

Abstract
Objective
The truncated mitochondrial progesterone receptor (PR-M) is homologous to nuclear PRs with the exception of an amino terminus hydrophobic membrane localization sequence, which localizes PR-M to mitochondria. Given the matrilineal inheritance of both spontaneous preterm birth (SPTB) and the mitochondrial genome, we hypothesized that (a) PR-M is polymorphic and (b) PR-M localization sequence polymorphisms could result in variable progesterone mitochondrial effects and variable responsiveness to progesterone prophylaxis.
Methods
Secondary analysis of DNA from women enrolled in a multicenter, prospective, study of 17 alpha-hydroxyprogesterone caproate (17OHPC) versus placebo for the prevention of recurrent SPTB. DNA was extracted from stored saliva.
Results
The PR-M localization sequence was sequenced on 344 patients. Sequences were compared with the previously published 48 base-pair sequence, and all were identical.
Conclusions
We did not detect genetic variation in the mitochondrial localization sequence of the truncated PR-M in a group of women at high risk for SPTB.
Keywords: mitochondria, prematurity, progesterone
Background
17 alpha-hydroxyprogesterone caproate (17OHPC) and other progesterone agents have been shown to reduce the recurrence of spontaneous preterm birth (SPTB) in high-risk women.1-3 Over 12% of infants are delivered preterm, accounting for 70% of neonatal morbidity and mortality in the United States.4 Some proportion of SPTB is inherited, as it frequently clusters in families, and maternal genotype appears to contribute to the risk of developing this complication.5-8 The mechanism by which progesterone reduces SPTB is unknown but may occur at the level of the progesterone receptor (PR).
The human PR (hPR) is a member of the steroid and thyroid receptor superfamily. The gene encoding this receptor is located on chromosome 11q22-23 and consists of 8 exons. Nuclear PRs exist primarily as 2 distinct isoforms, PR-A and PR-B, and have been found in gestational tissues including the amion and chorion.9,10 Progesterone binding induces conformational changes in the receptor, which lead to protein phosphorylation, dissociation from heat shock proteins, dimer formation, and nuclear transport of the active protein progesterone complex. These hormone-receptor complexes then bind to specific promoter regions of progesterone-responsive genes and act as transcription factors for these target genes, altering gene expression.11
Progesterone is also responsible for rapid, nongenomic actions that cannot be explained through traditional steroid-receptor binding. Nongenomic actions of progesterone have been previously shown to increase intracellular calcium and regulate the relaxation of intestinal and uterine muscle.12-14 Recently, a truncated progesterone receptor (mitochondrial progesterone receptor [PR-M]) has been described.15 The complementary DNA (cDNA) for PR-M was originally cloned from human aortic and adipose cDNA libraries. Sequence analysis shows a unique 5′ untranslated region, Kozack sequence, and coding sequence for the first 16 amino acids derived from intron sequence of the distal third intron of the PR gene. The remainder of the cDNA sequence is identical to exons 4 through 8 of the nuclear PR. The unique amino terminus results in a hydrophobic mitochondrial localization signal.16 The PR-M receptor and protein has been found in high levels in the myometrium of pregnant women. Nongenomic actions of progesterone (including uterine muscle relaxation) may be initiated through this type of receptor.17
We hypothesize that the hydrophobic localization sequence of PR-M is polymorphic. Given the matrilineal inheritance of PTB and the matrilineal inheritance of the mitochondrial genome, we further hypothesize that polymorphisms in the mitochondrial localization sequence of PR-M would correlate with the variable responsiveness to 17OHPC for the prevention of recurrent PTB.
Methods
This is a secondary analysis of women enrolled from September 1999 to February 2002 in a multicenter, prospective, double-blind, randomized controlled trial of 17OHPC versus placebo, conducted by the Eunice Kennedy Shriver National Institute of Child Health and Human Development (NICHD) Maternal Fetal Medicine Units (MFMU) Network.1 The trial enrolled 463 women with a singleton gestation who had at least 1 prior SPTB, and randomized them to receive weekly injections of either 17OHPC (n 310) or placebo (n = 153), beginning at 16 to 203/7 weeks gestation = and continuing until 366/7 weeks gestation or delivery. The trial demonstrated a reduction in the rate of recurrent PTB from 54.9% in the placebo group to 36.3% in the treatment group (P < .001).
