

Abstract
Training context can influence resistance to disruption under differing reinforcement schedules. With nonhumans, when relatively lean and rich reinforcement schedules are experienced in the context of a multiple schedule, greater resistance is found in the rich than the lean component, as described by behavioral momentum theory. By contrast, when the schedules are experienced in separated blocks of sessions (i.e., as single schedules), resistance is not consistently greater in either component. In the current study, two groups of 6 children with intellectual disabilities responded to stimuli presented in relatively lean or rich components. For both, reinforcers were delivered according to the same variable-interval reinforcement schedule; additionally, the rich component included the delivery of response-independent reinforcers. The Within group was trained on a multiple schedule in which lean and rich components alternated regularly within sessions; the Blocked group was trained on two single schedules in which sessions with either the lean or rich schedule were conducted in successive blocks. Disruption tests presented a concurrently available alternative stimulus disrupter signaling the availability of tangible reinforcers. All 6 Within participants showed greater resistance to disruption in the rich component, consistent with behavioral momentum theory. By contrast, there was no consistent or significant difference in resistance for Blocked participants. This finding is potentially relevant to the development of interventions in applied settings, where such interventions often approximate single schedules and include response-independent reinforcers.
Keywords: resistance to disruption, behavioral momentum, intellectual disability, children, touchscreen
A number of studies in both basic and applied research have documented a positive relation between the rate of reinforcement in a discriminated operant and the behavior's resistance to change (e.g., Mace et al., 1990; Nevin, 1974; Nevin, Tota, Torquato, & Shull, 1990; see Nevin & Grace, 2000a for a review). Nevin has described this relationship in terms of behavioral momentum (Nevin, 1992), an analogy between the physics of motion and behavioral persistence. In classical mechanics, the product of a moving body's mass and velocity determines its momentum. The degree to which an outside force can disrupt the motion of that moving body depends on its momentum; increasing mass while holding velocity constant increases resistance to change. Behavioral momentum theory suggests behavioral parallels in which response rate is analogous to velocity and a behavioral characteristic analogous to mass can be estimated by measuring the resistance of response rate to some disrupting operation (e.g., extinction, prefeeding).
Behavioral momentum is most often investigated in multiple schedule procedures in which presentations of distinct discriminative stimuli are alternated and each stimulus is associated with a different reinforcement rate. After a period of exposure to the multiple schedule, a disruption is arranged in both components by extinction, alternative reinforcement, prefeeding or other external variables that can be equally applied to both components. Research by Nevin and others has shown that resistance to disruption is greater in the component that has the richer reinforcement schedule (e.g., Nevin, 1974) and is primarily determined by stimulus–reinforcer contingencies (i.e., resistance to disruption may be independent of response rate; Nevin et al., 1990).
Within multiple-schedule and concurrent chain preparations, behavioral momentum effects have been demonstrated with a variety species, schedule values, and disrupters (e.g., Cohen, 1996; Harper, 1996; Mauro & Mace, 1996; McLean, Campbell-Tie, & Nevin, 1996; Nevin, 1992; see Nevin & Grace, 2000a for a review). Some have questioned whether similar effects are found in other preparations, such as single schedules (e.g., Cohen, 2000). Here, some data suggest that resistance to disruption is not related to reinforcer rate (Cohen, Riley, & Weigle, 1993; Cohen, 1998). Single schedules differ from multiple schedules in that only one reinforcement schedule is in effect for each session, rather than two (or more). When training is conducted under single schedule conditions, responding is not always more resistant to disruption in the richer of two single schedules. For example, Cohen (1998) compared resistance to prefeeding and extinction for both relatively lean and rich reinforcement schedules in rats under three different training conditions within-subjects. The manner in which the rats experienced each reinforcement schedule differed across conditions, but in each condition, both the lean and rich schedules were associated with a unique discriminative stimulus (constant vs. flashing houselight). In the Successive (or Blocked) condition, the rats experienced only one reinforcement schedule each session until stable responding was achieved, followed by disrupter tests. Then, training and disruption testing was conducted with the other schedule. In the Daily Alternation condition, only one reinforcement schedule was experienced per session, but sessions with the lean and rich schedules alternated daily. Disruption testing was conducted for each schedule successively. Finally, in the Within-session condition, components with each reinforcement schedule alternated throughout each session during both training and disruption testing (i.e., the typical multiple schedule paradigm). Resistance to disruption appeared to depend on both the training context and the disrupter used. When training was conducted in the multiple-schedule context, momentum effects were found with both disrupters, but when training was conducted in successive blocks (i.e., single schedules), there was little evidence consistent with behavioral momentum. When training alternated each schedule daily, momentum effects were found under prefeeding but not under extinction disrupter conditions.
