

Abstract
Previous research had shown that three closely related species of Lysobacter, i.e., Lysobacter antibioticus, Lysobacter capsici, and Lysobacter gummosus, were present in different Rhizoctonia-suppressive soils. However, the population dynamics of these three Lysobacter spp. in different habitats remains unknown. Therefore, a specific primer–probe combination was designed for the combined quantification of these three Lysobacter spp. using TaqMan. Strains of the three target species were efficiently detected with TaqMan, whereas related non-target strains of Lysobacter enzymogenes and Xanthomonas campestris were not or only weakly amplified. Indigenous Lysobacter populations were analyzed in soils of 10 organic farms in the Netherlands during three subsequent years with TaqMan. These soils differed in soil characteristics and crop rotation. Additionally, Lysobacter populations in rhizosphere and bulk soil of different crops on one of these farms were studied. In acid sandy soils low Lysobacter populations were present, whereas pH neutral clay soils contained high populations (respectively, <4.0–5.87 and 6.22–6.95 log gene copy numbers g−1 soil). Clay content, pH and C/N ratio, but not organic matter content in soil, correlated with higher Lysobacter populations. Unexpectedly, different crops did not significantly influence population size of the three Lysobacter spp. and their populations were barely higher in rhizosphere than in bulk soil.
Introduction
The genus Lysobacter is known for its strong lytic activities as well as production of secondary metabolites with antibiotic activity [26]. Several Lysobacter isolates have been described as potential biocontrol agent of a variety of plant pathogenic fungi, oomycetes, and bacteria [7, 9, 10, 14, 25]. The increased interest in this genus was followed by the isolation and description of several new species. Most recently, the presence of Lysobacter spp. in soil was correlated with disease suppression of Rhizoctonia solani [23, 24]. These suppressive soils contained members of three closely related species of Lysobacter, i.e., Lysobacter antibioticus, Lysobacter capsici, and Lysobacter gummosus. However, details about occurrence and population dynamics of Lysobacter spp. remain unknown.
A suitable selective medium is not available for Lysobacter spp. In early studies, Lysobacter spp. were determined as plaque-forming units (PFU) using media with cyanobacteria. Abundance in natural habitats was relatively low (500 PFU cm−3 soil, 400 PFU ml−1 freshwater) [26]. An extensive search for antagonistic bacteria, using an agar medium together with a fungus, showed the presence of antagonistic Lysobacter species in several soils [24]. Population densities of introduced Lysobacter enzymogenes strains were determined on plant roots in biocontrol experiments with plate counts and immunofluorescent colony staining [4, 13] and more recently by real-time PCR detection [19]. In several other studies, reviewed by Hayward et al., only one or few Lysobacter isolates were found in various niches (soil, rhizosphere, compost water, sludge, salamander skin) of different continents (Europe, Asia, and North America) [7], showing the general appearance of Lysobacter but not giving any quantitative information.
Quantitative information about the indigenous Lysobacter spp. in soil is scarce and factors influencing the population density are unknown. Now that Lysobacter spp. are gaining ecological importance, as potential biocontrol agents [7] as well as being involved in natural disease suppression [24], several key questions about the presence and abundance of Lysobacter populations arise. Is their population size influenced by soil factors, such as soil type, pH, texture, or organic matter? What is the influence of crop species, rotation, or agricultural measures? Have Lysobacter spp. preferred niches in soil, i.e., is their population size enhanced in rhizosphere soil in the vicinity of plant roots? A specific detection method to quantify Lysobacter populations in different habitats is required to answer such ecological key questions.
Currently, quantitative molecular detection techniques are increasingly applied to specifically quantify bacterial or fungal strains, species, genera, or larger taxonomic entities [15, 16, 19, 20, 27]. Meanwhile, recent improvements in extraction methods for DNA or RNA and the sufficient removal of inhibitory compounds with easy-to-use commercial kits has also made these techniques suitable for application on complex substrates such as soil.
This paper describes the design and application of a specific primer–probe combination for quantifying the three closely related species L. antibioticus, L. capsici, and L. gummosus simultaneously in one TaqMan system. The detection method was applied to assess the Lysobacter populations in soils of arable fields at different locations within the Netherlands during three subsequent years. These soils differed in soil characteristics and crop rotation. To study the influence of different crops on Lysobacter communities, rhizosphere and bulk soil, samples were taken from different crops on one farm where Lysobacter was known to be present. These samples were subjected to DNA and RNA analyses to compare total and active community sizes.
Methods
Cultivation of and DNA Extraction from Lysobacter Strains
Lysobacter strains were derived from the culture collection of Plant Research International (Table 1). Four type strains were obtained from DSMZ (Braunschweig, Germany) and two strains of L. enzymogenes were kindly supplied by D.Y. Kobayashi (Rutgers University, New Bunswick, NJ, USA) and G. Yuen (University of Nebraska, Lincoln, NE, USA). Genetic affiliation of the three target species is shown in Fig. 1. The strains, stored at −80°C in 10% glycerol, were cultured on nutrient poor agar medium R2A (BD Laboratories, Franklin Lakes, NJ, USA). The inoculated R2A plates were incubated 48 h at 25°C. Single colonies were picked and resuspended in 5 ml 1/10 strength Trypticase Soy Broth (TSB, BD Laboratories) and incubated 2 days at 25°C under rotation at 200 rpm. Genomic DNA was isolated from the cultures using the MasterPure™ Complete DNA Purification Kit (Epicentre Biotechnologies Madison, WI, USA) according to the manufacturer’s instructions. DNA was further purified using Wizard DNA clean-up (Promega, Madison, WI, USA) because of excess polysaccharides. To confirm product integrity and to estimate yield, DNA preparations were visualized by using standard 1.5% (w/v) agarose 0.5× TBE gel electrophoresis with ethidium bromide staining.
Table 1.
