

Abstract
Lysophosphatidylcholine is a bioactive lipid that regulates a large number of cellular processes and is especially present during the deposition and infiltration of inflammatory cells and deposition of atheromatous plaque. Such molecule is also present in saliva and feces of the hematophagous organism Rhodnius prolixus, a triatominae bug vector of Chagas disease. We have recently demonstrated that LPC is a modulator of Trypanosoma cruzi transmission. It acts as a powerful chemoattractant for inflammatory cells at the site of the insect bite, which will provide a concentrated population of cells available for parasite infection. Also, LPC increases macrophage intracellular calcium concentrations that ultimately enhance parasite invasion. Finally, LPC inhibits NO production by macrophages stimulated by live T. cruzi, and thus interferes with the immune system of the vertebrate host. In the present paper, we discuss the main signaling mechanisms that are likely used by such molecule and their eventual use as targets to block parasite transmission and the pathogenesis of Chagas disease.
1. Immune Response to Trypanosoma cruzi Infection in the Vertebrate Host
T. cruzi infects the vertebrate host through bite wounds produced in skin by a feeding bug or through the interaction of the parasite with conjunctival mucosa. Such interaction sometimes produces visible signs called Romaña's sign or chagoma inoculation. The histology of this initial site of infection is defined by an elevated number of mononuclear cells [1]. This first sign of infection suggests that T. cruzi can stimulate skin cells to produce mediators that trigger a local inflammatory response. Despite controversies about the mechanism of the pathogenesis of Chagas disease [2–5], until recently, some authors believed that the disease was limited to an acute phase, followed by a chronic phase that was considered an autoimmune disease, where the parasites would be physically linked to sites of inflammation in the heart and esophagus [6–8]. However, nowadays, the disease is considered multifactorial, with multiple and continuous interactions between pathogen and host [9]. After the incubation period of 2 to 3 weeks, infection with T. cruzi is manifested by the presence of a large number of parasites in the blood and tissues. Acute infection is accompanied by an excessive activation of the immune system that includes the production of high levels of cytokines, intense activation of T and B cells, lymphadenopathy, splenomegaly, and intense inflammation associated with tissue infection niches. The acute phase induces the development of an effective acquired immunity leading to the control of parasitemia. The chronic phase is considered lifelong and is associated with only a few parasites in the host. The beginning of chronic infection with T. cruzi is asymptomatic in most patients. However, with the advance of the disease, clinical manifestations become variable, ranging from no symptoms to the involvement of cardiovascular and/or gastrointestinal symptoms [10, 11]. Before the acquired immunity is established, the innate immune system appears to be essential for at least two important aspects of Chagas disease: control of replication of the parasite in the host tissue and progress of the inflammatory reaction. The latter, in turn, has been considered to be the main cause of tissue damage and dysfunction of certain organs in the host [11]. Some studies in experimental models of infection of T. cruzi suggest that the potent immune response to Th-1 CD4 and CD8 cells, with the production of specific inflammatory cytokines, such as interferon gamma (IFN-γ), tumour necrosis factor (TNF-α), and interleukin 12 (IL-12), as well as the production of reactive nitrogen species such as nitric oxide (NO), plays an important role in the control of parasitemia during the initial stage of the disease [4, 10–13]. Moreover, cells of innate immunity, such as natural killer (NK) cells, dendritic cells, and macrophages, are also key elements in the initial control of parasite replication [10–13].
In recent years, research on Chagas disease has focused on the investigation of the role of pathogen-associated molecular patterns (PAMPs) of protozoa, which are the targets of innate immune receptors. Also, the problem of identifying relevant receptors in innate immunity-parasite interactions during the evolution of the disease in the host has been addressed by several laboratories. This strategy ultimately aims at the development of therapeutic interventions through the use of PAMPs derived from parasites. Glycosyl-phosphatidyl-inositol (GPI) is the name given to the first glycoconjugate in T. brucei that was identified with the function of anchoring proteins on the cell surface [14–17]. PAMPs widely studied in T. cruzi are, in fact, GPI anchors. All evolutive forms of this parasite express on their surface GPI-anchored glycoproteins [14–17]. Some studies have identified GPI anchors isolated from trypomastigote-derived mucin-like glycoproteins (GPI-mucins) of T. cruzi as the molecules primarily responsible for stimulating the host immune system [18, 19]. Thus, T. cruzi GPI-mucins are able to activate macrophages and stimulate the production of proinflammatory cytokines, chemokines, and NO [20–22]. Innate immune response to T. cruzi has been studied extensively and is based on the activation of signaling pathways triggered by Toll-like receptors (TLRs). TLRs are proteins that recognize conserved motifs associated with several different pathogens; they trigger intracellular signaling cascades that ultimately lead to a complex host immune response [11, 12]. There are 10 TLRs described in humans and 12 in mice [11, 12]. Generally, the stimulus induced by GPI molecules occurs during the early phase of infection, where macrophages respond to trypomastigotes in a TLR-dependent mechanism and ultimately induce the production of IL-12 and TNF-α and trigger the responses of CD4 and CD8 cells through the production of IFN-γ [23]. Thus, macrophages activated by TNF-α and IFN-γ seem to have an important role in controlling parasite growth. Free GPI anchors or glycoinositolphospholipids (GIPLs) are also able to stimulate the host immune system. GIPLs are similar to GPIs but contain instead ceramide in their lipid moiety [18, 19].
TLRs 2, 4, and 9 are the major TLRs involved in innate immune response to T. cruzi [18, 24–29]. TLR2 has been identified as the main receptor responsible for macrophage activation by GPI mucins [18, 24–29]. According to Ropert and Gazzinelli [27], the receptor heterodimer composed of TLR2 and TRL6 is activated by GPI mucin and the CD14 coreceptor. Oliveira et al. [25] observed that GIPL from T. cruzi confers an inflammatory response via TLR4, promoting the recruitment of neutrophils into the peritoneal cavity of mice. Later, Medeiros et al. [26] demonstrated that this effect was partially dependent on the production of IL-1β. The genomic DNA of T. cruzi also plays an important role in proinflammatory response of the vertebrate host during infection, since TLR 9 is activated by CpG motifs from nonmethylated DNA [28, 29]. Besides the innate immune response mediated by TLRs, T. cruzi can also stimulate TLR-independent pathways that lead to the production of IFN-β and IFN-γ. In this case, this occurs due to a surge in intracellular calcium concentration which ultimately leads to the activation of calcineurin and calmodulin [30–32].
