

Abstract
Mutations in the renin-angiotensin system (RAS) genes are associated with congenital anomalies of the kidney and urinary tract (CAKUT). As angiotensin (Ang) II, the principal effector peptide growth factor of the RAS, stimulates ureteric bud (UB) branching in whole intact embryonic (E) metanephroi, defects in UB morphogenesis may be causally linked to CAKUT observed under conditions of disrupted RAS. In the present study, using the isolated intact UB (iUB) assay, we tested the hypothesis that Ang II stimulates UB morphogenesis by directly acting on the UB, identified Ang II target genes in the iUB by microarray and examined the effect of Ang II on UB cell migration in vitro. We show that isolated E11.5 mouse iUBs express Ang II AT1 and AT2 receptor mRNA. Treatment of E11.5 iUBs grown in collagen matrix gels with Ang II (10−5 M) increases the number of iUB tips after 48 hours of culture compared to control (4.8±0.4 vs. 2.4±0.2, p<0.01). A number of genes required for UB branching as well as novel genes whose role in UB development is currently unknown are targets of Ang II signaling in the iUB. In addition, Ang II increases UB cell migration (346±5.1 vs. 275±4.4, p<0.01 ) in vitro. In summary, Ang II stimulates UB cell migration and directly induces morphogenetic response in the iUB. We conclude that Ang II-regulated genes in the iUB may be important mediators of Ang II-induced UB branching. We hypothesize that Ang II-dependent cell movements play an important role in UB branching morphogenesis.
Keywords: kidney development, ureteric bud, angiotensin, Etv 4, Etv5
1. Introduction
Metanephric development begins when the nephric duct (ND) gives rise to the ureteric bud (UB) on embryonic (E) day E10.5 in mice and E28 in humans (Grobstein, 1953; Saxen, 1987; Ekblom, 1989; Costantini and Kopan, 2010). UB outgrowth from the ND is followed by its repetitive branching, a process called branching morphogenesis. Each UB tip can induce the adjacent metanephric mesenchyme (MM) to undergo mesenchymal-to-epithelial transition and form nephrons (from the glomerulus through the distal tubule) (Grobstein, 1953; Saxen, 1987; Ekblom, 1989; Costantini and Kopan, 2010). In turn, signals emanating from the MM control UB morphogenesis. Initial generations of UB branches ultimately undergo patterning to form the ureter, renal pelvis and calyces, whereas subsequent generations of UB branches will give rise to collecting ducts. Notably, defects in UB morphogenesis cause a spectrum of congenital anomalies of the kidney and urinary tract (CAKUT), the single most common cause of renal failure in children (NAPRTCS Annual Report, 2010).
The critical role of the renin-angiotensin system (RAS), a major regulator of the blood pressure and fluid/electrolyte homeostasis (Kobori et al., 2006), in kidney and renal collecting system morphogenesis is evident from occurrence of diverse forms of CAKUT in animals or humans as a result of RAS gene mutations. These forms of CAKUT include papillary and medullary hypodysplasia, hydronephrosis, renal tubular dysgenesis, collapsed collecting ducts, aberrant UB budding, duplicated collecting system and urinary concentrating defect (Niimura et al., 1995; Esther et al., 1996; Nagata et al., 1996; Oliverio et al., 1998; Nishimura et al., 1999; Oshima et al., 2001; Gribouval et al., 2005; Song and Yosypiv, 2011). Collectively, these findings indicate that UB and its derivatives, the collecting ducts, are principal target for the RAS.
Our previous work demonstrates that lack of endogenous Ang II in angiotensinogen (AGT)-deficient mice leads to defects in UB branching and that both Ang II AT1 (AT1R) and AT2 (AT2R) receptor signaling promotes UB branching morphogenesis in intact whole embryonic kidneys (Yosypiv et al., 2008; Yosypiv et al., 2008 Song et al., 2010a, 2010b, 2010c).
In this work, we tested the hypothesis that Ang II can directly regulate branching morphogenesis in the intact isolated UB (iUB) independent of confounding effects of the mesenchyme. Given the important role of cell movement in UB morphogenesis (Tang et al., 2002; Chi et al., 2009), we next examined the effect of Ang II on UB cell migration in vitro. In addition, we conducted a global analysis of mRNA expression in iUBs cultured with or without Ang II. The results demonstrate that Ang II, when applied directly to the iUB organ culture, stimulates UB branching and induces directional migration of cultured UB cells in vitro. Finally, we identified a UB cell-autonomous transcriptome downstream of Ang II signaling.
2. Results
2.1. Effect of Ang II on ureteric bud (UB) branching morphogenesis in iUB culture
In a previous study, we demonstrated that Ang II can induce a branching morphogenesis-like program in immortalized ureteric bud-derived cells cultured in collagen matrix gels in vitro (Iosipiv and Schroeder, 2003). In this study, we investigated the direct effects of Ang II on UB branching in a physiologically more relevant system which allows to isolate the effects of the mesenchyme and examine the specific effects of Ang II on division and elongation of ampullary structures, such as iUB culture (Qiao et al., 1999). Absense of mesenchymal contamination of the iUB preparation was confirmed by the lack of mesenchyme-specific marker, GDNF, expression in the iUB (Fig. 1A). Notably, iUBs express UB tip cell-specific marker, Ret, as well as Ang II AT1R and AT2R mRNA (Fig. 1A-C). Treatment of E11.5 iUBs grown ex vivo with Ang II (10−5 M) increased the number of UB tips compared to control after 48 hours of culture compared to control (Mean±SEM: 4.8±0.4 vs. 2.4±0.2, p<0.01) (Fig. 1D, E. Supplemental Fig. 4 and 5). These data demonstrate that Ang II can induce morphogenetic response in UB epithelia in UB cell-autonomous manner.
