

Abstract
The Arabidopsis GA3 cDNA was expressed in yeast (Saccharomyces cerevisiae) and the ability of the transformed yeast cells to metabolize ent-kaurene was tested. We show by full-scan gas chromatography-mass spectrometry that the transformed cells produce ent-kaurenoic acid, and demonstrate that the single enzyme GA3 (ent-kaurene oxidase) catalyzes the three steps of gibberellin biosynthesis from ent-kaurene to ent-kaurenoic acid.
GAs are an important group of plant growth regulators with roles in a number of plant growth and developmental processes (Hooley, 1994). Considerable progress has been made in isolating and characterizing the genes encoding enzymes of GA biosynthesis (Hedden and Kamiya, 1997), in particular the enzymes that synthesize ent-kaurene and the dioxygenases that catalyze the late steps of GA biosynthesis. The intermediate steps that oxidize ent-kaurene to GA12 are catalyzed by a number of Cyt P450 monooxygenases. Two Cyt P450 genes implicated in GA biosynthesis have been isolated. The maize Dwarf3 gene encodes a member of the CYP88 family of Cyt P450 functions (Winkler and Helentjaris, 1995), but although the dwarf3 mutant responds to GA, the point of the lesion in GA biosynthesis is unknown. The Arabidopsis GA3 gene encodes a Cyt P450 protein, which is a member of the CYP701 family (Helliwell et al., 1998). The ga3 mutant accumulates ent-kaurene and shows a growth response to ent-kaurenoic acid but not ent-kaurene, and only a slight response to ent-kaurenol (Helliwell et al., 1998). These data are consistent with GA3-encoding ent-kaurene oxidase, which has been proposed to catalyze the three-step oxidation of ent-kaurene to ent-kaurenoic acid (Fig. 1). A direct demonstration of this activity has not been made.
Figure 1.

The intermediates of the GA biosynthetic pathway from ent-kaurene to ent-kaurenoic acid.
The pea lh-2 mutant is also blocked in ent-kaurene oxidation. Using extracts from embryos, Swain et al. (1997) demonstrated that the mutant was not able to oxidize the radiolabeled intermediates ent-kaurene, ent-kaurenol, or ent-kaurenal, but was able to oxidize ent-kaurenoic acid. Extracts from wild-type plants could metabolize all four substrates. These data suggest that a single enzyme catalyzes these three reactions, although the mutation could be in a gene encoding a regulatory protein affecting all three steps. Other data also support the proposal that the oxidation of ent-kaurene to ent-kaurenoic acid is catalyzed by a single enzyme. Coolbaugh et al. (1978) showed that in wild cucumber (Marah macrocarpus) the inhibition of each of the three steps from ent-kaurene to ent-kaurenoic acid by ancymidol had the same kinetics, whereas inhibition of oxidation of ent-kaurenoic acid to ent-7α-hydroxykaurenoic acid was greater, suggesting that it is catalyzed by a different enzyme.
In this paper we describe the expression of the Arabidopsis GA3 cDNA in yeast (Saccharomyces cerevisiae) and show that the GA3 protein does catalyze the three-step oxidation of ent-kaurene to ent-kaurenoic acid.
MATERIALS AND METHODS
Expression in Yeast
The entire GA3 cDNA was amplified by PCR and inserted between the glyceraldehyde 3-phosphate dehydrogenase promoter and glyceraldehyde 3-phosphate dehydrogenase terminator in the pYE22 multiple cloning site vector, a modification of the pYE2211 vector (Ashikari et al., 1989) in which a polylinker containing EcoRI, SalI, KpnI, NotI, and BamHI restriction sites was inserted in place of the glyceraldehyde 3-phosphate dehydrogenase open reading frame. Plasmids were prepared from single colonies after the transformation of Escherichia coli and the cDNA insert was sequenced to confirm that the fragment would encode the precise protein encoded by the GA3 cDNA. The sequenced plasmid was then used to transform the yeast (Saccharomyces cerevisiae) strain G1315 by a lithium chloride method (Cullin and Pompon, 1988). The transformation mixture was plated on a minimal medium consisting of 0.67% (w/v) yeast nitrogen base without amino acids, 2% (w/v) Glc and 2% (w/v) agar to select for transformants. Yeast RNA was extracted by the hot acidic-phenol method (Ausubel et al., 1993). Yeast microsomes were prepared using an enzymatic digestion method (Pompon et al., 1996).
