

Abstract
A previous study reported preliminary results of enhanced processing of simple visual information in the form of faster reaction times, in female fragile X premutation carriers (fXPCs). In this study, we assessed manual and oral motor reaction times in 30 female fXPCs and 20 neurotypical (NT) controls. Participants completed two versions of the reaction time task; one version required a manual motor response and the other version required an oral motor response. Results revealed that the female fXPCs displayed faster reaction times for both manual and oral motor responses relative to NT controls. Molecular measures including CGG repeat length, FMR1 mRNA levels, and age were not associated with performance in either group. Given previously reported age and CGG repeat modulated performance on a magnitude comparison task in this same group of premutation carriers, results from the current study seem to suggest that female fXPCs may have spared basic psychomotor functionality.
Keywords: Psychomotor performance, Anxiety, Cortiospinal tract, Corticopontine tract, Motor cortex, X-linked genetic disease
INTRODUCTION
The FMR1 gene is polymorphic for the length of a CGG trinucleotide repeat in the 5′ untranslated region of the X chromosome. In the general population, there are <45 CGG repeats. Fragile X syndrome (FXS) is defined by the full mutation (i.e., >200 CGG repeats) and silenced transcription of the FMR1 gene due to the hypermethylation of the promoter. In the fragile X premutation there are 55–200 CGG repeats, which results in a three- to eight-fold increase in FMR1 mRNA levels in leukocytes and decreased FMR1 protein (FMRP) levels due to translational inefficiency of the mutant FMR1 mRNA (Garcia-Arocena & Hagerman, 2010). Female fragile X premutation carriers (fXPCs) differ from male fXPCs in that female fXPCs have a second, unmutated copy of the FMR1 gene that is expressed randomly in 50% of the cells. It has been estimated that 1 in 260 to 813 males and 1 in 113 to 259 females are fXPCs (Hagerman, 2008). Of note, some fXPCs develop a late onset (>50 years old) neurodegenerative disorder: fragile X-associated tremor/ataxia syndrome (FXTAS; Bourgeois et al., 2009).
Along with the recent development of genetic profiles for fXPCs, psychiatric features have begun to emerge in female fXPCs compared to female controls, such as higher rates of anxiety, depression, and attention problems (Bourgeois et al., 2011). However, whether asymptomatic (i.e., non-FXTAS) young, adult female fXPCs show cognitive impairments remains controversial. There have been a few reports suggesting that fXPCs show cognitive impairments (Cornish et al., 2008; Goodrich-Hunsaker et al., 2011). Other studies report that there is no evidence for neurocognitive implications in fXPCs younger than 50 years old (Hunter et al., 2008). To add to this controversy, there is evidence that young female fXPCs may actually outperform neurotypical control groups (NTs) in some domains. Specifically, young female fXPCs were shown to have faster reaction times on a battery of attention tasks compared to young NT adults who were not fragile X carriers (Steyaert, Borghgraef, & Fryns, 1994). However, these results were preliminary, and no other studies have reported similar findings.
In the current study, we measured both simple manual and oral motor speeded responses to visual stimuli. We also wanted to determine whether performance was differentially modulated in fXPCs and NTs across CGG repeat length, FMR1 mRNA levels, and age. Psychologists have described two major kinds of reaction time experiments: simple and choice reaction time experiments (Welford, 1980). Simple reaction time tasks assess the speed at which a participant can respond to a single external cue. Choice reaction time tasks involve multiple stimuli in which the participant is required to give a specific response that corresponds to each particular stimulus. Choice reaction time tasks require participants to make a decision, whereas simple reaction time tasks do not. A single-stimulus simple reaction time task, such as that used in the current study, is considered a fairly pure index of psychomotor speed. Because reaction times, along with error rates, are commonly used as outcome measures in cognitively demanding tasks, it is important to assess each participant’s baseline psychomotor speed when cognitive demands are minimized to account for the specific effects of cognitive demands upon performance.