2. Lysophosphatidylcholine and Modulation of NO Production and Host Immunity
Lysophospholipids such as lysophosphatidylcholine (LPC), sphyngosylphosphoryilcholine (SPC), lysophosphatidic acid (LPA), and sphingosine-1-phosphate (S1P) regulate a large number of cellular processes. LPC is a derivative of phosphatidylcholine (PC) that arises by the loss of a fatty acid through the action of a phospholipase A2 (PLA2) or by transferring it to cholesterol by the action of a cholesterol-acetyltransferase [33]. LPC is involved in several physiological events and is already known as a central molecule in several pathological states, but it is especially present during the deposition and infiltration of inflammatory cells and deposition of atheromatous plaque [34–36]. Research directed towards LPC has increased greatly since the finding that these molecules are involved in atherosclerosis [37]. The idea that various phospholipases secreted by circulating leukocytes participate in this pathology was soon proposed. Thus, the current model suggests that diabetes and hypercholesterolemia contribute to generate a large number of LDL particles in plasma that can undergo oxidation of unsaturated fatty acids, generating an oxidized particle (oxLDL). Since on average 50% of LDL fatty acids are arachidonic acid and linoleic acid, the chances of such an oxidative event are huge. The oxLDL is a potential cause of the increased expression of inflammatory markers such as TNF-α, MCP-1, and MCSF that will attract differentiating monocytes to the lesion site. In this sense, LPC is one of the most powerful chemotactic signals for macrophages and is also generated by cells in the apoptotic process as mentioned above. OxLDL particles are recognized by various secretory PLA2 in the plasma, including type IIA, V, and X. Our group showed for the first time the presence of phospholipids and lysophospholipids in saliva and feces of the hematophagous organism Rhodnius prolixus, a triatominae bug vector of Chagas disease [38]. The major lipids present in R. prolixus saliva are PC and LPC [38]. Salivary LPC is an additional antihemostatic molecule that is part of the pharmacological arsenal injected into the bite site to allow the insect to feed. It inhibits platelet aggregation and increases the production of NO in endothelial cells. Thus, LPC was initially described as a molecule with antiplatelet and vasodilatory activities, and a few years later, its effect as an immunomodulator of T.cruzi infection was demonstrated [38, 39].
The role of LPC as a modulator of T. cruzi transmission occurs by three main mechanisms summarized on Figure 1 and mentioned as follows.
Figure 1.
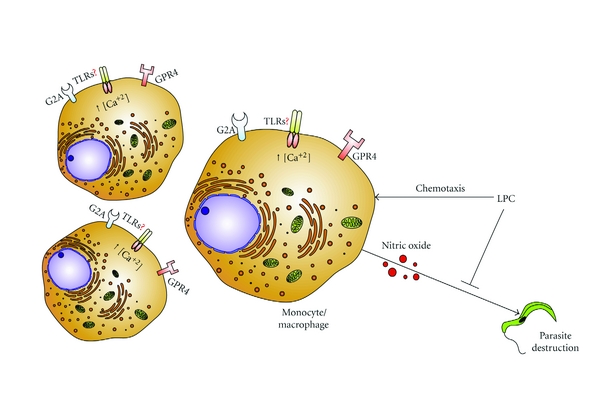
LPC-induced signaling on monocytes and macrophages. LPC is a signaling molecule that may act through different receptors on cell surface such as G2A and GPC4. Despite its description as a proinflammatory molecule, LPC-mediated signaling through TLRs is not demonstrated yet. LPC induces cell chemotaxis which ultimately increases the number of cells in the wound site. Also, LPC-treated cells undergo a decrease on NO synthesis when stimulated by parasite or LPS. Finally, a transient increase on intracellular calcium is also reported in such cells. These combined effects enhance the number of cells prone to T. cruzi invasion.
(1) LPC is a vector-derived molecule. It acts as a powerful chemoattractant for inflammatory cells at the site of the insect bite. This event will provide a concentrated population of cells available for T. cruzi infection [38, 39].
(2) LPC increases macrophage intracellular calcium concentrations that ultimately enhance parasite invasion.
(3) LPC inhibits NO production by macrophages stimulated by either live T. cruzi, LPS, or LPS in the presence of IFN-γ, and thus interferes with the immune system of the vertebrate host [39].
The above findings demonstrate that LPC is now a signaling molecule with effects beyond that of counteracting host blood hemostasis, since it acts as modulator of NO biology and parasite transmission [40–43]. Macrophages are intimately related to the establishment of acute infection with T. cruzi, since the success of the infection depends on the initial invasion of these cells [44–46]. This leads to the assumption that salivary LPC may facilitate the parasite infection, favoring not only insect feeding, but also preparing the environment for the arrival of the parasite, minutes or hours after the initial bug bite. Recent results obtained by our group demonstrated that injection of salivary LPC into host skin followed by parasite inoculation in the same site minutes later ultimately increases blood parasitemia from 3- to 6-fold in animals infected with T. cruzi. LPC's effect on parasitemia is mainly achieved by the activation of macrophage chemotaxis and immunosuppression of NO production induced by the parasite. We also showed an increase in the rate of association of the parasite with macrophages induced either by 500-fold diluted saliva or by LPC. This was the first demonstration of a potentiating factor of transmission of Chagas disease and the first implication of a lysophospholipid in an infectious disease [39].
The activation of receptors that recognize the parasite by the presence of specific structures on its surface stimulates host cells to produce TNF-α, IL-12, and NO, as mentioned above. Depending on the MyD88 adapter protein, TLRs 2, 4, and 9 have been implicated in the network used by the immune system of the mammalian host to control infection by T. cruzi [14–18]. Campos et al. [18, 23] were the first to demonstrate the involvement of TLR2 in the interaction between the parasite and host macrophages. The expression of TLR2 is essential for the induction of IL-12, TNF-α, and NO, and this receptor is activated by parasite-derived molecules such as GPI anchors, which have been isolated from the surface of trypomastigotes of T. cruzi [14–18]. The production of NO but not IL-12 by T. cruzi-exposed macrophages is not affected by bug saliva [39]. Curiously, in bone-marrow-derived macrophages obtained from TLR2-deficient mice, the production of IL-12 is largely suppressed by LPC. These data indicate that in some cell types, the production of this cytokine may be affected by this lysophospholipid through a TLR2-independent mechanism.