The reason why resistance to disruption sometimes differs under single versus multiple schedules is unclear, but the two types of schedules differ in more than one way. First, multiple schedules arrange conditions of differential reinforcement: Distinctive stimuli (e.g., red and green response keys) signal the rich and lean reinforcement conditions. By contrast, with single schedules there is only one stimulus condition (e.g., the entire experimental apparatus) when responding is reinforced and so nondifferential reinforcement is arranged. In addition, the differential and nondifferential reinforcement conditions may influence the development of stimulus control. In the multiple schedule, discriminative stimuli may come to control behavior because they are the only stimuli that distinguish the different reinforcement schedules. In the single schedule, the aspects of the stimulus context that gain stimulus control may be unclear. Finally, in the multiple schedule, there are relatively frequent alternations between signaled reinforcement schedules, but in the single schedule, there is not.
Cohen (1998) also demonstrated that reinforcer rate effects were not consistent with behavioral momentum predictions under single-schedule conditions even when stimulus control by an experimenter-defined discriminative stimulus was documented through other means (see Cohen, 1998 for an in-depth analysis of these issues). This finding suggests that the differences in resistance to disruption between single and multiple schedules may be related to frequent alternation of the rich and lean schedules (Nevin & Grace, 2000b).
Because behavioral momentum theory has implications for differential reinforcement of other/alternative behavior procedures that are frequently used in clinical settings (Ahearn, Clark, Gardenier, Chung, & Dube, 2003; Mace et al., 2010), demonstrations of behavioral momentum have been extended to human populations with intellectual disabilities in both basic and applied settings (reviewed in Dube, Ahearn, Lionello-DeNolf, & McIlvane, 2009). For example, Mace et al. (1990) taught two adult group-home residents to sort plastic dinnerware on the basis of color. Correct sorts of one color resulted in snack foods delivered at a relatively rich reinforcement rate while correct sorts of the other color resulted in foods delivered at a relatively lean rate. During test sessions, a videotape of a music/dance television program was played while the residents completed the sorting task. In tests, sorting rates fell for both tasks, but rates fell less for the task associated with the rich rate than the task associated with the lean rate. Similar results have been obtained in children with autism in a classroom setting (Parry-Cruwys et al., 2011).
Other studies using multiple-schedule procedures have been conducted in laboratory settings in which children with intellectual disabilities worked on computer-presented games or discrimination tasks with responding on a computer touchscreen (Dube, Mazzitelli, Lombard, & McIlvane, 2000; Dube & McIlvane, 2001; Dube, McIlvane, Mazzitelli, & McNamara, 2003; Lionello-DeNolf, Dube & McIlvane, 2010). Together, these studies provided evidence for behavioral momentum effects across different reinforcement schedule types (variable-interval, variable-ratio), types of reinforcers (tokens, food), tasks (free operant, discrete trial) and disrupters (alternative stimulus, movie plus prefeeding, and some evidence for response-independent food plus praise).
The influence of contextual factors on behavioral persistence may be of particular concern to clinicians interested in developing or augmenting interventions designed to increase behavioral persistence of desirable behavior and/or reduce persistence of undesirable behavior. The experiences of individuals in the natural environment seem likely to sometimes resemble single schedules and at other times resemble multiple schedules. Thus, interventions based on behavioral momentum theory may benefit from taking into account the ways in which reinforcement schedules in the target environment might affect behavioral persistence. To date, however, translational research on behavioral momentum with clinical populations has not examined single-schedule versus multiple-schedule contexts.
The purpose of the present experiment was to extend the work of Cohen (1998) to children with developmental and intellectual disabilities in a laboratory setting. All participants were given training and disruption tests with the same pair of rich-versus-lean reinforcement schedules, in a single-schedule format for those in a Blocked group, and in a multiple-schedule format for those in a Within group. As in our previous research (Dube et al., 2003; Lionello-DeNolf et al., 2010), the disrupter was a stimulus signaling a concurrently available alternative source of reinforcement. With multiple-schedule training (Within), behavioral momentum theory and the research summarized above predicts that responding would be disrupted more in the lean condition than the rich, relative to baseline. If results such as those of Cohen (1998) are translatable to clinical human participants, then those results predict no consistent differences in disruption relative to baseline in the lean and rich single-schedule training (Blocked) conditions.