METHODS
Participants
Participants were 50 females aged 21 to 42, including 20 neurotypical (NT) controls and 30 fragile X premutation carriers (fXPCs). The mean age (± SD) for fXPCs was 31.50 ±7.17 years and for NT controls was 34.08 ±4.57 years. The two groups did not differ in age, t =−1.56, p =.13, or Full-Scale IQ, t =−1.38, p =.18 (see Table 1). Participants were recruited through the NeuroTherapeutics Research Institute (NTRI) at the Medical Investigation of Neurodevelopmental Disorders (M.I.N.D.) Institute at the University of California, Davis Medical Center. Participants with any of the following were not included in our study: acute medical condition such as renal, liver, cardiac, or other disease that may be associated with brain atrophy or dysfunction; current or past history of major DSM-IV Axis I psychiatric disorder; history of head trauma; toxic encephalopathy, encephalitis, or bacterial meningitis; history of alcoholism or drug problem; use of current medication that affect cerebral blood flow (e.g., beta blockers); and presence of metal in the body. We also requested that all participants be off anti-depressant or anti-anxiety medication for 2 weeks before participation. This study was approved by the Institutional Review Board and conformed to institutional and federal guidelines for the protection of human participants. Written informed consent was obtained before participation from all participants.
Table 1.
Participant descriptive statistics and FMR1 measures
| Neurotypical control
|
Fragile X premutation carrier
|
t | p value | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Mean | SD | Range | n | Mean | SD | Range | n | |||
| Age | 31.50 | 7.17 | 21–41 | 20 | 34.08 | 4.57 | 23–42 | 30 | −1.56 | .13 |
| Full-Scale IQ | 111.6 | 12.32 | 89–129 | 15 | 117.14 | 11.78 | 101–144 | 22 | −1.38 | .18 |
| CGG repeat size | 30.07 | 1.22 | 28–32 | 15 | 96.17 | 19.85 | 67–143 | 30 | −12.81 | <.0001 |
| FMR1 mRNA level | 1.61 | 0.27 | 1.25–1.98 | 8 | 2.38 | 0.52 | 1.55–3.46 | 24 | −3.98 | <.0001 |
| Activation ratio | n/a | n/a | n/a | 0 | 55% | 16% | 23–87% | 30 | n/a | n/a |
Psychological Assessment
Full-Scale IQ (FSIQ) was measured using either the Wechsler Adult Intelligence Scale, third edition (WASI-III; Wechsler, 1997) or the Wechsler Abbreviated Scale of Intelligence (WASI; Wechsler, 1999). FSIQ data were available for 15/20 NT controls and 22/30 fXPCs.
Molecular Analysis
As previously described (Tassone, Pan, Amiri, Taylor, & Hagerman, 2008), genomic DNA was isolated from peripheral blood leucocytes using standard methods (Puregene Kit; Gentra Inc., Valencia, CA). Repeat size was determined using Southern blot and PCR amplification of genomic DNA. The activation ratio (AR), indicating the percent of cells that carry the normal allele on the active X chromosome, was calculated by Southern blot. All quantifications of FMR1 mRNA were performed using a 7900 Sequence detector (PE Biosystems).
Simple Reaction Time
The simple reaction time task was presented on a 2 GHz Intel Core 2 Duo HP Compaq dc7700 Small Form Factor PC equipped with 1 GB of RAM running SuperLab version 4.0.7b (Cedrus Corporation, San Pedro, CA). The stimuli included a drawing of a house that was 19 cm in height and 7.9 cm wide with an entrance that was 5.4 cm high and 3.2 cm wide on the monitor 60 cm from the participant. On each trial participants were asked to indicate as quickly as possible, by pressing a single button for the manual motor response version of the task or by speaking “GO” into a microphone for the oral motor version of the task, whenever a picture of a friendly alien appeared at the right or left side of the entrance. For the manual motor response version, participants used their dominant hand. The alien figure was 5.4 cm tall and 1.8 cm in the widest extent. This image remained on the screen until the participant responded. The version order was randomized across participants. Each version of the task consisted of 60 consecutive trials. Delays between trials were set to one of three intervals (400, 800, or 1200 ms), which were presented in random order to minimize anticipatory responses. Response time was recorded as the primary dependent variable.