Moreover, GIPLs from T. cruzi are TLR4 agonists with proinflammatory effects [25, 26]. We showed that NO production, induced by the parasite or by lipopolysaccharide (LPS), another ligand of TLR4, either in murine peritoneal macrophages or bone marrow-derived macrophages, is blocked in both cases by LPC even in the presence of IFN-γ in vitro [39]. The ability of LPC to reverse the induction of NO production in all cases, almost independently of the ligand type, suggests that this lysophospholipid must act by a unique pathway. In this regard, the receptors involved in cell signaling induced by LPC, in general, exhibit a certain promiscuity with respect to the ligand and vice versa. In the case of LPC, different receptors have been proposed for this molecule, including G2A, a G protein-coupled receptor, and GPR4, another important candidate [45, 47–49]. Despite the controversy generated in the literature due to the low reproducibility of the studies using radioactive LPC and its interaction with candidate receptors, the ability of G2A to bind fatty acids and protons is noteworthy [33]. Thus, G2A remains in the literature as the best-known receptor involved in the adaptation of the signal induced by LPC [47–49]. Moreover, the redistribution of G2A receptor itself and the exposure of TLR4 are influenced by LPC metabolism [33, 50]. In this case, the content of intracellular LPC is finely controlled by the activity of a lysophosphatidylcholine acyltransferase (LPCAT), an enzyme that uses LPC as a substrate and generates phospholipids as the product of its action. The treatment of monocytes with LPS activates this enzyme and increases the transport of TLR4 to membrane rafts in these cells [50]. Since the LPCAT inhibitor used, 5-hydroxyethyl 5′3′ thiophenol pyridine (HETP), increases the lysophospholipids/phospholipids ratios, it reverses the effect of LPS [50]. Thus, it seems appropriate to propose that in the presence of T. cruzi, one should conduct a map of the distribution of both receptors, G2A and TLR4, in the presence and in the absence of LPC using both proteomic and immunological methods [33, 50].
During programmed cell death, LPC is generated by a calcium-independent PLA2 activated by caspase-3. Thus, LPC acts as a chemotactic find-me signal that attracts the phagocyte to the apoptotic cells [49, 51, 52] and as an eat-me signal involving recruitment of complement proteins for recognition by phagocytes [49, 51, 52]. Such LPC-induced chemotaxis is very interesting, because in Chagas disease, the uptake of apoptotic cells by macrophages infected with T. cruzi stimulates parasite growth [53]. In addition, it has been shown that T. cruzi infective stages are able to generate lipid messengers, including LPC, that modulate host cell signaling [54]. Regarding adaptor molecules mobilized in response to LPC, it is known that in most cell types, there is the involvement of isoforms of protein kinase C [55]. Probably it is the type of isoform activated in each cell that directs the intensity and type of response triggered by LPC in that specific cell type. When combined with different types of TLRs and adapters, LPC-mediated signaling must produce a specific and still poorly understood repertoire of immunosuppression.
3. Vector Phospholipases and Eventual Target to Block T. cruzi Transmission
PLA2 is an enzyme family present in various organisms such as viruses, bacteria, plants, and animals. According to studies done on mammals, the action of PLA2 is important for the remodeling of cell membranes, lipid digestion, cell signaling, and immune defense of the host as well as production of various lipid mediators [56–61]. In insects, the phospholipases that have been studied are related to the venom injected into their prey, the physiology of digestion, immunity, and reproduction [61]. Among the published studies on phospholipase activity in arthropods are those reporting the presence of such enzymes in the salivary glands of Manduca sexta [62] and in the saliva and salivary glands of Amblyoma americanum [63, 64]. These studies have found a correlation between PLA2 activity and digestion. In addition, Zhu et al. [64] suggested another role for this activity, linking it to the production of prostaglandins, promotion of vasodilation and the suppression of inflammation and immunity. The production of prostacyclin may also lead to the inhibition of platelet aggregation and the induction of vasodilation. Furthermore, platelet-activating factor (PAF) acetyl hydrolase, a member of the GVII family of PLA2 enzymes, is a serine-dependent hydrolase that does not require Ca2+ for activity. This enzyme cleaves the acetyl group from the sn-2 position of the phospholipid, and in the case of PAF, there is the hydrolysis of the sn-2 ester bond, releasing acetate and biologically inactive lyso-PAF [65, 66]. Cat flea (Ctenocephalides felis) salivary gland homogenate has a PAF-acetylhydrolase activity, and the estimated amount of activity in a single pair of salivary glands (~5 pmol/min) is of the right order of magnitude to induce a localized anti-inflammatory/allergic effect [67]. Extracellular PAF is proinflammatory and acts via very high affinity G-coupled protein receptors, causing activation of platelets, neutrophils, and monocytes [66]. The hypothesis is that PAF-acetylhydrolase activities from saliva can downregulate inflammatory and immune reactions mediated by PAF released from host cells. This may happen as a reaction to injected cat flea saliva and may be interrupted by host grooming or scratching the locale of the bite. Besides these reports, phospholipases also have been identified in transcriptomes of saliva or salivary glands of some hematophagous arthropods such as the soft ticks Ornithodoros coriaceus [67, 68] and O. parkeri [69], of hard ticks such as Ixodes pacificus [70], and in insects such as Anopheles funestus [71], Phlebotomus arabicus [72], and Glossina morsitans [73].
Zeidner et al. [74] have suggested that aside from facilitating some tick digestive processes, it is possible that secretion of PLA2 into the feed site creates some protective barrier against bacteria that can be carried into the wound. They demonstrated that the borreliacidal activity found in A. americanum saliva is most probably due to the enzymatic effects of PLA2 and that it would directly and rapidly kill Borrelia burgdorgeri through the digestion of membrane lipids, composed by a majority of PC and phosphatidylglycerol. The authors hypothesize that high level of PLA2 enzymatic activity present in saliva is related to A. americanum's refractoriness to B. burgdorgeri. Other studies have demonstrated the importance of PLA2 in the infection processas elicited by pathogens such as Toxoplasma gondii, Cryptosporium parvum, Entamoeba histolytica, Leishmania amazonensis, and T. cruzi [75–77]. Moreover, Connelly and Kierszenbaum [44] showed that the presence of PLA2 significantly increased the association between T. cruzi and macrophages, and they suggest that this effect relies on alterations of the parasite membrane, since it was induced by pretreatment of parasite membranes with PLA2 but not macrophages. But nowadays, as cited above, the presence of PLA2 in salivary secretions of T. cruzi vectors implies LPC generation and its further involvement in the inflammatory process that occurs during infection.