Institutional Review Board (IRB) approval and subject consent for the original study, as well as future analyses such as this study, were obtained at each of the 19 participating Network sites by trained research nurses.1
As a part of the original trial protocol, maternal saliva samples were collected for future analyses. Saliva samples were originally labeled with unique, de-identified study codes and frozen at −20°C prior to DNA extraction in July and August 2008. Genomic DNA was extracted from samples using established methods (Puregene, Qiagen Systems, Valencia, California). DNA was then amplified using Invitrogen Platinum Taq DNA Polymerase (Invitrogen, Carlsbad, California); amplicons were verified by electrophoresis, and excess primers and nucleotides were removed by the ExoSAP-IT reaction (Affymetrix, Cleveland, Ohio). The purified polymerase chain reaction (PCR) product was then sequenced for the hydrophobic localization sequence of PR-M using BigDye Terminator v1.1 (Applied Biosystems, Carlsbad, California) and custom primers, designed for the sequence of interest. The resultant DNA sequence was then compared to the previously published sequence.15
Results
DNA was successfully extracted from stored saliva samples of 367 patients with a history of at least 1 prior SPTB. DNA was unable to be extracted in sufficient quantity from 96 of the 463 patients in the original study. Gel electrophoresis of the PCR amplification product demonstrated isolation of the 48 base-pair PR-M localization sequence (Figure 1). The localization sequence of PR-M was then successfully sequenced on 344 patients (Figure 2). All 344 samples were found to have identical DNA sequences. This sequence was then compared to the previously published 48 base-pair sequence and was found to be identical. No genetic variation was observed in this signal sequence.
Figure 1.
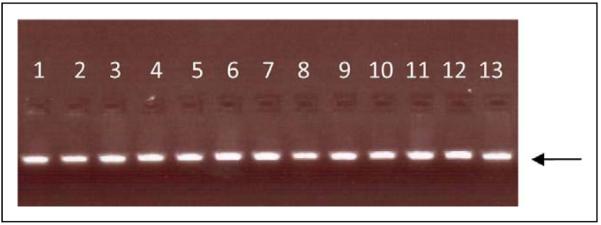
Gel electrophoresis (1.5% agarose gel) of the PR-M localization sequence demonstrating isolation of the 48 base-pair PCR amplification product used for sequencing. The arrow indicates the gel position of 50 base pairs (15 ng). Thirteen samples are shown, with 1 sample per lane. PR-M indicates mitochondrial progesterone receptor; PCR, polymerase chain reaction.
Figure 2.

The 16 novel amino acids (and 48 corresponding base pairs) comprising the hydrophobic localization sequence of progesterone-receptor M (PR-M). This sequence was identical in all 351 genotyped patients.
Discussion
The mitochondrial localization sequence of PR-M in 344 women with at least 1 prior SPTB is without variation and is genetically identical to the published sequence. Variation in responsiveness to 17OHPC for the prevention of recurrent PTB cannot be explained by examination of the PR-M mitochondrial localization sequence.
Numerous studies have demonstrated the actions of progesterone in the regulation of arterial and muscular relaxation throughout the body. Progesterone acts both through (1) traditional genomic pathways by binding a nuclear PR, forming a steroid receptor complex, and altering gene transcription and (2) nongenomic pathways, which are less clearly understood but are responsible for the rapid actions of this hormone and may be modulated through membrane receptors.
