

Abstract
Exclusion allows the detection of hidden food when confronted with the choice between an empty and a potentially baited food location. However, exclusion may be based on avoidance of the empty location without drawing inferences about the presence of the food in the baited location. So far, such inferences have been demonstrated in the great apes only: after seeing an experimenter eating one of two food types, which both had been hidden previously in two boxes, the apes were able to choose the box that still contained the other food type. African grey parrots are capable of exclusion, and we here assessed if they are capable of inference by exclusion. In our task, two different but equally preferred food items were hidden in full view of the birds under two opaque cups. Then, an experimenter secretly removed one food type and showed it to the bird. Similarly to the apes, one out of seven parrots significantly preferred the baited cup; control conditions rule out that its choice was based on associative learning or the use of olfactory cues. Thus, we conclude that—like the apes—some grey parrots are able to infer the location of a hidden food reward.
Keywords: African grey parrots, inference by exclusion, reasoning
1. Introduction
Choice by exclusion is defined as ‘the ability to base the choice of a target on the rejection of potential alternatives’ [1]; for instance, apes [2], monkeys [3–6], dogs [7–9] and ravens [10] selected the baited box in a two-choice procedure with two opaque boxes after they received information about the empty box only. However, this task can be solved by an avoidance of the empty box rather than an inference about the reward in the baited box. Only in a chimpanzee study by Premack & Premack [11], was it possible to rule out avoidance of the incorrect alternative. First, the experimenter hid an apple and a banana in two different boxes in full view of the ape; subsequently, the chimpanzee was allowed to witness the experimenter eating one of the fruits, which had been removed secretly, and was thereupon allowed to choose one box. Only one out of five individuals was reliably able to choose the still-baited box. This task was then replicated with all great ape species by Call [12], who found a similar degree of inter-individual differences. Thus, in general, great apes are able to draw inferences by exclusion, but it seems to be a demanding, not easy-to-solve task.
Nothing is known about ‘true’ inferential reasoning in such a food-finding task beyond the great apes, as it has never been replicated with a non-ape species. Recent data indicate that African grey parrots (Psittacus erithacus), who are well-known for their advanced cognitive abilities [13], are able to choose by exclusion in the visible as well as in the acoustic domain [14]. Therefore, we here tested whether African grey parrots are capable of ape-like true inferential reasoning, which will be important for our understanding of the cognitive evolution of mammals and birds.
2. Material and methods
The subjects were seven grey parrots (four males), housed in a parrot rescue centre in Vienna, Austria. The birds were of different and often not fully known history and were between approximately 7 and 25 years old. They were housed together with other not tested individuals in an indoor–outdoor aviary (3 × 5 m each) that was enriched with several perches and toys. Birds were fed with fruits in the morning and seeds in the evening. Testing was conducted between 9.00 and 13.00 h and each bird was tested in visual isolation (testing compartment: 120 × 70 cm and 210 cm height) on a fixed platform (60 × 35 cm at a height of 130 cm). The experimental set-up consisted of two identical opaque cups (9 cm height and 8 cm diameter) on a wooden, movable platform (40 × 23 cm) hanging from the ceiling at the same height but 15 cm away from the fixed platform (see electronic supplementary material, figure S1). Therefore, while the birds were positioned on the fixed platform, the experimenter could present the set-up out of their reach on the movable platform and could push it within their reach to allow them to choose.
(a). Pre-tests
(i). Choice training
This test was introduced to ensure that the birds were aware that they would have to make a choice. One piece of food (e.g. a seed or a piece of walnut, depending on known preferences of the birds) was placed on the movable platform and covered with an opaque cup. Simultaneously, the second cup was placed on the same platform at a distance of approximately 20 cm. Then, the platform was pushed forward to allow the bird to make a choice by touching a cup with its beak and retrieve a food item if correct. One session consisted of 10 trials; the training criterion to reach the next step was set to be correct on at least eight trials in two consecutive sessions.