PLA2 enzymes from snake venom induce a wide spectrum of pharmacological effects, including anticoagulant proprieties that can be mediated by hydrolysis of phospholipid or by a nonenzymatic mechanism, such as when PLA2 from Naja nigricollis venom binds factor Xa in the coagulation cascade through the specific anticoagulant site on its surface [78]. Our group is investigating a further role for the LPC present in the saliva of vectors, which we believe is related to muscular paralysis. Rigoni et al. [79] have shown that lysophospholipids, in particular LPC, can block the exocytosis of neurotransmitters, thus paralyzing the muscle. In this context, using the predator insect Belostoma anurum as model, we showed that the salivary LPC also has this property. Our hypothesis is that B. anurum uses lysophospholipids as a way to paralyze the prey while it feeds, since it makes an extraoral digestion [80]. We obtained similar results with LPC from R. prolixus, with less pronounced blockage of exocytosis. LPC action may be more local in order to avoid disturbing the host. Thus, the above data show that the presence of LPC generated by PLA2s in salivary secretions of predators and blood-sucking arthopods is widespread in the animal kingdom, and this molecule may be a surviving trace of ancient feeding habits.
Another aspect that should be emphasized is that PLA2s also generate free fatty acids that can be converted to eicosanoids. Eicosanoids are polyunsaturated fatty acids of 20 carbons that act as local mediators of short half-life; they are derived from arachidonic acid (20 : 4 n-6) or other polyunsaturated 20-carbon precursors (20 : 3 n-6 and 20 : 5 n-3). Arachidonic acid is esterified in phospholipids of plasma membranes, these being released by the action of PLA2. The biological action of arachidonic acid products requires its oxygenation, which can take place in three different ways: (a) via the cyclooxygenases that generate prostaglandins and thromboxanes, (b) via the lipooxygenases that generate leukotrienes and lipoxins, and (c) via the cytochrome P-450, which generates epoxides [81]. Physiological processes that usually involve autacoids, hormones, and growth factors may stimulate the release of arachidonic acid, as already widely described in mammals, as in mediating immune and inflammatory response of late vertebrates [82]. Recently, the involvement of thromboxane A2 (TXA2) in the process of vertebrate host infection by T. cruzi [83] was demonstrated. The same group showed that the eicosanoid TXA2 is prevalent in all life stages of the parasite. Thus, in infected mice, the parasite itself may account for 90% of the total TXA2 in plasma. In this regard, it is noteworthy that the production of TXA2 from arachidonic acid occurs by the cyclooxygenase pathway. Accordingly, results from our laboratory indicate that half of the fatty acids ingested along with blood are unsaturated and about half of them are arachidonic acid. So, if a pool of TXA2 is a prerequisite for the process of infection of host cells by any pathogen, that pool could be generated during the final stages of blood digestion in the vector at the expense of fatty acids released there. A triacylglycerol-lipase activity was identified in the gut lumen of blood-fed insects and is probably involved in the digestion of lipids from the blood meal. These lipase activities and also the metabolism and fate of lipids that are generated during digestion of ingested blood were studied and characterized [84]. However, neither the dynamic generation of free fatty acids in insects infected with T. cruzi nor their processing to TXA2 in the final stage of the digestive process has ever been assessed in any vector. Thus, an attractive model for the future might involve the silencing of a PLA2 gene in the saliva of Chagas disease vectors to obtain LPC-depleted or LPC-free saliva. The saliva of these insects would be expected to lower the rate of infection of the vertebrate host.
4. The Role of Host Plasma LPC in T. cruzi Infection
The original studies that implicated LPC in the pathogenesis of atherosclerosis tended to highlight the presence of this phospholipid in atheromatous lesions [37]. However, the origin and dynamics of the formation of this molecule remained unknown for many years. Wilensky et al. [85] identified the main enzyme responsible for the generation of LPC, Lp-PLA2, also known as PAF-acetyl hydrolase or phospholipase VIIA, which is secreted by leukocytes and associated with plasma lipoproteins, especially LDL. This enzyme recognizes and cleaves oxLDL and oxidized phospholipids, generating LPC and free fatty acid oxidation (oxNEFAS, or oxidized nonesterified fatty acids). The LPC, as previously mentioned, is a potent proinflammatory molecule capable of leukocyte recruitment and activation with induction of apoptosis. Demonstration of Lp-PLA2 in the necrotic core of atheromatous lesions and fibrous cap of vulnerable plaques supports our current views of the importance of this enzyme in atherogenesis. An important therapeutic option is the selective inhibition of Lp-PLA2. Wilensky et al. [85] showed that the administration of darapladib (GlaxoSmithKline) in experimental models selectively reduces the activity of this enzyme, attenuates the formation of LPC, and reduces the formation of atherosclerotic plaques with negative regulation of proinflammatory genes in macrophages and T lymphocytes. Chagas disease treatment aims to slow the progression of myocardial impairment caused by invasion of the patient's heart by the parasite. Some of the drugs used to treat Chagas disease cause changes in the patients plasma lipid profile, leading to high concentrations of LDL. During treatment, these patients are likely to present favorable conditions for LDL oxidation and generation of LPC, which will certainly trigger the proinflammatory phenotype, thereby maintaining levels of reinfection of myocardial cells. In this sense, it would be important to evaluate the generation of LPC in Chagas disease patients treated with various categories of drugs in order to verify the formation of this lipid mediator. Likewise, chronic treatment with the drug in healthy experimental animals should be carried out to identify any effects on the levels of LPC and subsequent susceptibility to infection by T. cruzi. In conclusion, darapladib may constitute a novel tool with dual use: to optimize the current therapeutic treatment of chronic chagasic patients and to experimentally modify the plasma levels of LPC in mice to determine whether the reduction of such levels decreases the susceptibly to infection by T. cruzi.
Acknowledgments
The studies on cell signaling in the authors' laboratories were supported by the Brazilian agencies Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), Fundação de Amparo à Pesquisa Carlos Chagas Filho (FAPERJ), and also by the funding program “Instituto Nacional de Ciência e Tecnologia em Entomologia Molecular”. The authors wish to express their gratitude to Mr. Nilson Porto who kindly drew the figure depicted in this paper. This work is dedicated to Dr. Martha Meriwether Sorenson who has reviewed with outstanding scientific criticism hundreds of manuscripts produced by the researchers of Instituto de Bioquímica Médica, UFRJ, Rio de Janeiro, Brazil, in the last 27 years.
