

Abstract
M cells are specialized epithelial cells mediating immune surveillance of the mucosal lumen by transepithelial delivery of antigens to underlying dendritic cells (DC). At least three M cell phenotypes are known in airways and intestine, but their developmental relationships are unclear. We used reporter transgenic mouse strains to follow the constitutive development of M cell subsets, and their acute induction by cholera toxin (CT). M cells overlying intestinal Peyer’s patches (PP), Isolated Lymphoid Follicles (ILF), and Nasal Associated Lymphoid Tissue (NALT) are induced by distinct settings, yet show convergent phenotypes, such as expression of a PGRP-S (PeptidoGlycan Recognition Protein – S) transgene reporter. By contrast, though PP, ILF, and Villous M cells are all derived from intestinal crypt stem cells, their phenotypes were clearly distinct; for example, PP M cells frequently appeared to form M cell-DC functional units while Villous M cells did not consistently engage underlying DC. B lymphocytes are critical to M cell function by forming a basolateral pocket and possible signaling through CD137; however, initial commitment to all M cell lineages is B lymphocyte and CD137 independent. CT causes induction of new M cells in the airway and intestine without cell division, suggesting transdifferentiation from mature epithelial cells. In contrast to intestinal PP M cells, CT-induced NALT M cells appear to be generated from ciliated Foxj1+PGRP-S+ cells, indicative of a possible pre-committed progenitor. In sum, constitutive and inducible differentiation of M cells is toward strictly defined context-dependent phenotypes suggesting specialized roles in surveillance of mucosal antigens.
Keywords: mucosal immunity, immune surveillance, epithelium
Introduction
M cells are specialized epithelial cells of the airway and intestinal mucosa that play a primary role in mucosal immune surveillance (1–3). Captured microparticles are transported (transcytosed) across the epithelium for uptake by underlying dendritic cells and stimulation of mucosal immunity. Most M cells are found in follicle-associated epithelium overlying organized mucosal lymphoid aggregates such as intestinal Peyer’s patches (PP) (4) and Nasal Associated Lymphoid Tissues (NALT) (5). M cells are also present over inducible lymphoid aggregates such as Isolated Lymphoid Follicles (ILF) (6) in the intestine and Bronchus-Associated Lymphoid Tissue (BALT) (7) in the lung. In addition, an inducible M cell population has been described at the tips of intestinal villi, called Villous M cells (8).
The regulation of M cell development and function is not well understood despite the assumed central role of M cells in mucosal immunity. In the constitutive mucosal lymphoid tissues, it would be reasonable to assume that lymphoid tissue inducer cells (LTi) responsible for the formation of Peyer’s patches (9) and associated stromal cells are also involved in M cell development. Accordingly, LTi-type TNF and lymphotoxin ligands have been reported to induce expression of genes in intestinal epithelium that were associated with follicle epithelium and M cells (10). Since the follicle epithelium turns over rapidly with a half-life of a few days (11), the regulation of M cell proportion among follicle epithelial cells requires dynamic regulation. Thus, the origin and induction of M cells has been subject to controversy. Various studies have supported the competing views that M cell induction is due to (a) a subset of crypt stem cells dedicated solely to M cell production (12–14) or (b) transdifferentiation from existing mature enterocytes (15; 16). The situation becomes more complex in settings where immune stimulation by bacteria (e.g., Strep. pneumoniae) (17) or their components (e.g., flagellin or cholera toxin) (18) can induce rapid production of new M cells; here, the kinetics of induction challenge the idea that crypt stem cells directly account for all new M cell production.
Studies have suggested that M cell development in the intestine is dependent on B lymphocytes; indeed, a characteristic feature of PP follicle M cells is the presence of a basolateral pocket usually containing at least one B lymphocyte (19; 20). Co-cultures between Caco-2 intestinal epithelium and B lymphocytes have been shown to induce an M cell phenotype (15; 21), and mice lacking B lymphocytes appear to lack M cell transcytosis function (20). However, in CD137-deficient mice that fail to develop M cell basolateral pockets and transcytosis function, the initial lineage commitment of progenitors to the M cell lineage appeared intact (22). Thus, it is possible that M cell development is a two step process with the first lineage commitment step being B cell independent, followed by a step that is B cell-dependent.
The induction of the M cell phenotype is no less complex in the case of mouse NALT M cells. The apical membrane of intestinal M cells appears membranous (hence the name “M” cell) compared to neighboring cells with an actin-based brush border (4). In contrast, airway M cells, also with a membranous apical surface, are surrounded by cells with long tubulin-based cilia (5). Instead of a rapid turnover and crypt stem cell origin, airway epithelial cells are generated from a dispersed population of basal cells, and have half-lives on the order of weeks to months (23–26). Despite their distinct origins and turnover, airway and intestinal M cells show strikingly similar phenotype and functions, suggesting that convergent or overlapping genetic programs are induced.
Materials and Methods
Animals
B cell-deficient mutant mice on the BALB/c background (C.129S2-Igh-6tm1Cgn/J; referred to as “Igh-6” in this report), CX3CR1-EGFP knock-in and Foxj1-EGFP transgenic, and BALB/c mice were obtained from The Jackson Laboratory. CD137-deficient mice on the BALB/c background were provided by Dr. B.S. Kwon. All mice were bred in the UC Riverside vivarium under SPF conditions and were handled in accordance with institutional IACUC and NIH guidelines.
Transgene Construction
To identify the PGRP-S promoter, 5kb of sequence upstream of the PGRP-S gene and PGRP-S intron sequence was obtained from the UCSC genome Bioinformatics database (http://genome.ucsc.edu/), and screened for putative promoter elementsusing the TRANSFAC Transcription Factor Binding Sites Database (http://gene-regulation.com/cgi-bin/pub/databases/transfac/search.cgi), with results in supplemental data. 4kb of candidate promoter sequence was then cloned by PCR from mouse genomic DNA using these primers: 5′-TTCCCAAG-CTTGCTGATAGCCTTTCTAAAACTG-3′ and 5′-GTTCTCAGTGCTATCCATGCTGGACGTGC-CGCACAGG-3′. Based on conventional splice site sequences as well as the size of the intron, the second intron from PGRP-S was also cloned using the primers 5′-GTAAGTACATCCTGGACTG-GGGATGA-3′ and 5′-CTGCAAGGGGAGGGACAAGATACAGA-3′, and inserted into the dsRed express2 coding sequence by overlap PCR. The insertion site of dsRed express2 was decided according to the conservative splice site AG at 5′ exon and G at 3′ exon. The DsRed express2 fragment was obtained from the pCMV-dsRed-Express2 vector (Clontech) and was inserted downstream of the PGRP promoter fragment by overlap PCR. The BGH polyA site fragment, obtained from pCDNA 3.1(+) (Invitrogen) was added to the end of the dsRed coding sequence. Expression of dsRed with the PGRP-S intron (in pCDNA) was checked by transfection into Caco-2BBe cells before cloning downstream of the PGRP-S promoter. To generate transgenic mice the 5.5 kb insert in pGEM was excised from the plasmid backbone by HindIII/KpnI digestion, gel purified and injected into fertilized oocytes of CB6 F2 mice. Founders were identified by PCR analysis of DNA isolated from tail clips using primers specific for DsRed express2 (5′-CGACATCCCCGACTACAAGAAGC-3′ and 5′-CTTCAGCTTCAGCGC-CTTGTGGAT-3′). Founders were bred to BALB/c wild type mice.