Data Analysis
Data measured either manual or oral motor reaction time. Within each version, trials with reaction times greater than or less than 3 times the interquartile range or less than or equal to 150 ms (anticipatory responses) were excluded from the analyses. Results were calculated as the median of reaction times across each condition delay. Repeated-measures analysis of variance (ANOVA) with delay (400, 800, 1200 ms) as the within-participant factor and group (NT and fXPC) as the between-participant factor were performed. Correlations between simple reaction time and age (for both groups) and molecular variables (fXPCs only) were computed within groups. For the correlation analyses, simple reaction times were calculated as the median of reaction across all trials and condition delays.
Degrees of freedom were adjusted using the Welch procedure for one-way ANOVAs when the equality of variance assumption was violated. For repeated-measures ANOVAs, Greenhouse-Geisser corrections were used to correct for violations of the sphericity assumption. The reaction times were log-transformed to better meet the assumptions of the model. Analyses were conducted using SPSS and a p value <.05 was considered statistically significant.
RESULTS
Molecular Analyses
Molecular data were available for 15/20 NT controls and 30/30 fXPCs.
Descriptive statistics of CGG repeat size, FMR1 mRNA, and activation ratios are reported in Table 1. Within the fXPCs, as expected, FMR1 mRNA level was positively associated with CGG repeat size, Pearson’s r =.42, p (one-tailed) =.02. The partial correlation between CGG and FMR1 mRNA after accounting for the activation ratio in the fXPCs was much stronger, Pearson’s r =.55, p (one-tailed) =.003. In the female fXPCs, we found no significant correlations between CGG repeat size or FMR1 mRNA with age.
Simple Reaction Time
The simple reaction time tasks assessed participants’ abilities to make speeded responses while eliminating all cognitive demands to perform accurately. Responses from two female NTs and one female fXPC were identified as outliers (i.e., greater than or less than 3 times the interquartile range for each group) on the oral version of the task, so they were removed from those analyses only. A repeated-measures ANOVA showed that female fXPCs were faster to respond on the manual (F(1,48) =10.00; p =.003) and oral (F(1,45) = 6.76; p =.01) motor reaction time task than female NTs (see Figure 1). For both task versions, reaction times decreased as the delay between trials increased (p <.0001), but did not differ between the groups, p >.28. This indicates that both groups of participants could anticipate upcoming trials. Overall median reaction times across all trials for each task version within each group were used to assess the association between performance and age (both groups) and molecular variables (in the female fXPCs). There were no significant associations.
Fig. 1.
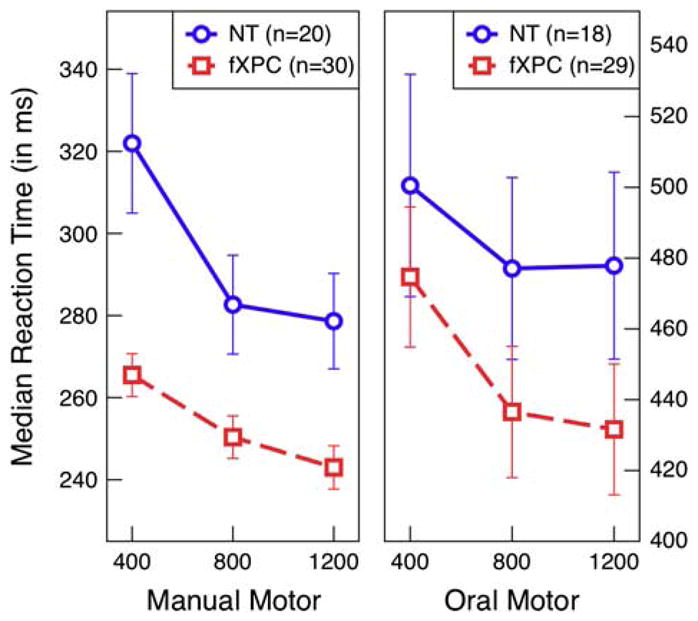
Oral motor reaction times ranged from 248 to 782 ms (M =461.24, SEM =7.92) and were overall slower than manual motor reaction times, which ranged from 203 to 515 ms (M =268.14, SEM =3.42). Group analyses of manual motor and oral reaction time show that female fXPCs, as a group, responded faster than female NT controls, p =.003 and p =.01, respectively. For both task versions, reaction times decreased as the delay between trials were longer, p <.0001, but did not differ between the groups, p >.28. Error bars represent standard error of the mean.
