

Abstract
Quantifiers are very common in everyday speech, but we know little about their cognitive basis or neural representation. The present study examined comprehension of three classes of quantifiers that depend on different cognitive components in patients with focal neurodegenerative diseases. Patients evaluated the truth-value of a sentence containing a quantifier relative to a picture illustrating a small number of familiar objects, and performance was related to MRI grey matter atrophy using voxel-based morphometry. We found that patients with corticobasal syndrome (CBS) and posterior cortical atrophy (PCA) are significantly impaired in their comprehension of Cardinal Quantifiers (e.g. “At least three birds are on the branch”), due in part to their deficit in quantity knowledge. MRI analyses related this deficit to temporal-parietal atrophy found in CBS/PCA. We also found that patients with behavioral variant frontotemporal dementia (bvFTD) are significantly impaired in their comprehension of Logical Quantifiers (e.g. “Some the birds are on the branch”), associated with a simple form of perceptual logic, and this correlated with their deficit on executive measures. This deficit was related to disease in rostral prefrontal cortex in bvFTD. These patients were also impaired in their comprehension of Majority Quantifiers (e.g. “At least half of the birds are on the branch”), and this too was correlated with their deficit on executive measures. This was related to disease in the basal ganglia interrupting a frontal-striatal loop critical for executive functioning. These findings suggest that a large-scale frontal-parietal neural network plays a crucial role in quantifier comprehension, and that comprehension of specific classes of quantifiers may be selectively impaired in patients with focal neurodegenerative conditions in these areas.
Keywords: comprehension, quantifier, parietal, frontal, corticobasal, frontotemporal dementia
INTRODUCTION
Generalized quantifiers such as “at least 3 X,” “less than half of the X,” and “some X” are extraordinarily common in day-to-day speech. Yet, we know little about the cognitive and neural basis for quantifiers. In this study, we evaluated several classes of quantifiers in patients with focal neurodegenerative disease to investigate the cognitive and neural basis for difficulty understanding quantifiers.
Studies of lexical semantic memory generally have focused on words that refer to objects. A major recent concern of this research has been to determine the contribution of sensory-motor features of objects to the mental representation of word meaning (Barsalou, Simmons, Barbey, & Wilson, 2003; Martin, 2007). By comparison, quantifier meaning emphasizes several components that do not depend on the sensory-motor system (Clark & Grossman, 2007). Foremost among these is that knowledge of quantity and magnitude appears to play a central role in meaning associated with the quantifier segment of the lexicon. For example, quantifier meaning depends on mapping a truth-value to a set of objects or to a quantity of a mass substance. This and several other components of quantifier meaning appear to play a relatively prominent role in each of the three major classes of quantifiers that we discuss below and evaluate comparatively in this study.
Consider first cardinal quantifiers like “at least three X.” This class of quantifiers is thought to depend largely on knowledge of quantity or magnitude (Keenan, Westersthål, van Benthem, & ter Meulen, 1997; van Benthem, 1986). Knowledge of a precise number in a statement like “There are three birds on the branch” can be demonstrated by matching the cardinality of the number to the quantity of the named objects in a picture. Thus, using a one-to-one correspondence rule as in counting, the states of “1 bird,” “2 birds,” and then “3 birds” can be matched precisely to the number of birds in the picture. To assess the truth-value of a statement with a cardinal quantifier like “There are at least three birds on the branch,” a listener must assess the states of “1 bird”, “2 birds,” and “3 birds,” and then determine whether there are additional states of magnitude on a number line beyond those associated with the state of three birds on the branch.
Contrast cardinal quantifiers with majority quantifiers. Majority quantifiers like “at least half of the X” appear to depend in part on knowledge of quantity, but this class of quantifiers also appears to involve executive resources such as working memory and strategic processing (McMillan, Clark, Moore, DeVita, & Grossman, 2005). In a sentence like “At least half of the birds are on the branch,” for example, a magnitude component determines the cardinality corresponding to the subset of target objects “birds on the branch” in a picture. This quantity is maintained in working memory while the cardinality corresponding to “half” of all of the birds in the picture is established. The cardinality of “half” can be established by counting pairs of objects, dividing the total number of objects by two, or several other methods. Then a comparison can be made between the quantity of target objects and the quantity corresponding to “half” of all of the objects. If the quantity of the subset exceeds the quantity of half, then the truth-value of the statement is satisfied.
Another class of quantifiers is known as “logical” or “Aristotelean.” Logical quantifiers like “some” and “all” do not seem to involve number knowledge per se. Instead, a statement like “There are some birds on the branch” appears to depend in part on a simple system of perceptual logic that establishes dichotomously the presence or absence of a single target object (Troiani, Peelle, Clark, & Grossman, 2008). As soon as one bird is seen on the branch, then the truth-value of “some” is satisfied. This statement is no truer when two birds are on the branch, and is falsified only when no birds are on the branch. Conversely, the truth-value of a statement like “All of the birds are on the branch” is satisfied only if every one of the birds is on the branch, is falsified even if one bird is not on the branch, and is no more false when two birds are not on the branch.
Previous work suggested that comprehension of these classes of quantifiers is selectively compromised in patients with focal neurodegenerative diseases (McMillan, Clark, Moore, & Grossman, 2006; Troiani, et al., 2008). Thus, patients with corticobasal syndrome (CBS) had difficulty judging whether a sentence containing a quantifier accurately describes isolated objects presented as a spatial array or a sequence. We reasoned that these patients are significantly impaired with cardinal quantifiers in part because of their deficit with number knowledge (Halpern, et al., 2003; Halpern, et al., 2004; Koss, et al., 2010), although impairments of visuospatial processing or working memory needed to track a series of objects may have interfered with performance. We addressed some of these potential confounds in a recent study that assessed quantifiers in a more naturalistic, strictly verbal context. Non-aphasic CBS patients thus were impaired at judging the accuracy of sentences containing quantifiers concerned with familiar concepts such as the number of days in a week or the number of inches in a foot (Troiani, Clark, & Grossman, 2011). However, we have not directly compared quantifier comprehension with a similar evaluation of precise number knowledge, nor have we verified that the patients can map a sentence containing a quantifier to a naturalistic picture of a scene. The present study extended this work by examining quantifier comprehension with a sentence-picture matching task that uses the same set of naturalistic visual scenes to assess different classes of quantifiers as well as precise numbers. We also examine performance involving sentence stimuli without a quantifier or a number to verify that language and visual-perceptual deficits cannot explain patients’ deficits. Because posterior cortical atrophy (PCA) is associated with disease in temporal-parietal cortical regions as well (Alladi, et al., 2007; Hof, Vogt, Bouras, & Morrison, 1997; Nestor, Caine, Fryer, Clarke, & Hodges, 2003; Victoroff, Ross, Benson, & Verity, 1994), we also evaluated performance in some patients with PCA.
Logical quantifiers, by comparison, are selectively compromised in patients with behavioral variant frontotemporal dementia (bvFTD) (Troiani, et al., 2008). This appears to be related in part to their executive resource limitations (Kramer, Jurik, & Sha, 2003; Libon, et al., 2007). Patients with bvFTD and Alzheimer’s disease (AD) are impaired with majority quantifiers (McMillan, et al., 2006; Troiani, et al., 2008). While these patients have modest difficulty with numbers, their deficit instead appears to be related to their executive difficulty (Kramer, et al., 2003; Libon, et al., 2008; Libon, et al., 2007). CBS patients also may have some difficulty with majority quantifiers, due in part to the number component of this class of quantifiers (McMillan, et al., 2006; Troiani, et al., 2008).
From the perspective of our model (Clark & Grossman, 2007), these patterns of quantifier impairment are related largely to the anatomic distribution of disease associated with these focal neurodegenerative conditions, as illustrated schematically in Figure 1. Each of these diseases thus selectively interrupts a large-scale frontal-parietal network responsible for quantifier processing in a different way. Imaging (Grossman, et al., 2004; Whitwell, et al., 2007) and histopathological (Josephs, et al., 2006; Murray, et al., 2007) studies show that disease in CBS involves at least parietal and posterior temporal regions, as well as the basal ganglia. A recent clinical-pathological case series demonstrated a disorder of number knowledge in patients with focal parietal atrophy due to autopsy-proven corticobasal degeneration (Pantelyat et al., 2011). Patient (Ashkenazi, Henik, Ifergane, & Shelef, 2008; Cappelletti, Barth, Fregni, Spelke, & Pascual-Leone, 2007; Cappelletti, Muggleton, & Walsh, 2009; Dellatolas, Deloche, Basso, & Claros-Salinas, 2001; Gobel, Calabria, Farne, & Rossetti, 2006; Martory, et al., 2003), fMRI (Ansari, Dhital, & Siong, 2006; Castelli, Glaser, & Butterworth, 2006; Piazza, Mechelli, Price, & Butterworth, 2006; Shuman & Kanwisher, 2004) and transcranial magnetic stimulation (TMS) (Cappelletti, et al., 2007; Cappelletti, et al., 2009) studies emphasize that inferior parietal and posterior-superior temporal regions, including the intraparietal sulcus, play a crucial role in processing magnitude and quantity. This is consistent with classical localizationist teaching about acalculia. fMRI studies of healthy adults also relate comprehension of cardinal quantifiers to temporal-parietal activation (McMillan, et al., 2005; Troiani, et al., 2008). Since temporal-parietal cortex supports the processing of quantity, and since quantity plays a critical role in cardinal quantifiers, our model predicts that temporal-parietal disease should interfere with the comprehension of cardinal quantifiers in CBS and PCA (McMillan, et al., 2006; Troiani, et al., 2008).