Cholera toxin administration
Cholera toxin (Calbiochem) was reconstituted in PBS to a final concentration of 1μg/μl. Mice were starved for 12 hours before cholera toxin administration. The dose of Cholera toxin (15 μg) was based on the studies of Terahara et al. (18) and was delivered either by gavage (in 200μl PBS) or intranasally (in 20μl PBS, 10μl each side). Intranasal volume (10μl per side) was chosen to insure accurate dosing. After 48 hours, mice were humanely killed and dissected for analysis.
Nucleoside analogue labeling and staining
Mice were injected i.p. with 200μg of the nucleoside analogue 5-ethynyl-2′-deoxyuridine (EdU, Invitrogen) in 200 μl PBS at the same time cholera toxin was administered. Small intestine, Peyer’s patch, and NALT were harvested at 48h after injection. Dissected tissues were fixed in 4% formaldehyde/PBS for 15 min, followed by washing with 3% BSA/PBS twice. Tissue was then permeabilized with 0.5% Triton X-100/PBS for 20 m and blocked with 6% BSA/PBS for another 30 m before staining with Alexa Fluor 488 azide Click it reaction cocktail (Invitrogen) for 30 m at room temperature. Tissue was washed with 3% BSA/PBS twice again, and used for immunohistochemistry staining as described below.
Immunohistochemistry and Confocal microscopy
For cryostat sections, dissected NALT or PP tissue was fixed in 4% paraformaldehyde/PBS, flash-frozen for cryostat sections and stained in sections. For whole tissue mounts, dissected tissues were fixed in 4% paraformaldehyde/PBS and then stained in whole tissue fragments. For both cryostat sections and whole mounts, tissues were then permeabilized in PBS, 0.1% Tween and blocked in 0.1% Tween in Casein solution. After that, tissues were stained with antibodies to GP2 (Medical and Biological Laboratories) and Rhodamine, Fluorescein or biotinylated Ulex Europaeus Agglutinin 1 Lectin (UEA-1) (Vector), followed by secondary reagents Alexa Fluor 488 conjugated anti-rat antibody, or streptavidin conjugated Alexa Fluor 647. Post-fixation was done with 4% paraformaldehyde/PBS. Tissue was then mounted with Prolong Gold antifade reagent (Invitrogen) and DAPI as a nuclear counterstain. Images were obtained using a BD CARVII Confocal Imager (BD Biosystems) on a Zeiss Axio Observer inverted microscope. Hardware control (microscope, confocal and digital camera (Diagnostic Instrument Xplorer – XS)) used BD IPLab Imaging Software. Image Z resolution was further optimized with Volocity software (PerkinElmer). M cell numbers were counted for every image and then normalized by surface area, measured by Volocity software.
Scanning Electron Microscopy (SEM)
For Scanning EM, tissues were dissected and fixed in 2.5% glutaraldehyde (Electron Microscopy Sciences) for 3h and 1% OSO4 (Thermo) in PBS for anther 3h. Tissue was then prepared by critical point drying and gold sputter coating. Samples were examined with an FEI XL30 field emission scanning electron microscopy (SEM) at 5 kV.
Statistics
Comparison of M cell numbers between CT treated and non-treated mice were performed using 2-tailed, non-parametric Mann-Whitney test. The nonparametric test was chosen because counts were presented as numbers per defined area rather than absolute counts, though t-tests gave similar results. Comparison of M cell numbers among non-treated, CT gavaged, and CT intranasally treated mice were performed using one way ANOVA (non parametric) Kruskal-Wallis test followed by Dunn’s multiple comparison post test. All statistical analyses were performed using Prism software (GraphPad).
Results
Expression of PGRP-S-dsRed transgene in neutrophils, and M cells of Peyer’s patch follicle epithelium and Nasal Associated Lymphoid Tissue
One challenge to the study of M cell development and function has been the lack of genetic markers of M cell differentiation. In the mouse, identification has depended on the lectin UEA-1 specific for fucose displayed largely on the apical surface of M cells. The association with a specific fucosyltransferase gene has been described (18), though it is not clear that this gene’s expression is unique to M cells. We recently identified the peptidoglycan recognition protein PGRP-S as expressed specifically by Peyer’s patch follicle associated M cells (27), in addition to its known expression by neutrophils (28–30). After an analysis of upstream genomic sequences, we identified a candidate promoter fragment, which we used to drive expression of a reporter red fluorescent protein transgene, called PGRP-S-dsRed (Supplemental Data).
PGRP-S-dsRed transgenic mice expressed red fluorescence in all neutrophils as expected, but also in a small population of F4/80+ CD11blo cells presumably in the macrophage/myeloid lineage (Supplemental Data). Most importantly, red fluorescence was strong in UEA-1-staining epithelial cells in Peyer’s patch follicle epithelium (Figure 1A,B). Although neutrophils can be found within normal mucosal lymphoid tissues, they are not usually found within follicle epithelium; since the morphology of the bone marrow derived cells can be clearly distinguished from M cells, dsRed expression appears to be a promising marker for follicle epithelium M cells.
Figure 1.

Expression of PGRP-S-dsRed transgene in M cells of Peyer’s patch follicle epithelium. (A). Peyer’s patch from PGRP-S-dsRed mice showing that within the follicle epithelium, nearly all the UEA-1 (green) positive M cells express dsRed fluorescence in the cytoplasm.
(B). Peyer’s patch section showing UEA-1 positive (green) M cells expressing dsRed in the cytoplasm (red, arrow). Relative to the neighboring crypt (upper left), apical UEA-1 staining appears slightly earlier than cytoplasmic dsRed fluorescence (asterisk).
(C). Statistical analysis of UEA-1+ PGRP-S-dsRed+ cells in regions of follicle epithelium. Images from whole mount preparations of Peyer’s patches were analyzed, defining the inner two-thirds by drawing radii from the center of each follicle and calculating the two-third point to draw a new oval. Each point represents a single follicle.
(D) Peyer’s patch section showing cytoplasmic dsRed correlating well with apical staining for gp2 (green, arrow), though with nearby crypt at the far left, weak dsRed fluorescence (asterisk) usually slightly preceded the appearance of gp2. Toward the right, stronger dsRed dominates the double-positive fluorescence.