The action of progesterone varies among tissues. For example, in the gallbladder, progesterone acts through nongenomic-signaling pathways including tyrosine kinase and cyclic adenosine monophosphate activity.18 In vascular smooth muscle, PR variation has been noted, and theorized to be one potential cause of variable responsiveness and action in response to progesterone.19 In uterine smooth muscle cells, progesterone decreases calcium influx and thus uterine contractility.14,20 The tissue specificity of progesterone action is likewise thought to mediated by tissue-specific PR expression and function.21
Mitochondria are vital cell components involved with cellular energy and regulation of cellular apoptosis. Each mitochondrion has its own genome consisting of double-stranded mitochondrial DNA. Traditionally, mitochondrial action was thought to be regulated by second messengers. However, recent studies have confirmed the presence of steroid receptors (including glucocorticoid and estrogen receptors) in the mitochondrion.22-25 Neither PR isoform A nor B has been shown to localize to the mitochondria.26 However, the PR-M receptor is a recently described receptor and has recently been shown to localize to the mitochondrial membrane.16 Additionally, PR-M messenger RNA (mRNA) levels vary among different tissues in the body, with the highest levels found in muscular tissues including the heart and myometrium.27
Despite localization of PR-M to the mitochondria, along with the known roles of mitochondria in the production and utilization of cellular energy and progesterone in muscle contractility, the PR-M hydrophobic mitochondrial localization sequence is not polymorphic and does not appear to contribute toward the variable responsiveness to progesterone for preterm birth prevention. Future studies should investigate other potential mechanisms of action of progesterone, which might include detailed sequencing of the known genomic PRs and correlating these findings with clinical outcomes.
Acknowledgments
The authors wish to thank the following subcommittee members who participated in protocol development and coordination between clinical research centers (Allison Northen, RN, BSN), protocol/ data management and statistical analysis (Yinglei Lai, PhD), and protocol development and oversight (Michael W. Varner, MD and Mark B. Klebanoff, MD). In addition to the authors, other members of the Eunice Kennedy Shriver National Institute of Child Health and Human Development Maternal-Fetal Medicine Units Network are as follows:
University of Utah—M. Varner (University of Utah Health Sciences Center), E. Taggart (University of Utah Health Sciences Center), M. Belfort (Intermountain Healthcare)
University of Alabama at Birmingham—A. Northen, J. Hauth
Brown University—H. Silver, J. Tillinghast
Case Western Reserve University—P. Catalano, C. Milluzzi
University of Chicago—A.H. Moawad, P. Jones, M. Lindheimer
University of Cincinnati—N. Elder, T. Siddiqi
Columbia University—M. D’Alton, V. Pemberton
University of Pittsburgh—S. Caritis, M. Cotroneo, K. Lain
University of Miami—C. Alfonso, S. Beydoun
University of North Carolina, Chapel Hill—K. Dorman, K. Moise
Northwestern University—G. Mallet, M. Socol
The Ohio State University—F. Johnson, M. Landon
University of Tennessee—R. Ramsey
University of Texas at San Antonio—O. Langer, S. Nicholson
The University of Texas Health Science Center at Houston—M. C. Day, L. Gilstrap
University of Texas Southwestern Medical Center—J. Gold, G. Wendel
Drexel University—M. DiVito, J. Tolosa
Wake Forest University Health Sciences—E. Mueller-Heubach, M. Swain
Wayne State University—G. Norman, Y. Sorokin
The George Washington University Biostatistics Center— A. Das, E. Thom, S. Gilbert
Eunice Kennedy Shriver National Institute of Child Health and Human Development—M. Klebanoff, D. McNellis, S. Tolivaisa
MFMU Network Steering Committee Chair (Vanderbilt University Medical Center)—S. Gabbe
DNA extraction and analysis were conducted by Taueret Laboratories, Salt Lake City, UT. Taueret received a copy of the draft manuscript but had no input into the analysis of the data or the content of the manuscript.
Funding
The author(s) disclosed receipt of the following financial support for the research and/or authorship of this article: grants from the Eunice Kennedy Shriver National Institute of Child Health and Human Development (HD27860, HD36801, HD27917, HD21414, HD27861, HD27869, HD27905, HD34208, HD34116, HD21410, HD27915, HD34136, HD34210, HD34122, HD40500, HD40544, HD34116, HD40560, HD40512) and its contents do not necessarily represent the official view of NICHD or NIH.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no conflicts of interest with respect to the authorship and/or publication of this article.
Author’s Notes
Sources of the work: University of Utah Department of Obstetrics and Gynecology, Eunice Kennedy Shriver National Institute of Child Health & Human Development Maternal-Fetal Medicine Unit Network. This article was presented as a poster at the 2009 Society for Gynecologic Investigation in Glasgow, Scotland.