(ii). Preference test
This test was introduced to ensure that the test performance would not be influenced by food preferences. We visibly placed two different food items (the same as in the training) simultaneously on the movable platform and covered them with the two cups. Then, the bird was allowed to choose and retrieve one food item. The position of the food types was semi-randomized with the stipulation that the same food type was not placed on the same side for more than three consecutive trials. Two sessions were conducted, each consisting of 10 trials. Birds were advanced to the test if they did not show a preference for one food type, i.e. did not select one item more than 13 times over the two sessions.
(b). Tests
In the test phase, these two equally preferred food items were then placed simultaneously on the movable platform and covered with the two cups.
(i). Visible condition
Standing equidistantly between the cups and in full view of the subjects, the experimenter looked straight ahead, lifted the left cup, manipulated the food, returned the cup, lifted the right cup, manipulated the food and returned the cup. One manipulation consisted of taking the food, showing it to the bird and putting it back on the board. The other manipulation consisted of taking the food, showing it to the bird and removing it. The order of manipulations was randomized, but no food type was removed more than three times in a row and no side remained baited in more than three consecutive trials. Afterwards the bird was allowed to make a choice and received the reward if correct or saw the empty cup if not.
(ii). Invisible condition
This condition was identical to the visible condition, with the exception that after covering the rewards with the cups an opaque barrier was placed between the bird and the cups and the two manipulations were performed behind this barrier. Then, the barrier was removed and the experimenter showed the item she had taken away and put it into her pocket.
The birds first received either the visible or the invisible condition. If the birds showed a significant preference for the baited cup in the invisible condition, two control conditions were conducted.
(iii). Olfaction control
Behind the barrier, the experimenter hid one piece of food (per trial one of the two food types) underneath one of the cups. Without any information about the food location, the bird was then allowed to choose one cup.
(iv). Association control
To test if the birds used the associative rule ‘always choose the cup with the food type not having been shown’, we visibly placed the two food items on the board, covered them with the cups and then took—in full view of the subject—one piece of food (of the same type as one of the food items being hidden) out of the experimenters' pocket, showed it to the bird and put it back in the pocket. Afterwards the bird was allowed to choose.
In all conditions, the birds received three sessions with 10 trials per session for a total of 30 trials in each condition. Birds received only one session per day. If a bird left the testing area during testing, the session was continued on the following day (see the electronic supplementary material).
3. Results
The parrots received 22.9 ± 4.9 (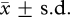 , range 20–30) trials of choice training. In the preference test, no bird chose one food type more than 12 times (binomial-tests: p ≥ 0.503). In the test, the birds significantly preferred the baited cup in the visible (one-sample t-test: n = 7, d.f. = 6, t = 7.772, p < 0.001), but not in the invisible condition (t = 1.430, p = 0.203; figure 1). On an individual level, all birds selected the baited cup in the majority of the trials in the visible condition and three birds were significantly above chance level (p ≤ 0.043; table 1 and the electronic supplementary material, results). In the invisible condition, one out of seven birds was significantly above chance (binomial-test: p = 0.005; all others: p ≥ 0.200; table 1). The successful bird (a female of approx. 13 years) chose at chance level in the olfaction and in the association control (binomial-test: p = 0.856 in both cases). When comparing her performance in the first and last 15 trials, we found no difference in the visible condition (McNemar-test: n = 15, p > 0.999) or in the olfaction (p = 0.625) and association control (p = 0.508), but a significant improvement in the invisible condition (p = 0.016).