References
- 1.Dias JC. Cecílio Romaña, Romaña’s sign and Chagas’ disease. Revista da Sociedade Brasileira de Medicina Tropical. 1997;30(5):407–413. doi: 10.1590/s0037-86821997000500012. [DOI] [PubMed] [Google Scholar]
- 2.Andrade ZA. Pathogenesis of Chagas’ disease. Research in Immunology. 1991;142(2):126–129. doi: 10.1016/0923-2494(91)90021-a. [DOI] [PubMed] [Google Scholar]
- 3.Tarleton RL. Chagas disease: a role for autoimmunity? Trends in Parasitology. 2003;19(10):447–451. doi: 10.1016/j.pt.2003.08.008. [DOI] [PubMed] [Google Scholar]
- 4.Golgher D, Gazzinelli RT. Innate and acquired immunity in the pathogenesis of Chagas disease. Autoimmunity. 2004;37(5):399–409. doi: 10.1080/08916930410001713115. [DOI] [PubMed] [Google Scholar]
- 5.Andrade LO, Andrews NW. Opinion: the Trypanosoma cruzi—host-cell interplay: location, invasion, retention. Nature Reviews Microbiology. 2005;3(10):819–823. doi: 10.1038/nrmicro1249. [DOI] [PubMed] [Google Scholar]
- 6.Jones EM, Colley DG, Tostes S, Lopes ER, Vnencak-Jones CL, McCurley TL. Amplification of a Trypanosoma cruzi DNA sequence from inflammatory lesions in human chagasic cardiomyopathy. American Journal of Tropical Medicine and Hygiene. 1993;48(3):348–357. doi: 10.4269/ajtmh.1993.48.348. [DOI] [PubMed] [Google Scholar]
- 7.Benvenuti LA, Roggério A, Freitas HFG, Mansur AJ, Fiorelli A, Higuchi ML. Chronic American trypanosomiasis: parasite persistence in endomyocardial biopsies is associated with high-grade myocarditis. Annals of Tropical Medicine and Parasitology. 2008;102(6):481–487. doi: 10.1179/136485908X311740. [DOI] [PubMed] [Google Scholar]
- 8.Vago AR, Macedo AM, Oliveira RP, et al. Kinetoplast DNA signatures of Trypanosoma cruzi strains obtained directly from infected tissues. The American Journal of Pathology. 1996;149(6):2153–2159. [PMC free article] [PubMed] [Google Scholar]
- 9.Dutra WO, Gollob KJ. Current concepts in immunoregulation and pathology of human Chagas disease. Current Opinion in Infectious Diseases. 2008;21(3):287–292. doi: 10.1097/QCO.0b013e3282f88b80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Rassi A, Jr., Rassi A, Marin-Neto JA. Chagas disease. The Lancet. 2010;375(9723):1388–1402. doi: 10.1016/S0140-6736(10)60061-X. [DOI] [PubMed] [Google Scholar]
- 11.Junqueira C, Caetano B, Bartholomeu DC, et al. The endless race between Trypanosoma cruzi and host immunity: lessons for and beyond Chagas disease. Expert Reviews in Molecular Medicine. 2010;12, article e29 doi: 10.1017/S1462399410001560. [DOI] [PubMed] [Google Scholar]
- 12.Takeuchi O, Akira S. Pattern recognition receptors and inflammation. Cell. 2010;140(6):805–820. doi: 10.1016/j.cell.2010.01.022. [DOI] [PubMed] [Google Scholar]
- 13.Brener Z, Gazzinelli RT. Immunological control of Trypanosoma cruzi infection and pathogenesis of Chagas’ disease. International Archives of Allergy and Immunology. 1997;114(2):103–110. doi: 10.1159/000237653. [DOI] [PubMed] [Google Scholar]
- 14.Ferguson MAJ. The structure, biosynthesis and functions of glycosylphosphatidylinositol anchors, and the contributions of trypanosome research. Journal of Cell Science. 1999;112(17):2799–2809. doi: 10.1242/jcs.112.17.2799. [DOI] [PubMed] [Google Scholar]
- 15.Ferguson MAJ, Low MG, Cross GAM. Glycosyl-sn-1,2-dimyristylphosphatidylinositol is covalently linked to Trypanosoma brucei variant surface glycoprotein. The Journal of Biological Chemistry. 1985;260(27):14547–14555. [PubMed] [Google Scholar]
- 16.Almeida IC, Camargo MM, Procópio DO, et al. Highly purified glycosylphosphatidylinositols from Trypanosoma cruzi are potent proinflammatory agents. The EMBO Journal. 2000;19(7):1476–1485. doi: 10.1093/emboj/19.7.1476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Almeida IC, Gazzinelli RT. Proinflammatory activity of glycosylphosphatidylinositol anchors derived from Trypanosoma cruzi: structural and functional analyses. Journal of Leukocyte Biology. 2001;70(4):467–477. [PubMed] [Google Scholar]
- 18.Campos MAS, Almeida IC, Takeuchi O, et al. Activation of toll-like receptor-2 by glycosylphosphatidylinositol anchors from a protozoan parasite. The Journal of Immunology. 2001;167(1):416–423. doi: 10.4049/jimmunol.167.1.416. [DOI] [PubMed] [Google Scholar]
- 19.Coelho PS, Klein A, Talvani A, et al. Glycosylphosphatidylinositol-anchored mucin-like glycoproteins isolated from Trypanosoma cruzi Trypomastigotes induce in vivo leukocyte recruitment dependent on MCP-1 production by IFN-γ-primed-macrophages. Journal of Leukocyte Biology. 2002;71(5):837–844. [PubMed] [Google Scholar]
- 20.Gutierrez FRS, Mineo TWP, Pavanelli WR, Guedes PMM, Silva JS. The effects of nitric oxide on the immune system during Trypanosoma cruzi infection. Memórias do Instituto Oswaldo Cruz. 2009;104(1):236–245. doi: 10.1590/s0074-02762009000900030. [DOI] [PubMed] [Google Scholar]
- 21.Aliberti JCS, Machado FS, Souto JT, et al. β-chemokines enhance parasite uptake and promote nitric oxide-dependent microbiostatic activity in murine inflammatory macrophages infected with Trypanosoma cruzi . Infection and Immunity. 1999;67(9):4819–4826. doi: 10.1128/iai.67.9.4819-4826.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Shoda LKM, Kegerreis KA, Suarez CE, et al. DNA from protozoan parasites Babesia bovis, Trypanosoma cruzi, and T. brucei is mitogenic for B lymphocytes and stimulates macrophage expression of interleukin-12, tumor necrosis factor alpha, and nitric oxide. Infection and Immunity. 2001;69(4):2162–2171. doi: 10.1128/IAI.69.4.2162-2171.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Campos MA, Closel M, Valente EP, et al. Impaired production of proinflammatory cytokines and host resistance to acute infection with Trypanosoma cruzi in mice lacking functional myeloid fifferentiation factor 88. The Journal of Immunology. 2004;172(3):1711–1718. doi: 10.4049/jimmunol.172.3.1711. [DOI] [PubMed] [Google Scholar]