FIGURE 1.
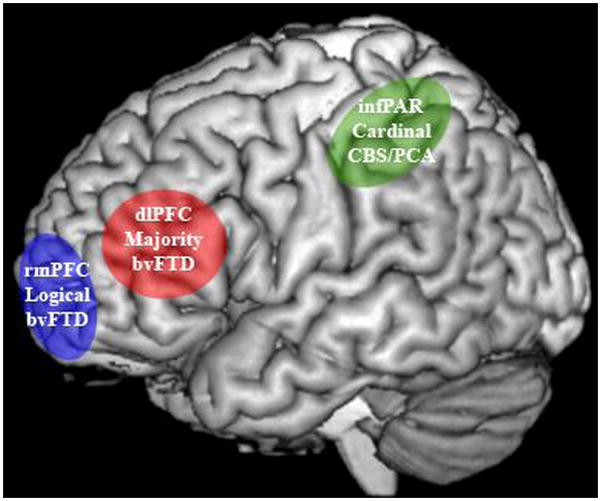
DIAGRAM OUTLINING THE NEUROCOGNITIVE NETWORK FOR QUANTIFIER COMPREHENSION1
NOTE
1. Marked areas represent anatomic regions that play a central role for a specific aspect of quantifier meaning that is compromised by disease in a particular patient group. Green: inferior parietal cortex critical for quantity and magnitude knowledge that is compromised in CBS; blue: rostral medial prefrontal cortex critical for dichotomous perceptual decision-making that is compromised in bvFTD; red: dorsolateral prefrontal cortex critical for working memory and strategic planning that is compromised in bvFTD.
By comparison, patients with bvFTD have disease that involves rostral and medial portions of prefrontal cortex (rmPFC). This is seen in imaging studies (Grossman, et al., 2004; Kipps, Hodges, Fryer, & Nestor, 2009) and assessments of regional histopathologic burden (Forman, et al., 2006; Grossman, et al., 2007). One fMRI study of logical quantifiers in healthy adults showed rmPFC activation (Troiani, et al., 2008). Likewise, fMRI studies of healthy adults relate activations for decisions based on simple perceptual logic to rmPFC (Burgess, Dumontheil, & Gilbert, 2007; Gilbert, Frith, & Burgess, 2005). Since this facet of executive control contributes crucially to comprehension of logical quantifiers, our model predicts that disease in rostral and medial portions of PFC should interfere with comprehension of this class of quantifiers (Troiani, et al., 2008). However, disease in this anatomic distribution should not necessarily interfere with comprehension of cardinal quantifiers, just as parietal disease in CBS should not compromise comprehension of logical quantifiers.
Finally, bvFTD is also associated with disease that includes ventral and dorsal portions of lateral PFC as well as the striatum (Forman, et al., 2006; Grossman, et al., 2007; Grossman, et al., 2004; Kipps, et al., 2009). Disease in this distribution interrupts a frontal-striatal loop that appears to play a critical role in executive resources like working memory and strategic comparisons in studies of patients with brain damage (Baddeley, Della Sala, Papagno, & Spinnler, 1997; Brown, Soliveri, & Jahanshahi, 2000) and fMRI studies of healthy adults (Badre & Wagner, 2004; Smith & Jonides, 2002). fMRI studies of healthy adults show lateral PFC activation during comprehension of majority quantifiers (McMillan, et al., 2005; Troiani, et al., 2008). Since executive resources play a central role in the comprehension of majority quantifiers, our model predicts that executive deficits in bvFTD should interfere with majority quantifier comprehension (McMillan, et al., 2006; Troiani, et al., 2008) as well.
METHODS
Subjects
We studied 27 non-aphasic, right-handed English-speakers with a neurodegenerative disease. Fourteen patients had the clinical diagnosis of bvFTD, and 13 patients had CBS (n = 10) or PCA (n = 3). bvFTD subjects were diagnosed according to published criteria (McKhann, et al., 2001; Rascovsky, et al., 2007). They had a progressive disorder of social comportment and personality characterized by disinhibition, ritualistic behavior, difficulty with initiation, lack of empathy, and impoverished insight into their difficulties. While there are no published consensus clinical criteria for the diagnosis of CBS, these patients were diagnosed according to published expert opinions (Riley, Lang, Litvan, Goetz, & Lang, 2000) and clinical-pathological studies of patients with autopsy-proven CBD (Boeve, et al., 1999; Grimes, Lang, & Bergeron, 1999; Murray, et al., 2007). These patients had an asymmetric extrapyramidal syndrome involving features such as rigidity, dystonia and “alien limb” phenomena, together with progressive ideomotor apraxia and cortical sensory loss. Likewise, there are no published clinical consensus criteria for PCA, and we based the diagnosis on clinical-pathological cases emphasizing the presence of progressive visuospatial difficulty without memory impairment (Alladi, et al., 2007; Hof, et al., 1997; Nestor, et al., 2003; Victoroff, et al., 1994). Patients with other cognitive deficits such as progressive aphasia or an amnestic syndrome were excluded, and patients with a neurologic condition such as stroke or hydrocephalus, a primary psychiatric disorder or a medical condition causing cognitive difficulty were excluded. Clinical and demographic criteria are summarized in Table 1. We also studied 12 healthy, right-handed native English-speaking seniors. All subjects participated in accordance with an informed consent procedure approved by the University of Pennsylvania Institutional Review Board.
TABLE 1.
MEAN ± S.D. CLINICAL AND DEMOGRAPHIC CHARACTERISTICS, AND PERFORMANCE ON EXECUTIVE MEASURES1
| Age | Education | MMSE | Summary Executive Z Score | FAS Z Score | Stroop Formula Z Score | Trails Formula Z Score | |
|---|---|---|---|---|---|---|---|
| HEALTHY SENIORS | 65.9 ± 8.8 | 15.0 ± 2.7 | 28.9 ± 1.0 | - | - | - | - |
| CORTICOBASAL SYNDROME/POSTERIOR CORTICAL ATROPHY | 65.4 ± 8.4 | 15.6 ± 4.6 | 25.9 ± 3.3* | −1.67 ± 1.6 | −1.24 ± 0.9 | −2.74 ± 1.7* | −1.15 ± 2.6 |
| BEHAVIORAL VARIANT FRONTOTEMPORAL DEMENTIA | 64.9 ± 8.8 | 14.6 ± 2.6 | 23.2 ± 4.6* | −1.54 ± 1.2 | −1.47 ± 1.2 | −2.17 ± 1.3* | −1.23 ± 1.4 |
NOTE
* Indicates significant difference from healthy seniors at p<0.05.
We examined the participants with a brief neuropsychological battery. Performance is summarized in Table 1 as well. All neuropsychological tests could not be administered to all participants because of limited patient availability or technical difficulty. We indicate in parentheses the numbers of participants in each group who were evaluated with each measure.
Letter Fluency (FAS)
Participants stated as many unique F, A, and S words as possible during one minute for each letter. Intrusions and perseverations were not included in total scores. The average number of words per minute was calculated by combining the total number of F, A, and S words and dividing by 3.
Stroop color-name reading
Participants named the color of the font of 50 color names, where the color of the font did not match the named color (e.g. “RED” printed in green font). A Stroop Formula was calculated as total correct answers minus incorrect answers (self-corrected and uncorrected) and then divided by the time to completion (maximum = 300 sec).
Trails B
Participants used a pencil to draw a line connecting an ascending series of alternating letters and numbers (A-1-B-2…) randomly arrayed on an 8×11” sheet of paper. A Trails Formula was calculated as total correct answers minus incorrect answers (self-corrected and uncorrected) and then divided by the time to completion (maximum = 300 sec).
Patient group z-scores were calculated for all neuropsychological data relative to performance by a local group of matched healthy seniors (n=25). A total executive score was calculated by averaging z-scores from FAS, Stroop and Trails B.
Materials
Subjects were presented with 100 brief, grammatically simple statements and asked to judge the truth-value of each statement relative to a colored picture illustrating a naturalistic scene containing a small number of familiar objects. We used a single picture as a stimulus for a set of sentences containing the three classes of quantifiers. All of the sentence stimuli are provided in the Appendix. Each sentence contained familiar words referring to familiar, countable objects illustrated in the picture (e.g. birds, cows). A total of three types of quantifier statements were presented in 20 trials each, where half of the statements for each type of quantifier were true statements and half were false.
To minimize the risk of confounding poor comprehension of quantifiers with difficulty due to limited quantity knowledge per se, we used small numbers within the subitizing range that are better understood by CBS patients than numbers in the larger, counting range (Halpern, Clark, Moore, Cross, & Grossman, 2007). Each picture thus contained between two and five objects, and the targeted subset of objects differed from the remainder by one. To show that there is no difference in performance as a function of numerosity, we compared performance within each group for each quantifier for stimuli with a smaller numerosity (i.e. ≤3) compared to stimuli with a larger numerosity (i.e. ≥4). None of these differences were significant (all contrasts p>0.05). Moreover, the use of the same picture for a set of sentences containing different quantifiers underlined that any difference in comprehension is due to the particular quantifier in a statement rather than the quantity or nature of the objects in a sentence. The statements were matched for length across conditions by the addition of neutral adjectives.
The three types of statements included: cardinal, logical and majority quantifiers. Cardinal quantifiers [1] depend largely on quantity knowledge.
[1] There are fewer than three birds on the branch
There are more than two cows in the barn
Logical quantifiers [2] depend on a simple, perceptual logic that involves decisions about dichotomous stimulus properties.
[2] Some of the books are on the shelf
Not all the cows are in the barn
Majority quantifiers [3] depend on quantity knowledge as well as executive resources such as working memory.