Enzyme Assays
Single colonies of transformed yeast and untransformed controls were used to inoculate 50-mL cultures in a yeast peptone dextrose medium containing 1% (w/v) Bacto yeast extract, 2% (w/v) Bacto (Difco, Detroit, MI) peptone, and 2% (w/v) Glc. After growing overnight, 0.5 mL of each culture was removed; the yeast was pelleted and resuspended in 0.5 mL of a reaction mixture containing 100 mm Tris-HCl pH 7.5, 0.5 mm NADPH, and 0.5 mm FAD, according to the method of Hazebroek et al. (1993). The substrates added were 25 μg of ent-kaurene, 5 μg of ent-kaurenol, or 20 μg of [17,17-2H2]-ent-kaurenal. The substrates were dissolved in 100% methanol before they were added to the reaction mixture; the final methanol concentration in the reaction mixture was 5%. The reactions were incubated for 1 h at 30°C, with shaking at 150 rpm. At the end of the incubation the reaction mixture was extracted, once with 0.5 mL of hexane and twice with 0.5 mL of ethyl acetate. The organic fractions were then pooled and dried using a Speed-Vac (Savant Instruments, Farmingdale, NY) before derivatization for GC-MS. Assays with yeast microsomal fractions were carried out as described above with 100 μg of microsomal protein replacing the yeast cells.
Analysis of ent-Kaurene Metabolites by GC-MS
For analysis by GC-MS some metabolites of ent-kaurene require methylation or trimethylsilylation. Dried samples were dissolved in 50 μL of methanol and methylated with excess diazomethane (about 200 μL), after which the samples were redried. Trimethylsilylation was carried out by the addition of 5 μL each of pyridine and N,O-bis(TMS)trifluoroacetamide plus 1% trimethylchlorosilane (Alltech Associates, Deerfield, IL).
Samples were injected onto a BPX-5 column (25-m × 0.22-mm i.d. [SGE, Austin, TX]) with a 0.25-μm-thick 5% phenyl (equivalent) polysilphenylene-siloxane stationary phase and analyzed in a full-scan mode. A second injection of the samples was made onto a HP-1 column (25-m × 0.2-mm i.d., [Hewlett-Packard]) with a 0.33-μm dimethyl polysiloxane stationary phase. Both columns were nonpolar. The HP-1 column achieved better separation of the ent-kaurenoic acid and the ent-kaurenol in particular and also further confirmed the identities of all metabolites. GC conditions were as described by Green et al. (1997). We co-injected all of the sample (1 μL) with a series of hydrocarbons derived from Parafilm (Gaskin et al., 1971) for KRI determination, and used authentic standards (from L.N. Mander, Australian National University, Canberra, and J.D. Metzger, Ohio State University, Columbus) for a comparison. We also compared the full scans with a PC-based spectral library (Gaskin and MacMillan, 1991).
RESULTS
Yeast colonies were picked after a transformation with the GA3 cDNA construct. We then performed RNA gel-blot analysis to identify the transformed yeast cell line with the highest expression of the GA3 mRNA (Fig. 2) and used this cell line for the subsequent analysis of GA3 enzyme activity.
Figure 2.
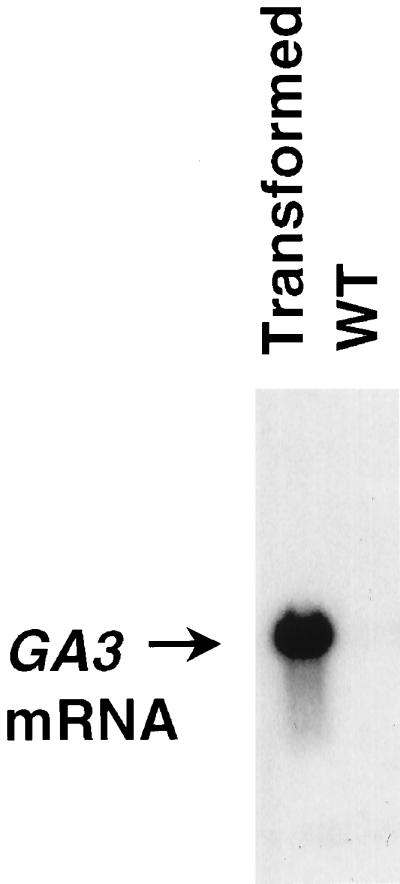
RNA gel blot of total RNA from nontransformed yeast and yeast transformed with the GA3 expression construct probed with a GA3-specific probe. WT, Wild type.