Among cells in the Peyer’s patch follicle epithelium, expression of dsRed showed a specific pattern of coincidence with UEA-1+ epithelial cells suggesting a consistent association with late commitment to the M cell lineage. UEA-1 staining is predominantly at the apical membrane of M cells while the dsRed is cytoplasmic, so in whole mounts of Peyer’s patches, the UEA-1 and dsRed fluorescence within the same cell are sufficiently separated so that a coincident fluorescence signal (e.g., green+red = yellow) depends on z-stack movies to illustrate the correlation between apical UEA-1 and cytoplasmic dsRed (Supplemental Data). At the margins of the follicle epithelium, newly generated M cells (identified by UEA-1 binding) migrate from the adjacent intestinal crypts, so early M cell lineage commitment and maturation likely occurs in this zone. By quantifying the coincidence of apical UEA-1 staining with cytoplasmic dsRed expression, we found that in the outer third of the follicle dome, fewer than 50% of the UEA-1+ cells also expressed dsRed. By contrast, UEA-1+ cells within the central two-thirds of the dome of the follicle epithelium showed greater than 99% coincidence with cytoplasmic dsRed expression (Figure 1C). Thus, once PP M cell commitment has already occurred at the margin of the follicle dome, dsRed expression highly correlated with M cell differentiation thereafter. UEA-1 staining consistently preceded expression of dsRed; in turn, dsRed expression coincided with or slightly preceded the expression of another M cell-associated gene, gp2 (31; 32) (Figure 1D). Together, these results suggest that lineage commitment (defined by UEA-1 lectin binding) and the two genetic markers of M cell development (PGRP-S/dsRed and gp2) are sequentially coordinated at the margin of the follicle epithelium.
Peyer’s patch M cell lineage commitment and PGRP-S reporter expression is B lymphocyte- and CD137-independent
While late expression of PGRP-S and the dsRed transgene suggests association with commitment to the M cell lineage, it is not necessarily associated with M cell function despite the implicit utility of the peptidoglycan binding of PGRP-S for M cell capture of bacteria. Indeed, we recently reported that in CD137-deficient mice, M cell lineage commitment (i.e., appearance of UEA-1+ cells) appeared to be intact, but M cell transcytosis function in both NALT and PP was absent. Associated with this defect was the failure to develop the characteristic B lymphocyte basolateral pockets, suggesting a requirement for a CD137-CD137L signal between M cells and B lymphocytes for pocket formation and M cell functional development. This was also consistent with the observation that in B cell-deficient mice, M cell function was absent (19; 20).
Therefore, to test whether PGRP-S and dsRed expression are specific to lineage commitment and not M cell function, we backcrossed the PGRP-S-dsRed transgene to CD137-deficient (Figure 2A) and B lymphocyte deficient Igh-6 (Figure 2B) mice. In both backcrosses we confirmed previous studies (19; 20) that UEA-1+ cells were still present in PP follicle epithelium. Due to the lack of B lymphocyte formation of basolateral pockets in both backcrosses, the UEA-1+ cells were unusually narrow; however, more importantly, all of them were positive for cytoplasmic dsRed expression. Thus, the M cell specific PGRP-S gene was still induced in UEA-1+ cells despite the lack of basolateral pocket formation and M cell transcytosis function in both knockout models. This result suggests that the M cell genetic program that is initiated prior to basolateral pocket formation is fixed during initial lineage commitment whether transcytosis function develops or not. In addition, it supports the interesting possibility that acquisition of transcytosis function is dependent more on cytoskeletal changes associated with the formation of the basolateral pocket rather than any M cell specific genetic program.
Figure 2.

PGRP-S reporter expression in CD137KO and B lymphocyte deficient Igh-6 mice. CD137KO/PGRP-S-DsRed (A) or Igh-6/PGRP-S-DsRed (B) Peyer’s patch sections show UEA-1+ (Green) M cells expressing dsRed fluorescence (Red) in the cytoplasm (arrow).
M cell - dendritic cell interactions suggest functional units in Peyer’s patch
While M cell function may be dependent on basolateral pocket B lymphocytes, their critical cellular interaction in immune surveillance is their delivery of lumenal antigens to subepithelial antigen presenting cells. Therefore, to examine the relationship between M cells and underlying dendritic cells, we crossed the PGRP-S-dsRed transgene to the CX3CR1-EGFP reporter transgene that expresses a green fluorescent protein in myeloid cells such as DC (33). Because the dsRed and EGFP proteins fill the cytoplasm of the M cells and DC, their full morphology can be visualized by confocal microscopy. Interestingly, most red fluorescent PP M cells appeared to have a close association with an underlying green fluorescent DC, and in many cases dendritic processes can be identified extending up along the basolateral pocket of the M cells, where the local contact points between the cells appear as yellow fluorescence (Figure 3). This one-to-one association suggests the formation of an M cell-DC functional unit, though it may also reflect the high density of myeloid cells in the Peyer’s patch subepithelial zone.
Figure 3.
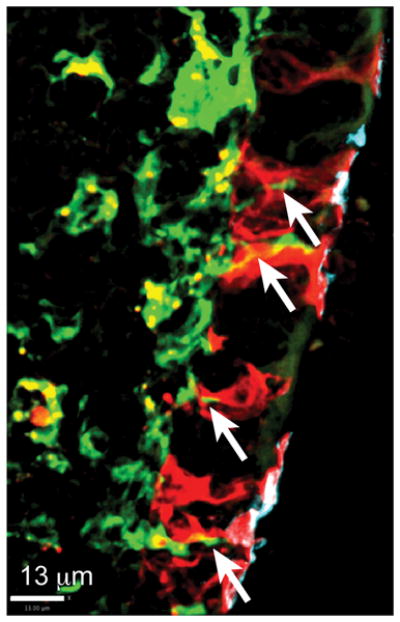
M cell - dendritic cell association. Section of CX3CR1-EGFP/PGRP-S-dsRed double transgenic Peyer’s patch follicle epithelium shows M cells with apical staining with UEA-1+ (cyan), cytoplasmic dsRed+ (red) in close contact with EGFP+ dendritic cells (green) under each M cell, where dendritic processes extend into M cells (arrows).
Cholera toxin induces de novo M cell transdifferentiation from enterocytes on Peyer’s patch
Several immune stimuli can activate the generation of new M cells within hours to days (16–18; 34). This phenomenon has been controversial, with various studies arguing that the induction is due to (a) rapid activation of M cell production from crypt stem cells, (b) acute transdifferentiation from mature enterocytes, or alternatively, (c) increased activity of existing M cells (35). It is possible that under different experimental conditions, one or more of these mechanisms may be acting at the same time. In our studies we focused only on the actual numbers of UEA-1+ M cells across the PP follicle epithelium, and we found that cholera toxin induced the development of up to 30% more M cells across the PP follicle epithelium within 48 hours (Figure 4A–C). Despite the increased numbers of M cells in the follicle epithelium, they maintained a dispersed distribution across the epithelium, in contrast to the clustering of M cells seen with villous M cells (see below). The effect of CT on intestinal M cells was similar whether the CT was administered intranasally or orally (Figure 4C); this could be due to oral ingestion of CT draining from nasal administration, although a direct or indirect systemic effect through cytokines such as TNFα (10) or RANKL (36) may also be responsible.