METHOD
Participants
Twelve students from two schools for children with developmental disabilities participated in the study. Gender, chronological age, clinical diagnosis, and mental-age equivalent scores from the Peabody Picture Vocabulary Test-III (PPVT; Dunn & Dunn, 1997) are listed in Table 1. The PPVT was administered by research assistants and clinical diagnoses were obtained from student records.
Table 1.
Participants' chronological ages, clinical diagnoses, and PPVT Mental Age Equivalent (MAE) scores.

Apparatus
All sessions took place in a quiet room that contained a table, chairs, a computer equipped with a touchscreen monitor, and a token dispenser. The researcher sat behind and slightly to the side of the participant throughout each session.
Stimuli were digital depictions of balloons, wrapped gifts, and television sets, each approximately 1.5 cm square in size. The balloons were orange, presented on a blue background, and appeared on the left third of the screen. The gifts were blue with a red bow, presented on an orange background, and appeared on the right third of the screen. The televisions were brown and gray, presented on a gray background, and appeared on the center third of the screen. Five identical copies of the stimuli appeared on the active portion of the touchscreen during each component. The stimuli were animated such that the participants tapped “moving targets” on the monitor screen (Dube et al., 2003).
The defined response was a tap to the screen. A “hit” was recorded if the tap was within the boundaries of a stimulus. Feedback was provided with each hit. The stimulus disappeared with a “popping” sound and another identical stimulus appeared in a new location. When a reinforcer was scheduled, a hit to the stimulus resulted in its disappearance in an animated explosion with distinctive sounds for each type of stimulus. During rich components, some reinforcers were delivered on a variable time (VT) schedule, independently of responding; these reinforcers were also accompanied by the animated explosion and sounds, and the location of the explosion was within the component stimulus area but uncorrelated with any specific component stimulus location. Concurrently, a reinforcer was delivered by the researcher or the token dispenser. The computer software recorded all responses to the touchscreen (hits plus touches to the background) and the number of reinforcers delivered.
Reinforcers were tokens or food items. Four children (DDA, EBG, MIB, YCD) earned poker-chip tokens which were dispensed automatically. Tokens were traded at the end of each session for snack foods, money, or access to a computer game. The remaining children earned food reinforcers that were delivered by the researcher, who placed the food item on the table in front of the participant. Reinforcers were initially selected on recommendations from the participants' classroom teachers. From those food items identified, four were chosen and presented to the participant in a 36-trial, two-item per trial, forced-choice preference test (Fisher et al., 1992). The most frequently chosen items were used in the study; these included candy, chips, and fresh fruit.
Procedure
The experimental protocol consisted of 34 sessions. For half the participants, the balloons were associated with the lean reinforcement schedule and the gifts were associated with the rich reinforcement schedule; this was reversed for the remaining half of participants. The balloon and the gift stimuli always appeared in separate components. For all participants, the television stimuli served as the alternative stimulus disrupter. Each session consisted of five or six 1-min components separated by a 10-s intercomponent interval (ICI).
Pretraining
In pretraining sessions, the balloon or the gift stimuli were presented in separate components and responses to both stimuli were reinforced on a fixed-interval 5 s schedule. There were six components per session and each participant was required to complete a minimum of two sessions.
Baseline training was conducted in three stages (see Table 2), in which the reinforcement rates associated with the balloon and gift stimuli differed. In Baseline 1 sessions, there were six components, the lean reinforcement rate was variable interval (VI) 10 s and the rich reinforcement rate was VI 10 s with a superimposed VT 7 s schedule (VI 10 s + VT 7 s). Each participant completed four such sessions. In Baseline 2 sessions, there were six components, the lean reinforcement rate was VI 12 s, and the rich reinforcement rate was VI 12 s + VT 6 s. Each participant completed 12 sessions. Each Baseline 3 session consisted of five components. The first component of each of these was presentation of the alternative stimulus (television) and responses to it were reinforced on a VI 8 s schedule. The remaining four components consisted of presentations of the lean and rich component stimuli as described for Baseline 2. All VI schedules included nine schedule values calculated as in Fleshler and Hoffman (1962).
Table 2.
Reinforcement schedules associated with each stimulus during baseline and test phases.