Isolates used in this study
| Species | Isolate nr. | EMBL Accession | Origin | Reference |
|---|---|---|---|---|
| Lysobacter antibioticus | 3.2.10 | AM941209 | Bulk soil; Pietersbierum, NL | [24] |
| L. antibioticus | 1.4.11 | AM941210 | Bulk soil; Strijen, NL | [24] |
| L. antibioticus | 4.1.2 | FR822759 | Bulk soil; Marknesse, NL | [24] |
| L. antibioticus | 4A.2.4 | FR822760 | Bulk soil; Marknesse, NL | [24] |
| L. antibioticus | 9.4.19 | AM941219 | Bulk soil; Strijen, NL | [24] |
| L. antibioticus | 11.2.4 | AM941208 | Bulk soil; Marknesse, NL | [24] |
| L. antibioticus | 12.3.1 | AM941206 | Bulk soil; Engwierum, NL | [24] |
| L. antibioticus | 12.4.1 | AM941218 | Bulk soil; Marknesse, NL | [24] |
| L. antibioticus | 66 | FN398325 | Bulk soil; Zwaagdijk, NL | [23] |
| L. antibioticus | 76 | FR822755 | Bulk soil; Zwaagdijk, NL | [23] |
| L. antibioticus | 156 | FN398326 | Potato; Marknesse, NL | [31] |
| L. antibioticus | DSM 2044T | AB019582 | Culture collection | |
| L. capsici | 1.3.3 | FN357195 | Bulk soil; Strijen, NL | [22] |
| L. capsici | 6.2.3 | FN357196 | Bulk soil; Hoensbroek, NL | [22] |
| L. capsici | 10.4.5 | FN357197 | Bulk soil; Marknesse, NL | [22] |
| L. capsici | 55 | FN357198 | Bulk soil; Zwaagdijk, NL | [22] |
| L. capsici | 177 | FN398327 | Potato; Marknesse, NL | [31] |
| L. capsici | YC5194T (DSM 19286T) | EF488749 | Culture collection | |
| L. gummosus | 3.2.11 | FN600120 | Bulk soil; Pietersbierum, NL | [24] |
| L. gummosus | 1A.1.1 | FR822756 | Bulk soil; Engwierum, NL | [24] |
| L. gummosus | 1A.1.10 | FR822757 | Bulk soil; Engwierum, NL | [24] |
| L. gummosus | 1A.1.13 | FR822758 | Bulk soil; Engwierum, NL | [24] |
| L. gummosus | 2.4.7 | AM941212 | Bulk soil; IJzendijke, NL | [24] |
| L. gummosus | 3.3.1 | AM941214 | Bulk soil; Pietersbierum, NL | [24] |
| L. gummosus | 5.1.5 | FR822761 | Bulk soil; Hensbroek, NL | [24] |
| L. gummosus | 10.1.1 | FR822762 | Bulk soil; IJzendijke, NL | [24] |
| L. gummosus | 11.3.5 | AM941213 | Bulk soil; Marknesse, NL | [24] |
| L. gummosus | 13.3.12 | AM941220 | Bulk soil; Pietersbierum, NL | [24] |
| L. gummosus | DSM 9680T | AB161361 | Culture collection | |
| L. enzymogenes | 3.1T8 | AM930380 | Cucumber rhizosphere; NL | [5] |
| L. enzymogenes | 1.1.4 | FN398321 | Bulk soil; Bakel, NL | [19] |
| L. enzymogenes | C3 | AY074793 | Turfgrass leaf | [29] |
| L. enzymogenes | N4.7 | U89956 | Agricultural soil | [29] |
| L. enzymogenes | DSM 2034T | AJ298291 | Culture collection | |
| Xanthomonas campestris pv. campestris | ATCC 33913T | AE012505 | Culture collection |
Figure 1.

Phylogenetic tree constructed using 16S rRNA gene sequences available in GenBank database employing the neighbor-joining method. The three Lysobacter target strains 1.3.3, 3.2.10, and 3.2.11 and the non-target strain 3.1T8 are presented in bold; type strains of Lysobacter and related other species are presented with their strain number and the accession number in parentheses. Numbers at the nodes indicate percentage bootstrap values, >70%, from 1000 resamplings of the data. Scale bar presents sequence difference
Design of Primers and Probe for Real-Time PCR Detection of L. gummosus, L. antibioticus, and L. capsici
Beacon designer 5.0 software (Premier Biosoft, Palo Alto, CA, USA) was used to design the primers and probe for the simultaneous detection of L. gummosus, L. antibioticus, and L. capsici based on their 16S rRNA sequences. L. enzymogenus, a closely related species, was used as non-target. Sequences of four strains of different species derived from the culture collection of Plant Research International, as well as sequences of the corresponding type strains were used for this purpose (Fig. 2). To increase hybridization specificity and enhance the probe discrimination between single nucleotide polymorphisms, a Locked Nucleic Acid (LNA™) dual label probe was developed (P_Lyso_guanx1-5′ TGACATCCACGGAACTTTC 3′, underlined letter is a LNA base), labeled at 5′ end with the fluorescent dye 6-FAM and at 3′ end with the quencher BHQ1. Following the probe design, a selective reverse primer (Rv_Lyso_guanxl-5′ TGCAGCACCTGTCTCAC 3′) and a non-selective forward primer (Fw_Lyso_guanxl-5′ CAACGCGAAGAACCTTACC 3′) were designed using FastPCR V3.7.43 (University of Helsinki, Finland). Figure 2 depicts the primer and the probe sequences developed to detect L. antibioticus, L. capsici, and L. gummosus for real-time PCR. The probe and the primers used in this study were synthesized by Biolegio (Nijmegen, the Netherlands).
Figure 2.

Alignment of 16S rRNA fragment of target species L. antibioticus (La), L. capsici (Lc), L.gummosus (Lg) and a related non-target species L. enzymogenes (Le). For each species the sequence of one isolate from the PRI collection as well as from the corresponding type strain was used. Location of primers (dark grey box) and TaqMan probe (light grey box) are marked
Specificity of the primer–probe combination was tested in silico with Visual OMP (DNA software, Ann Arbor, MI, USA) by comparing the theoretical 89-bp product of the probe and primer combination with sequences retrieved from public databases. In addition, specificity of this primer–probe combination was tested in duplicate with the TaqMan assay on DNA of in total 29 target and six non-target strains (Table 1).
Real-Time PCR Procedure
TaqMan assays to detect L. antibioticus, L. capsici, and L. gummosus were performed in 96-well plates with 3-μl samples in a total volume of 25 μl. Reaction mixtures contained 12.5 μl 2× Premix Ex Taq and 0.5 μl ROX reference Dye (both Takara Bio Inc, Otsu, Japan), 600 nM forward and reverse primer, and 100 nM FAM-labeled probe. Amplification reactions were performed on a 7500 Real-Time PCR system (Applied Biosystems, Foster City, CA, USA). Amplification was carried out 15 s at 95°C and 60 s at 60°C for 40 cycles. Data analysis was carried out with Sequence Detection System software (Applied Biosystems).
Size of the PCR products of several samples was checked on a 2% Agarose gel (Tebu-Bio, Le Perray en Yvelines, France).
Quantification of Lysobacter spp. and Total Bacterial Populations
Standard curves for quantification of Lysobacter spp. were made from genomic DNA of the target strains
L. antibioticus 3.2.10, L. capsici 1.3.3, and L.
gummosus 3.2.11 described in Table 1. Purified DNA (described before) was quantified using a ND-1000
spectrophotometer (Nanodrop Technologies, Wilmington, DE, USA) and the number of
gene copies per microliter was calculated. Using tenfold increment, the standards
concentrations were adjusted from 107 to
101 gene copies per microliter for each species.