[3] Less than half the pumpkins are in the truck
More than half the birds are on the branch
We also administered two sets of control items. These used the same pictures as the quantifier sentences. In the first set, 20 sentences (half correct, half incorrect) probed a precise number [4] in the picture.
[4] There are three birds on the branch
There are two cows in the barn
Additionally, 20 control statements [5] containing no quantifier or number component served to probe comprehension of the visual pictures and sentence content (half correct, half incorrect).
[5] There are birds on the branch
There are books on the shelf
All stimuli were displayed using a Dell Inspiron 1100 laptop. E-Prime v2.0 presentation software controlled stimulus presentation and recorded response accuracy. After a 500 msec fixation target, a statement appeared at the bottom of the screen for 3000 msec. Text was displayed in Arial 18 font. Next, the statement remained at the bottom of the screen and a picture appeared above the text for another 10,000 msec. Participants were asked to judge whether the sentence described the picture correctly. “Yes” or “no” responses were recorded by pressing one of two buttons on the computer keyboard. Prior to the experiment, participants were given a practice run consisting of 10 trials. During the practice trials, incorrect responses were corrected and explained by the experimenter in order to ensure comprehension of task instructions. All patients appeared to understand the task.
Statistical analyses were performed with SPSS v12.0 (Statistical Package for the Social Sciences, Chicago, IL). We used t-tests to evaluate between-group and within-group performance. Because of ceiling effects in control subjects and in patients for conditions where performance was relatively preserved, we adjusted the degrees of freedom to account for the unequal distributions. We used a Spearman nonparametric correlation to relate cardinal quantifier comprehension to a measure of quantity knowledge – our “precise” condition described above – because performance on the precise condition was at ceiling in bvFTD. We also correlated neuropsychological performance on the executive measures with comprehension of logical and majority quantifiers.
Imaging methods
High resolution structural MRI scans were available in 16 patients, including 8 patients with CBS/PCA and 8 patients with bvFTD. T1-weighted MRI images were acquired with a Siemens 3T Trio scanner with 1-mm slice thickness and a 192 × 256 matrix using an MPRAGE protocol (TR = 1620 ms, TE = 3 ms, flip angle = 15°, resolution = .9766 × .9766 × 1 mm). We used PipeDream (https://sourceforge.net/projects/neuropipedream/) and Advanced Normalization Tools (ANTS, http://www.picsl.upenn.edu/ANTS/) tools to perform the most stable and reliable multivariate imaging normalization and structure-specific processing currently available (Avants, Epstein, Grossman, & Gee, 2008; Avants, et al., 2010; Klein, et al., 2009). PipeDream deforms each individual dataset into a standard local template space in a canonical stereotactic coordinate system. Core processing involves mapping T1 structural MRI to a population-specific, unbiased, average-shape and -appearance image at 1mm3 resolution derived from a representative local population consisting of 25 healthy seniors and 25 FTLD patients (Kim, et al., 2008). The pipeline begins by registering the subject image to the local template, after which the subject space can be mapped directly to MNI space by combining the subject-to-template and template-to-MNI transformations. The coordinate deformation is diffeomorphic – that is, smooth and invertible, symmetric so that it is not biased towards the reference space for computing the mappings, and topology-preserving to capture the large deformation necessary to aggregate images in a common space (Avants & Gee, 2004; Avants, et al., 2010). Next, three-tissue (grey matter, white matter, and cerebrospinal fluid) segmentation is performed in subject space using the Atropos tool in ANTS. Prior probability images for each tissue class, previously defined in the local template, are warped into the subject image to guide the segmentation. These algorithms allow template-based priors to guide cortical segmentation and compute grey matter density (Das, Avants, Grossman, & Gee, 2009). Grey matter density images are smoothed in SPM5 (http://www.fil.ion.ucl.ac.uk/spm/software/spm5) using a 4 mm full-width half-maximum Gaussian kernel to minimize individual gyral variations.
In SPM5, two sample t-tests identified areas of significant cortical atrophy in CBS and bvFTD by contrasting grey matter density maps between each patient group and 40 healthy, age-matched controls. An explicit mask defined by a grey matter prior probability map in SPM5 limited the analysis to voxelwise comparisons within grey matter. The analysis of atrophy contrasting CBS/PCA and control groups was performed at a p<0.001 level, including all clusters surviving a 400 voxel extent criterion, and a voxel-level height threshold criterion within each cluster of p<0.001. There was a substantial discrepancy in the total number of significantly atrophic voxels in CBS/PCA and bvFTD patients relative to controls at the p<0.001 level, and we sought to minimize the risk that this difference would bias regression analyses relating comprehension performance to grey matter atrophy. Thus, we report the atrophy analysis of bvFTD relative to controls conducted at p<0.005 so that the total number of atrophic voxels is relatively equivalent in the contrasts of the two patient groups relative to controls. SPM5 was used to perform regression analyses relating comprehension accuracy for each class of quantifier to grey matter atrophy. We interpreted these regressions only in areas surviving a statistical map consisting of the area of significant grey matter atrophy in each group because it is only these areas that are abnormal in the patients. Each of these analyses was conducted at p<0.05, unless otherwise stated, and we used statistical criterion of a peak voxel z-score > 3.09 (equivalent to p<0.001) and a 200 voxel cluster extent.
RESULTS
Behavioral Results
As summarized in Figure 2, CBS/PCA patients had a selective deficit for cardinal quantifiers compared to healthy seniors (t=2.75, df= 17.9; p=0.013). The deficit for cardinal quantifiers was evident in both CBS (mean ±SE = 82.5±18.7% correct) and PCA (mean ±SE = 61.7±18.9% correct) subgroups of this cohort. Moreover, only CBS/PCA patients (mean ±SE = 76.9 ±15% correct) differed significantly from healthy seniors (mean ±SE = 96.7±5.7% correct) in their comprehension of precise stimuli that assess knowledge of quantity (t=2.17, df=15.7; p=0.046). We also found a significant correlation between comprehension of cardinal quantifiers and quantity knowledge as measured by comprehension of the precise number stimuli (s=0.563; df=13; p=0.045). bvFTD patients equaled healthy seniors in their comprehension of cardinal quantifiers (p>0.05). bvFTD patients (mean ±SE = 94.3±8.1% correct) also did not differ from controls in the performance with precise stimuli (p>0.05). Finally, there was no correlation between cardinal comprehension accuracy and precise number knowledge in bvFTD.
FIGURE 2.
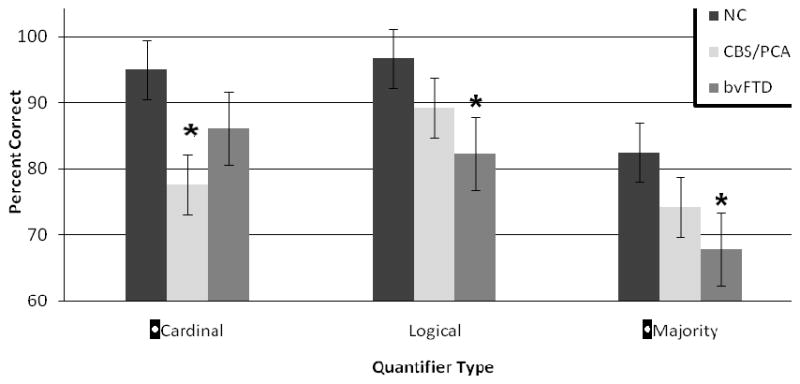
MEAN (±S.D.) QUANTIFIER COMPREHENSION ACCURACY IN CORTICOBASAL SYNDROME/POSTERIOR CORTICAL ATROPHY AND BEHAVIORAL VARIANT FRONTOTEMPORAL DEMENTIA1
NOTE
1. * indicates a significant difference (p<0.05) relative to controls.
By comparison, Figure 2 shows that bvFTD patients were impaired in their comprehension of logical quantifiers (t=2.77, df=13.9; p=0.015) and majority quantifiers (t=2.38, df=21.1; p=0.027) relative to healthy seniors. We also found a significant correlation between comprehension of logical quantifiers and the summary measure of executive functioning (s=0.658; df=11; p=0.028). A correlation between majority quantifier comprehension and the summary measure of executive functioning approached significance (s=0.590, df=11, p=0.056). Additionally, comprehension of logical quantifiers and majority quantifiers were significantly correlated with each other (s=0.649; df=11; p=0.012). However, the CBS/PCA group did not differ statistically from the healthy seniors group in their comprehension of logical and majority quantifiers (p>0.05), and there was no correlation between comprehension of logical and majority quantifiers and performance on measures of executive functioning in CBS/PCA patients (p>0.05).
It is noteworthy that healthy seniors (mean ±SE = 93.3±7.2% correct), CBS/PCA (mean ±SE = 90.4 ±10% correct) and bvFTD (mean ±SE = 84.3 ±14% correct) groups did not differ statistically in their comprehension of control stimuli that do not contain a quantifier or a number.
Imaging Results
Figure 3 shows the distribution of significant cortical atrophy, and the peaks of significant clusters are summarized in Table 2. Figure 3 Panel A shows that the CBS/PCA group had significant atrophy in a parietal and posterior temporal distribution bilaterally, extending anteriorly into the peri-Sylvian region. The basal ganglia also showed atrophy bilaterally. By comparison, Figure 3 Panel B shows that cortical atrophy in bvFTD patients was most prominent in a frontal distribution bilaterally, including lateral, medial and ventral frontal regions. Atrophy extended into the right temporal lobe as well.
FIGURE 3.