Participants were divided into two groups, Blocked and Within, such that the mean mental-age equivalent (MAE) score (in years) from the PPVT was similar. The mean MAE scores were 5.23 (SD = 2.7) and 4.95 (SD = 2.6) for the Blocked and Within groups, respectively. Both groups received the same amount of baseline training with the lean and rich component stimuli, but differed in the manner in which the components were presented (see Table 3). For the Blocked group, all sessions consisted of only the lean or only the rich component. That is, all six components of the Baseline 1 and 2 and the final four components of the Baseline 3 sessions involved presentation of only one stimulus (e.g., balloons) and one schedule. When baseline training was completed with those stimuli, a series of six disrupter test sessions was conducted (procedure described below). Then, the baseline training protocol was repeated using the other stimulus (e.g., gifts) and followed by a second six-session disrupter series. For BHH, COT, and MHF, the balloon stimuli were associated with the rich schedule and the gift stimuli were associated with the lean schedule, and vice versa for ELK, LLO, and MIB. Training and disrupter testing was completed with the balloon stimuli first and followed by training and disrupter testing with the gift stimuli for COT, LLO, and MHF, and vice versa for BHH, ELK, and MIB.
Table 3.
Training and testing sequence for the Blocked and Within groups.

By contrast, the Within group experienced multiple schedules with presentations of both stimuli within each session. The components alternated regularly within each session and the component presented first (balloons or gifts) alternated daily. Then, a series of six disrupter-test sessions was conducted. For JET, YCD, and ZCH, the balloon stimuli were associated with the rich schedule and the gift stimuli were associated with the lean schedule, and vice versa for DDA, EBG, and WGB.
Disrupter tests
Each disrupter test session consisted of five components. The first component was presentation of the alternative stimulus on the center of the screen. For the Blocked group, components 2–4 were presentations of either the lean or the rich stimulus as in Baseline 3. The final component was the disrupter test: The alternative stimulus was presented concurrently with the lean or the rich component stimulus. Responses to both the component and alternative stimuli continued to be reinforced as in Baseline 3 (i.e., VI 8 s for the alternative stimulus).
For the Within group, components 2 and 3 were presentations of the lean and the rich component stimuli as in Baseline 3 and components 4 and 5 were disrupter tests. In the test components, the alternative stimulus was presented with the lean component stimulus for one test and with the rich component stimulus for the other test; responses to both the component and alternative stimuli continued to be reinforced as in Baseline 3. Within each session, lean and rich components always alternated, including both baseline and test (e.g., baseline lean, baseline rich, disruption lean, disruption rich). Across disruption-test sessions, the initial test component alternated between lean or rich. By the end of the training and testing sequence, participants in each group had experienced the same number of baseline and disruption components with each stimulus.
Data analysis
Response rates were calculated for each session by dividing the total number of responses per component by the total duration in min per component. Time taken to consume the foods or collect tokens was not deducted from the total time in each component. Relative resistance to disruption was assessed by calculating disruption/baseline response rate ratios for the lean and rich components separately. For the Blocked group, the baseline data were mean responses per min in components 2–4 (the baseline components of the disrupter-test session), and disruption data were responses per min in component 5 (lean with disrupter or rich with disrupter). For the Within group, the baseline data were mean responses per min in components 2 and 3 (one baseline lean and one baseline rich component) and disruption data were responses per min in components 4 and 5 (one lean with disrupter and one rich with disrupter). A disruption/baseline ratio was calculated for each of the six test sessions for each participant, and then the ratios were averaged to obtain a single measure per participant for the test series.
RESULTS
Pretraining
All participants except JET and LLO responded to the touchscreen during the pretraining sessions without prompting. For JET and LLO, hand-over-hand guidance was used to prompt appropriate responding to the stimuli until each participant completed one session without prompting. JET received a total of 6 pretraining sessions, and LLO received a total of 10, and each received an equal number of pretraining components with the balloon and gift stimuli. All the remaining participants completed the pretraining sessions in the minimum of two sessions.
Baseline Training
Table 4 shows average responses per min for each participant for the lean and rich components, average reinforcers obtained in each component and obtained reinforcer rate ratios (rich divided by lean) for the Baseline 3 sessions conducted just prior to each test series. Obtained reinforcer rates were approximately 2 to 5 times greater in the rich than lean condition for all participants, and did not differ systematically between groups. The average obtained reinforcer rate ratio was 3.68 for the Blocked group and 3.02 for the Within group. Response rates varied across participants (but did not vary systematically between groups) and were greater in the lean condition, with two exceptions: MIB (Blocked) and WGB (Within). Thus, the added VT reinforcers in the rich condition were similarly effective in decreasing response rates for both groups.