Real-time PCR was performed as described in the previous paragraph. The
amplification efficiency (E) was calculated by the formula: 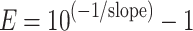 . Additionally, the standard curve from the target isolate
L. gummosus was compared with a dilution
series of DNA extracted from cells of the same isolate, which had been quantified
by plate counting on R2A.
. Additionally, the standard curve from the target isolate
L. gummosus was compared with a dilution
series of DNA extracted from cells of the same isolate, which had been quantified
by plate counting on R2A.
TaqMan reactions were performed on 3 μl DNA preparations from soil samples which were diluted 10 or 50 times to avoid inhibition. A standard curve was included in every TaqMan run. Measurements of negative (undetermined) samples were repeated. Data analysis was carried out with the Applied Biosystems software; the baseline was set automatically. From the obtained cycle threshold (C t) values from the DNA dilution series and the DNA extracts of soil samples, the gene copy numbers of the target organism were calculated per gram of soil sample.
Inhibition of TaqMan reactions with DNA preparations of soil samples (10 or 50 times diluted) was tested in separate TaqMan runs with the gfp-coding gene assay developed as internal control by Klerks et al. [12].
The total bacterial population in soil samples was quantified with real-time PCR (SYBRGreen) following the method of Fierer et al. [3]. Reactions were performed on 3 μl DNA preparations from soil samples which were diluted 100 times.
Recovery of Inoculated L. gummosus Cells from Soil by Plate Counting and TaqMan
L. gummosus was cultured in TSB during 2 days at 25°C. Inoculum (1 × 109 washed cells) or water (control) was added to soil portions (2 g) of a clay (farm A) and a sandy soil (farm F) (three replicates per soil type). Part of the treated soil was frozen (−20°C), part was analyzed directly for colony-forming units. For plate counts, soil was suspended by shaking 1 g soil in a 250 ml Erlenmeyer flasks with 10 g gravel (2–4 mm) and 99 ml 0.1% NaPP (tetra-sodium diphosphate) at 460 rpm for 10 min. Tenfold dilution series were made in 1/4-strength Ringers solution (one tablet Ringers (Oxoid) in 500 ml distilled water, autoclaved at 121°C) and plated in triplicate on R2A plates. Total bacteria and Lysobacter colonies (distinguished by its colony morphology) were enumerated after 2 days incubation at 25°C. DNA was extracted from frozen soil as described by Postma et al. [24] using a Mo Bio Ultra Clean soil DNA isolation kit (Mo Bio Laboratories, BIOzymTC, Landgraaf, the Netherlands) and analyzed by TaqMan as described in the previous paragraph.
Soil Samples of Different Locations
Arable fields of 10 organic farms at different locations within the Netherlands, having different soil characteristics and crop rotations, were sampled in August of 2004, 2005, and 2006 [24]. Sandy and marine clay soils were sampled in different regions of the Netherlands, in three and five different provinces, respectively (Fig. 3). These farms were part of a network with organic farmers [28]. From each field, four independent soil samples of 1.5 l were taken at 3–20 cm depth with minimally 10 m distance between each sampling site. In total, 144 soil samples (31 fields each with four replicates) were subjected to DNA extraction with the Mo Bio Ultra Clean soil DNA isolation kit and stored at −20°C until use.
Figure 3.

Location of sampled farms in different provinces of the Netherlands
Soil Samples of Different Crops from One Farm
Rhizosphere and bulk soil samples were taken from different crops on farm A (Engwierum, the Netherlands) in June 2008. Fields with a grass–clover mixture, red beet, onion, potato, and wheat were sampled in four replicates per field. Individual plants with adhering soil were harvested. Rhizosphere soil was defined as the soil that remained attached to roots after removal from field and gentle shaking. Remaining soil was defined as bulk soil. Subsequently, the rhizosphere soil was carefully removed from the roots with forceps and a spatula. These samples were subjected to DNA extraction.
September 2008, additional rhizosphere and bulk soil samples were taken on farm A in order to analyze DNA as well as RNA of the microbial community. Sampled crops were the grass–clover mixture, red beet, and pumpkin (onion, wheat and potato were already harvested). All soil samples were frozen immediately following sampling at field location with dry ice, transported to the lab, and stored at −80°C until DNA and RNA extraction.
Extraction of DNA and RNA from Soil Samples
Soil genomic DNA was isolated from 0.5 g (wet weight) rhizosphere or bulk soil using the Mo Bio Ultra Clean soil DNA isolation kit. The RNA isolation of soil samples was, from sampling until cDNA synthesis, performed under RNAse-free conditions. The working area and materials reserved for RNA handling were treated with RNAse decontamination solution (RNaseZap®, Ambion). Total RNA was extracted from 0.5 g (wet weight) using the RNeasy Mini Kit (Qiagen) according to the manufacturer’s specifications using 900 μl instead of 450 μl Buffer RTL. Total RNA was resuspended in 50 μl of RNAse-free water. Total RNA was quantified using the NanoDrop ND-1000 Spectrophotometer (Bio-Rad Laboratories Inc., the Netherlands).
RNA samples were reverse-transcribed using Moloney Murine Leukemia Virus reverse transcriptase with low RNAse H activity (200 μ/μl, RevertAid™ First Strand cDNA Synthesis Kit, M-MuLVRT, Fermentas) using random hexaminer primers (0.2 μg/μl) according to the manufacture’s protocol (RevertAid™ First Strand cDNA Synthesis Kit, Fermentas). The reaction mixture (20 μl) contained 5 μl of template RNA.
Populations of L. antibioticus, L. capsici, and L. gummosus in the soil samples were quantified in the DNA and cDNA extracts from rhizosphere and bulk soil samples using TaqMan as described previously.
Statistical Analysis
Statistical analyses were performed with GenStat Release 12.1 (PC/Windows XP) (Rothamsted Experimental Station, Harpenden, UK). Differences among populations of Lysobacter spp. in different soil samples were assessed using analysis of variance (ANOVA). Gene copy numbers of Lysobacter spp. were analyzed after logarithmic transformation. Negative TaqMan reactions were left out of the statistical analysis. After ANOVA, least significant differences were calculated at a significance level of P = 0.05. Multiple regression analyses were performed on mean values per field with generalized linear models to analyze the influence of environmental factors on the Lysobacter population size.
Results
Specificity of the Primer–Probe Combination
All target strains without any exception reacted perfectly with the primer–probe combination; C t values were in all cases between 12 and 18. The non-target of another genus (i.e., Xanthomonas) had a similar C t value (36) as the water control. The strains of the closely related L. enzymogenes species reacted to some extent: C t values were between 30 and 35. A 17-fold higher cycle threshold for this related species implies that only very high population numbers of this related non-target will interact with measurements of the target strains (17 cycles correspond with 2.5 105). Besides, L. enzymogenes was never isolated from one of the soils used in this study [24]
Quantification of Lysobacter spp.
