2007;3(12):e184. doi: 10.1371/journal.ppat.0030184. [DOI] [PMC free article] [PubMed] [Google Scholar]