DISCUSSION
As a group, the young adult female neurotypical controls (NTs) had comparable manual motor and oral motor reaction times to age- and gender-matched controls reported in previously published studies (Der & Deary, 2006; Steyaert et al., 1994). With the reaction times of the NTs in the present study replicating earlier reports, the finding that the young, asymptomatic (i.e., non-FXTAS) female fragile X premutation carriers (fXPCs) performed faster than the NTs on both versions of the simple reaction time task is unexpected. The results from the current study are consistent with a previous study that also reported faster simple reaction times in female fXPCs relative to NTs (Steyaert et al., 1994). Steyaert et al. (1994) assessed various aspects of attention as measured by the de Sonneville Visual Attention Tasks (SVAT). The SVAT consists of eight different tasks, including a simple manual motor reaction time task, the Finger Motor Response Exercise (FMSE). The female fXPCs had overall faster manual reaction times compared to NTs (Steyaert et al., 1994). The results of the current study replicate these findings and extend the results to include oral motor reaction times as well. These findings taken together demonstrate that female fXPCs have enhanced basic psychomotor speed relative to NT controls.
In remains unclear why reaction times would be faster in female fXPCs. One hypothesis as to why basic psychomotor speed is enhanced in female fXPCs is that there may be a neural reorganization that allows for faster processing on the simple reaction time task, but results in cognitive impairments in other domains. Basic psychomotor speed as measured by simple reaction time tasks depends largely upon the corticospinal tract (pyramidal system) with contributions coming from the supplementary motor area, premotor cortex, somatosensory cortex, cerebellum, parietal lobe, and cingulate gyrus (Kolb & Whishaw, 2009). Several of these regions are effected in female fXPCs with FXTAS, such as reduced cerebellar volume (Adams et al., 2007) and parietal gyral atrophy (Hagerman et al., 2004). Similar findings have emerged in female fXPCs without FXTAS. We have detected widespread white matter changes throughout the corticospinal and corticopontine tracts, implicating the motor system, in this same group of female fXPCs (manuscript in preparation). Moreover, these changes also appear to be impacted by age and CGG repeat length. Therefore, these behavioral differences in the current study may be the result of subtle underlying neural changes occurring preclinically, long before symptoms of FXTAS develop. At present, the time course of this reorganization is unknown, although a preliminary study comparing fXPC children to NTs found no group differences on measures of intellectual functioning, academic achievement, or visuomotor integration (Myers, Mazzocco, Maddalena, & Reiss, 2001). Whether these features develop at some critical point in response to environmental factors, or if they develop along a continuous trajectory of development that is increasingly different from NTs remains unclear.
Another hypothesis is that anxiety levels may be modulating basic psychomotor speed. There is a high prevalence of anxiety reported in female fXPCs (Bourgeois et al., 2011). Nettelbeck (1973) reported that anxious personality types have faster reaction times. The heightened anxiety levels reported in female fXPCs may stem from being carriers of the fragile X premutation allele, which has many lifelong implications. When female fXPCs transmit the premutation allele to their children, the CGG repeat length almost always expands and greater than 50% of the time will expand into the full mutation range (>200 CGG repeats; Nolin et al., 2003). The stress of raising a child with FXS could certainly exacerbate elevated anxiety levels in female fXPCs (Bourgeois et al., 2011). Additionally, many of the female fXPCs are daughters of men with the late-onset (>50 years old) neurodegenerative disorder, fragile X-associated tremor/ataxia syndrome (FXTAS), which they may also eventually develop. Individuals who have a parent with FXTAS or a child with FXS have concerns about these, which can predispose them to have heightened anxiety. Given these life circumstances, female fXPCs may have a heightened anxiety that is contributing to their increased basic psychomotor speed. In the current study, anxiety levels were not assessed, so unfortunately no direct correlation can be drawn between anxiety levels and psychomotor speed. Related to anxiety, increased arousal may facilitate reaction times as well. Studies are currently underway to quantify anxiety and arousal (e.g., pupillometry) in these participants.