References
- 1.Meis PJ, Klebanoff M, Thom E, et al. Prevention of recurrent preterm delivery by 17 alpha-hydroxyprogesterone caproate. N Engl J Med. 2003;349(13):2379–2385. doi: 10.1056/NEJMoa035140. [DOI] [PubMed] [Google Scholar]
- 2.Facchinetti F, Paganelli S, Comitini G, Dante G, Volpe A. Cervical length changes during preterm cervical ripening: effects of 17-alpha-hydroxyprogesterone caproate. Am J Obstet Gynecol. 2007;196(5):453.e1–453.e4. doi: 10.1016/j.ajog.2006.09.009. [DOI] [PubMed] [Google Scholar]
- 3.Fonseca EB, Celik E, Parra M, Singh M, Nicolaides KH, Fetal Medicine Foundation Second Trimester Screening Group Progesterone and the risk of preterm birth among women with a short cervix. N Engl J Med. 2007;357(5):462–469. doi: 10.1056/NEJMoa067815. [DOI] [PubMed] [Google Scholar]
- 4.Martin JA, Hamilton BE, Sutton PD, Ventura SJ, Menacker F, Munson ML. Births: final data for 2003. Natl Vital Stat Rep. 2005;54(2):1–116. [PubMed] [Google Scholar]
- 5.Porter TF, Fraser AM, Hunter CY, Ward RH, Varner MW. The risk of preterm birth across generations. Obstet Gynecol. 1997;90(1):63–67. doi: 10.1016/S0029-7844(97)00215-9. [DOI] [PubMed] [Google Scholar]
- 6.Ward K, Argyle V, Meade M, Nelson L. The heritability of preterm delivery. Obstet Gynecol. 2005;106(6):1235–1239. doi: 10.1097/01.AOG.0000189091.35982.85. [DOI] [PubMed] [Google Scholar]
- 7.Esplin MS, O’Brien E, Fraser A, et al. Estimating recurrence of spontaneous preterm delivery. Obstet Gynecol. 2008;112(3):516–523. doi: 10.1097/AOG.0b013e318184181a. [DOI] [PubMed] [Google Scholar]
- 8.Boyd HA, Poulsen G, Wohlfahrt J, Murray JC, Feenstra B, Melbye M. Maternal contributions to preterm delivery. Am J Epidemiol. 2009;170(11):1358–1364. doi: 10.1093/aje/kwp324. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Mills AA, Yonish B, Feng L, Schomberg DW, Heine RP, Murtha AP. Characterization of progesterone receptor isoform expression in fetal membranes. Am J Obstet Gynecol. 2006;195(4):998–1003. doi: 10.1016/j.ajog.2006.06.040. [DOI] [PubMed] [Google Scholar]
- 10.Merlino AA, Welsh TN, Tan H, et al. Nuclear progesterone receptors in the human pregnancy myometrium: evidence that parturition involves functional progesterone withdrawal mediated by increased expression of progesterone receptor-A. J Clin Endocrinol Metab. 2007;92(5):1927–1933. doi: 10.1210/jc.2007-0077. [DOI] [PubMed] [Google Scholar]
- 11.Attardi BJ, Zeleznik A, Simhan H, Chiao JP, Mattison DR, Caritis SN. Comparison of progesterone and glucocorticoid receptor binding and stimulation of gene expression by progesterone, 17-alpha hydroxyprogesterone caproate, and related progestins. Am J Obstet Gynecol. 2007;197(6):599.e1–599.e7. doi: 10.1016/j.ajog.2007.05.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Godeau JF, Schorderet-Slatkine S, Hubert P, Baulieu EE. Induction of maturation in Xenopus laevis oocytes by a steroid linked to a polymer. Proc Natl Acad Sci U S A. 1978;75(5):2353–2357. doi: 10.1073/pnas.75.5.2353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Bielefeldt K, Waite L, Abooud F, Conklin J. Nongenomic effects of progesterone on human intestinal smooth muscle cells. Am J Physiol. 1996;271(2 pt 1):G370–G376. doi: 10.1152/ajpgi.1996.271.2.G370. [DOI] [PubMed] [Google Scholar]