Figure 4.

Cholera toxin induces de novo M cell differentiation from enterocytes on Peyer’s patch follicle epithelium. Peyer’s patch whole mounts from BALB/c mice shows UEA-1+ M cells before (A) and after (B) cholera toxin treatment, and insets show cross sections of epithelium to show the normal morphology of the M cells (“G” indicates goblet cell). SEM images below show M cells on Peyer’s patch follicle epithelium of BALB/c mice before (A bottom) and after cholera toxin (B bottom). M cells are apparent as cells lacking brush border microvilli. (C), Peyer’s patch follicle M cell density in control (n=42), cholera toxin gavaged (n=50) and cholera toxin intranasally delivered (n=24). Each measurement represents one follicle, three mice per treatment group. M cell density is significantly increased in the treated groups (P<0.001). The columns at the right show similar CT-induced M cell numbers when only the inner 2/3 of the follicle is counted, suggesting that induction is uniform across the follicle dome. (D), CD137KO Peyer’s patch follicle M cell density in control (n=41) versus cholera toxin gavaged (n=47). (E), Igh-6 mice Peyer’s patch follicle M cell density in control (n=12) versus cholera toxin gavaged (n=12). Here, cholera toxin induced a significant increase in M cell density (P=0.0194). (F) Peyer’s patch from cholera toxin-treated BALB/c mice given EdU at the same time. EdU (red) labeled nearby crypt cells (arrow) but not the UEA-1 positive (green) M cells.
The induction of higher numbers of M cells by CT was not significant in the context of the CD137KO; however, here the non-treated M cell levels were already considerably higher than the background levels on wild type BALB/c (Figure 4D), suggesting that the knockout phenotype was associated with some chronic stimulation of M cells. Indeed, Lee et al. (37) reported that CD137KO mice show evidence for some immune hyper-reactivity, which might result in an increased basal level of mucosal stimulation of M cell production. In the case of B cell-deficient Igh-6 mice, CT induction of new M cells over follicle epithelium was clearly evident despite the absence of B cells or CD137 (Figure 4E). Thus, CT induction of new M cell lineage cells was largely along the lines of first step lineage commitment, independent of B cells.
The induction of new M cells was rapid, but sustained, consistent with the normal half-life of intestinal epithelium. The kinetics of this rapid increase suggested that the new M cells were generated from mature follicle enterocytes rather than from new cells emerging from the nearby crypts. We examined this question in two ways. First, we administered CT along with the nucleoside analogue EdU. Within the 48 hours of the M cell induction, EdU uptake was clearly evident within the neighboring intestinal crypts, but not in any of the follicle epithelial cells further from the crypts, including the UEA-1+ dsRed+ cells (Figure 4F). Thus, any acutely differentiated M cells appear to arise by direct transdifferentiation from mature enterocytes. Second, we looked at where the new M cells appeared on the follicle dome. Consistent with the EdU study, the increased density of M cells was similar when quantified separately for both the outer third of the follicle and the inner two thirds, indicating that the new M cells appeared uniformly across the dome and not just among newly formed cells coming from the crypts (Figure 4C).
Phenotypic and functional differences in CT-induced villous M cells
The apparent transdifferentiation of mature enterocytes to an M cell phenotype is especially prominent in the case of villous M cell development. In untreated mice, villous M cells are infrequently present at the tips of the intestinal villi (Figure 5A), but in response to CT, villi with clusters of UEA-1+ M cells are more easily identified (Figure 5B). Considering their location exclusively at the tips of villi, it was unlikely that they were newly generated from crypt stem cell precursors; accordingly, CT-induced villous M cells did not label with EdU during the induction period (Figure 5B), suggesting that these are also the product of transdifferentiation from mature enterocytes. Scanning electron microscopy confirmed the loss of brush border microvilli (Figure 5A,B). In contrast to PP follicle M cells, they were consistently found in clusters rather than dispersed as single M cells. Some individual villous M cells showed contacts with underlying DC (Figure 5C, arrows), though this was much less frequent compared to Peyer’s patch. Consistent with the differences in gene expression phenotype, the PGRP-S-dsRed transgene was also not expressed in these M cells (Figure 5C). As with follicle M cells, CT induction was not dependent on the presence of B cells or CD137 (not shown).
Figure 5.

Cholera toxin induces villous M cell differentiation from enterocytes. (A), Villous epithelium from untreated BALB/c mice stained with UEA-1 (green) showing villous M cells infrequently at the tips of the villi. Blue: nuclei. (B), Whole mount villous epithelium from BALB/c mice gavaged with cholera toxin, also injected with EdU at the same time. Inset shows induced villous M cells without EdU. The picture shows increased numbers of UEA-1+ villous M cells (green) clustered together at the villus tips. However, EdU (red) mainly labeled nearby crypt cells (arrow). SEM images of villous M cells in control villi (A bottom) and after cholera toxin (B bottom) showing villous M cells clustered at the tips of villi of mice. (C), Villous sections from control and cholera toxin gavaged CX3CR1-GFP/PGRP-S-dsRed double transgenic mice, taken with 20x and 40x objectives. UEA-1 positive villous M cells (cyan) did not express dsRed fluorescence, but occasional contacts between M cells and underlying myeloid cells (green) could be seen (arrows). Nuclei (blue or grey).
Isolated Lymphoid Follicle (ILF) M cells resemble Peyer’s patch M cells in PGRP-S-dsRed expression
As villous M cells are inducible by acute stimuli such as cholera toxin, other mucosal lymphoid tissues such as ILF can also be induced by immunological stimulation. In contrast to villous M cells, ILF have organized lymphoid tissues below the epithelium (38; 39), are dependent on lymphoid tissue inducer cells (LTi) (40; 41), and appear to be the consequence of chronic (rather than acute) stimulation so are found more frequently in older animals (42). Thus, it would be expected that ILF should develop epithelial M cells that are more similar to PP M cells rather than villous M cells. This was confirmed in older (5–12 month) PGRP-S-dsRed mice, where the small number of UEA-1+ epithelial cells overlying ILF (found in sections of intestine after PP were removed) also expressed dsRed (Figure 6A,B).
Figure 6.