In another study, Panayiotou and Vrana (2004) reported that self-awareness and anxiety about one’s performance could also enhance performance on simple tasks in which the participant can easily be successful. However, if the task requires a large speed/accuracy tradeoff, then anxiety about one’s performance does not enhance performance. Female fXPCs, in general, have several motivating reasons to perform well. First, their participation and the outcomes from research studies will hopefully provide a direct benefit to themselves and their family. Second, although female fXPCs are less affected than their male counterparts, research suggests that they may have subtle cognitive impairments (Cornish et al., 2008; Goodrich-Hunsaker et al., 2011), and thus female fXPCs may be self-conscious and as a result perform at the absolute best of their abilities in compensation, even during simple tasks. Therefore, female fXPCs would have enhanced performance on relatively easy tasks like the simple reaction time task, but poorer performance on more cognitively demanding tasks.
The results of the current study are unexpected and warrant further investigation to determine why female fXPCs have faster simple reaction times compared to female NTs. Historically, female fXPCs have appeared cognitively unaffected; however, since female fXPCs do in fact appear to have enhanced basic psychomotor speed, past research reporting no group difference in cognitively demanding tasks (Hunter et al., 2008) may not be null results. Reaction times for the cognitively demanding component of the task could actually be slower, given that simple reaction time is faster in female fXPCs compared to female NTs, thereby producing no group difference. Thus, this study underscores the importance of controlling for simple reaction time in cognitively demanding tasks to identify group differences in performance. Additionally, further studies into the underlying mechanisms of increased psychomotor speed in fXPCs may aid in early detection and prevention of neurodegenerative disease and may be able to identify targets for potential neurotherapeutic interventions.
Acknowledgments
This work was supported by National Institute of Health (NIH) grants: NIA RL1 AG032119, NINDS RL1 NS062412, and NIDA TL1 DA024854. This work was also made possible by a Roadmap Initiative grant (UL1 DE019583) from the National Institute of Dental and Craniofacial Research (NIDCR) in support of the NeuroTherapeutics Research Institute (NTRI) consortium. The NIH had no further role in study design; in the collection, analysis and interpretation of data; in the writing of the report; and in the decision to submit the study for publication. We thank the participants who made this work possible and Siddharth Srivastava for critical review and comments.
References
- Adams JS, Adams PE, Nguyen D, Brunberg JA, Tassone F, Zhang W, Hagerman RJ. Volumetric brain changes in females with fragile X-associated tremor/ataxia syndrome (FXTAS) Neurology. 2007;69(9):851–859. doi: 10.1212/01.wnl.0000269781.10417.7b. [DOI] [PubMed] [Google Scholar]
- Bourgeois JA, Coffey SM, Rivera SM, Hessl D, Gane LW, Tassone F, Hagerman RJ. A review of fragile X premutation disorders: Expanding the psychiatric perspective. The Journal of Clinical Psychiatry. 2009;70(6):852–862. doi: 10.4088/JCP.08m04476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bourgeois JA, Seritan AL, Casillas EM, Hessl D, Schneider A, Yang Y, Hagerman RJ. Lifetime prevalence of mood and anxiety disorders in fragile X premutation carriers. The Journal of Clinical Psychiatry. 2011;72(2):175–182. doi: 10.4088/JCP.09m05407blu. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cornish KM, Li L, Kogan CS, Jacquemont S, Turk J, Dalton A, Hagerman PJ. Age-dependent cognitive changes in carriers of the fragile X syndrome. Cortex. 2008;44(6):628–636. doi: 10.1016/j.cortex.2006.11.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Der G, Deary IJ. Age and sex differences in reaction time in adulthood: Results from the United Kingdom Health and Lifestyle Survey. Psychology and Aging. 2006;21(1):62–73. doi: 10.1037/0882-7974.21.1.62. [DOI] [PubMed] [Google Scholar]