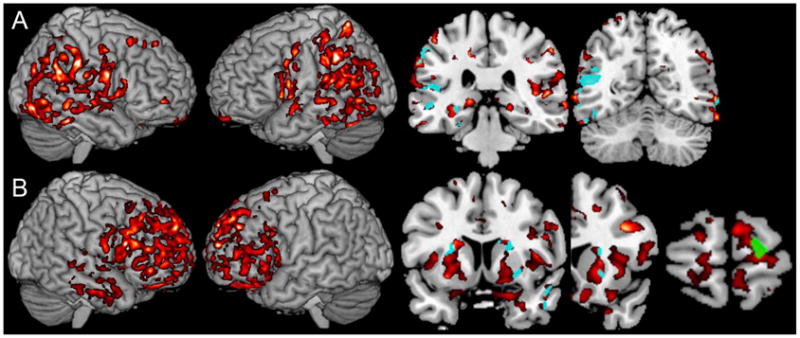
GREY MATTER ATROPHY AND REGRESSION ANALYSES FOR QUANTIFIER COMPREHENSION ACCURACY IN CORTICOBASAL SYNDROME AND BEHAVIORAL VARIANT FRONTOTEMPORAL DEMENTIA1
NOTE
1. Panel A: Corticobasal syndrome and posterior cortical atrophy. The left portion of the panel shows significant cortical atrophy in CBS/PCA; the right portion of the panel shows regions of cortical atrophy related to cardinal quantifier comprehension difficulty by regression analysis (aqua) in coronal slices at y=−31 and y=−59; Panel B: Behavioral variant frontotemporal dementia. The left portion of the panel shows significant cortical atrophy in bvFTD; the right portion of the panel shows regions of cortical atrophy related to logical quantifier comprehension difficulty by regression analysis (green) at coronal slice y=66; aqua shows regions of cortical atrophy related to majority quantifier comprehension difficulty by regression analysis at coronal slices y=18 and y=8.
TABLE 2.
IMAGING COORDINATES FOR GREY MATTER ATROPHY AND REGRESSION ANALYSES RELATING ATROPHY TO QUANTIFIER COMPREHENSION ACCURACY
| Anatomic Locus (Brodmann Area) | Coordinates | z-score | Cluster Size (voxels) | ||
|---|---|---|---|---|---|
| x | y | z | |||
| CORTICOBASAL SYNDROME/POSTERIOR CORTICAL ATROPHY: GREY MATTER ATROPHY | |||||
| R lateral parietal (40) | 61 | −22 | 34 | 6.16 | 16270 |
| R lateral parietal (19) | 29 | −42 | −3 | 5.55 | 2704 |
| R medial parietal (31) | 11 | −45 | 31 | 5.03 | 524 |
| R parietal-occipital (19) | 50 | −72 | −5 | 5.52 | 5801 |
| R parietal-occipital (18) | 23 | −76 | −6 | 4.48 | 558 |
| L temporal-parietal (39/22) | −46 | −53 | 13 | 5.84 | 22007 |
| L lateral parietal (7) | −25 | −52 | 66 | 5.59 | 1113 |
| L lateral parietal (40) | −48 | −28 | 52 | 4.81 | 752 |
| R dorsal frontal (8) | 17 | 42 | 45 | 4.76 | 403 |
| R dorsal frontal (8) | 36 | 21 | 51 | 4.49 | 428 |
| R ventral frontal (11/47) | 30 | 30 | −17 | 4.19 | 486 |
| R medial frontal (10) | 5 | 52 | 11 | 5.02 | 444 |
| R medial frontal (11) | 2 | 52 | −20 | 4.20 | 511 |
| R ventral frontal (11/10) | 32 | 49 | −6 | 4.86 | 419 |
| L medial frontal (11) | −9 | 16 | −27 | 3.88 | 418 |
| L insula | −22 | 14 | 14 | 5.69 | 5637 |
| L dorsal cingulate (24/31) | −14 | −16 | 44 | 5.42 | 604 |
| R dorsal temporal (21/22) | 50 | −27 | −1 | 5.51 | 670 |
| R ventral temporal (21/20) | 62 | −34 | −15 | 5.42 | 1120 |
| L middle temporal (21) | −58 | 2 | −9 | 4.64 | 539 |
| L ventral temporal (21/20) | −67 | −36 | −13 | 4.61 | 683 |
| L parahippocampal (36/35) | −29 | −23 | −21 | 5.46 | 2791 |
| R precuneus (31) | 17 | −65 | 27 | 4.44 | 673 |
| L caudate | 22 | 18 | 3 | 5.55 | 7663 |
| L thalamus | −17 | −30 | 2 | 4.77 | 525 |
| CORTICOBASAL SYNDROME/POSTERIOR CORTICAL ATROPHY: REGRESSION FOR CARDINAL QUANTIFIER COMPREHENSION ACCURACY | |||||
| R temporal-parietal (37) | 60 | −57 | −6 | 4.08 | 321 |
| R temporal-parietal (37) | 38 | −46 | −11 | 3.12 | 238 |
| R occipital (18) | 42 | −85 | −2 | 3.88 | 719 |
| R occipital (18) | 18 | −82 | −2 | 3.70 | 567 |
| L parietal (40) | −56 | −31 | 45 | 4.05 | 362 |
| L temporal-parietal (39/22) | −48 | −42 | 14 | 4.08 | 1948 |
| L temporal-parietal (21/22) | −65 | −49 | 5 | 3.94 | 361 |
| L parietal-occipital (39/19) | −48 | −72 | 15 | 3.98 | 370 |
| L temporal-occipital (37/19) | −56 | −61 | −6 | 4.13 | 414 |
| L temporal-occipital (19) | −46 | −69 | −7 | 4.41 | 1466 |
| L temporal-occipital (37/19) | −48 | −67 | 8 | 3.88 | 247 |
| L temporal (21) | −50 | −34 | 2 | 4.15 | 467 |
| L parahippocampal (36) | −28 | −25 | −17 | 3.77 | 317 |
| L parahippocampal (36/19) | −22 | −41 | −4 | 3.72 | 370 |
| L insula (42/22) | −55 | −31 | 20 | 3.37 | 1221 |
| B. BEHAVIORAL VARIANT FRONTOTEMPORAL DEMENTIA | |||||
|---|---|---|---|---|---|
| Anatomic Locus (Brodmann Area) | Coordinates | z-score | Cluster Size (voxels) | ||
| x | y | z | |||
| BEHAVIORAL VARIANT FRONTOTEMPORAL DEMENTIA: GREY MATTER ATROPHY | |||||
| Bilateral frontal (10/32) | 24 | 47 | 13 | 5.95 | 93671 |
| R frontal (11) | 32 | 49 | −6 | 4.64 | 1659 |
| BEHAVIORAL VARIANT FRONTOTEMPORAL DEMENTIA: REGRESSION FOR LOGICAL QUANTIFIER COMPREHENSION ACCURACY | |||||
| R frontal (10) | 17 | 62 | 7 | 2.97 | 258 |
| BEHAVIORAL VARIANT FRONTOTEMPORAL DEMENTIA: REGRESSION FOR MAJORITY QUANTIFIER COMPREHENSION ACCURACY | |||||
| R temporal (22/21) | 55 | −27 | 3 | 3.58 | 3365 |
| R frontal (8) | |||||
| R insula | 32 | 3 | −1 | 3.56 | 801 |
| R putamen | 25 | 6 | 14 | 3.48 | 532 |
| L putamen | −23 | −2 | 7 | 3.94 | 747 |
We related quantifier comprehension accuracy to grey matter atrophy in CBS/PCA with regression analyses. Figure 3 Panel A shows a significant association of cardinal quantifier comprehension accuracy (aqua) with atrophy in parietal and temporal regions bilaterally in a distribution that overlaps with their significant temporal-parietal atrophy. Although the regression analysis examining logical quantifier comprehension accuracy was not significant in bvFTD at the p=0.05 level, we pursued a more liberal analysis at p<0.1 to examine our a priori hypothesis relating logical quantifier comprehension to rostral prefrontal cortex. Figure 3 Panel B shows that logical quantifier comprehension accuracy in bvFTD (green) was related to rostral prefrontal atrophy. We also used regression to relate comprehension of majority quantifiers (aqua) to cortical atrophy in bvFTD. When the regression was performed at p<0.05, we found statistically significant associations with frontal insula and putamen regions, and the lateral temporal region of the right hemisphere. When we examined our hypothesis at the p<0.1 level, the regression analysis revealed an additional association in the lateral prefrontal region of the right hemisphere.
DISCUSSION
Quantifiers are extraordinarily common in our daily speech, yet we know little about the cognitive and neural basis for this segment of our vocabulary. Previous findings used judgments of sequences or spatial arrays of isolated objects to suggest a deficit in quantifier comprehension. This was recently extended to the comprehension of familiar verbal concepts like days in a week and inches in a foot. The present study examined three classes of quantifiers as well as precise numbers using pictures of familiar objects in a natural context in patients with CBS and PCA who have parietal disease and in patients with bvFTD who have prefrontal disease. The results of the present study are consistent with our multi-component model of quantifier comprehension. One class of quantifier, known as cardinal quantifiers, depends in large part on knowledge of quantity. Cardinal quantifiers are compromised in patients with CBS and PCA who have degraded number knowledge due to their temporal-parietal disease. Another class of quantifiers, logical quantifiers, depends in large part on a simple form of perceptual logic, and these are compromised in patients with bvFTD who have disease in rostral regions of PFC. A final class of quantifiers, known as majority quantifiers, depends on executive resources such as working memory and strategic comparisons, and these too are impaired in bvFTD due in part to disease that interferes with functioning of the frontal-striatal loop. Additionally, we emphasize that a language disorder is unlikely to explain the deficit in these non-aphasic patients as they did not differ from healthy seniors in their comprehension of control stimuli that do not contain a quantifier or a number. We discuss each of these classes of quantifiers below.