- 24.Martin D, Tarleton R. Generation, specificity, and function of CD8+ T cells in Trypanosoma cruzi infection. Immunological Reviews. 2004;201:304–317. doi: 10.1111/j.0105-2896.2004.00183.x. [DOI] [PubMed] [Google Scholar]
- 25.Oliveira AC, Peixoto JR, de Arrada LB, et al. Expression of functional TLR4 confers proinflammatory responsiveness to Trypanosoma cruzi glycoinositolphospholipids and higher resistance to infection with T. cruzi . The Journal of Immunology. 2004;173(9):5688–5696. doi: 10.4049/jimmunol.173.9.5688. [DOI] [PubMed] [Google Scholar]
- 26.Medeiros MM, Peixoto JR, Oliveira AC, et al. Toll-like receptor 4 (TLR4)-dependent proinflammatory and immunomodulatory properties of the glycoinositolphospholipid (GIPL) from Trypanosoma cruzi . Journal of Leukocyte Biology. 2007;82(3):488–496. doi: 10.1189/jlb.0706478. [DOI] [PubMed] [Google Scholar]
- 27.Ropert C, Gazzinelli RT. Regulatory role of toll-like receptor 2 during infection with Trypanosoma cruzi . Journal of Endotoxin Research. 2004;10(6):425–430. doi: 10.1179/096805104225006507. [DOI] [PubMed] [Google Scholar]
- 28.Gazzinelli RT, Denkers EY. Protozoan encounters with Toll-like receptor signalling pathways: implications for host parasitism. Nature Reviews Immunology. 2006;6(12):895–906. doi: 10.1038/nri1978. [DOI] [PubMed] [Google Scholar]
- 29.Bafica A, Santiago HC, Goldszmid R, Ropert C, Gazzinelli RT, Sher A. Cutting edge: TLR9 and TLR2 signaling together account for MyD88-dependent control of parasitemia in Trypanosoma cruzi infection. The Journal of Immunology. 2006;177(6):3515–3519. doi: 10.4049/jimmunol.177.6.3515. [DOI] [PubMed] [Google Scholar]
- 30.Hemmi H, Takeuchi O, Kawai T, et al. A Toll-like receptor recognizes bacterial DNA. Nature. 2000;408(6813):740–745. doi: 10.1038/35047123. [DOI] [PubMed] [Google Scholar]
- 31.Chessler AD, Ferreira LR, Chang TH, Fitzgerald KA, Burleigh BA. A novel IFN regulatory factor 3-dependent pathway activated by trypanosomes triggers IFN-beta in macrophages and fibroblasts. The Journal of Immunology. 2008;181(11):7917–7924. doi: 10.4049/jimmunol.181.11.7917. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Chessler ADC, Unnikrishnan M, Bei AK, Daily JP, Burleigh BA. Trypanosoma cruzi triggers an early type I IFN response in vivo at the site of intradermal infection. The Journal of Immunology. 2009;182(4):2288–2296. doi: 10.4049/jimmunol.0800621. [DOI] [PubMed] [Google Scholar]
- 33.Kabarowski JH. G2A and LPC: regulatory functions in immunity. Prostaglandins and Other Lipid Mediators. 2009;89(3-4):73–81. doi: 10.1016/j.prostaglandins.2009.04.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Wang L, Radu CG, Yang LV, Bentolila LA, Riedinger M, Witte ON. Lysophosphatidylcholine-induced surface redistribution regulates signaling of the murine G protein-coupled receptor G2A. Molecular Biology of the Cell. 2005;16(5):2234–2247. doi: 10.1091/mbc.E04-12-1044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Wang J, Zhang Y, Wang H, et al. Potential mechanisms for the enhancement of HERG K+ channel function by phospholipid metabolites. British Journal of Pharmacology. 2004;141(4):586–599. doi: 10.1038/sj.bjp.0705646. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Drobnik W, Liebisch G, Audebert FX, et al. Plasma ceramide and lysophosphatidylcholine inversely correlate with mortality in sepsis patients. The Journal of Lipid Research. 2003;44(4):754–761. doi: 10.1194/jlr.M200401-JLR200. [DOI] [PubMed] [Google Scholar]
- 37.Kugiyama K, Kerns SA, Morrisett JD, Roberts R, Henry PD. Impairment of endothelium-dependent arterial relaxation by lysolecithin in modified low-density lipoproteins. Nature. 1990;344(6262):160–162. doi: 10.1038/344160a0. [DOI] [PubMed] [Google Scholar]
- 38.Golodne DM, Monteiro RQ, Graça-Souza AV, Silva-Neto MAC, Atella GC. Lysophosphatidylcholine acts as an anti-hemostatic molecule in the saliva of the blood-sucking bug Rhodnius prolixus . The Journal of Biological Chemistry. 2003;278(30):27766–27771. doi: 10.1074/jbc.M212421200. [DOI] [PubMed] [Google Scholar]
- 39.Mesquita RD, Carneiro AB, Bafica A, et al. Trypanosoma cruzi infection is enhanced by vector saliva through immunosuppressant mechanisms mediated by lysophosphatidylcholine. Infection and Immunity. 2008;76(12):5543–5552. doi: 10.1128/IAI.00683-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Murugesan G, Rani MRS, Gerber CE, et al. Lysophosphatidylcholine regulates human microvascular endothelial cell expression of chemokines. Journal of Molecular and Cellular Cardiology. 2003;35(11):1375–1384. doi: 10.1016/j.yjmcc.2003.08.004. [DOI] [PubMed] [Google Scholar]
- 41.Lauber K, Blumenthal SG, Waibel M, Wesselborg S. Clearance of apoptotic cells: getting rid of the corpses. Molecular Cell. 2004;14(3):277–287. doi: 10.1016/s1097-2765(04)00237-0. [DOI] [PubMed] [Google Scholar]
- 42.Radu CG, Yang LV, Riedinger M, Au M, Witte ON. T cell chemotaxis to lysophosphatidylcholine through the G2A receptor. Proceedings of the National Academy of Sciences of the United States of America. 2004;101(1):245–250. doi: 10.1073/pnas.2536801100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Yang LV, Radu CG, Wang L, Riedinger M, Witte ON. Gi-independent macrophage chemotaxis to lysophosphatidylcholine via the immunoregulatory GPCR G2A. Blood. 2005;105(3):1127–1134. doi: 10.1182/blood-2004-05-1916. [DOI] [PubMed] [Google Scholar]