Yeast Expressing GA3 Are Able to Metabolize ent-Kaurene to ent-Kaurenoic Acid
Yeast cells expressing the GA3 mRNA and control cells of untransformed yeast were incubated with ent-kaurene. Immediately after the incubation the cells were extracted and prepared for GC-MS. Authentic ent-kaurene, ent-kaurenol, ent-kaurenal, and ent-kaurenoic acid, derivatized where appropriate, were also injected to generate KRIs and ion spectra for these compounds.
Ion current peaks in the extracts from GA3-expressing and nontransformed yeast were initially compared with a library of spectra. ent-Kaurene could be identified in the extracts from both the nontransformed and the GA3-expressing yeast. In the extracts from GA3-expressing yeast, peaks were present that were putatively assigned as ent-kaurene, ent-kaurenol, ent-kaurenal, and ent-kaurenoic acid (Fig. 3). None of these peaks was present in the extracts from nontransformed yeast. In experiments in which the yeast cells expressed a different P450 cDNA, pumpkin CYP88A2 (C.A. Helliwell and E.J. Dennis, unpublished results), only ent-kaurene could be detected. KRIs were calculated for the putatively identified peaks and compared with those of authentic standards; this calculation was carried out using data from both the BPX-5 and HP-1 columns (Table I). We found that the KRIs for the putative peaks did not differ significantly from those of the authentic compounds. Confirmation of the identity of the putative peaks was provided by a comparison of the relative abundances of characteristic ions of the putative and reference compounds (Table I). The HP-1 column gave the best separation of the compounds, particularly of ent-kaurenol and ent-kaurenoic acid.
Figure 3.

Relative abundances of the base peak ions (as in Table I) of ent-kaurenal (286), ent-kaurenoic acid (316), and ent-kaurenol (270) between 15 and 18 min of retention time on the HP-1 column for extracts of GA3-expressing (A) and nontransformed (B) yeast after feeding of ent-kaurene.
Table I.
Identification of metabolites after feeding ent-kaurene to yeast transformed with the Arabidopsis GA3 clone
| Putativeaand Reference Compoundb | KRI
|
Characteristic
Ions
|
||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| BPX-5 | HP-1 | Relative Abundance of Base Peakc | ||||||||||
| % | ||||||||||||
| 360 (M+) | 345 | 270 | 257 | 255 | 241 | 227 | 187 | 175 | 161 | |||
| ent-Kaurenol TMS | 2244 | 2330 | 25 | 7 | 100 | 80 | 21 | 11 | 22 | 10 | 19 | 23 |
| ent-Kaurenol TMS std. | 2244 | 2329 | 23 | 7 | 100 | 72 | 24 | 14 | 14 | 11 | 23 | 26 |
| 286 (M+) | 271 | 257 | 243 | 225 | 215 | 199 | 187 | 161 | ||||
| ent-Kaurenal | 2231 | 2257 | 100 | 11 | 35 | 61 | 27 | 19 | 34 | 41 | 42 | |
| ent-Kaurenal std. | 2233 | 2258 | 100 | 9 | 35 | 62 | 27 | 12 | 35 | 36 | 34 | |
| 316 (M+) | 301 | 273 | 257 | 256 | 241 | 213 | 199 | 187 | 159 | |||
| ent-Kaurenoic acid methyl ester | 2242 | 2286 | 100 | 29 | 55 | 92 | 38 | 69 | 29 | 18 | 27 | 22 |
| ent-Kaurenoic acid methyl ester std. | 2242 | 2285 | 100 | 26 | 52 | 93 | 37 | 69 | 29 | 19 | 19 | 21 |
Samples were analyzed, following appropriate derivatization, by full-scan GC-MS on both BPX-5 and HP-1 capillary columns.
Comparison was with unlabeled standards (std.), and all samples were coinjected with parafilm so that KRI values could be compared.
Data are from full scan on an HP-1 column.