Separate standard curves of the three target Lysobacter strains were evaluated in repeated TaqMan runs. The prepared standard curves were within the range of −3.1 to −3.6, corresponding with a PCR efficiency between 80% and 110%, and a recommended r 2 value of above 0.98 [1, 27]. Thus, all target strains showed appropriate reactions with real-time PCR. The standard curve prepared with L. gummosus showing an efficiency of 98%; slope, −3.38; and r 2, 1.00 was used throughout the study as standard.
Taking sample size and dilution steps into account, lowest measured gene copy numbers of Lysobacter spp. in soil samples with TaqMan were about 104; thus, 4 log g−1 soil is regarded as minimum detection limit.
Comparing the standard curve of L. gummosus with a dilution series of DNA template from freshly cultured cells of the same isolate, showed that the copy numbers were about 1.5–2 times higher than the cell numbers (data not presented). Regression analyses between both dilution series resulted in a significant fit (F < 0.001): log (number of gene copies) = 0.07 + 1.05 log(CFU), accounting for 99.9% of the variance.
Presence of the 89-bp PCR products was confirmed for several different soil samples on agarose gel (results not shown).
Recovery of Inoculated L. gummosus Cells from Soil by Plate Counting and TaqMan
High numbers of L. gummosus cells were added to the soils to be able to enumerate the introduced bacteria among the indigenous populations. Recovered CFU in both soils were not statistically different with the inoculum. With TaqMan, 8.00 and 8.57 log Lysobacter cells g−1 soil were detected in sand and clay soil, respectively, compared to 8.65 log Lysobacter cells which had been introduced (Table 2). Thus, sandy soil showed a lower recovery than marine clay (23% compared to 86%). TaqMan values in the non-inoculated soils were comparable with the indigenous Lysobacter populations presented in Table 3.
Table 2.
Recovery of L. gummosus cells after their introduction in two soil types with plate counts and TaqMan
| Soil type | Inoculum (10 log Lysobacter/g soil) | Plate counts (10log Lysobacter/g soil) | Recovery (%)b | TaqMan (10log Lysobacter/g soil) | Recovery (%) b |
|---|---|---|---|---|---|
| Marine clay (farm A) | – | <3.00 | 6.36 | ||
| Marine clay (farm A) | 8.65 | 8.80 | 152 | 8.57 | 86 |
| Sandy soil (farm F) | – | <3.00 | 5.88 | ||
| Sandy soil (farm F) | 8.65 | 8.69 | 113 | 8.00 | 23 |
| LSDa | 0.25 | 0.10 |
aLeast significant differences (LSD) at P = 0.05; three replicates per treatment
bRecovery (%) is expressed as 100 × numbers detected in soil just after inoculation/numbers in the inoculum
Table 3.
Soil characteristics for bulk soil (3–20 cm depth) collected at arable fields of 10 organic farms in 2004, 2005, and 2006 at different locations within the Netherlands, ordered from low to high Lysobacter spp. populations (gene numbers expressed as log g−1 soil)
| Year | Farm codea | Crop | Soil typeb | Clay particles (<2 μm) (%)b | pH-KClb | Organic matter (%)b | C/N quotientb | Lysobacter gene copies (log g−1 soil)d | Total bacteria gene copies (log g−1 soil) | Lysobacter percentage of total bacteria |
|---|---|---|---|---|---|---|---|---|---|---|
| 2004 | C | Grass–clover | Sandy soil | 2.0 | 5.2 | 10.2 | 26.1 | <4.00 (4/3) | 8.27 | – |
| 2005 | C | Wheat | Sandy soil | 1.5 | 5.1 | 10.8 | 18.4 | 4.71 (2/1) | 8.87 | 0.01 |
| 2005 | F | Grass–clover | Sandy soil | 2.0 | 4.9 | 3.8 | 16.7 | 5.57 | 9.12 | 0.03 |
| 2004 | F | Triticale, grass–clover | Sandy soil | 2.0 | 4.7 | 2.8 | 18.9 | 5.69 (2/0) | 9.11 | 0.04 |
| 2006 | J | Pumpkin | Sandy soil | 2.5 | 4.9 | 1.7 | 23.2 | 5.71 | 8.40 | 0.21 |
| 2006 | F | Wheat, winter radish | Sandy soil | 1.5 | 5.0 | 2.6 | 19.0 | 5.75 (1/0) | 8.43 | 0.21 |
| 2004 | J | Pumpkin | Sandy soil | 1.8 | 5.4 | 1.9 | 21.3 | 5.76 (3/1) | 8.84 | 0.08 |
| 2005 | J | Alfalfa | Sandy soil | 1.8 | 5.4 | 2.4 | 18.1 | 5.87 (2/1) | 8.05 | 0.67 |
| 2006 | A | Red beet | Marine clay | 12.3 | 7.4 | 1.9 | 12.7 | 6.22 | 9.11 | 0.13 |
| 2005 | A | Carrot | Marine clay | 12.0 | 7.5 | 2.3 | 9.2 | 6.25 | 8.21 | 1.09 |
| 2006 | Aa | Grass-clover | Marine clay | 24.3 | 7.4 | 2.7 | 10.1 | 6.27 | 8.86 | 0.26 |
| 2006 | I | Pea | Marine clay | 28.0 | 7.4 | 2.5 | 11.9 | 6.37 | 8.94 | 0.27 |
| 2005 | I | Sugar beet | Marine clay | 27.3 | 7.6 | 2.7 | 10.1 | 6.37 | 8.61 | 0.58 |
| 2006 | D | Potato | Marine clay | 13.8 | 7.4 | 3.0 | 13.3 | 6.44 | 9.30 | 0.14 |
| 2005 | B | Carrot | Marine clay | 13.5 | 7.3 | 2.5 | 9.7 | 6.44 (1/0) | 8.54 | 0.80 |
| 2006 | Ga | Grass-clover | Marine clay | 15.5 | 7.4 | 4.0 | 12.7 | 6.47 | 9.34 | 0.13 |
| 2006 | Ea | Grass-clover | Marine clay | 23.5 | 7.2 | 8.9 | 11.9 | 6.48 | 9.18 | 0.20 |
| 2005 | G | Wheat | Marine clay | 14.0 | 5.9 | 12.0 | 11.7 | 6.51 (1/0) | 8.30 | 1.60 |
| 2006 | E | Cabbage | Marine clay | 26.3 | 7.2 | 7.5 | 11.1 | 6.51 | 9.01 | 0.32 |
| 2006 | Da | Grass-clover | Marine clay | 14.3 | 7.6 | 2.9 | 13.6 | 6.51 | 9.13 | 0.24 |
| 2005 | D | Broccoli | Marine clay | 13.3 | 7.5 | 3.5 | 12.6 | 6.54 | 8.56 | 0.95 |
| 2005 | H | Celery | Marine clay | 20.3 | 7.5 | 3.5 | 9.8 | 6.57 (1/0) | 8.88 | 0.49 |
| 2004 | A | Wheat | Marine clay | 11.5 | 7.4 | 1.7 | 13.6 | 6.57 (1/0) | 8.72 | 0.70 |
| 2004 | B | Cauliflower | Marine clay | 13.8 | 7.3 | 1.6 | 12.4 | 6.58 (1/0) | 8.72 | 0.71 |
| 2005 | E | Wheat | Marine clay | 25.8 | 7.2 | 4.1 | 10.6 | 6.63 | 8.53 | 1.25 |
| 2004 | I | Grass-clover | Marine clay | 29.3 | 7.4 | 1.9 | 15.7 | 6.64 (1/0) | 8.33 | 2.03 |
| 2006 | G | Parsnip, pumpkin | Marine clay | 16.3 | 7.0 | 9.4 | 12.7 | 6.65 | 9.59 | 0.11 |