Table 4.
Baseline average responses (reinforcers) per minute and obtained reinforcer rate ratios for Baseline 3 sessions prior to the disrupter test.

Disrupter Tests
During disrupter-test sessions, all the participants responded to the alternative stimulus when it was presented in isolation (Component 1). During disruption components (the lean or rich stimulus presented with the alternative stimulus), all the participants responded to both stimuli, except for ZCH who did not respond to the alternative stimulus in the rich condition but did in the lean condition. Response rates in all disruption components were reduced compared to baseline components, but the effect differed across groups (see Figure 1). In the figure, a value of 1.0 indicates equal responding in disruption compared to baseline whereas a value less than 1.0 indicates less responding in disruption relative to baseline. For the Blocked group, the mean disruption/baseline response rate ratios were 0.616 and 0.515 for the lean and rich components, respectively. For the Within group, the mean disruption/baseline response rate ratios were 0.389 and 0.851 for the lean and rich components, respectively. A Wilcoxon Signed-Ranks test, one-tailed, with Type 1 error level set at .05, indicated significantly greater disruption in the lean than the rich component for the Within group (p = .014) but not for the Blocked group (p = .173). (The lean/rich difference for the Within group remains significant if the Type 1 error level is adjusted for multiple comparisons; .05/2 = .025.) At the group level, therefore, there was no significant difference in disruption for the Blocked group and substantially greater disruption in the lean condition for the Within group, as predicted by behavioral momentum theory.
Fig 1.
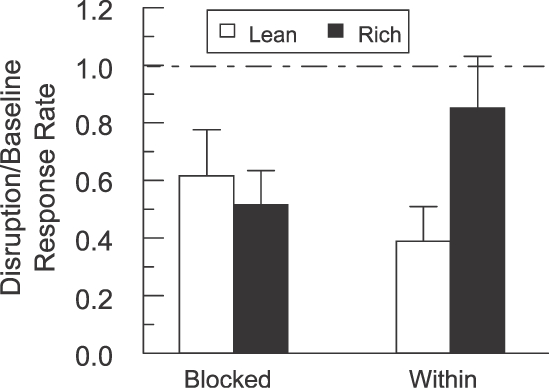
Mean disruption/baseline response rate ratios during disruption with the alternative stimulus for the Blocked and Within groups. The dashed line at 1.0 indicates equal responding in baseline and disruption. Whiskers on each bar represent standard error of the mean. Responses to the alternative stimulus are not shown.
Figure 2 summarizes the data from the alternative stimulus disrupter test for individual participants. Error bars represent standard error over the six disrupter-test sessions for each participant with two exceptions. Participant ZCH (Within group) completed only four test sessions because he graduated from the school where the study was being conducted, and Participant YCD (Within group) completed only five test sessions due to experimenter error. Although DDA completed all six test sessions, during four sessions he did not respond to the rich-component stimulus during Baseline components and data for the rich condition from those sessions are not included in the figure (data from the lean condition from those sessions are included). The data in Figure 2 indicate that the alternative stimulus was a generally effective disrupter with four exceptions: BHH (lean condition) and LLO, JET, and ZCH (rich condition). Moreover, the pattern of disruption observed at the group level was also observed at the individual subject level. For the Blocked group, 4 of the 6 participants showed greater disruption in the rich than the lean condition. In contrast, all 6 participants in the Within group showed greater disruption in the lean than the rich condition.
Fig 2.

Mean disruption/baseline response rate ratios during disruption with the alternative stimulus for individual subjects. The dashed line at 1.0 indicates equal responding in baseline and disruption. Whiskers on each bar represent standard error of the mean. Responses to the alternative stimulus are not shown.
DISCUSSION
Participants were trained to respond to distinct stimuli associated with different reinforcement rates (rich or lean) and then were given alternative-stimulus disrupter tests. For the Within group, components with each reinforcement schedule alternated regularly during each experimental session (i.e., a multiple schedule). For the Blocked group, the reinforcement schedules were experienced separately in two series of successive sessions (i.e., as single schedules). The data from the Within group demonstrated reliable behavioral momentum effects: For all 6 participants disruption to component-stimulus responding was greater in the lean condition than the rich condition when the alternative stimulus was presented and group means were significantly different. By contrast, data from the Blocked group did not demonstrate reliable behavioral momentum effects: Only 2 of 6 participants showed greater disruption in the lean than the rich component and group means did not differ significantly. The results from the Blocked group can thus be contrasted with the broader literature showing evidence for greater disruption in overall leaner contexts within multiple schedules (Lionello-DeNolf et al., 2010; Mace et al., 1990; Nevin, 1974; Nevin & Grace, 2000a). The notable difference between those reports and the Blocked group in this study is method of exposure to the two training contexts. The results, however, are consistent with Cohen (1998) and Cohen et al. (1993) with rats and pigeons, and the present results extends those findings to children with intellectual disabilities.