- 44.Connelly MC, Kierszenbaum F. Modulation of macrophage interaction with Trypanosoma cruzi by phospholipase A2-sensitive components of the parasite membrane. Biochemical and Biophysical Research Communications. 1984;121(3):931–939. doi: 10.1016/0006-291x(84)90766-6. [DOI] [PubMed] [Google Scholar]
- 45.Denkers EY, Butcher BA. Sabotage and exploitation in macrophages parasitized by intracellular protozoans. Trends in Parasitology. 2005;21(1):35–41. doi: 10.1016/j.pt.2004.10.004. [DOI] [PubMed] [Google Scholar]
- 46.Kollien AH, Schaub GA. The development of Trypanosoma cruzi in triatominae. Parasitology Today. 2000;16(9):381–387. doi: 10.1016/s0169-4758(00)01724-5. [DOI] [PubMed] [Google Scholar]
- 47.Kabarowski JHS, Zhu K, Le LQ, Witte ON, Xu Y. Lysophosphatidylcholine as a ligand for the immunoregulatory receptor G2A. Science. 2001;293(5530):702–705. doi: 10.1126/science.1061781. [DOI] [PubMed] [Google Scholar]
- 48.Zhu K, Baudhuin LM, Hong G, et al. Sphingosylphosphorylcholine and lysophosphatidylcholine are ligands for the G protein-coupled receptor GPR4. The Journal of Biological Chemistry. 2001;276(44):41325–41335. doi: 10.1074/jbc.M008057200. [DOI] [PubMed] [Google Scholar]
- 49.Peter C, Waibel M, Radu CG, et al. Migration to apoptotic “find-me” signals is mediated via the phagocyte receptor G2A. The Journal of Biological Chemistry. 2008;283(9):5296–5305. doi: 10.1074/jbc.M706586200. [DOI] [PubMed] [Google Scholar]
- 50.Jackson SK, Abate W, Parton J, Jones S, Harwood JL. Lysophospholipid metabolism facilitates Toll-like receptor 4 membrane translocation to regulate the inflammatory response. Journal of Leukocyte Biology. 2008;84(1):86–92. doi: 10.1189/jlb.0907601. [DOI] [PubMed] [Google Scholar]
- 51.Lauber K, Bohn E, Kröber SM, et al. Apoptotic cells induce migration of phagocytes via caspase-3-mediated release of a lipid attraction signal. Cell. 2003;113(6):717–730. doi: 10.1016/s0092-8674(03)00422-7. [DOI] [PubMed] [Google Scholar]
- 52.Kim SJ, Gershov D, Ma X, Brot N, Elkon KB. I-PLA2 activation during apoptosis promotes the exposure of membrane lysophosphatidylcholine leading to binding by natural immunoglobulin M antibodies and complement activation. The Journal of Experimental Medicine. 2002;196(5):655–665. doi: 10.1084/jem.20020542. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Freire-de-Lima CG, Nascimento DO, Soares MBP, et al. Uptake of apoptotic cells drives the growth of a pathogenic trypanosome in macrophages. Nature. 2000;403(6766):199–203. doi: 10.1038/35003208. Erratum in: Nature, vol. 404, no. 6780, pp. 904, 2000. [DOI] [PubMed] [Google Scholar]
- 54.Belaunzarán ML, Wainszelbaum MJ, Lammel EM, et al. Phospholipase A1 from Trypanosoma cruzi infective stages generates lipid messengers that activate host cell protein kinase c. Parasitology. 2007;134(4):491–502. doi: 10.1017/S0031182006001740. [DOI] [PubMed] [Google Scholar]
- 55.Oishi K, Raynor RL, Charp PA, Kuo JF. Regulation of protein kinase C by lysophospholipids: potential role in signal transduction. The Journal of Biological Chemistry. 1988;263(14):6865–6871. [PubMed] [Google Scholar]
- 56.Murakami M, Taketomi Y, Girard C, Yamamoto K, Lambeau G. Emerging roles of secreted phospholipase A2 enzymes: lessons from transgenic and knockout mice. Biochimie. 2010;92(6):561–582. doi: 10.1016/j.biochi.2010.03.015. [DOI] [PubMed] [Google Scholar]
- 57.Burke JE, Dennis EA. Phospholipase A2 structure/function, mechanism, and signaling. The Journal of Lipid Research. 2009;50:237–242. doi: 10.1194/jlr.R800033-JLR200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Suckling K. Phospholipase A2s: developing drug targets for atherosclerosis. Atherosclerosis. 2010;212(2):357–366. doi: 10.1016/j.atherosclerosis.2010.03.011. [DOI] [PubMed] [Google Scholar]
- 59.Boyanovsky BB, Webb NR. Biology of secretory phospholipase A2 . Cardiovascular Drugs and Therapy. 2009;23(1):61–72. doi: 10.1007/s10557-008-6134-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Stanley D. The non-venom insect phospholipases A2 . Biochimica et Biophysica Acta. 2006;1761(11):1383–1390. doi: 10.1016/j.bbalip.2006.05.011. [DOI] [PubMed] [Google Scholar]
- 61.Rana RL, Wyatt Hoback W, Rahim NAA, Bedick J, Stanley DW. Pre-oral digestion: a phospholipase A2 associated with oral secretions in adult burying beetles, Nicrophorus marginatus . Comparative Biochemistry and Physiology B. 1997;118(2):375–380. [Google Scholar]
- 62.Tunaz H, Stanley DW. Phospholipase A2 in salivary glands isolated from tobacco hornworms, Manduca sexta . Comparative Biochemistry and Physiology B. 2004;139(1):27–33. doi: 10.1016/j.cbpc.2004.05.010. [DOI] [PubMed] [Google Scholar]
- 63.Bowman AS, Gengler CL, Surdick MR, et al. A novel phospholipase A2 activity in saliva of the lone star tick, Amblyomma americanum (L.) Experimental Parasitology. 1997;87(2):121–132. doi: 10.1006/expr.1997.4201. [DOI] [PubMed] [Google Scholar]
- 64.Zhu K, Bowman AS, Dillwith JW, Sauer JR. Phospholipase A2 Activity in Salivary Glands and Saliva of the Lone Star Tick (Acari: Ixodidae) during Tick Feeding. Journal of Medical Entomology. 1998;35(4):500–504. doi: 10.1093/jmedent/35.4.500. [DOI] [PubMed] [Google Scholar]
- 65.Derewenda ZS, Ho YS. PAF-acetylhydrolases. Biochimica et Biophysica Acta. 1999;1441(2-3):229–236. doi: 10.1016/s1388-1981(99)00158-4. [DOI] [PubMed] [Google Scholar]