| 2004 | E | Shallot, winter radish | Marine clay | 27.5 | 7.2 | 6.8 | 11.6 | 6.67 | 8.83 | 0.69 |
| 2004 | D | Onion | Marine clay | 11.8 | 7.5 | 2.9 | 18.4 | 6.79 | 8.69 | 1.26 |
| 2004 | G | Brussels sprouts | Marine clay | 19.0 | 6.8 | 10.3 | 13.4 | 6.95 (1/0) | 9.54 | 0.26 |
| 2004 | H | Cabbage | Marine clay | 20.5 | 7.4 | 2.9 | 12.2 | 6.95 (1/0) | 8.67 | 1.90 |
| LSDc | 2.4 | 0.4 | 1.5 | 2.3 | 0.51 | ns |
a“a” is a field from the same farm but with grass–clover
bData are compiled from previous research [24]
cLeast significant differences (LSD) at P = 0.05; four replicates per treatment; ns no significant differences
dBetween brackets: number of undetectable samples/number of samples with inhibition; 104 is estimated to be the detection limit
Effect of Soil Characteristics on Lysobacter spp. Populations
Populations of the species L. antibioticus, L. capsici, and L. gummosus were quantified using the Lysobacter-specific TaqMan in bulk soil samples collected in 2004, 2005, and 2006 in arable fields of 10 organic farms (Table 3). Sandy soils from farms C, F, and J had the lowest values of Lysobacter spp.; data ranged from undetectable (<4.0) up to 5.87 log gene copies per gram of soil. In clay soils, numbers of Lysobacter were in all cases higher than in sandy soils ranging from 6.22 to 6.95 log gene copies per gram of soil. This difference in Lysobacter populations between clay and sand (32- and 12.5-fold between minimum and maximum values of clay and sand, respectively) is larger than the difference in the recovery of the inoculated cells in both soils (fourfold). Moreover, the part of the total bacteria (real-time, SYBRGreen) that is detected to be Lysobacter was in general much higher in clay than in sandy soil (Table 3). Mean percentage of Lysobacter in sand and clay were 0.18% and 0.70%, respectively.
Several of the sandy soil samples (tenfold diluted) had negative TaqMan reactions. Part of them (six out of 14) showed inhibition of the TaqMan reaction, as tested with the gfp-coding gene method. Of the clay samples, only few (eight out of 92) samples showed a negative reaction, even though their replicates showed high Lysobacter numbers (Table 3, see data between brackets). None of these samples showed inhibition of the TaqMan reaction using the gfp-coding gene method. Thus, the negative reactions in clay soils were not due to failure of the method, but due to low numbers present in the sample. The variation between replicates resulted in an LSD value of 0.51 (negative samples were excluded from ANOVA).
The influence of the different environmental factors on the Lysobacter populations was analyzed using multiple regression analyses. Most important factors contributing to the density of the combined L. antibioticus, L. capsici, and L. gummosus populations were soil type and the sampled field (F < 0.001). No significant differences in Lysobacter densities were recorded across the years of sampling (F = 0.500) and the crop grown in the field (F = 0.898).
Physical, chemical, and biological soil parameters measured from the same soil samples in a previous study [24] were used to analyze the influence of these factors on the Lysobacter populations. Multiple regression analyses between these soil characteristics and Lysobacter populations assessed using TaqMan, showed that these populations strongly correlated with the pH, percent clay particles, and C/N quotient in the soil (all F < 0.001). Sandy soils, with low Lysobacter populations, were low in percent clay, low in pH, and high in C/N quotient (Table 3). Organic matter content in the soils did not correlate with Lysobacter populations (F = 0.236). Also soil suppressiveness against R. solani (data presented in previous study [24]) did not correlate with the Lysobacter populations measured with TaqMan (F = 0.484).
Effect of Specific Crops on Lysobacter spp. Populations
Different crops were sampled from fields of one selected farm (A) with a marine clay soil in June and September 2008. None of these clay samples showed inhibition of the TaqMan reaction. Populations of the species L. antibioticus, L. capsici, and L. gummosus in bulk soil ranged from 6.77 to 7.29 log gene copies per gram of soil in June (Table 4). Only onion bulk soil had a higher value of 7.74 log gene copies per gram. Rhizosphere populations ranged from 7.17 to 7.78 log gene copies per gram of soil in June. On average, populations were higher in the rhizosphere soil compared to bulk soil; 7.58 and 7.26 log gene copies per gram, respectively, soil for all crops (Table 4).
Table 4.
Lysobacter populations (gene copies expressed as log g−1 soil and% Lysobacter between brackets) in rhizosphere and bulk soil of different crops sampled June 2008 on farm A
| Crop | Bulk soil | Rhizosphere soil |
|---|---|---|
| Grassa | 7.29 (0.14)c | 7.17 (0.10) |
| Clovera | – | 7.56 (0.16) |
| Onion | 7.74 (0.40) | 7.66 (0.35) |
| Potato | 6.77 (0.04) | 7.78 (0.26) |
| Red beet | 7.24 (0.14) | 7.57 (0.19) |
| Wheat | 6.99 (0.07) | 7.64 (0.28) |
| Mean of crops | 7.26 (0.13) | 7.58 (0.21) |
| LSDb | 0.20 | |
aGrass and clover were grown together as a mixtures in one field
bLeast significant differences (LSD) at P = 0.05; crop as replicate, each with four subsamples
cPercent Lysobacter = 100 × mean number of Lysobacter/mean number of total bacteria
Bulk soil and rhizosphere soil samples taken in September 2008 were all between 6.60 and 7.19 log gene copies per gram of soil, with no significant difference between rhizosphere and bulk soil (Table 5).
Table 5.