Extending the length of incubation time increased the abundance of the ent-kaurenoic acid ions for the GA3-expressing yeast (data not shown). ent-Kaurenoic acid accumulation was approximately linear over a 2-h incubation. The abundances of the intermediates ent-kaurenol and ent-kaurenal were approximately 5-fold lower than ent-kaurenoic acid and did not vary greatly over the 2-h incubation, presumably because these intermediates were metabolized to ent-kaurenoic acid.
In experiments in which microsomes (approximately 0.2 mg of protein) prepared from GA3-expressing and nontransformed yeast were assayed, no metabolism of ent-kaurene was observed in preparations from nontransformed cells. In the incubations of the microsomes from GA3-expressing cells, ent-kaurenol was detected but not ent-kaurenal or ent-kaurenoic acid.
Yeast Expressing GA3 Metabolize ent-Kaurenol and ent-Kaurenal
To confirm that the GA3-expressing yeast cells were catalyzing all three steps of the GA biosynthetic pathway from ent-kaurene to ent-kaurenoic acid, both GA3-expressing and nontransformed yeast cells were incubated with ent-kaurenol or [17,17-2H2]-ent-kaurenal (Table II). Neither ent-Kaurenoic acid nor [17,17-2H2]-ent-kaurenoic acid was detected in extracts from the nontransformed yeast incubated with ent-kaurenol or [17,17-2H2]-ent-kaurenal. The extracts from the GA3-expressing yeast incubated with ent-kaurenol or [17,17-2H2]-ent-kaurenal contained ent-kaurenoic acid or [17,17-2H2]-ent-kaurenoic acid, respectively. [17,17-2H2]-ent-Kaurenal was not detected in reactions where it was included as a substrate, which could be due to nonenzymatic oxidation or metabolism by the yeast cells to a product other than ent-kaurenoic acid.
Table II.
Summary of intermediates detected after a 1-h incubation of yeast expressing the GA3 cDNA with ent-kaurene, ent-kaurenol, and ent-kaurenal
| Feed | Intermediates Detected by GC-MS
|
|||
|---|---|---|---|---|
| ent-Kaurene | ent-Kaurenol | ent-Kaurenal | ent-Kaurenoic acid | |
| ent-Kaurene | Yes | Yes | Yes | Yes |
| ent-Kaurenol | n.d.a | Yes | n.d. | Yes |
| ent-Kaurenal | n.d. | n.d. | n.d. | Yes |
n.d., Not detected.
DISCUSSION
We have expressed the Arabidopsis GA3 gene CYP701A3 in yeast and shown that the transformed cells were able to carry out the three-stage oxidation of ent-kaurene to ent-kaurenoic acid. The cells were also able to catalyze the oxidation of the intermediates ent-kaurenol and ent-kaurenal to ent-kaurenoic acid. These data showed that all three oxidation steps were enzymatically catalyzed and not due to spontaneous oxidation. The results confirmed previous evidence that GA3 encodes ent-kaurene oxidase. These earlier studies were based on growth responses to fed intermediates and measurements of ent-kaurene accumulation in the ga3-1 mutant (Helliwell et al., 1998).
Our results demonstrated that yeast was a suitable system for analyzing the function of this GA biosynthetic enzyme. It may now be possible to determine which step of GA biosynthesis is catalyzed by the maize Dwarf3 protein using this expression system. In the case of ent-kaurene oxidase, whole-yeast cells were a better system than the purified microsomal fraction of the cells. Whole cells expressing GA3 consistently gave metabolism of ent-kaurene to ent-kaurenoic acid, whereas the microsomes carried out the single-step conversion to ent-kaurenol only. The ability of the microsomes to metabolize ent-kaurenol and ent-kaurenal was not tested. A possible explanation for the difference in activity between whole cells and microsome preparations is that ent-kaurene oxidase was unstable in the microsome preparations. The ent-kaurene oxidase from the fungus Gibberella fujikuroi has been reported to lose activity rapidly in assays using crude lysates (Ashman et al., 1990). Another possibility is that in purification of the microsomal fraction, a cofactor essential for the oxidation of ent-kaurenol was lost, but the oxidation of ent-kaurene to ent-kaurenol could still proceed.
Our successful expression of the GA3 protein in a functional form in yeast is the first direct demonstration, to our knowledge, of the activity of a Cyt P450 enzyme of the GA biosynthesis pathway.