PGRP-S-dsRed expression in ILF M cells. After removal of Peyer’s patches, intestinal sections from five to twelve month old PGRP-S-dsRed mice were examined for ILF M cells. A, Apical staining for UEA-1 (green) identifies M cells that also express dsRed (red, arrows). B220 staining (cyan), follicle (asterisk). Nuclei (blue). B, In double transgenic PGRP-S-dsRed/CX3CR1-EGFP mouse, cells were found in the intestine with apical UEA-1+ staining (cyan) and cytoplasmic dsRed (red, arrows). One M cell shows contact with an underlying dendritic cell (EGFP/green, with yellow arrow) and a small follicle (asterisk). Nuclei (blue).
NALT M cells express PGRP-S-dsRed transgene and can also develop from Foxj1+ ciliated progenitors
M cells in the mouse are also are present in airway epithelium over NALT. In contrast to intestinal M cells residing among enterocytes with apical brush border microvilli (formed by actin bundles), NALT M cells are found among epithelium with apical cilia (formed by tubulin filaments). Despite these distinct origins, airway and intestinal M cells could show similarities in their development and genetic program. For example, similar to intestinal M cells, we found that NALT M cell initial lineage commitment is also independent of B cells and CD137 (22). However, in view of the different developmental origins of M cell populations in the airway versus intestine, the M cell phenotype may be alternatively viewed as a convergent development toward a common functional phenotype. For example, in contrast to intestinal epithelium, airway epithelial cells are generated from basal cell precursors (23; 24), which give rise to both ciliated airway epithelium and Clara cells. Ciliated airway epithelium expresses the transcription factor Foxj1, associated with terminal differentiation, and they have a very long half life, on the order of weeks or months (23; 25; 26; 43). In response to chemical injury, new Clara cells are quickly formed, and evidence suggests that they come from the basal cells and not from transdifferentiation of mature Foxj1+ ciliated epithelium. Although this has not been directly studied in the case of NALT epithelium, we expected similarities in Foxj1 expression and epithelial cell turnover.
As with the intestine, the PGRP-S-dsRed reporter was expressed in a subset of airway epithelium, but unlike PP, expression was not limited to UEA-1+ cells (Figure 7A). CT (both intranasal or gavage administration) can also induce rapid development of new M cells in the NALT epithelium (Figure 7B), and here too transdifferentiation from neighboring mature ciliated epithelial cells may be involved. In Foxj1-EGFP transgenic mice (26), NALT ciliated epithelial cells expressed the EGFP reporter, while UEA-1+ M cells were all negative for EGFP (Figure 7C). CT treatment significantly increased the number of NALT M cells, and they were all EGFP negative (Figure 7D). Expression of the PGRP-S-dsRed transgene in the NALT was different from the pattern seen in the intestine, as many dsRed+ cells were UEA-1 negative, indicating that they were not mature M cells (Figure 7C). Interestingly, in Foxj1-EGFP/PGRP-S-dsRed double transgenic mice, numerous UEA-1 negative cells were found expressing both EGFP and dsRed, apparent as yellow fluorescence (Figure 7C). When these mice were induced by intranasal CT, these double expressing cells were lost, and seemed to be replaced by new NALT M cells that were UEA-1+ and dsRed+ (Figure 7D,E), as the cell counts showed that the double expressing cells were similar in number to the incremental increase in M cells (Figure 7E). These numbers are consistent with the new M cells arising from dsRed+EGFP+ cells that lost Foxj1(EGFP) expression but retained PGRP-S(dsRed) during the transition to UEA-1+ M cells. None of the NALT epithelial cells labeled with EdU during the induction (not shown), so proliferation of progenitors, whether basal cells or existing M cells, was unlikely. Thus, in contrast to the injury-induced production of Clara cells, at least some of the NALT M cell production may come from mature progenitors expressing differentiation associated genes from two distinct lineages (PGRP-S and Foxj1).
Figure 7.

NALT M cells can develop from Foxj1+ progenitors. (A) NALT from PGRP-S-dsRed transgenic mice shows UEA-1+ NALT M cells (cyan) among epithelial cells expressing dsRed (red). (B) NALT M cell density in control (n=22), cholera toxin gavaged (n=20) and cholera toxin intranasally delivered (n=19). Each count represents one image, three mice per treatment group. M cell density was significantly increased in the treated groups (***, P<0.001). (C) NALT from Foxj1-GFP/PGRP-S-dsRed double transgenic mice. All the UEA-1+Foxj1negative NALT M cells (cyan) and some of the UEA-1negative Foxj1+ ciliated epithelial cells (green) expressed dsRed (red, arrow). (D) NALT from Foxj1-GFP/PGRP-S-dsRed double transgenic mice treated with intranasal cholera toxin. After treatment, UEA-1+ NALT M cell numbers increased; moreover, there were no longer UEA-1-Foxj1+ ciliated epithelial cells (green) expressing dsRed fluorescence. (E). NALT M cell density count in Foxj1-GFP/PGRP-S-dsRed double transgenic mice showing that UEA-1negative Foxj1-GFP/PGRP-S-DsRed double expressing cells in control mice were similar in number to the increased number of new M cells in the cholera toxin treated mice. Counts were from individual confocal images (n=4 for UEA-1+ and UEA-1- untreated mice, n=6 for CT-treated mice), two mice per group. (***, P<0.001)
Discussion
The results of our studies here, along with other recent reports, support the identification of at least three distinct mucosal M cell subsets: PP/ILF follicle epithelium M cells, inducible villous M cells, and NALT M cells. A recent report identified a fourth type of M cell, a “respiratory” M cell (44), present in the upper airway, which is likely to be an airway analogue of the villous M cell. Given the distinct morphological and functional differences among the subsets, we propose to define broad categories of M cells as “organized” (PP, ILF, and NALT) versus “sporadic” (villous and respiratory). The organized M cells are found in epithelium overlying organized mucosal lymphoid tissues, and they appear to form functional units with underlying lymphoid tissue dendritic cells. Despite the differences in their developmental origins and life span, one characteristic shared by these cells is their dispersed single-cell distribution across the face of the follicle epithelium. In addition, they express a subset of specific genes that appear helpful to function in microbial uptake, including PGRP-S (27) and gp2 (31).
By contrast, the sporadic subsets are scattered in various sites along the mucosal epithelium, not associated with underlying organized lymphoid tissues. Because of their scattered distribution, it can be difficult to provide consistent quantification, but it appears that inducible M cells are dependent on immune stimuli to develop in significant numbers. This may be driven in part by the inducible stromal cell expression of RANKL, as direct administration of RANKL was very effective in inducing significant numbers of villous M cells (36). Due to the appearance of inducible villous M cells away from lymphoid tissues, any associations with underlying dendritic cells appeared random at best. Interestingly, in contrast to the dispersion of organized M cells, sporadic M cells could be found in dense clusters, such as at the tips of intestinal villi. Finally, function-associated genes PGRP-S and gp2 were not expressed by these cells, which appears to correlate with some reduced transcytosis function.