DNA and RNA-based detection of Lysobacter populations (resp. gene or transcript copies expressed as log g−1 soil and% Lysobacter between brackets) in rhizosphere (Rhiz) and bulk soil of different crops sampled in September 2008 on farm A
| Crop | DNA-based analyses | RNA-based analyses | ||
|---|---|---|---|---|
| Bulk | Rhiz | Bulk | Rhiz | |
| Grassa | 7.14 (0.24)c | 7.19 (0.79) | 5.08 (0.03) | 4.46 (0.16) |
| Clovera | – | 7.02 (0.33) | – | 5.89 (0.62) |
| Red beet | 7.01 (0.15) | 6.60 (0.24) | 6.01 (0.21) | 7.78 (0.70) |
| Pumpkin | 6.76 (0.25) | 6.88 (0.06) | 4.91 (0.06) | 6.54 (2.15) |
| Mean of crops | 7.00 (0.23) | 6.92 (0.25) | 5.24 (0.09) | 6.17 (0.63) |
| LSDb | 0.24 | 0.84 | ||
aGrass and clover were grown together as a mixtures in one field
bLeast significant differences (LSD) at P = 0.05; crop as replicate, each with four subsamples
cPercent Lysobacter = 100 × mean number of Lysobacter/mean number of total bacteria
In June, the part of the total bacterial population that was detected to be Lysobacter was 0.13% and 0.21% on average for the different crops in the bulk and rhizosphere soil, respectively. In September, the % Lysobacter was 0.23% and 0.25%, respectively. It seems that the fluctuation of the percentage of Lysobacter is bigger among plant species than between rhizosphere and bulk soil.
Active Part of Lysobacter spp. Populations
RNA was extracted from soil samples of different crops in September 2008 and compared with DNA-based analyses of the L. antibioticus, L. capsici, and L. gummosus community. None of these samples showed inhibition of the TaqMan reaction, as tested with the gfp-coding gene method. RNA-based analyses were on average 5.24 log transcript copies per gram in bulk soil (Table 5). In rhizosphere, large variations were present between the crops: ranging from 4.46 to 7.78 log transcript copies per gram for grass and pumpkin, respectively. The average for all crops in rhizosphere was 6.17. Consistent lower values were observed in RNA (cDNA)-based versus DNA-based Lysobacter TaqMan measurements, except for red beet rhizosphere where the RNA values were higher than those of DNA measurements. Roughly, the activity, i.e., RNA/DNA ratio, of the Lysobacter population was 1–10%.
The active part of Lysobacter within the active total bacterial population was 0.09% and 0.63% on average for the different crops in the bulk and rhizosphere soil, respectively.
Discussion
A specific primer–probe combination was successfully designed to simultaneously quantify three closely related Lysobacter spp. using TaqMan. These species, i.e., L. antibioticus, L. capsici, and L. gummosus, form one cluster within the Lysobacter genus (Fig. 1). They were all three isolated from several Rhizoctonia-suppressive soils in the Netherlands and could strongly inhibit mycelium growth of this fungal plant pathogen [24]. Specificity of this detection system was demonstrated by in silico evaluation of the theoretical 89-bp product of the primer–probe combination, as well as by running TaqMan with strains of the three target species and several strains of the closely related L. enzymogenes, which had a much higher (17-fold) cycle threshold, and a nontarget Xanthomonas which did not show any positive reaction. These four tested species are the main species recovered from soil up to now. Although we had never isolated L. enzymogenes from one of the used soils in the current experiment. Other Lysobacter species have not been detected in soil regularly; they are either water-borne (e.g., Lysobacter brunescens) or described on the basis of one or few isolates.
For quantification of Lysobacter in the soil samples, a standard curve with genomic DNA of L. gummosus strain 3.2.11 was used. Some assumptions (e.g., size of the Lysobacter genome) had to be made for calculating the gene copy numbers in the soil samples. As a consequence, the absolute values are an estimation, but the measured values can be compared with each other to study the influence of several factors in soil on the population dynamics of Lysobacter. Gene copy numbers were 1.5–2 times higher than the values obtained by cell estimations via CFU counts. Recovery of freshly grown Lysobacter cells immediately after their introduction indicated that the detection efficiency of TaqMan in sandy soil and clay soil were approximately 23% and 86%, whereas recovery by plate counts was >100%. The reason for these differences in efficiencies is not clear, but a cell division during the extraction procedure to detect CFU can be an explanation for the high recovery with plate counts.
The influence of soil type on the population size of Lysobacter was remarkable. The three sandy soils contained low Lysobacter populations in all 3 years of sampling, ranging from undetectable (<104) to 5.87 log gene copies per gram bulk soil. In clay soils, the numbers were always higher: 6.22–6.95 log gene copies per gram bulk soil. Even if the difference in recovery between sandy and clay soil as was found with inoculated Lysobacter cells is taken into account, clay soils seem to contain larger Lysobacter populations than sandy soils. Several soil characteristics significantly correlated with the size of the Lysobacter populations, i.e., more Lysobacter gene copies were detected in soils with high pH, high percentage clay, and a lower C/N ratio. However, organic matter content in soil appeared to have no influence on the Lysobacter population size. In our previous study, antagonistic Lysobacter spp. had only been isolated from clay soils and not from sandy soils with low pH [24]. In other studies, Lysobacter spp. was isolated from agricultural soils and sand dunes in Scotland where the pH was not below 6 [26]. This strong effect of soil type on the detected Lysobacter populations is in line with other studies on microbial community composition, where soil type had a larger effect than land use on community composition [2, 6]. Another study showed that the bacterial community was largely driven by the pH and soil texture [17].
It was hypothesized that also the presence and type of crop would influence the Lysobacter population, since in a previous study we isolated about five times more Lysobacter isolates from fields with a grass–clover crop than from fields of the same farm with another crop [24]. However, TaqMan analyzes of the same samples (farm A, D, E, and G in 2006) resulted in similar population sizes of Lysobacter spp. in grass–clover and other crops. Also, new samples taken from fields with different crops, all from farm A, did not show higher Lysobacter populations when grass and clover were grown. Moreover, the vicinity of plant roots often did not enhance Lysobacter populations. Lysobacter populations were higher in rhizosphere than in the bulk soil in only a few cases. For this reason, we speculate that Lysobacter is not a genuine rhizosphere bacterium, such as fluorescent Pseudomonas spp. which can occur in much higher populations in the rhizosphere compared to bulk soil [18, 30]. Lysobacter spp. is known for their capacity to degrade various biomacromolecules; they proliferate on chitin and decompose nematodes, bacteria, yeasts, fungi, etc. [26]. Some isolates of the three target Lysobacter spp. do not proliferate on citric and malic acid and on glucose and maltose [21, 22], which are important components of plant exudates [11]. Consequently, Lysobacter spp. is probably not stimulated directly by plant exudates as is the case for Pseudomonas spp.