, range 20–30) trials of choice training. In the preference test, no bird chose one food type more than 12 times (binomial-tests: p ≥ 0.503). In the test, the birds significantly preferred the baited cup in the visible (one-sample t-test: n = 7, d.f. = 6, t = 7.772, p < 0.001), but not in the invisible condition (t = 1.430, p = 0.203; figure 1). On an individual level, all birds selected the baited cup in the majority of the trials in the visible condition and three birds were significantly above chance level (p ≤ 0.043; table 1 and the electronic supplementary material, results). In the invisible condition, one out of seven birds was significantly above chance (binomial-test: p = 0.005; all others: p ≥ 0.200; table 1). The successful bird (a female of approx. 13 years) chose at chance level in the olfaction and in the association control (binomial-test: p = 0.856 in both cases). When comparing her performance in the first and last 15 trials, we found no difference in the visible condition (McNemar-test: n = 15, p > 0.999) or in the olfaction (p = 0.625) and association control (p = 0.508), but a significant improvement in the invisible condition (p = 0.016).
Figure 1.

Percentage of correct choices in the visible and invisible condition. The horizontal line indicates the chance level. The box plot shows median and quartiles. The whiskers represent the 10% and 90% range, dots indicate the 5% and 95% range.
Table 1.
Individual performances given as percentage of correct choices. Significant performances (according to a Binomial-test) are highlighted in bold (f = female; m = male).
| individual | sex | visible | invisible | olfaction | association |
|---|---|---|---|---|---|
| Awisa | f | 70.0 | 76.7 | 46.7 | 56.7 |
| Cocohan | m | 63.3 | 50.0 | — | — |
| Kasi | f | 63.3 | 60.0 | — | — |
| Leo | m | 60.0 | 53.3 | — | — |
| Maja | f | 76.7 | 46.7 | — | — |
| Moritz | m | 73.3 | 63.3 | — | — |
| Rocky | m | 66.7 | 43.3 | — | — |
4. Discussion
Our study shows that African grey parrots can use partial information about the removed food item not only to choose by exclusion based on avoidance, but that they are capable of true inferential reasoning. However, whereas several subjects solved the task when they could see the experimenter's actions, only one individual was able to solve the task when it had to infer the outcome of the experimenter's actions; still, these findings are in line with the inter-individual differences found in the great apes [2,11]. The birds were clearly motivated to obtain the reward, even though side biases occurred (see the electronic supplementary material). As they were highly attentive and never refrained from making a choice, we are confident that the success of only a single bird in the invisible condition is not owing to motivational issues; rather, it supports the assumption that these inference tasks are not trivial but cognitively demanding. Again, this result demonstrates substantial inter-individual differences in grey parrot cognitive performance [15] and that the level of performance depends on subtle differences between tasks [13,16].
Importantly, the successful bird significantly improved her performance only within the invisible condition. This improvement does not necessarily point to associative learning, as the bird performed at chance level in the association control. We suggest that the improvement is owing to the fact that the bird did not comprehend the relevance of the food presentation at the beginning of the test. This is plausible in particular because this bird was first tested in the invisible condition. Probably, the bird needed a few trials to become acquainted with the test. Thus, we suggest that this bird's performance was not based on learning instead of reasoning, but rather that the bird learned to reason about the experimenter's action to solve the task.
Acknowledgements
The experiments comply with Austrian law.
The authors thank the FWF (P20528-B17) for funding. Special thanks to Nadja Ziegler and the ARGE Papageienschutz for granting access to their grey parrots. Not the least, we thank all the animal keepers and especially Judith Schmidt.