Cardinal quantifier comprehension in CBS/PCA
Cardinal quantifiers like “at least 3 X” are well understood in formal semantic terms (Keenan, et al., 1997; van Benthem, 1986), but a mechanistic account from a neurocognitive perspective has been addressed only recently (Clark & Grossman, 2007). According to this approach, a sequence of quantity states must be examined, and this depends on a sense of number knowledge. Quantity is a distinct semantic domain that involves magnitude and numerosity representations independent of the objects or states that are being enumerated (Dehaene, 1997; Halpern, et al., 2004). The truth-value of a cardinal quantifier ultimately is tested when there is a comparison between the quantity specified in the quantifier and the state of the world that is being evaluated.
Previous work in clinically-diagnosed and autopsy-confirmed cases has shown that non-aphasic patients with CBS have difficulty with numbers (Halpern, et al., 2003; Halpern, et al., 2004; Koss, et al., 2010; Pantelyat, et al., 2011). We are aware of only one previous assessment of quantity in an individual patient with PCA (Delazer, Kramer, Zanarian, Donnemiller, & Benke, 2006). This case study described preserved processing for some aspects of number knowledge in PCA. We examined quantity as a control condition in the participants of the present study, and we confirmed that patients with CBS as well as the three patients with PCA have significant difficulty judging a small quantity of familiar objects. The discrepancy between our observation and that of Delazer and colleagues may be due in part to differences in patient assessment or the anatomic distribution of disease in individual patients. It does not appear that this deficit in CBS and PCA can be easily attributed to a visual-perceptual deficit or difficulty understanding the meaning of the objects mentioned in the materials used in this study since participants were as accurate as healthy seniors at evaluating the same pictured materials with sentences that do not contain a quantity. This deficit appreciating precise quantity appears to be relatively specific to CBS and PCA since bvFTD patients were not significantly impaired in their comprehension of precise quantity. Only a small number of PCA patients participated in this study. Additional work with larger numbers of PCA patients is needed to resolve the discrepancy with the single previously reported case and characterize number knowledge in these patients.
In this context, we found that the comprehension of cardinal quantifiers in non-aphasic patients with CBS/PCA tracks their poor comprehension of precise quantity. Thus, these patients were significantly impaired in their comprehension of cardinal quantifiers compared to controls. Moreover, comprehension of cardinal quantifiers in CBS/PCA patients was significantly correlated with their accuracy judging precise numbers. The selective nature of this deficit is emphasized by the observation that these patients did not differ from controls in their comprehension of logical quantifiers, presumably because logical quantifiers do not depend on knowledge of quantity. These observations are consistent with previous work showing a deficit understanding cardinal quantifiers in CBS (McMillan, et al., 2006; Troiani, et al., 2008; Troiani, et al., 2010). The present study extends difficulty with cardinal quantifiers in CBS and PCA to impaired judgments of familiar objects in naturalistic visual scenes.
We also examined the neuroanatomic basis for difficulty understanding cardinal quantifiers in CBS/PCA. Studies of healthy subjects using fMRI (Ansari, et al., 2006; Castelli, et al., 2006; Piazza, et al., 2006; Shuman & Kanwisher, 2004), and correlations of performance with anatomic deficits in patients and with temporary suppression of local cortical functioning in controls with transcortical magnetic stimulation (Ashkenazi, et al., 2008; Cappelletti, et al., 2007; Cappelletti, et al., 2009; Dellatolas, et al., 2001; Gobel, et al., 2006; Martory, et al., 2003), have emphasized the association of quantity and number processing with inferior parietal and posterior temporal regions. Rare fMRI studies of healthy adults also emphasize the role of temporal-parietal regions in the comprehension of cardinal quantifiers (McMillan, et al., 2005; Troiani, et al., 2008). These findings are consistent with quantitative neuroimaging assessments showing atrophy in temporal-parietal regions in patients with CBS and PCA (Grossman, et al., 2004; Hof, et al., 1997; Nestor, et al., 2003; Whitwell, et al., 2007). Likewise, histopathologic assessments of the anatomic distribution of disease burden in both CBS (Josephs, et al., 2006; Murray, et al., 2007; Pantelyat, et al., 2011) and PCA (Alladi, et al., 2007; Hof, et al., 1997; Victoroff, et al., 1994) have shown disease in the parietal lobe and posterior temporal regions.
Quantitative assessment of volumetric MRI in the present study confirmed previous observations of temporal-parietal atrophy in CBS/PCA patients. Moreover, we used regressions in voxel-based morphometric analyses to relate impaired comprehension of cardinal quantifiers to temporal-parietal atrophy. We found that difficulty with cardinal quantifier comprehension was associated with atrophy in posterior temporal and parietal regions in patients in the CBS/PCA group. The results of the regression analysis also extended more ventrally in the temporal lobe than fMRI studies of quantifier comprehension in healthy adults, and this may reflect a difference in study methods. The relationship with visual association cortex in the ventral temporal lobe thus may be due in part to the use of stimuli containing visual-perceptual pictures and printed alphabetic sentences, and processing these stimulus attributes depends in part on these visual association areas. Unlike fMRI, we cannot subtract out these attributes of the stimulus materials with the imaging technique used in this study. Given the potential controversy associated with our observation of a deficit in PCA, it would have been useful as well to examine this subgroup of patients separately. Unfortunately, the number of patients with PCA was too small to perform atrophy and regression analyses in this subgroup alone, and it will be important for future studies to relate quantity knowledge and cardinal quantifier comprehension directly to temporal-parietal regions in a larger group of patients with PCA.
Logical quantifier comprehension in bvFTD
The pattern of difficulty in bvFTD stands in contrast to the impairment seen in patients with CBS/PCA. While non-aphasic bvFTD patients do not have difficulty understanding cardinal quantifiers, bvFTD patients do have limited executive resources (Kramer, et al., 2003; Libon, et al., 2008; Libon, et al., 2007). One kind of executive resource may involve a simple form of perceptual logic that depends in part on dichotomous decisions about objects that are being selectively attended (Burgess, et al., 2007; Gilbert, et al., 2005). This appears to be crucial to the comprehension of logical quantifiers. Quantifiers like “some X” and “not all X” thus depend largely on detecting the presence or absence of a single instance of a target category. This kind of simple perceptual logic has been associated with “Aristotelean” or logical quantifiers such as these in a previous study of patients with bvFTD (Troiani, et al., 2008). This cannot be attributed easily to a language deficit since these patients are not aphasic and had normal performance on the control measure assessing comprehension of the same materials without numbers or quantifiers. Likewise, normal performance on these control materials emphasizes that these patients do not have impaired vigilance or difficulty with other task-related demands.
Consistent with the possibility that comprehension of logical quantifiers depends in part on executive resources in bvFTD, we found that these patients are significantly impaired in their comprehension of logical quantifiers. Moreover, difficulty understanding logical quantifiers correlated with performance on measures of executive functioning in these patients. Difficulty with logical quantifiers also appeared to be fairly specific since bvFTD patients were relatively unimpaired in their comprehension of cardinal quantifiers, and only bvFTD patients were impaired at understanding logical quantifiers. This is consistent with a double dissociation compared to the CBS/PCA group of patients who are relatively impaired in their comprehension of cardinal quantifiers but appear to have preserved understanding of logical quantifiers. This double dissociation must be interpreted cautiously since the dissociation involves performance of patient groups relative to control subjects rather than comparisons of the two patient groups directly to each other. Future work will be needed to confirm the double dissociation in larger groups of subjects.
One previous study examined the neuroanatomic basis for the comprehension of logical quantifiers. This fMRI study of healthy adults showed recruitment of rostral PFC (Troiani, et al., 2008). Several other fMRI studies have associated this brain region with the form of simple perceptual logic that appears to contribute to processing this kind of quantifier (Burgess, et al., 2007; Gilbert, et al., 2005). Quantitative MRI studies have shown significant atrophy in this anatomic distribution in bvFTD (Grossman, et al., 2007; Grossman, et al., 2004; Kipps, et al., 2009), and histopathologic assessments of the anatomic distribution of disease have demonstrated that rostral prefrontal regions are frequently compromised in patients with bvFTD (Forman, et al., 2006; Grossman, et al., 2007). We found significant atrophy in this area in bvFTD. Moreover, we found an association between comprehension of logical quantifiers and atrophy of rostral prefrontal cortex. This was evident when the regression analysis was conducted at a more liberal p<0.1 level, possibly because logical quantifiers are highly familiar and relatively easy to process. Another factor contributing to the weak but predicted association of logical quantifier comprehension with rostral PFC in bvFTD may have been the relatively small number of patients with available imaging. Nevertheless, this association was unique to bvFTD and was not seen in CBS/PCA, just as temporal-parietal disease was not seen in bvFTD. Additional work is needed to confirm our findings in a larger group of bvFTD patients.
Majority quantifier comprehension in bvFTD
We also examined comprehension of majority quantifiers. These too appear to depend on executive resources, although the resources are different from those implicated in logical quantifiers. Here, a quantity must be maintained in an active state in working memory while a second quantity is computed. Then a strategic comparison between the two quantities must be performed to evaluate the truth-value of the quantified statement. Patients with bvFTD were significantly impaired at understanding majority quantifiers. Moreover, they had deficits on executive measures, and comprehension of majority quantifiers in bvFTD correlated with performance on these executive measures. Previous work also found difficulty understanding majority quantifiers in bvFTD (McMillan, et al., 2006; Troiani, et al., 2008), and this work has shown a correlation of impaired majority quantifier comprehension with executive resource limitations as well.