- 14.Fomin V, Cox B, Word R. Effect of progesterone on intracellular Ca2+ homeostasis on human myometrial smooth muscle cells. Am J Physiol. 1999;276(2 pt 1):C379–C385. doi: 10.1152/ajpcell.1999.276.2.C379. [DOI] [PubMed] [Google Scholar]
- 15.Saner KJ, Welter BH, Zhang F, et al. Cloning and expression of a novel, truncated, progesterone receptor. Mol Cell Endocrinol. 2003;200(1-2):155–163. doi: 10.1016/s0303-7207(02)00380-5. [DOI] [PubMed] [Google Scholar]
- 16.Garde RV, Saner CN, Price TM. Discovery of a truncated progesterone receptor (PR-M) in the mitochondria; Plenary Oral Presentation, Wyeth Presidential Award, Society for Gynecological Investigation at the 53rd Annual Meeting; Toronto, Canada. March 2006. [Google Scholar]
- 17.Gavrilova-Jordan LP, Schomberg D, Feng L, Jayes FL, Murtha A, Price TM. Expression of a truncated progesterone receptor isoform (PR-M) in pregnant and laboring myometrium. Reprod Sci. 2009;16(3 suppl):114A. [Google Scholar]
- 18.Kline LW, Karpinski E. Progesterone inhibits gallbladder motility through multiple signaling pathways. Steroids. 2005;70(9):673–679. doi: 10.1016/j.steroids.2005.03.011. [DOI] [PubMed] [Google Scholar]
- 19.Hodges YK, Richer JK, Horwitz KB, Horwitz LD. Variant estrogen and progesterone receptor messages in human vascular smooth muscle. Circulation. 1999;99(20):2688–2693. doi: 10.1161/01.cir.99.20.2688. [DOI] [PubMed] [Google Scholar]
- 20.Morishita S. Prompt effect of progesterone on the adrenergic response of smooth muscles. Jpn J Pharmacol. 1986;42(2):289–296. doi: 10.1254/jjp.42.289. [DOI] [PubMed] [Google Scholar]
- 21.Mesiano S. Myometrial progesterone responsiveness. Semin Reprod Med. 2007;25(1):5–13. doi: 10.1055/s-2006-956771. [DOI] [PubMed] [Google Scholar]
- 22.Demonacos C, Tsawdaroglou NC, Djordjevic-Markovic, et al. Import of the glucocorticoid receptor into rat liver mitochondria in vivo and in vitro. J Steroid Biochem Mol Biol. 1993;46(3):401–413. doi: 10.1016/0960-0760(93)90231-k. [DOI] [PubMed] [Google Scholar]
- 23.Psarra AM, Solakidi S, Trougakos IP, Margaritis LH, Spyrou G, Sekeris CE. Glucocorticoid receptor isoforms in human hepatocarcinoma HepG2 and SaOS-2 osteosarcoma cells: presence of glucocorticoid receptor beta in nucleoli. Int J Biochem Cell Biol. 2005;37:2544–2558. doi: 10.1016/j.biocel.2005.06.015. [DOI] [PubMed] [Google Scholar]
- 24.Monje P, Boland R. Subcellular distribution of native estrogen receptor alpha and beta isoforms in rabbit uterus and ovary. J Cell Biochem. 2001;82(3):467–479. doi: 10.1002/jcb.1182. [DOI] [PubMed] [Google Scholar]
- 25.Chen JQ, Delannoy M, Cooke C, Yager JD. Mitochondrial localization of ERα and ERβ in human MCF7 cells. Am J Physiol Endocrinol Metab. 2004;286(6):E1011–E1022. doi: 10.1152/ajpendo.00508.2003. [DOI] [PubMed] [Google Scholar]
- 26.Gavrilova-Jordan LP, Price TM. Actions of steroids in mitochondria. Semin Reprod Med. 2007;25(3):154–164. doi: 10.1055/s-2007-973428. [DOI] [PubMed] [Google Scholar]
- 27.Gavrilova-Jordan LP, Zhang L, Schomberg D, Jayes FL, Feng L, Price TM. Expression of a truncated progesterone receptor (PRM) correlates with mitochondrial content in human tissues. Reprod Sci. 2009;16(3 suppl):189A. [Google Scholar]