The procedure of the current study differed from Cohen (1998) in several ways. First, Cohen employed a within-subjects design and the current experiment employed a between-subjects design. The results of the current study are consistent with those of the within-subject design. Second, Cohen used prefeeding and extinction as disrupters. Our study extended the generality of Cohen's findings to situations in which the disrupter is a concurrently presented alternative stimulus that signals the availability of reinforcement.
Another way in which the current investigation differed from that of Cohen (1998) is that Cohen included a third condition in which sessions with the lean and rich schedules alternated on a daily basis. In that condition, behavioral momentum effects were found when prefeeding was used as a disrupter but not when extinction was used. Those data raise the question of whether differential disruption in rich versus lean contexts may be related to the interaction of two variables: the frequency of alternation between two schedules and the type of disrupter. Ahearn et al. (2003) provided evidence that session-to-session alternation can be sufficient to produce behavioral momentum effects in children with autism. In that study, 3 children were exposed to richer versus leaner reinforcer contexts in alternating sessions, and levels of stereotypy were measured during disruption by presentation of alternative stimuli, access to toys that competed with the stereotypy. The data for all participants indicated that stereotypy was more persistent in the richer sessions than the leaner sessions.
The results in the Blocked condition of the present study may at first appear to contrast with those of a recent translational study reporting greater resistance to extinction following richer than following leaner blocked conditions (Mace et al., 2010). Participants were 3 children with developmental disabilities and the target responses were severe problem behavior. The three conditions included blocked sessions of (a) baseline with reinforcement for problem behavior, (b) baseline plus differential reinforcement of alternative behavior (DRA), and (c) extinction for all responses. Mace et al. found that resistance to extinction was greater following a block of baseline-plus-DRA sessions than following a block of baseline-only sessions. Because of procedural differences, however, a comparison of the present study with Mace et al. is not straightforward. One obvious difference between the two studies is the disrupter: alternative reinforcement versus extinction. A second difference concerns the procedure for creating the richer context: in the present study a superimposed VT schedule of response-independent reinforcers, and in Mace et al. a concurrent DRA contingency with response requirements that were incompatible with the problem behavior (e.g., appropriate manual toy play as an alternative to pulling others' hair). We note that the DRA condition in Mace et al. (2010) is very similar to the disrupter condition in the present study: the presentation of an alternative reinforced response that competes with the target response. From this perspective, the comparison of interest between the two studies would be the decrease in response rates from baseline to disruption in the present study, and from baseline to baseline-plus-DRA in Mace et al. Each participant in Mace et al., however, received only one baseline to baseline-plus-DRA transition and thus a within-subject analysis of disruption as a function of baseline reinforcer rate cannot be conducted. A between-subject analysis is also impractical because of differences in responses and reinforcers across subjects. We do note, however, that the Mace et al. data do not seem inconsistent with the present results. Participant Tom had the highest baseline reinforcement rate (106/hr, fixed ratio 1 for food stealing) but the least resistance to disruption in baseline-plus-DRA (baseline-plus-DRA/baseline = 24/106 = .23); Jackie had the lowest baseline reinforcement rate (60/hr, VI 60 s for aggression) but the greatest resistance to disruption in baseline-plus-DRA (257/427 = .60).
Taken together, the results from the present study, Ahearn et al. (2003), and Mace et al. (2010) illustrate the need for continuing translational research in behavioral momentum in individuals with intellectual and developmental disabilities. Promising topics for such research include disruption by extinction versus reinforced alternative, and the possibility of an interaction between rich/lean alternation frequency and disrupter type.