We did not find statistically significant impairment understanding majority quantifiers in CBS/PCA relative to healthy controls in the present study, although this condition was the most difficult for the CBS/PCA group. In previous work, by comparison, we found that CBS patients are significantly impaired in their comprehension of majority quantifiers (McMillan, et al., 2006; Troiani, et al., 2008). We reasoned previously that CBS patients have difficulty with majority quantifiers because of the quantity component of majority quantifiers. There are several possible explanations for the discrepancy between previous work and the current study. One possibility is related to a difference in the materials used in the present study compared to previous studies. The materials of the present study involved naturalistic pictures, and processing these may have required relatively modest executive resources compared to a series of isolated object pictures without a context. Another possibility is that we generally used smaller numbers in the present study, typically in the subitizing range. Previous work showed that these smaller numbers are processed more accurately than single digit numbers within the counting range, even though the smaller numbers are not subitized and are more difficult for CBS patients than controls (Halpern, et al., 2007). A final possibility is that patients with CBS may have executive deficits (Graham, Bak, & Hodges, 2003), related in part to the observation of some frontal disease in these patients (Josephs, et al., 2006; Murray, et al., 2007), although patients with PCA tend to have minimal frontal disease. It is thus possible that we did not find a deficit with majority quantifiers in the present study because we combined CBS and PCA into a single group, thereby lessening the frontal disease burden in this group overall. Additional work is needed in larger groups of patients with more varied stimulus materials to evaluate these discrepant findings in more detail.
fMRI studies of majority quantifiers in healthy adults have shown activation that includes extensive regions of lateral PFC (McMillan, et al., 2005; Troiani, et al., 2008). This corresponds to areas of the brain that are activated in fMRI studies of working memory and difficult strategic comparisons (Badre & Wagner, 2004; Smith & Jonides, 2002). bvFTD patients have considerable atrophy in PFC and the striatum in quantitative MRI studies (Grossman, et al., 2007; Grossman, et al., 2004; Kipps, et al., 2009). Autopsy studies of bvFTD also show disease in PFC and the striatum (Forman, et al., 2006; Grossman, et al., 2007). It is unsurprising from this perspective that bvFTD patients have difficulty understanding majority quantifiers. Nevertheless, we found only modest evidence relating performance on majority quantifiers directly to prefrontal atrophy. This may have been due in part to the fact that this particular group of patients did not have robust atrophy in dorsolateral regions of prefrontal cortex. At a more liberal level of significance, regression analyses did demonstrate an association between majority quantifier comprehension and predicted atrophy in lateral PFC of bvFTD patients. We did find a significant relationship between comprehension of majority quantifiers and basal ganglia disease. This may reflect the interruption of a frontal-striatal cognitive loop important for executive-mediated cognitive difficulties. Additional work is needed to relate majority quantifiers to more precisely to prefrontal-striatal disease in bvFTD.
In sum, we demonstrated disruption of a large-scale neural network important for the representation of quantifier meaning. Unlike the representation of word meaning for objects, quantifier meaning is minimally contingent on representations of sensory-motor features. Instead, quantifier meaning appears to depend largely on a sense of magnitude and quantity. Moreover, the neural basis for quantifier meaning overlaps minimally with the hypothesized neuroanatomic network thought to be important for the representation of word meaning for objects (Barsalou, et al., 2003). Word meaning for objects is centered in part on a network involving modality-specific association cortices (Martin, 2007). Instead, the representation of quantifier meaning appears to be based in large part on magnitude and quantity knowledge that depends on inferior parietal cortex, with additional support as needed from portions of prefrontal cortex. This raises the possibility that there are multiple neural mechanisms for representing long-term knowledge about our world in lexical semantic memory.
RESEARCH HIGHLIGHTS.
This report assesses the cognitive and neural basis for a common element of our vocabulary – quantifiers.
Cognitive components contributing to quantifier comprehension are thought to include knowledge of quantity, a simple form of perceptual logic, and executive resources such as working memory and strategic decision-making.
A large-scale frontal-parietal neural network is thought to support quantifier comprehension.
This study demonstrates selective impairment of the cognitive components implicated in quantifier comprehension in patients with focal neurodegenerative disease that compromises frontal and parietal brain regions.
Acknowledgments
Portions of this work were presented at annual meetings of the American Academy of Neurology, 2010, and Society for Neuroscience, 2009. This work was supported in part by grants from NIH (NS44266, AG17586, AG15116, NS53488, and AG32953).
APPENDIX. STIMULUS SENTENCES FOR QUANTIFIER AND CONTROL CONDITIONS
Cardinal
The blonde lady is holding more than three flowers.
The strong boy is carrying more than three boxes.
There is at least one broom in the closet.
There are more than three crayons in the box.
There are fewer than three birds in the cage.
There are more than three pillows on the bed.
There are fewer than two penguins on the igloo.
There are less than five cans on the counter.
The young man is carrying fewer than five books.
There are at least two ants on the hill.
There are greater than two bananas in the tree.
There are at least four bottles on the table.
There are at least two apples in the tree.
There are greater than two pumpkins in the truck.
There are fewer than two cows in the pasture.
The intelligent woman is holding at least two pencils.
The happy girl is holding at least one cup.
The happy boy is holding more than four balloons.
There are greater than three shoes on the rack.
There are less than four people sitting on the floor.
There are more than four shoes on the rack.
There are less than two crayons in the box.
There are greater than three people sitting on the floor.
There are more than six cans on the counter.
There is less than one banana in the tree.
The intelligent woman is holding more than three pencils.
The young man is carrying fewer than four books.
There are at least three penguins on the igloo.
There are at least five bottles on the table.
There are more than two birds in the cage.
There are more than six pillows on the bed.
There are greater than three ants on the hill.
The happy boy is holding less than three balloons.
The happy girl is holding more than two cups.
There are greater than four brooms in the closet.
The blonde lady is holding less than two flowers
The strong boy is carrying at least four boxes.
There are more than five pumpkins in the truck.
There are at least three cows in the pasture.
There are fewer than three apples in the tree.
Logical
Not all of the ants are on the hill.
There are some orange pumpkins in the red truck.
The intelligent business woman is holding some sharpened pencils.
The happy brunette girl is holding some blue cups.
Not all of the pillows are on the bed.
Not all of the cows are in the pasture.
There are some silver cans on the kitchen counter.
There are some polite people sitting on the floor.
The happy, brunette boy is holding some pink balloons.
The strong, determined boy is carrying some cardboard boxes
The blonde lady is holding some beautiful purple flowers.
The studious, young man is carrying some green books.
Not all of the crayons are in the box.
Not all of the shoes are on the rack.
Not all of the bananas are in the tree.
Not all of the apples are in the tree.
Not all of the bottles are on the table.
Not all of the penguins are on the igloo.
There are some blue birds in the metal cage.
Not all of the brooms are in the closet.
The blonde lady is holding some beautiful purple flowers.
The studious, young man is carrying some green books.
Not all of the pillows are on the bed.
Not all of the ants are on the hill.
The strong, determined boy is carrying some cardboard boxes
The happy brunette girl is holding some blue cups.
Not all of the cows are in the pasture.
There are some orange pumpkins in the red truck.
Not all of the bananas are in the tree.
Not all of the apples are in the tree.
Not all of the shoes are on the rack.
The happy, brunette boy is holding some pink balloons.
There are some silver cans on the kitchen counter.
The intelligent business woman is holding some sharpened pencils.
Not all of the penguins are on the igloo.
There are some polite people sitting on the floor.
Not all of the crayons are in the box.
There are some blue birds in the metal cage.
Not all of the bottles are on the table.
Not all of the brooms are in the closet.
Majority
Fewer than half the penguins are on the igloo.
The boy is holding at least half of the balloons.
Fewer than half the shoes are on the rack.
More than half the bottles are on the table.
Less than half the cows are in the pasture.
Less than half the birds are in the cage.
The lady is holding at least half the flowers.
The girl is holding less than half the cups.
More than half the people are sitting on the floor.
Less than half the pumpkins are in the truck.
The woman is holding more than half the pencils.
Greater than half the pillows are on the bed.
Less than half the brooms are in the closet.
Fewer than half the crayons are in the box.
At least half the cans are on the counter.
Less than half the ants are on the hill.
At least half the bananas are in the tree.
The man is carrying more than half the books.
The boy is carrying less than half the boxes.
Fewer than half the apples are in the tree.
The man is carrying less than half the books.
The lady is holding less than half the flowers.
The girl is holding at least half of the cups
The boy is holding fewer than half the balloons.
More than half the apples are in the tree.
More than half the ants are on the hill.
At least half the shoes are on the rack.
Fewer than half the bottles are on the table.
More than half the birds are in the cage.
Less than half the bananas are in the tree.
The boy is carrying at least half the boxes.
Greater than half the penguins are on the igloo.
Greater than half the cows are in the pasture.
Less than half the people are sitting on the floor.
Greater than half the crayons are in the box.
At least half the pumpkins are in the truck.
At least half of the brooms are in the closet.
Half of the soft pillows are on the big bed.
Less than half the cans are on the counter.
The woman is holding less than half the pencils.
Precise
There are three orange pumpkins in the red truck.
There are four colored crayons in the open box.
There are four silver cans on the kitchen counter.
There are four blue shoes on the metal rack.
The studious, young man is carrying four green books.
There are five glass bottles on the round table.
There are three red apples in the tall tree.
There is one fat penguin on the cold igloo.