ACKNOWLEDGMENTS
We thank Ying Luo for technical assistance and Professors L.N. Mander and J.D. Metzger for providing GA intermediates.
Abbreviations:
- KRI
Kovat's retention index
- TMS
trimethylsilyl
LITERATURE CITED
- Ashikari T, Kiuchi-Goto N, Tanaka Y, Shibano Y, Amachi T, Yoshizumi H. High expression and efficient secretion of Rhizopus oryzae glucose amylase in the yeast Saccharomyces cerevisiae. Appl Microbiol Biotechnol. 1989;30:515–520. [Google Scholar]
- Ashman PJ, Mackenzie A, Bramley PM. Characterization of ent-kaurene oxidase activity from Gibberella fujikoroi. Biochim Biophys Acta. 1990;1036:151–157. doi: 10.1016/0304-4165(90)90027-t. [DOI] [PubMed] [Google Scholar]
- Ausubel FM, Brent R, Kingston RE, Moore DD, Seidman JG, Smith JA, Struhl K (1993) Current Protocols in Molecular Biology. John Wiley and Sons, New York, pp 13.12.1–13.12.3
- Coolbaugh RC, Hirano SS, West CA. Studies on the specificity and site of action of α-cyclopropyl-α-[p-methoxyphenyl]-5-pyrimidine methyl alcohol (ancymidol), a plant growth regulator. Plant Physiol. 1978;62:571–576. doi: 10.1104/pp.62.4.571. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cullin C, Pompon D. Synthesis of functional mouse cytochromes P-450 P1 and chimeric P-450 P3–1 in the yeast Saccharyomyces cerevisiae. Gene. 1988;65:203–217. doi: 10.1016/0378-1119(88)90457-x. [DOI] [PubMed] [Google Scholar]
- Gaskin P, MacMillan J (1991) GC-MS of the Gibberellins and Related Compounds: Methodology and a Library of Spectra. Cantock's Enterprises, Bristol, UK. Data converted to MS Chemstation format by Steve Croker
- Gaskin P, MacMillan J, Firn RD, Pryce RJ. “Parafilm:” a convenient source of n-alkanes for determination of gas chromatographic retention indices. Phytochemistry. 1971;10:1155–1157. [Google Scholar]
- Green LS, Faergestad EM, Poole A, Chandler PM. Grain developmental mutants of barley. α-Amylase production during grain maturation and its relation to endogenous gibberellic acid content. Plant Physiol. 1997;114:203–212. doi: 10.1104/pp.114.1.203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hazebroek JP, Metzger JD, Mansager ER. Thermoinductive regulation of gibberellin metabolism in Thlaspi arvense L. II. Cold induction of enzymes in gibberellin biosynthesis. Plant Physiol. 1993;102:547–552. doi: 10.1104/pp.102.2.547. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hedden P, Kamiya Y. Gibberellin biosynthesis: enzymes genes and their regulation. Annu Rev Plant Physiol Plant Mol Biol. 1997;48:431–460. doi: 10.1146/annurev.arplant.48.1.431. [DOI] [PubMed] [Google Scholar]
- Helliwell CA, Sheldon CC, Olive MR, Walker ARW, Zeevaart JAD, Peacock WJ, Dennis ES. Cloning of the Arabidopsis ent-kaurene oxidase gene GA3. Proc Natl Acad Sci USA. 1998;95:9019–9024. doi: 10.1073/pnas.95.15.9019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hooley R. Gibberellins: perception, transduction and responses. Plant Mol Biol. 1994;26:1529–1555. doi: 10.1007/BF00016489. [DOI] [PubMed] [Google Scholar]
- Pompon D, Lauerat B, Bronine A, Urban P. Yeast expression of animal and plant P450s in optimized redox environments. Methods Enzymol. 1996;272:51–64. doi: 10.1016/s0076-6879(96)72008-6. [DOI] [PubMed] [Google Scholar]
- Swain SM, Reid JB, Kamiya Y. Gibberellins are required for embryo growth and seed development in pea. Plant J. 1997;12:1329–1338. [Google Scholar]
- Winkler RG, Helentjaris T. The maize Dwarf3 gene encodes a cytochrome P450-mediated early step in gibberellin biosynthesis. Plant Cell. 1995;7:1307–1317. doi: 10.1105/tpc.7.8.1307. [DOI] [PMC free article] [PubMed] [Google Scholar]