Cohen (1998) suggested that one reason his single-schedule results differed from other studies that have shown greater resistance to disruption in rich than lean schedules under single-schedule conditions (e.g., Church & Raymond, 1967) may be related to the range of reinforcement rates used (see also Nevin & Grace, 2000b). That is, the difference between the rich and lean schedules may not have been great enough in Cohen's experiments. In both Cohen et al. (1993) and Cohen (1998), the rich reinforcement schedule was VI 30 s and the lean was VI 120 s, a 4∶1 programmed reinforcer rate ratio. Similar to Cohen, the programmed reinforcer rate ratio in the single-schedule (Blocked) condition of the current study was 3∶1 and the obtained rate ratios ranged from 2.8 to 4.8 (see Table 4). By contrast, in Church and Raymond, different groups of rats were trained on either a lean VI 5 min schedule or a rich VI 12 s schedule, a 25∶1 programmed reinforcer rate ratio. Greater disruption in the lean than the rich condition was found only in Church and Raymond. These findings raise another question of whether differential disruption in rich versus lean contexts may be related to the interaction of two variables: the frequency of alternation between two schedules and the degree of disparity between those two schedules, another promising topic for further research.
Research on the relation between the frequency of rich/lean reinforcer rate alternations and reinforcer rate effects on behavioral persistence may have implications for clinical interventions. Special education settings may include some situations that more closely resemble single schedules (e.g., lunch in the cafeteria, swimming) and others that are more like multiple schedules (e.g., alternation between brief periods of group and individual instruction, or group games that alternate between turn-taking and waiting for one's next turn) in which reinforcement rates differ (e.g., individual instruction is likely to include a higher rate of teacher-mediated reinforcement than group instruction). Would adaptive behavior established in the leaner context of the alternating situation be more readily disrupted than the same behavior and reinforcer schedule in a single-schedule context? Would interventions designed to decrease the persistence of problem behavior via manipulation of reinforcer rates be more effective if they included frequent alternations between the context in which the problem behavior occurs and a context associated with a richer reinforcement rate? Alternatively, if the therapeutic goal were to increase the persistence of an adaptive behavior associated with a lean reinforcement rate, would the intervention be more successful if conditions were arranged so that there were relatively longer periods between exposures to richer contexts? Future work may explore these possibilities.
Applications based on behavioral momentum theory have the potential to increase the effectiveness of current interventions and to enhance the development of new ones. If continued research shows that reinforcer rate effects on behavioral persistence are sensitive to interactions among (a) the frequency of alternation between stimuli that signal relatively richer and leaner reinforcement contexts, (b) the type of disrupter, and (c) the degree of disparity between the richer and leaner schedules, then it will be important to assess these elements within the structure of the clinical context when developing interventions.
Acknowledgments
This research was supported by NICHD grants HD033802, HD046666, and HD04147. The contents of this paper are solely the responsibility of the authors and do not necessarily represent the official views of NICHD. We thank Emily Wheeler, Melissa Loeb, and Raseeka Premchander for their assistance in data collection; and Bill McIlvane for his advice on the experimental approach.
REFERENCES
- Ahearn W.H, Clark K.M, Gardenier N.C, Chung B.I, Dube W.V. Persistence of stereotypic behavior: Examining the effects of external reinforcers. Journal of Applied Behavior Analysis. 2003;36:439–448. doi: 10.1901/jaba.2003.36-439. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Church R.M, Raymond G.A. Influence on the schedule of positive reinforcement on punished responding. Journal of Comparative and Physiological Psychology. 1967;63:329–332. doi: 10.1037/h0024382. [DOI] [PubMed] [Google Scholar]
- Cohen S.L. Behavioral momentum of typing behavior in college students. Journal of Behavioral Analysis and Therapy. 1996;1:36–51. [Google Scholar]
- Cohen S.L. Behavioral momentum: The effects of the temporal separation of rates of reinforcement. Journal of the Experimental Analysis of Behavior. 1998;69:29–47. doi: 10.1901/jeab.1998.69-29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cohen S.L. Behavioral momentum: Issues of generality. Behavioral and Brain Sciences. 2000;23:95–96. [Google Scholar]