The number of Lysobacter gene copies in clay bulk soil samples was on average 7.0 log gene copies per gram soil, whereas the antagonistic Lysobacter isolates had been estimated previously to be 5.5 log CFU g−1 soil (approximately 0.3% of culturable bacteria) [24]. Similarly, Huijsdens et al. [8] demonstrated that quantification by real-time PCR yielded approximately 100-fold higher numbers of Escherichia coli in the human gut than quantification by plate counts. Higher numbers of gene copies than antagonistic isolates can be expected if: (1) not all cells are antagonistic, (2) not all DNA originates from culturable bacterial cells, (3) more than one gene copy is present per bacterium, and (4) bacteria occur clustered in the soil environment. In contrast to soil with freshly inoculated target cells, the indigenous Lysobacter population might contain poorly culturable cells, or even dead intact DNA. Another crucial point would be the presence of Lysobacter cells which are not antagonistic against Rhizoctonia. This would also explain the lack of correlation between Lysobacter gene copy numbers and Rhizoctonia disease suppression.
In addition to the size of the population, bacterial activity will influence beneficial aspects such as disease suppression. For this reason the activity of the Lysobacter population was assessed by comparing DNA and RNA quantities of Lysobacter spp. Although the percentage of RNA/DNA was roughly 1–10%, it did not support the enhanced suppressiveness in grass–clover crops; i.e., the activity of Lysobacter was not higher in grass or clover than in other crops.
One not-yet-discussed result is the large variation which sometimes occurred between replicate samples of 0.5 g soil which had been taken from larger soil samples. In several fields, high populations of Lysobacter were present in three replicates, where the fourth replicate had undetectable populations. This was not due to inhibition in the TaqMan reaction, which had been checked by the gfp assay. Also during isolation of antagonistic Lysobacter isolates [24] large variations in isolate numbers between replicates were found. Probably, cells of Lysobacter appear in clusters in the soil, which makes research on the ecology of Lysobacter more complicated.
This paper describes the first attempt to quantify Lysobacter spp. in different soils and in the vicinity of different crops. Conclusions which can be drawn from these first experiments about the habitat of L. antibioticus, L. capsici, and L. gummosus are: (1) sandy soil with a low pH is unfavorable, (2) soil factors stimulating Lysobacter populations are clay content, pH, and C/N ratio, but not organic matter content, (3) equal and sometimes higher populations are present in rhizosphere compared to bulk soil, (4) the effect of the crop is unclear, since antagonistic isolates were previously found to be higher in grass–clover crops, but total gene copy numbers assessed with TaqMan were not influenced by these crops.
Acknowledgments
We thank the organic farmers for allowing us to take soil samples. Leo van Overbeek, Lia Groenenboom-de Haas, Saskia Burgers, and Els Nijhuis are acknowledged for fruitful advice on the manuscript and on statistical methods, and for preparation of the phylogenetic tree. This research was supported by the Dutch Ministry of Agriculture, Nature and Food Quality in the research program BO-07.
Open Access
This article is distributed under the terms of the Creative Commons Attribution Noncommercial License which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
References
- 1.Anonymous (2008) Definition of minimum performance requirements for analytical methods of GMO testing. European Network of GMO Laboratories (ENGL). Available from: http://gmo-crl.jrc.ec.europa.eu/doc/Min_Perf_Requirements_Analytical_methods.pdf. Accessed on: 13 Oct 2008
- 2.Bossio DA, Scow KM, Gunapala N, Graham KJ. Determinants of soil microbial communities: effects of agricultural management, season, and soil type on phospholipid fatty acid profiles. Microb Ecol. 1998;36:1–12. doi: 10.1007/s002489900087. [DOI] [PubMed] [Google Scholar]
- 3.Fierer N, Jackson JA, Vilgalys R, Jackson RB. Assessment of soil microbial community structure by use of taxon-specific quantitative PCR assays. Appl Environ Microbiol. 2005;71:4117–4120. doi: 10.1128/AEM.71.7.4117-4120.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Folman LB, De Klein MJEM, Postma J, van Veen JA. Production of antifungal compounds by Lysobacter enzymogenes isolate 3.1T8 under different conditions in relation to its efficacy as a biocontrol agent of Pythium aphanidermatum in cucumber. Biol Control. 2004;31:145–154. doi: 10.1016/j.biocontrol.2004.03.008. [DOI] [Google Scholar]
- 5.Folman LB, Postma J, Van Veen JA. Characterisation of Lysobacter enzymogenes (Christensen and Cook 1978) strain 3.1T8, a powerful antagonist of fungal diseases of cucumber. Microbiol Res. 2003;158:107–115. doi: 10.1078/0944-5013-00185. [DOI] [PubMed] [Google Scholar]
- 6.Girvan MS, Bullimore J, Pretty JN, Osborn AM, Ball AS. Soil type is the primary determinant of the composition of the total and active bacterial communities in arable soils. Appl Environ Microbiol. 2003;69:1800–1809. doi: 10.1128/AEM.69.3.1800-1809.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Hayward AC, Fegan N, Fegan M, Stirling GR. Stenotrophomonas and Lysobacter: ubiquitous plant-associated gamma-proteobacteria of developing significance in applied microbiology. J Appl Microbiol. 2010;108:756–770. doi: 10.1111/j.1365-2672.2009.04471.x. [DOI] [PubMed] [Google Scholar]
- 8.Huijsdens XW, Linskens RK, Mak M, Meuwissen SGM, Vandenbroucke-Grauls CMJE, Savelkoul PHM. Quantification of bacteria adherent to gastrointestinal mucosa by real-time PCR. J Clin Microbiol. 2002;40:4423–4427. doi: 10.1128/JCM.40.12.4423-4427.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Islam MT, Hashidoko Y, Deora A, Ito T, Tahara S. Suppression of damping-off disease in host plants by the rhizoplane bacterium Lysobacter sp. strain SB-K88 is linked to plant colonization and antibiosis against soilborne peronosporomycetes. Appl Environ Microbiol. 2005;71:3786–3796. doi: 10.1128/AEM.71.7.3786-3796.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Ji G-H, Wei L-F, He Y-Q, Wu Y-P, Bai X-H. Biological control of rice bacterial blight by Lysobacter antibioticus strain 13–1. Biol Control. 2008;45:288–296. doi: 10.1016/j.biocontrol.2008.01.004. [DOI] [Google Scholar]