The blonde lady is holding four beautiful purple flowers.
The happy, brunette girl is holding two blue cups.
There are three nice people sitting on the floor.
The happy, brunette boy is holding five pink balloons.
There are three yellow bananas in the big tree.
There are two blue birds in the metal cage.
There are two tall brooms in the open closet.
There are four soft pillows on the big bed.
The strong, determined boy is carrying four cardboard boxes.
There are three red ants on the round hill.
The intelligent, business woman is holding three sharpened pencils.
There is one spotted cow in the open pasture.
The happy, brunette girl is holding two blue cups.
There are three fat penguins on the cold igloo.
There are two colored crayons in the open box.
There are two blue birds in the metal cage.
The intelligent business woman is holding one sharpened pencil.
There are six soft pillows on the big bed.
The studious, young man is carrying four green books.
There is one red ant on the round hill.
There are three lazy cows in the open pasture.
The blonde lady is holding two beautiful purple flowers.
There are two blue shoes on the metal rack.
There are five orange pumpkins in the red truck.
The happy, brunette boy is holding three pink balloons.
There are five glass bottles on the round table.
There are four tall brooms in the open closet.
There are five red apples in the tall tree.
There are three polite people sitting on the floor.
There is one yellow banana in the big tree.
There are six silver cans on the kitchen counter.
The strong, determined boy is carrying two cardboard boxes.
Control
There are glass bottles on the round, wooden table.
The intelligent business woman is holding thin, sharpened pencils.
There are happy, spotted cows in the open pasture.
There are silver soup cans on the kitchen counter.
There are bright, colored crayons in the open box.
There are small blue birds in the metal cage
There are soft, purple pillows on the big bed.
The happy, brunette boy is holding pink, floating balloons.
The blonde young lady is holding beautiful purple flowers
The happy, brunette girl is holding small, blue cups.
There are fat penguins on the cold, icy igloo.
There are tied, blue shoes on the metal rack.
There are polite people sitting on the hard floor.
There are small, red ants on the round hill.
The strong, determined boy is carrying large, cardboard boxes.
There are ripe, yellow bananas in the big tree.
There are tall brooms in the wide, open closet.
There are fat orange pumpkins in the red truck.
There are tasty, red apples in the tall tree.
The studious, young man is carrying big, green books
There are sweet, juicy acorns in the tall tree.
There are green, jumpy frogs on the round hill.
The happy, brunette boy is holding pretty, blooming flowers.
There are old, worn hats on the metal rack.
The blonde young lady is holding clanky, silver pots.
There are elegant, brown horses in the open pasture.
The strong, determined boy is carrying flimsy, plastic bags.
There are cotton shirts in the wide, open closet.
There are big, hefty boxes on the kitchen counter.
There are plump, juicy strawberries in the big tree.
The studious, young man is carrying colorful, bright posters.
The intelligent business woman is holding bulky, black staplers.
There are long, plastic rulers in the open box.
The happy, brunette girl is holding round, porcelain plates.
There are fun, plastic toys on the big bed.
There are hungry seals on the cold, icy igloo.
There are fat, juicy watermelons in the red truck.
There are brown dogs sitting on the hard floor.
There are plastic cups on the round, wooden table.
There are big, green lizards in the metal cage.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Alladi S, Xuereb J, Bak T, Nestor P, Knibb J, Patterson K, et al. Focal cortical presentations of Alzheimer’s disease. Brain. 2007;130(10):2636–2645. doi: 10.1093/brain/awm213. [DOI] [PubMed] [Google Scholar]
- Ansari D, Dhital B, Siong SC. Parametric effects of numerical distance on the intraparietal sulcus during passive viewing of rapid numerosity changes. Brain Research. 2006;1067(1):181–188. doi: 10.1016/j.brainres.2005.10.083. [DOI] [PubMed] [Google Scholar]
- Ashkenazi S, Henik A, Ifergane G, Shelef I. Basic numerical processing in left intraparietal sulcus (IPS) acalculia. Cortex. 2008;44(4):439–448. doi: 10.1016/j.cortex.2007.08.008. [DOI] [PubMed] [Google Scholar]
- Avants B, Gee JC. Geodesic estimation for large deformation anatomical shape averaging and interpolation. Neuroimage. 2004;23(Supplement 1):S139–S150. doi: 10.1016/j.neuroimage.2004.07.010. [DOI] [PubMed] [Google Scholar]
- Avants B, Yushkevich PA, Pluta J, Minkoff D, Korczykowski M, Detre J, et al. The optimal template effect in hippocampus studies of diseased populations. Neuroimage. 2010 doi: 10.1016/j.neuroimage.2009.09.062. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Avants BB, Epstein CL, Grossman M, Gee J. Symmetric diffeomorphic image registration with cross-correlation: Evaluating automated labeling of elderly and neurodegenerative brain. Med Image Anal. 2008;12:26–41. doi: 10.1016/j.media.2007.06.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baddeley AD, Della Sala S, Papagno C, Spinnler H. Dual-task performance in dysexecutive and nondysexecutive patients with a frontal lesion. Neuropsychology. 1997;11:187–194. doi: 10.1037//0894-4105.11.2.187. [DOI] [PubMed] [Google Scholar]
- Badre D, Wagner AD. Selection, integration, and conflict monitoring: Assessing the nature and generality of prefrontal cognitive control mechanisms. Neuron. 2004;41:473–487. doi: 10.1016/s0896-6273(03)00851-1. [DOI] [PubMed] [Google Scholar]
- Barsalou LW, Simmons WK, Barbey AK, Wilson CD. Grounding conceptual knowledge in modality-specific systems. Trends in Cognitive Sciences. 2003;7(2):84–91. doi: 10.1016/s1364-6613(02)00029-3. [DOI] [PubMed] [Google Scholar]
- Boeve BF, Maraganore DM, Parisi JE, Ahlskog JE, Graff-Radford NR, Caselli RJ, et al. Pathologic heterogeneity in clinically diagnosed corticobasal degeneration. Neurology. 1999;53:795–800. doi: 10.1212/wnl.53.4.795. [DOI] [PubMed] [Google Scholar]
- Brown RG, Soliveri P, Jahanshahi M. Executive processes in Parkinson’s disease -- random number generation and response suppression. Neuropsychologia. 2000;36:1355–1362. doi: 10.1016/s0028-3932(98)00015-3. [DOI] [PubMed] [Google Scholar]
- Burgess PW, Dumontheil I, Gilbert SJ. The gateway hypothesis of rostral prefrontal cortex (area 10) function. Trends in Cognitive Sciences. 2007;11(7):290–298. doi: 10.1016/j.tics.2007.05.004. [DOI] [PubMed] [Google Scholar]
- Cappelletti M, Barth H, Fregni F, Spelke E, Pascual-Leone A. rTMS over the intraparietal sulcus disrupts numerosity processing. Experimental Brain Research. 2007;179(4):631–642. doi: 10.1007/s00221-006-0820-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cappelletti M, Muggleton N, Walsh V. Quantity without numbers and numbers without quantity in the parietal cortex. Neuroimage. 2009;46(2):522–529. doi: 10.1016/j.neuroimage.2009.02.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Castelli F, Glaser DE, Butterworth B. Discrete and analogue quantity processing in the parietal lobe: A functional MRI study. Proceedings of the National Academy of Sciences of the United States of America. 2006;103(12):4693–4698. doi: 10.1073/pnas.0600444103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clark R, Grossman M. Number sense and quantifier comprehension. Topos. 2007;26:51–62. [Google Scholar]
- Das SR, Avants BB, Grossman M, Gee J. Registration based cortical thickness measurement. Neuroimage. 2009;45:867–879. doi: 10.1016/j.neuroimage.2008.12.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dehaene S. The Number Sense. New York, NY: Oxford University Press; 1997. [Google Scholar]
- Delazer M, Kramer E, Zanarian L, Donnemiller E, Benke T. Number processing in posterior cortical atrophy - A neuropsychological case study. Neuropsychologia. 2006;44:36–51. doi: 10.1016/j.neuropsychologia.2005.04.013. [DOI] [PubMed] [Google Scholar]
- Dellatolas G, Deloche G, Basso A, Claros-Salinas D. Assessment of calculation and number processing using the EC301 battery: Cross-cultural normative data and application to left- and right-brain damaged patients. Journal of the International Neuropsychological Society. 2001;7:840–859. [PubMed] [Google Scholar]
- Forman MS, Farmer J, Johnson JK, Clark CM, Arnold SE, Coslett HB, et al. Frontotemporal dementia: Clinicopathological correlations. Annals of Neurology. 2006;59:952–962. doi: 10.1002/ana.20873. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gilbert SJ, Frith CD, Burgess PW. Involvement of rostral prefrontal cortex in selection between stimulus-oriented and stimulus-independent thought. European Journal of Neuroscience. 2005;21(5):1423–1431. doi: 10.1111/j.1460-9568.2005.03981.x. [DOI] [PubMed] [Google Scholar]