References
- 1.Schloegl C. 2011. What you see is what you get-reloaded: can jackdaws (Corvus monedula) find hidden food through exclusion? J. Comp. Psychol. 125, 162–174 10.1037/a0023045 (doi:10.1037/a0023045) [DOI] [PubMed] [Google Scholar]
- 2.Call J. 2004. Inferences about the location of food in the great apes (Pan paniscus, Pan troglodytes, Gorilla gorilla, and Pongo pygmaeus). J. Comp. Psychol. 118, 232–241 10.1037/0735-7036.118.2.232 (doi:10.1037/0735-7036.118.2.232) [DOI] [PubMed] [Google Scholar]
- 3.Hill A., Collier-Baker E., Suddendorf T. 2011. Inferential reasoning by exclusion in great apes, lesser apes, and spider monkeys. J. Comp. Psychol. 125, 91–103 10.1037/a0020867 (doi:10.1037/a0020867) [DOI] [PubMed] [Google Scholar]
- 4.Sabbatini G., Visalberghi E. 2008. Inferences about the location of food in Capuchin monkeys (Cebus apella) in two sensory modalities. Anim. Behav. 122, 156–166 10.1037/0735-7036.122.2.156 (doi:10.1037/0735-7036.122.2.156) [DOI] [PubMed] [Google Scholar]
- 5.Schmitt V., Fischer J. 2009. Inferential reasoning and modality dependent discrimination learning in olive baboons (Papio hamadryas anubis). J. Comp. Psychol. 123, 316–325 10.1037/a0016218 (doi:10.1037/a0016218) [DOI] [PubMed] [Google Scholar]
- 6.Petit O., Call J., Thierry B. 2005. Inference about food location in Tonkean macaques. Primate Rep. 72, 76 [Google Scholar]
- 7.Aust U., Range F., Steurer M., Huber L. 2008. Inferential reasoning by exclusion in pigeons, dogs and humans. Anim. Cogn. 11, 587–597 10.1007/s10071-008-0149-0 (doi:10.1007/s10071-008-0149-0) [DOI] [PubMed] [Google Scholar]
- 8.Bräuer J., Kaminski J., Riedel J., Call J., Tomasello M. 2006. Making inference about the location of hidden food: social dog, causal ape. J. Comp. Psychol. 120, 38–47 10.1037/0735-7036.120.1.38 (doi:10.1037/0735-7036.120.1.38) [DOI] [PubMed] [Google Scholar]
- 9.Erdöhegyi A., Topal J., Viranyi Z., Miklosi A. 2007. Dog-logic: inferential reasoning in a two-way choice task and its restricted use. Anim. Behav. 74, 725–737 10.1016/j.anbehav.2007.03.004 (doi:10.1016/j.anbehav.2007.03.004) [DOI] [Google Scholar]
- 10.Schloegl C., Dierks A., Gajdon G. K., Huber L., Kotrschal K., Bugnyar T. 2009. What you see is what you get? Exclusion performances in ravens and keas. PLoS ONE 4, 1–12 10.1371/journal.pone.0006368 (doi:10.1371/journal.pone.0006368) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Premack D., Premack A. J. 1994. Levels of causal understanding in chimpanzees and children. Cognition 50, 347–362 10.1016/0010-0277(94)90035-3 (doi:10.1016/0010-0277(94)90035-3) [DOI] [PubMed] [Google Scholar]
- 12.Call J. 2006. Inference by exclusion in the great apes: the effect of age and species. Anim. Cogn. 9, 393–403 10.1007/s10071-006-0037-4 (doi:10.1007/s10071-006-0037-4) [DOI] [PubMed] [Google Scholar]
- 13.Pepperberg I. M. 1999. The alex studies: cognitive and communicative abilities in grey parrots. Cambridge, MA: Harvard University Press [Google Scholar]
- 14.Schmidt J., Kotrschal K., Schloegl C. In preparation Exclusion abilities in African grey parrots (Psittacus erithacus). [Google Scholar]
- 15.Pepperberg I. M. 2007. Individual differences in grey parrots (Psittacus erithacus): effects of training. J. Ornithol. 148, 161–168 10.1007/s10336-007-0162-0 (doi:10.1007/s10336-007-0162-0) [DOI] [Google Scholar]
- 16.Giret N., Peron F., Lindova J., Tichotova L., Nagle L., Kreutzer M., Tymr F., Bovet D. 2010. Referential learning of French and Czech labels in African grey parrots (Psittacus erithacus): different methods yield contrasting results. Behav. Process. 85, 90–98 10.1016/j.beproc.2010.06.010 (doi:10.1016/j.beproc.2010.06.010) [DOI] [PubMed] [Google Scholar]