The situation becomes more complex as we consider the acute induction of new M cells by cholera toxin treatment. The numbers of M cells belonging to both the organized and sporadic subsets increased significantly. However, the striking result here was that the phenotype of the new PP and NALT M cells conformed strictly to the local phenotype; thus, new PP and NALT M cells all expressed the PGRP-S-dsRed transgene, while new villous M cells were all negative for the reporter transgene expression. Similarly, the dispersed single-cell distribution of M cells across the face of the PP and NALT epithelium was retained despite the increased density.
The cholera toxin-inducible M cells in both airway and intestine all appeared to be rapidly produced without evidence for cell division (i.e., EdU uptake) or recent crypt origin. The simplest explanation for the development of these cells is that they are the product of transdifferentiation from mature enterocytes or ciliated airway epithelium. Interestingly, the finding of double expressing Foxj1-EGFP/PGRP-S-dsRed cells in the NALT epithelium raises the possibility that latent M cell precursors may already exist among mature mucosal epithelium. That is, a subset of differentiated ciliated airway epithelium may specifically retain the ability to convert to an M cell phenotype, losing their apical cilia. Unfortunately, there is no evidence for a similar follicle epithelium subset on Peyer’s patches, since all PGRP-S-dsRed expressing epithelial cells were already also UEA-1+.
An important implication of the distinct phenotypes and functions of the organized versus sporadic M cell subsets is that the differential uptake of specific types of microbes will change as the balance of M cell subsets changes (8). Since various inflammatory and infectious conditions (such as the microbial components described above) can trigger the production of the sporadic phenotype, the nature of the mucosal immune response can change as local tissue responses change. There are two principal direct differences in the contributions of organized versus sporadic M cells: First, since sporadic M cells are less often associated with underlying dendritic cells and organized lymphoid tissues, uptake through these cells is far less likely to promote the production of secretory IgA, as the isotype switching of B cells is mainly associated with the organized mucosal lymphoid tissues (41; 45; 46). Second, the sporadic M cells do not express the sets of genes associated with selective microbe uptake (e.g., gp2 and PGRP-S), so along with their relative inefficiency, they will enable a different array of microbes to cross the mucosal barrier. Since any potential immune response is more likely to be initiated in draining lymphoid tissues such as the mesenteric lymph node, production of secretory IgA is likely to be less prominent, with more dependence on IgG and T cell responses.
While our studies were concerned with microbial (e.g., cholera toxin) triggers of M cell development, other settings involving mucosal inflammation (and TNF/lymphotoxin cytokines) should also have an effect on the various M cell subsets present. However, since clear genetic markers for M cells have not been available until recently (at least for mouse), this question has not been closely studied in situations such as models of inflammatory bowel disease or ileitis. Here, the PGRP-S-dsRed transgenic strain should be useful in identifying differential induction of M cell subsets in vivo. The differential microbial uptake by M cell subsets may have an important effect on the progression or persistence of intestinal inflammation, in part through the differences in the induced mucosal immune response. As the relationship between M cell function and mucosal immunity becomes more clearly defined, these questions will begin to be addressed.
Finally, we found that the PGRP-S promoter provided a useful reporter of M cell lineage commitment, corresponding to the expression of the peptidoglycan recognition protein in M cells. It is therefore interesting to consider the more narrow question of whether peptidoglycan has any direct influence on M cell development. PGRP-S is a secreted protein known for its potential antimicrobial activity (47; 48) but it has no known signaling function; a PGRP-S-deficient mouse has not been examined for M cell development or function (28). Importantly, several other proteins expressed in mucosal epithelium can signal in response to peptidoglycan, including TLR2, NOD2, and other PGRP family members such as PGRP-L, PGRP-Iα, and PGRP-Iβ (30). Chabot et al. (49) found that peptidoglycan could induce a very rapid (45–90 minute) recruitment of dendritic cells into follicle epithelium and an increase in M cell particle uptake. This effect was absent in TLR2-deficient mice, so no other peptidoglycan recognition proteins appear to be involved in this response. Though not directly examined by Chabot et al., the effect may be transient; we studied M cell numbers and particle uptake in TLR2-deficient mice and found no significant differences (V. Gusti, D. Lo, unpublished). Thus, at least in this context, peptidoglycan and TLR2 signaling might not influence M cell lineage commitment or long term function.
Supplementary Material
Acknowledgments
The authors thank Larry Ostrowski and Brigid Hogan for discussions on Foxj1 studies, and Marian Neutra for discussions on the PGRP-S-dsRed transgenic mice. We thank the UC Irvine Transgenic Mouse Facility and Tom Fielder for production of the transgenic mice.
Abbreviations
- DC
dendritic cells
- CT
cholera toxin
- PP
Peyer’s patch
- NALT
Nasal Associated Lymphoid Tissue
- BALT
Bronchus Associated Lymphoid Tissue
- ILF
Isolated Lymphoid Follicles
- UEA-1
Ulex Europaeus Agglutinin 1
- PGRP-S
PeptidoGlycan Recognition Protein – S
- EdU
5-ethynyl-2′-deoxyuridine
Footnotes
This work was supported by NIH grants AI63426 and AI73689 to DDL.
Disclosures: The authors have no conflicts of interest.