- 11.Kamilova F, Kravchenko LV, Shaposhnikov AI, Azarova T, Makarova N, Lugtenberg B. Organic acids, sugars, and L-tryptophane in exudates of vegetables growing on stonewool and their effects on activities of rhizosphere bacteria. Mol Plant-Microbe Interact. 2006;19:250–256. doi: 10.1094/MPMI-19-0250. [DOI] [PubMed] [Google Scholar]
- 12.Klerks MM, Zijlstra C, Van Bruggen AHC. Comparison of real-time PCR methods for detection of Salmonella enterica and Escherichia coli O157:H7, and introduction of a general internal amplification control. J Microbiol Meth. 2004;59:337–349. doi: 10.1016/j.mimet.2004.07.011. [DOI] [PubMed] [Google Scholar]
- 13.Kobayashi DY, El-Barrad NEH. Selection of bacterial antagonists using enrichment cultures for the control of summer patch disease in Kentucky bluegrass. Curr Microbiol. 1996;32:106–110. doi: 10.1007/s002849900019. [DOI] [Google Scholar]
- 14.Kobayashi DY, Reedy RM, Palumbo JD, Zhou JM, Yuen GY. A clp gene homologue belonging to the Crp gene family globally regulates lytic enzyme production, antimicrobial activity, and biological control activity expressed by Lysobacter enzymogenes strain C3. Appl Environ Microbiol. 2005;71:261–269. doi: 10.1128/AEM.71.1.261-269.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Köhl J, Groenenboom-de Haas BH, Kastelein P, Rossi V, Waalwijk C. Quantitative detection of pear-pathogenic Stemphylium vesicarium in orchards. Phytopathology. 2009;99:1377–1386. doi: 10.1094/PHYTO-99-12-1377. [DOI] [PubMed] [Google Scholar]
- 16.Köhl J, van Tongeren OFR, Groenenboom-de Haas BH, van Hoof RA, Driessen R, van der Heijden L. Epidemiology of dark leaf spot caused by Alternaria brassicola and A. brassicae in organic seed production of cauliflower. Plant Pathol. 2010;59:358–367. doi: 10.1111/j.1365-3059.2009.02216.x. [DOI] [Google Scholar]
- 17.Lauber CL, Strickland MS, Bradford MA, Fierer N. The influence of soil properties on the structure of bacterial and fungal communities across land-use types. Soil Biol Biochem. 2008;40:2407–2415. doi: 10.1016/j.soilbio.2008.05.021. [DOI] [Google Scholar]
- 18.Miller HJ, Liljeroth E, Henken G, Van Veen JA. Fluctuations in the fluorescent pseudomonad and actinomycete populations of rhizosphere and rhizoplane during the growth of spring wheat. Can J Microbiol. 1990;36:254–258. doi: 10.1139/m90-044. [DOI] [Google Scholar]
- 19.Nijhuis EH, Pastoor R, Postma J. Specific detection of Lysobacter enzymogenes (Christensen and Cook 1978) strain 3.1T8 with TaqMan® PCR. J Appl Microbiol. 2010;108:1155–1166. doi: 10.1111/j.1365-2672.2009.04519.x. [DOI] [PubMed] [Google Scholar]
- 20.Nybroe O, Brandt KK, Nicolaisen MH, Sørensen J. Methods to detect and quantify bacteria in soil. In: van Elsas JD, Jansson JK, Trevors JT, editors. Modern soil microbiology. 2. Boca Raton: CRC Press; 2007. pp. 283–316. [Google Scholar]
- 21.Park JH, Kim R, Aslam Z, Jeon CO, Chung YR. Lysobacter capsici sp. nov., with antimicrobial activity, isolated from the rhizosphere of pepper, and emended description of the genus Lysobacter. Int J Syst Evol Microbiol. 2008;58:387–392. doi: 10.1099/ijs.0.65290-0. [DOI] [PubMed] [Google Scholar]
- 22.Postma J, Nijhuis EH, Yassin AF. Genotypic and phenotypic variation among Lysobacter capsici strains from Rhizoctonia-suppressive soils. Syst Appl Microbiol. 2010;33:232–235. doi: 10.1016/j.syapm.2010.03.002. [DOI] [PubMed] [Google Scholar]
- 23.Postma J, Scheper RWA, Schilder MT. Effect of successive cauliflower plantings and Rhizoctonia solani AG 2–1 inoculations on disease suppressiveness of a suppressive and a conducive soil. Soil Biol Biochem. 2010;42:804–812. doi: 10.1016/j.soilbio.2010.01.017. [DOI] [Google Scholar]
- 24.Postma J, Schilder MT, Bloem J, van Leeuwen-Haagsma WK. Soil suppressiveness and functional diversity of the soil microflora in organic farming systems. Soil Biol Biochem. 2008;40:2394–2406. doi: 10.1016/j.soilbio.2008.05.023. [DOI] [Google Scholar]
- 25.Postma J, Stevens LH, Wiegers GL, Davelaar E, Nijhuis EH. Biological control of Pythium aphanidermatum in cucumber with a combined application of Lysobacter enzymogenes strain 3.1T8 and chitosan. Biol Control. 2009;48:301–309. doi: 10.1016/j.biocontrol.2008.11.006. [DOI] [Google Scholar]
- 26.Reichenbach H (1992) The genus Lysobacter. In: Balows A, Trüper HG, Dworkin M, Harder W, Schleifer K-H (eds) The Prokaryotes 2nd ed, vol. IV, pp 3256–3275
- 27.Smith CJ, Osborn AM. Advantages and limitations of quantitative PCR (Q-PCR)-based approaches in microbial ecology. FEMS Microbiol Ecol. 2009;67:6–20. doi: 10.1111/j.1574-6941.2008.00629.x. [DOI] [PubMed] [Google Scholar]
- 28.Sukkel W, van Leeuwen-Haagsma W, van der Wel C. BIOM, organic farmers network for research, development and innovation. Odense: Joint Organic Congress; 2006. [Google Scholar]
- 29.Sullivan RF, Holtman MA, Zylstra GJ, White JF, Jr, Kobayashi DY. Taxonomic positioning of two biological control agents for plant diseases as Lysobacter enzymogenes based on phylogenetic analysis of 16S rDNA, fatty acid composition and phenotypic characteristics. J Appl Microbiol. 2003;94:1079–1086. doi: 10.1046/j.1365-2672.2003.01932.x. [DOI] [PubMed] [Google Scholar]
- 30.Van Elsas JD, Trevors JT, Starodub ME, Van Overbeek LS. Transfer of plasmid RP4 between pseudomonads after introduction into soil; influence of spatial and temporal aspects of inoculation. FEMS Microbiol Ecol. 1990;73:1–12. doi: 10.1111/j.1574-6968.1990.tb03919.x. [DOI] [Google Scholar]
- 31.Van Overbeek L, Van Elsas JD. Effects of plant genotype and growth stage on the structure of bacterial communities associated with potato (Solanum tuberosum L.) FEMS Microbiol Ecol. 2008;64:283–296. doi: 10.1111/j.1574-6941.2008.00469.x. [DOI] [PubMed] [Google Scholar]