- 66.Cheeseman MT, Bates PA, Crampton JM. Preliminary characterisation of esterase and platelet-activating factor (PAF)-acetylhydrolase activities from cat flea (Ctenocephalides felis) salivary glands. Insect Biochemistry and Molecular Biology. 2001;31(2):157–164. doi: 10.1016/s0965-1748(00)00113-2. [DOI] [PubMed] [Google Scholar]
- 67.Francischetti IMB, Meng Z, Mans BJ, et al. An insight into the salivary transcriptome and proteome of the soft tick and vector of epizootic bovine abortion, Ornithodoros coriaceus . Journal of Proteomics. 2008;71(5):493–512. doi: 10.1016/j.jprot.2008.07.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Francischetti IMB, Mans BJ, Meng Z, et al. An insight into the sialome of the soft tick, Ornithodorus parkeri . Insect Biochemistry and Molecular Biology. 2008;38(1):1–21. doi: 10.1016/j.ibmb.2007.09.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Mans BJ, Andersen JF, Francischetti IMB, et al. Comparative sialomics between hard and soft ticks: implications for the evolution of blood-feeding behavior. Insect Biochemistry and Molecular Biology. 2008;38(1):42–58. doi: 10.1016/j.ibmb.2007.09.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Francischetti IMB, Pham VM, Mans BJ, et al. The transcriptome of the salivary glands of the female western black-legged tick Ixodes pacificus (Acari: Ixodidae) Insect Biochemistry and Molecular Biology. 2005;35(10):1142–1161. doi: 10.1016/j.ibmb.2005.05.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Calvo E, Dao A, Pham VM, Ribeiro JMC. An insight into the sialome of Anopheles funestus reveals an emerging pattern in anopheline salivary protein families. Insect Biochemistry and Molecular Biology. 2007;37(2):164–175. doi: 10.1016/j.ibmb.2006.11.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Hostomská J, Volfová V, Mu J, et al. Analysis of salivary transcripts and antigens of the sand fly Phlebotomus arabicus . BMC Genomics. 2009;10, article 282 doi: 10.1186/1471-2164-10-282. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Alves-Silva J, Ribeiro JMC, van den Abbeele J, et al. An insight into the sialome of Glossina morsitans morsitans . BMC Genomics. 2010;11(1, article 213) doi: 10.1186/1471-2164-11-213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Zeidner N, Ullmann A, Sackal C, et al. A borreliacidal factor in Amblyomma americanum saliva is associated with phospholipase A2 activity. Experimental Parasitology. 2009;121(4):370–375. doi: 10.1016/j.exppara.2009.01.002. [DOI] [PubMed] [Google Scholar]
- 75.Vargas-Villarreal J, Olvera-Rodríguez A, Mata-Cárdenas BD, Martínez-Rodríguez HG, Said-Fernández S, Alagón-Cano A. Isolation of an Entamoeba histolytica intracellular alkaline phospholipase A2 . Parasitology Research. 1998;84(4):310–314. doi: 10.1007/s004360050401. [DOI] [PubMed] [Google Scholar]
- 76.Passero LFD, Laurenti MD, Tomokane TY, Corbett CEP, Toyama MH. The effect of phospholipase A2 from Crotalus durissus collilineatus on Leishmania (Leishmania) amazonensis infection. Parasitology Research. 2008;102(5):1025–1033. doi: 10.1007/s00436-007-0871-6. [DOI] [PubMed] [Google Scholar]
- 77.Bertello LE, Alves MJM, Colli W, de Lederkremer RM. Evidence for phospholipases from Trypanosoma cruzi active on phosphatidylinositol and inositolphosphoceramide. Biochemical Journal. 2000;345(1):77–84. [PMC free article] [PubMed] [Google Scholar]
- 78.Kini RM. Structure-function relationships and mechanism of anticoagulant phospholipase A2 enzymes from snake venoms. Toxicon. 2005;45(8):1147–1161. doi: 10.1016/j.toxicon.2005.02.018. [DOI] [PubMed] [Google Scholar]
- 79.Rigoni M, Caccin P, Gschmeissner S, et al. Neuroscience: equivalent effects of snake PLA2 neurotoxins and lysophospholipid-fatty acid mixtures. Science. 2005;310(5754):1678–1680. doi: 10.1126/science.1120640. [DOI] [PubMed] [Google Scholar]
- 80.Silva-Cardoso L, Caccin P, Magnabosco A, et al. Paralytic activity of lysophosphatidylcholine from saliva of the waterbug Belostoma anurum . Journal of Experimental Biology. 2010;213(19):3305–3310. doi: 10.1242/jeb.041954. [DOI] [PubMed] [Google Scholar]
- 81.Stanley-Samuelson DW, Jensen E, Nickerson KW, Tiebel K, Ogg CL, Howard RW. Insect immune response to bacterial infection is mediated by eicosanoids. Proceedings of the National Academy of Sciences of the United States of America. 1991;88(3):1064–1068. doi: 10.1073/pnas.88.3.1064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Stanley-Samuelson DW, Pedibhotla VK. What can we learn from prostaglandins and related eicosanoids in insects? Insect Biochemistry and Molecular Biology. 1996;26(3):223–234. doi: 10.1016/0965-1748(95)00092-5. [DOI] [PubMed] [Google Scholar]
- 83.Ashton AW, Mukherjee S, Nagajyothi FNU, et al. Thromboxane A2 is a key regulator of pathogenesis during Trypanosoma cruzi infection. The Journal of Experimental Medicine. 2007;204(4):929–940. doi: 10.1084/jem.20062432. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Grillo LAM, Majerowicz D, Gondim KC. Lipid metabolism in Rhodnius prolixus (Hemiptera: reduviidae): role of a midgut triacylglycerol-lipase. Insect Biochemistry and Molecular Biology. 2007;37(6):579–588. doi: 10.1016/j.ibmb.2007.03.002. [DOI] [PubMed] [Google Scholar]
- 85.Wilensky RL, Shi Y, Mohler ER, et al. Inhibition of lipoprotein-associated phospholipase A2 reduces complex coronary atherosclerotic plaque development. Nature Medicine. 2008;14(10):1059–1066. doi: 10.1038/nm.1870. [DOI] [PMC free article] [PubMed] [Google Scholar]