References
- 1.Kraehenbuhl JP, Neutra MR. Epithelial M cells: differentiation and function. Annu Rev Cell Dev Biol. 2000;16:301–332. doi: 10.1146/annurev.cellbio.16.1.301. [DOI] [PubMed] [Google Scholar]
- 2.Neutra MR, Frey A, Kraehenbuhl JP. Epithelial M cells: gateways for mucosal infection and immunization. Cell. 1996;86:345–348. doi: 10.1016/s0092-8674(00)80106-3. [DOI] [PubMed] [Google Scholar]
- 3.Neutra MR, Pringault E, Kraehenbuhl JP. Antigen sampling across epithelial barriers and induction of mucosal immune responses. Annu Rev Immunol. 1996;14:275–300. doi: 10.1146/annurev.immunol.14.1.275. [DOI] [PubMed] [Google Scholar]
- 4.Owen RL, Jones AL. Epithelial cell specialization within human Peyer’s patches: an ultrastructural study of intestinal lymphoid follicles. Gastroenterology. 1974;66:189–203. [PubMed] [Google Scholar]
- 5.Hameleers DM, van der Ende M, Biewenga J, Sminia T. An immunohistochemical study on the postnatal development of rat nasal-associated lymphoid tissue (NALT) Cell Tissue Res. 1989;256:431–438. doi: 10.1007/BF00218901. [DOI] [PubMed] [Google Scholar]
- 6.Rosner AJ, Keren DF. Demonstration of M cells in the specialized follicle-associated epithelium overlying isolated lymphoid follicles in the gut. J Leukoc Biol. 1984;35:397–404. doi: 10.1002/jlb.35.4.397. [DOI] [PubMed] [Google Scholar]
- 7.Gebert A, Hach G. Vimentin antibodies stain membranous epithelial cells in the rabbit bronchus-associated lymphoid tissue (BALT) Histochemistry. 1992;98:271–273. doi: 10.1007/BF00271042. [DOI] [PubMed] [Google Scholar]
- 8.Jang MH, Kweon MN, Iwatani K, Yamamoto M, Terahara K, Sasakawa C, Suzuki T, Nochi T, Yokota Y, Rennert PD, Hiroi T, Tamagawa H, Iijima H, Kunisawa J, Yuki Y, Kiyono H. Intestinal villous M cells: an antigen entry site in the mucosal epithelium. Proc Natl Acad Sci USA. 2004;101:6110–6115. doi: 10.1073/pnas.0400969101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Finke D, Acha-Orbea H, Mattis A, Lipp M, Kraehenbuhl J. CD4+CD3-cells induce Peyer’s patch development: role of alpha4beta1 integrin activation by CXCR5. Immunity. 2002;17:363–373. doi: 10.1016/s1074-7613(02)00395-3. [DOI] [PubMed] [Google Scholar]
- 10.Wang J, Lopez-Fraga M, Rynko A, Lo DD. TNFR and LTbetaR agonists induce follicle-associated epithelium and M cell specific genes in rat and human intestinal epithelial cells. Cytokine. 2009;47:69–76. doi: 10.1016/j.cyto.2009.05.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Creamer B. The turnover of the epithelium of the small intestine. Br Med Bull. 1967;23:226–230. doi: 10.1093/oxfordjournals.bmb.a070561. [DOI] [PubMed] [Google Scholar]
- 12.Bye WA, Allan CH, Trier JS. Structure, distribution, and origin of M cells in Peyer’s patches of mouse ileum. Gastroenterology. 1984;86:789–801. [PubMed] [Google Scholar]
- 13.Gebert A, Posselt W. Glycoconjugate expression defines the origin and differentiation pathway of intestinal M-cells. J Histochem Cytochem. 1997;45:1341–1350. doi: 10.1177/002215549704501003. [DOI] [PubMed] [Google Scholar]
- 14.Gebert A, Fassbender S, Werner K, Weissferdt A. The development of M cells in Peyer’s patches is restricted to specialized dome-associated crypts. Am J Pathol. 1999;154:1573–1582. doi: 10.1016/S0002-9440(10)65410-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Kernéis S, Bogdanova A, Kraehenbuhl JP, Pringault E. Conversion by Peyer’s patch lymphocytes of human enterocytes into M cells that transport bacteria. Science. 1997;277:949–952. doi: 10.1126/science.277.5328.949. [DOI] [PubMed] [Google Scholar]
- 16.Meynell HM, Thomas NW, James PS, Holland J, Taussig MJ, Nicoletti C. Up-regulation of microsphere transport across the follicle-associated epithelium of Peyer’s patch by exposure to Streptococcus pneumoniae R36a. FASEB J. 1999;13:611–619. doi: 10.1096/fasebj.13.6.611. [DOI] [PubMed] [Google Scholar]
- 17.Borghesi C, Regoli M, Bertelli E, Nicoletti C. Modifications of the follicle-associated epithelium by short-term exposure to a non-intestinal bacterium. J Pathol. 1996;180:326–332. doi: 10.1002/(SICI)1096-9896(199611)180:3<326::AID-PATH656>3.0.CO;2-6. [DOI] [PubMed] [Google Scholar]
- 18.Terahara K, Nochi T, Yoshida M, Takahashi Y, Goto Y, Hatai H, Kurokawa S, Jang MH, Kweon MN, Domino SE, Hiroi T, Yuki Y, Tsunetsugu-Yokota Y, Kobayashi K, Kiyono H. Distinct fucosylation of M cells and epithelial cells by Fut1 and Fut2, respectively, in response to intestinal environmental stress. Biochemical and Biophysical Research Communications. 2011;404:822–828. doi: 10.1016/j.bbrc.2010.12.067. [DOI] [PubMed] [Google Scholar]
- 19.Mach J, Hshieh T, Hsieh D, Grubbs N, Chervonsky A. Development of intestinal M cells. Immunol Rev. 2005;206:177–189. doi: 10.1111/j.0105-2896.2005.00281.x. [DOI] [PubMed] [Google Scholar]
- 20.Golovkina TV, Shlomchik M, Hannum L, Chervonsky A. Organogenic role of B lymphocytes in mucosal immunity. Science. 1999;286:1965–1968. doi: 10.1126/science.286.5446.1965. [DOI] [PubMed] [Google Scholar]
- 21.Kernéis S, Caliot E, Stubbe H, Bogdanova A, Kraehenbuhl J, Pringault E. Molecular studies of the intestinal mucosal barrier physiopathology using cocultures of epithelial and immune cells: a technical update. Microbes Infect. 2000;2:1119–1124. doi: 10.1016/s1286-4579(00)01266-1. [DOI] [PubMed] [Google Scholar]
- 22.Hsieh EH, Fernandez X, Wang J, Hamer M, Calvillo S, Croft M, Kwon BS, Lo DD. CD137 Is Required for M Cell Functional Maturation but Not Lineage Commitment. Am J Pathol. 2010;177:666–676. doi: 10.2353/ajpath.2010.090811. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Rawlins EL, Hogan BLM. Ciliated epithelial cell lifespan in the mouse trachea and lung. Am J Physiol Lung Cell Mol Physiol. 2008;295:L231–234. doi: 10.1152/ajplung.90209.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Rock JR, Onaitis MW, Rawlins EL, Lu Y, Clark CP, Xue Y, Randell SH, Hogan BLM. Basal cells as stem cells of the mouse trachea and human airway epithelium. Proc Natl Acad Sci USA. 2009;106:12771–12775. doi: 10.1073/pnas.0906850106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Rawlins EL, Ostrowski LE, Randell SH, Hogan BLM. Lung development and repair: contribution of the ciliated lineage. Proc Natl Acad Sci USA. 2007;104:410–417. doi: 10.1073/pnas.0610770104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Ostrowski LE, Hutchins JR, Zakel K, O’Neal WK. Targeting expression of a transgene to the airway surface epithelium using a ciliated cell-specific promoter. Mol Ther. 2003;8:637–645. doi: 10.1016/s1525-0016(03)00221-1. [DOI] [PubMed] [Google Scholar]