- Garcia-Arocena D, Hagerman PJ. Advances in understanding the molecular basis of FXTAS. Human Molecular Genetics. 2010;19(R1):R83–R89. doi: 10.1093/hmg/ddq166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goodrich-Hunsaker NJ, Wong LM, McLennan Y, Srivastava S, Tassone F, Harvey D, Simon TJ. Young adult female fragile X premutation carriers show age- and genetically-modulated cognitive impairments. Brain and Cognition. 2011;75(3):255–260. doi: 10.1016/j.bandc.2011.01.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hagerman PJ. The fragile X prevalence paradox. Journal of Medical Genetics. 2008;45(8):498–499. doi: 10.1136/jmg.2008.059055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hagerman RJ, Leavitt BR, Farzin F, Jacquemont S, Greco CM, Brunberg JA, Hagerman PJ. Fragile-X-associated tremor/ataxia syndrome (FXTAS) in females with the FMR1 premutation. American Journal of Human Genetics. 2004;74(5):1051–1056. doi: 10.1086/420700. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hunter JE, Allen EG, Abramowitz A, Rusin M, Leslie M, Novak G, Sherman SL. No evidence for a difference in neuropsychological profile among carriers and noncarriers of the FMR1 premutation in adults under the age of 50. American Journal of Human Genetics. 2008;83(6):692–702. doi: 10.1016/j.ajhg.2008.10.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kolb B, Whishaw IQ. Fundamentals of human neuropsychology. 6. New York, NY: Worth Publishers; 2009. [Google Scholar]
- Myers GF, Mazzocco MMM, Maddalena A, Reiss AL. No widespread psychological effect of the fragile X premutation in childhood: Evidence from a preliminary controlled study. Journal of Developmental and Behavioral Pediatrics. 2001;22(6):353–359. doi: 10.1097/00004703-200112000-00001. [DOI] [PubMed] [Google Scholar]
- Nettelbeck T. Individual differences in noise and associated perceptual indices of performance. Perception. 1973;2(1):11–21. doi: 10.1068/p020011. [DOI] [PubMed] [Google Scholar]
- Nolin SL, Brown WT, Glicksman A, Houck GE, Jr, Gargano AD, Sullivan A, Sherman SL. Expansion of the fragile X CGG repeat in females with premutation or intermediate alleles. American Journal of Human Genetics. 2003;72(2):454–464. doi: 10.1086/367713. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Panayiotou G, Vrana SR. The role of self-focus, task difficulty, task self-relevance, and evaluation anxiety in reaction time performance. Motivation and Emotion. 2004;28(2):171–196. [Google Scholar]
- Steyaert J, Borghgraef M, Fryns JP. Apparently enhanced visual information processing in female fragile X carriers: Preliminary findings. American Journal of Medical Genetics. 1994;51(4):374–377. doi: 10.1002/ajmg.1320510415. [DOI] [PubMed] [Google Scholar]
- Tassone F, Pan R, Amiri K, Taylor AK, Hagerman PJ. A rapid polymerase chain reaction-based screening method for identification of all expanded alleles of the fragile X (FMR1) gene in newborn and high-risk populations. The Journal of Molecular Diagnostics. 2008;10(1):43–49. doi: 10.2353/jmoldx.2008.070073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wechsler D. WAIS-III: Wechsler adult intelligence scale. San Antonio, TX: Psychological Corporation; 1997. [Google Scholar]
- Wechsler D. Wechsler abbreviated scale of intelligence. San Antonio, TX: Psychological Corporation; 1999. [Google Scholar]
- Welford AT. Choice reaction time: Basic concepts. In: Welford AT, Brebner JMT, editors. Reaction times. New York, NY: Academic Press; 1980. pp. 73–128. [Google Scholar]