- Cohen S.L, Riley D.S, Weigle P.A. Tests of behavioral momentum in simple and multiple schedules with rats and pigeons. Journal of the Experimental Analysis of Behavior. 1993;60:255–291. doi: 10.1901/jeab.1993.60-255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dube W.V, Ahearn W.H, Lionello-DeNolf K.M, McIlvane W.J. Behavioral momentum: Translational research and developmental disabilities. The Behavior Analyst Today. 2009;10:238–253. doi: 10.1037/h0100668. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dube W.V, Mazzitelli K, Lombard K.M, McIlvane W.J. Assessing behavioral momentum in humans with mental retardation and unstable baselines. Experimental Analysis of Human Behavior Bulletin. 2000;18:6–11. [Google Scholar]
- Dube W.V, McIlvane W.J. Behavioral momentum in computer-presented discriminations in individuals with severe mental retardation. Journal of the Experimental Analysis of Behavior. 2001;75:15–23. doi: 10.1901/jeab.2001.75-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dube W.V, McIlvane W.J, Mazzitelli K, McNamara B. Reinforcer rate effects and behavioral momentum in individuals with developmental disabilities. American Journal of Mental Retardation. 2003;108:134–143. doi: 10.1352/0895-8017(2003)108<0134:RREABM>2.0.CO;2. [DOI] [PubMed] [Google Scholar]
- Dunn L.M, Dunn L.M. Peabody Picture Vocabulary Test-III. Circle Pines, MN: American Guidance Service; 1997. [Google Scholar]
- Fisher W, Piazza C, Bowman L.G, Hagopian L.P, Owens J.C, Slevin I. A comparison of two approaches for identifying reinforcers for persons with severe to profound disabilities. Journal of Applied Behavior Analysis. 1992;25:491–498. doi: 10.1901/jaba.1992.25-491. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fleshler M, Hoffman H.S. A progression for generating variable-interval schedules. Journal of the Experimental Analysis of Behavior. 1962;5:529–530. doi: 10.1901/jeab.1962.5-529. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harper D.N. Response-independent food delivery and behavioral resistance to change. Journal of the Experimental Analysis of Behavior. 1996;65:549–560. doi: 10.1901/jeab.1996.65-549. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lionello-DeNolf K.M, Dube W.V, McIlvane W.J. Evaluation of resistance to change under different disrupter conditions in children with autism and severe intellectual disability. Journal of the Experimental Analysis of Behavior. 2010;93:369–383. doi: 10.1901/jeab.2010.93-369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mace F.C, Lalli J.S, Shea M.C, Lalli E.P, West B.J, Roberts M, Nevin J.A. The momentum of human behavior in a natural setting. Journal of the Experimental Analysis of Behavior. 1990;54:163–172. doi: 10.1901/jeab.1990.54-163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mace F.C, McComas J.J, Mauro B.C, Progar P.R, Taylor B.A, Ervin R, Zangrillo A.N. DRA increases resistance to extinction: Clinical demonstration, animal modeling, and clinical test of one solution. Journal of the Experimental Analysis of Behavior. 2010;93:349–367. doi: 10.1901/jeab.2010.93-349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mauro B.C, Mace F.C. Differences in the effect of Pavolvian contingencies upon behavioral momentum using auditory versus visual stimuli. Journal of the Experimental Analysis of Behavior. 1996;65:389–399. doi: 10.1901/jeab.1996.65-389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McLean A.P, Campbell-Tie P, Nevin J.A. Resistance to change as a function of Stimulus–reinforcer and location–reinforcer contingencies. Journal of the Experimental Analysis of Behavior. 1996;66:169–191. doi: 10.1901/jeab.1996.66-169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nevin J.A. Response strength in multiple schedules. Journal of the Experimental Analysis of Behavior. 1974;21:389–408. doi: 10.1901/jeab.1974.21-389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nevin J.A. An integrative model for the study of behavioral momentum. Journal of the Experimental Analysis of Behavior. 1992;57:301–316. doi: 10.1901/jeab.1992.57-301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nevin J.A, Grace R.C. Behavioral momentum and the Law of Effect. Behavioral and Brain Sciences. 2000a;23:73–130. doi: 10.1017/s0140525x00002405. [DOI] [PubMed] [Google Scholar]
- Nevin J.A, Grace R.C. Behavioral momentum: Empirical, theoretical, and metaphorical issues. Behavioral and Brain Sciences. 2000b;23:117–125. doi: 10.1017/s0140525x00002405. [DOI] [PubMed] [Google Scholar]
- Nevin J.A, Tota M.E, Torquato R.D, Shull R.L. Alternative reinforcement increases resistance to change: Pavlovian or operant contingencies. Journal of the Experimental Analysis of Behavior. 1990;53:359–379. doi: 10.1901/jeab.1990.53-359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parry-Cruwys D.E, Neal C.M, Ahearn W.H, Wheeler E.E, Premchander R, Loeb M.B, Dube W.V. Resistance to disruption in a classroom setting. Journal of Applied Behavior Analysis. 2011;44:363–367. doi: 10.1901/jaba.2011.44-363. [DOI] [PMC free article] [PubMed] [Google Scholar]