- 27.Lo D, Tynan W, Dickerson J, Mendy J, Chang HW, Scharf M, Byrne D, Brayden D, Higgins L, Evans C, O’Mahony DJ. Peptidoglycan recognition protein expression in mouse Peyer’s Patch follicle associated epithelium suggests functional specialization. Cell Immunol. 2003;224:8–16. doi: 10.1016/s0008-8749(03)00155-2. [DOI] [PubMed] [Google Scholar]
- 28.Cho JH, I, Fraser P, Fukase K, Kusumoto S, Fujimoto Y, Stahl GL, Ezekowitz RAB. Human peptidoglycan recognition protein S is an effector of neutrophil-mediated innate immunity. Blood. 2005;106:2551–2558. doi: 10.1182/blood-2005-02-0530. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Dziarski R. Peptidoglycan recognition proteins (PGRPs) Mol Immunol. 2004;40:877–886. doi: 10.1016/j.molimm.2003.10.011. [DOI] [PubMed] [Google Scholar]
- 30.Dziarski R, Gupta D. Review: Mammalian peptidoglycan recognition proteins (PGRPs) in innate immunity. Innate Immun. 2010;16:168–174. doi: 10.1177/1753425910366059. [DOI] [PubMed] [Google Scholar]
- 31.Hase K, Kawano K, Nochi T, Pontes GS, Fukuda S, Ebisawa M, Kadokura K, Tobe T, Fujimura Y, Kawano S, Yabashi A, Waguri S, Nakato G, Kimura S, Murakami T, Iimura M, Hamura K, Fukuoka SI, Lowe AW, Itoh K, Kiyono H, Ohno H. Uptake through glycoprotein 2 of FimH(+) bacteria by M cells initiates mucosal immune response. Nature. 2009;462:226–230. doi: 10.1038/nature08529. [DOI] [PubMed] [Google Scholar]
- 32.Terahara K, Yoshida M, Igarashi O, Nochi T, Pontes GS, Hase K, Ohno H, Kurokawa S, Mejima M, Takayama N, Yuki Y, Lowe AW, Kiyono H. Comprehensive gene expression profiling of Peyer’s patch M cells, villous M-like cells, and intestinal epithelial cells. J Immunol. 2008;180:7840–7846. doi: 10.4049/jimmunol.180.12.7840. [DOI] [PubMed] [Google Scholar]
- 33.Jung S, Aliberti J, Graemmel P, Sunshine MJ, Kreutzberg GW, Sher A, Littman DR. Analysis of fractalkine receptor CX(3)CR1 function by targeted deletion and green fluorescent protein reporter gene insertion. Mol Cell Biol. 2000;20:4106–4114. doi: 10.1128/mcb.20.11.4106-4114.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Chabot SM, Shawi M, Eaves-Pyles T, Neutra MR. Effects of flagellin on the functions of follicle-associated epithelium. J Infect Dis. 2008;198:907–910. doi: 10.1086/591056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Gebert A, Steinmetz I, Fassbender S, Wendlandt K-H. Antigen transport into Peyer’s patches: increased uptake by constant numbers of M cells. Am J Pathol. 2004;164:65–72. doi: 10.1016/S0002-9440(10)63097-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Knoop KA, Kumar N, Butler BR, Sakthivel SK, Taylor RT, Nochi T, Akiba H, Yagita H, Kiyono H, Williams IR. RANKL is necessary and sufficient to initiate development of antigen-sampling M cells in the intestinal epithelium. J Immunol. 2009;183:5738–5747. doi: 10.4049/jimmunol.0901563. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Lee SW, Vella AT, Kwon BS, Croft M. Enhanced CD4 T cell responsiveness in the absence of 4-1BB. J Immunol. 2005;174:6803–6808. doi: 10.4049/jimmunol.174.11.6803. [DOI] [PubMed] [Google Scholar]
- 38.Newberry RD, Lorenz RG. Organizing a mucosal defense. Immunol Rev. 2005;206:6–21. doi: 10.1111/j.0105-2896.2005.00282.x. [DOI] [PubMed] [Google Scholar]
- 39.McDonald KG, McDonough JS, Wang C, Kucharzik T, Williams IR, Newberry RD. CC chemokine receptor 6 expression by B lymphocytes is essential for the development of isolated lymphoid follicles. Am J Pathol. 2007;170:1229–1240. doi: 10.2353/ajpath.2007.060817. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Lügering A, Ross M, Sieker M, Heidemann J, Williams IR, Domschke W, Kucharzik T. CCR6 identifies lymphoid tissue inducer cells within cryptopatches. Clin Exp Immunol. 2010;160:440–449. doi: 10.1111/j.1365-2249.2010.04103.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Tsuji M, Suzuki K, Kitamura H, Maruya M, Kinoshita K, Ivanov II, Itoh K, Littman DR, Fagarasan S. Requirement for lymphoid tissue-inducer cells in isolated follicle formation and T cell-independent immunoglobulin A generation in the gut. Immunity. 2008;29:261–271. doi: 10.1016/j.immuni.2008.05.014. [DOI] [PubMed] [Google Scholar]
- 42.McDonald KG, Leach MR, Huang C, Wang C, Newberry RD. Aging impacts isolated lymphoid follicle fevelopment and function. Immun Ageing. 2011;8:1. doi: 10.1186/1742-4933-8-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.You Y, Huang T, Richer EJ, Schmidt JEH, Zabner J, Borok Z, Brody SL. Role of f-box factor foxj1 in differentiation of ciliated airway epithelial cells. Am J Physiol Lung Cell Mol Physiol. 2004;286:L650–657. doi: 10.1152/ajplung.00170.2003. [DOI] [PubMed] [Google Scholar]
- 44.Kim DY, Sato A, Fukuyama S, Sagara H, Nagatake T, Kong IG, Goda K, Nochi T, Kunisawa J, Sato S, Yokota Y, Lee CH, Kiyono H. The airway antigen sampling system: respiratory m cells as an alternative gateway for inhaled antigens. J Immunol. 2011;186:4253–4262. doi: 10.4049/jimmunol.0903794. [DOI] [PubMed] [Google Scholar]
- 45.Craig SW, Cebra JJ. Peyer’s patches: an enriched source of precursors for IgA-producing immunocytes in the rabbit. J Exp Med. 1971;134:188–200. doi: 10.1084/jem.134.1.188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Husband AJ, Gowans JL. The origin and antigen-dependent distribution of IgA-containing cells in the intestine. J Exp Med. 1978;148:1146–1160. doi: 10.1084/jem.148.5.1146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Tydell CC, Yount N, Tran D, Yuan J, Selsted ME. Isolation, characterization, and antimicrobial properties of bovine oligosaccharide-binding protein. A microbicidal granule protein of eosinophils and neutrophils. J Biol Chem. 2002;277:19658–19664. doi: 10.1074/jbc.M200659200. [DOI] [PubMed] [Google Scholar]
- 48.Tydell CC, Yuan J, Tran P, Selsted ME. Bovine peptidoglycan recognition protein-S: antimicrobial activity, localization, secretion, and binding properties. J Immunol. 2006;176:1154–1162. doi: 10.4049/jimmunol.176.2.1154. [DOI] [PubMed] [Google Scholar]
- 49.Chabot S, Wagner JS, Farrant S, Neutra MR. TLRs regulate the gatekeeping functions of the intestinal follicle-associated epithelium. J Immunol. 2006;176:4275–4283. doi: 10.4049/jimmunol.176.7.4275. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.