- Gobel SM, Calabria M, Farne A, Rossetti Y. Parietal rTMS distorts the mental number line: Simulating ‘spatial’ neglect in healthy subjects. Neuropsychologia. 2006;44:860–868. doi: 10.1016/j.neuropsychologia.2005.09.007. [DOI] [PubMed] [Google Scholar]
- Graham NL, Bak T, Hodges JR. Corticobasal degeneration as a cognitive disorder. Movement Disorders. 2003;18:1224–1232. doi: 10.1002/mds.10536. [DOI] [PubMed] [Google Scholar]
- Grimes DA, Lang AE, Bergeron CB. Dementia as the most common presentation of corticobasal ganglionic degeneration. Neurology. 1999;53:1969–1974. doi: 10.1212/wnl.53.9.1969. [DOI] [PubMed] [Google Scholar]
- Grossman M, Libon DJ, Forman MS, Massimo L, Wood E, Moore P, et al. Distinct antemortem profiles in patients With pathologically defined frontotemporal dementia. Arch Neurol. 2007;64(11):1601–1609. doi: 10.1001/archneur.64.11.1601. [DOI] [PubMed] [Google Scholar]
- Grossman M, McMillan C, Moore P, Ding L, Glosser G, Work M, et al. What’s in a name: Voxel-based morphometric analyses of MRI and naming difficulty in Alzheimer’s disease, frontotemporal dementia, and corticobasal degeneration. Brain. 2004;127:628–649. doi: 10.1093/brain/awh075. [DOI] [PubMed] [Google Scholar]
- Halpern C, Clark R, McMillan C, Dennis K, Moore P, Grossman M. Calculation difficulty in neurodegenerative diseases. Journal of the Neurological Sciences. 2003;208:31–38. doi: 10.1016/s0022-510x(02)00416-1. [DOI] [PubMed] [Google Scholar]
- Halpern C, Clark R, Moore P, Cross K, Grossman M. Too much to count on: Impaired very small numbers in corticobasal degeneration. Brain and Cognition. 2007;64(2):144–149. doi: 10.1016/j.bandc.2007.01.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Halpern C, Glosser G, Clark R, Gee JC, Moore P, Dennis K, et al. Dissociation of numbers and objects in corticobasal degeneration and semantic dementia. Neurology. 2004;62:1163–1169. doi: 10.1212/01.wnl.0000118209.95423.96. [DOI] [PubMed] [Google Scholar]
- Hof PR, Vogt BA, Bouras C, Morrison JH. Atypical form of Alzheimer’s disease with prominent posterior cortical atrophy: A review of lesion distribution and circuit disconnection in cortical visual pathways. Vision Research. 1997;37:3609–3625. doi: 10.1016/S0042-6989(96)00240-4. [DOI] [PubMed] [Google Scholar]
- Josephs KA, Petersen RC, Knopman DS, Boeve BF, Whitwell JL, Duffy JR, et al. Clinicopathologic analysis of frontotemporal and corticobasal degenerations and PSP. Neurology. 2006;66:41–48. doi: 10.1212/01.wnl.0000191307.69661.c3. [DOI] [PubMed] [Google Scholar]
- Keenan EL, Westersthål D, van Benthem J, ter Meulen A. Handbook of Logic and Language. Cambridge, MA: MIT Press; 1997. Generalized quantifiers in linguistics and logic; pp. 837–893. [Google Scholar]
- Kim J, Avants B, Patel S, Whyte J, BHC, Pluta J, et al. Structural consequences of diffuse traumatic brain injury: A large deformation tensor-based morphometry study. 2008;39:1014–1026. doi: 10.1016/j.neuroimage.2007.10.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kipps CM, Hodges JR, Fryer TD, Nestor PJ. Combined magnetic resonance imaging and positron emission tomography brain imaging in behavioural variant frontotemporal degeneration: refining the clinical phenotype. Brain. 2009;132(9):2566–2578. doi: 10.1093/brain/awp077. [DOI] [PubMed] [Google Scholar]
- Klein A, Andersson J, Ardekani BA, Ashburner J, Avants B, Chiang M, et al. Evaluation of 14 nonlinear deformation algorithms applied to human brain MRI registration. Neuroimage. 2009;46:786–802. doi: 10.1016/j.neuroimage.2008.12.037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koss S, Clark R, Vesely L, Weinstein J, Anderson C, Richmond L, et al. Numerosity impairment in corticobasal syndrome. Neuropsychology. 2010;24:476–492. doi: 10.1037/a0018755. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kramer JH, Jurik J, Sha SJ. Distinctive neuropsychological patterns of frontotemporal dementia, semantic dementia, and Alzheimer’s Disease. Cognitive and Behavioral Neurology. 2003;16:211–218. doi: 10.1097/00146965-200312000-00002. [DOI] [PubMed] [Google Scholar]
- Libon DJ, Xie S, Wang X, Massimo L, Moore P, Vesely L, et al. Neuropsychological decline in frontotemporal lobar degeneration: A longitudinal analysis. Neuropsychology. 2008;23:337–346. doi: 10.1037/a0014995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Libon DJ, Xie SX, Moore P, Farmer J, Antani S, McCawley G, et al. Patterns of neuropsychological impairment in frontotemporal dementia. Neurology. 2007;68:369–375. doi: 10.1212/01.wnl.0000252820.81313.9b. [DOI] [PubMed] [Google Scholar]
- Martin A. The representation of object concepts in the brain. Annu Rev Psychol. 2007;58:25–45. doi: 10.1146/annurev.psych.57.102904.190143. [DOI] [PubMed] [Google Scholar]
- Martory MD, Mayer E, Pegna AJ, Aninoni JM, Landis T, Khateb A. Pure global acalculia following a left subangular lesion. Neurocase. 2003;9:319–328. doi: 10.1076/neur.9.4.319.15549. [DOI] [PubMed] [Google Scholar]
- McKhann G, Trojanowski JQ, Grossman M, Miller BL, Dickson D, Albert M. Clinical and pathological diagnosis of frontotemporal dementia: Report of a work group on frontotemporal dementia and Pick’s disease. Archives of Neurology. 2001;58:1803–1809. doi: 10.1001/archneur.58.11.1803. [DOI] [PubMed] [Google Scholar]
- McMillan CT, Clark R, Moore P, DeVita C, Grossman M. Neural basis for generalized quantifier comprehension. Neuropsychologia. 2005;43:1729–1737. doi: 10.1016/j.neuropsychologia.2005.02.012. [DOI] [PubMed] [Google Scholar]
- McMillan CT, Clark R, Moore P, Grossman M. Quantifier comprehension in corticobasal degeneration. Brain and Cognition. 2006;62:250–260. doi: 10.1016/j.bandc.2006.06.005. [DOI] [PubMed] [Google Scholar]
- Murray R, Neumann M, Forman MS, Farmer J, Massimo L, Rice A, et al. Cognitive and motor assessment in autopsy-proven corticobasal degeneration. Neurology. 2007;68:1274–1283. doi: 10.1212/01.wnl.0000259519.78480.c3. [DOI] [PubMed] [Google Scholar]
- Nestor PJ, Caine D, Fryer TD, Clarke J, Hodges JR. The topography of metabolic deficits in posterior cortical atrophy (the visual variant of Alzheimer’s disease) with FDG-PET. Journal of Neurology, Neurosurgery, and Psychiatry. 2003;74:1521–1529. doi: 10.1136/jnnp.74.11.1521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pantelyat A, Gross RG, Moore P, Dreyfuss M, Schuck T, Irwin DJ, Trojanowski JQ, Grossman M. Acalculia in autopsy proven corticobasal degeneration. Neurology. 2011;76:S61–63. doi: 10.1212/WNL.0b013e31820c34ca. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Piazza M, Mechelli A, Price CJ, Butterworth B. Exact and approximate judgements of visual and auditory numerosity: An fMRI study. Brain Research. 2006;1106(1):177–188. doi: 10.1016/j.brainres.2006.05.104. [DOI] [PubMed] [Google Scholar]
- Rascovsky K, Hodges JR, Kipps CM, Johnson JK, Seeley WW, Mendez MF, et al. Diagnostic criteria for the behavioral variant of frontotemporal dementia (bvFTD): current limitations and future directions. Alzheimer’s Disease and Associated Disorders. 2007;21:514–518. doi: 10.1097/WAD.0b013e31815c3445. [DOI] [PubMed] [Google Scholar]
- Riley DE, Lang AE, Litvan I, Goetz CG, Lang AE. Corticobasal Degeneration and Related Disorders. Philadelphia: Lippincott Williams and Wilkins; 2000. Clinical diagnostic criteria; pp. 29–34. [Google Scholar]
- Shuman M, Kanwisher N. Numerical Magnitude in the Human Parietal Lobe: Tests of Representational Generality and Domain Specificity. Neuron. 2004;44(3):557–569. doi: 10.1016/j.neuron.2004.10.008. [DOI] [PubMed] [Google Scholar]
- Smith EE, Jonides J. Executive control and thought. Fundamental Neuroscience 2002 [Google Scholar]
- Troiani V, Clark R, Grossman M. Impaired verbal comprehension of quantifiers in corticobasal syndrome. Neuropsychology. 2011;25:159–165. doi: 10.1037/a0021448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Troiani V, Peelle JE, Clark R, Grossman M. Is it logical to count on quantifiers? Dissociable neural networks underlying numerical and logical quantifiers. Neuropsychologia. 2008;47:104–111. doi: 10.1016/j.neuropsychologia.2008.08.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Benthem J. Essays in Logical Semantics. Dordrecht, the Netherlands: D. Reidel Publishing Company; 1986. [Google Scholar]
- Victoroff J, Ross W, Benson DF, Verity MA. Posterior cortical atrophy: Neuropathological correlations. Archives of Neurology. 1994;51:269–274. doi: 10.1001/archneur.1994.00540150063018. [DOI] [PubMed] [Google Scholar]
- Whitwell JL, Jack CR, Jr, Parisi JE, Knopman DS, Boeve BF, Petersen RC, et al. Rates of cerebral atrophy differ in different degenerative pathologies. Brain. 2007;130(4):1148–1158. doi: 10.1093/brain/awm021. [DOI] [PMC free article] [PubMed] [Google Scholar]