Figure 1.
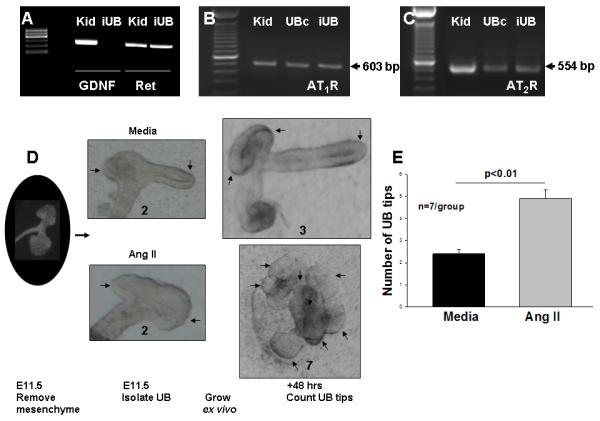
A: Isolated intact E11.5 ureteric buds (iUB) express Ret (300 bp), but not GDNF (291 bp), mRNA. E12.5 mouse kidneys (Kid) express both Ret and GDNF mRNA. B and C: iUB, immortalized UB cells (UBc) and E12.5 mouse kidneys express angiotensin (Ang) II AT1R and AT2R mRNA. D: Effect of Ang II on branching of iUBs. iUBs were dissected on E11.5 and grown in collagen matrix gels in the presence of GDNF, FGF1 and media (control) or GDNF, FGF1 and Ang II (10−5 M) for 48 hours. Numbers below each iUB image show number of UB tips (marked with arrows) at each time point. E: Bar graph shows the effect of treatments on the number of UB tips after 48 hours of culture. The number of iUB tips in higher in Ang II-treated iUBs compared with media control (Mean±SEM).
2.2. Effect of Ang II on UB cell migration in vitro
To gain further insight into the cellular mechanisms by which Ang II stimulates UB branching, we examined the effect of Ang II on UB cell movement in vitro using transwell migration assay. Ang II increased the number of cells that migrated through the filter compared with control (media) (Mean±SEM: 346±5.1 vs. 275±4.4, p=0.003) (Fig. 2A). These results demonstrate a direct stimulatory effect of Ang II on directional migration of UB cells in vitro.
Figure 2.
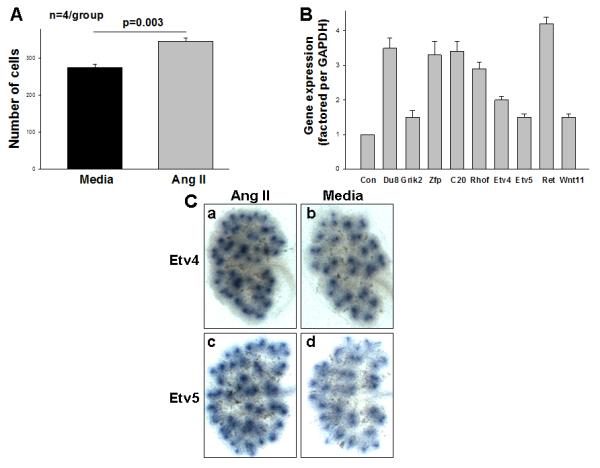
A: Bar graph shows that the number of ureteric bud-derived cells (UB cells) that migrated across the filter is higher in the presence of angiotensin (Ang) II compared to control (media) (Mean±SEM). B: Quantitative real-time reverse transcription-polymerase chain reaction confirming the changes in gene expression observed in the microarray analysis in isolated E11.5 ureteric buds treated with Ang II or media (control). Media values are normalized to 1 (Con- control) and data are presented as relative-fold difference. Du- Dusp8, Zfp- Zfp42, C20- Ccl20. C: Ang II upregulates Etv4 and Etv5 mRNA expression in E12.5 mouse metanephroi that were grown ex vivo for 24 h in the presence of Ang II (10−5 M) or media (control). After 24 h in culture, kidney explants were processed for whole-mount in situ hybridization. Representative images demonstrate that treatment with Ang II increases Etv4 (a) and Etv5 (c) mRNA expression in the UB compared to control (media, b, d).
2.3. A screen for Ang II-regulated genes in the UB
Given that only a few genes regulated by Ang II during UB morphogenesis were identified (Yosypiv et al., 2008), we conducted a global analysis of gene expression in iUBs cultured with or without Ang II. RNA was extracted from two pools of iUBs grown with Ang II and two pools of iUBs grown in the absence of Ang II, and surveyed with Agilent mouse GE4X44K chips. Of the ~44,000 transcripts represented on the GE4X44K array, 1057 were upregulated by two-fold or more with Ang II, 24 by 4-fold or more, whereas 18 were downregulated by 4-fold or more. Table 1 lists select genes most highly upregulated or downregulated by Ang II (p<0.001 to <0.05 vs. without Ang II). The genes upregulated by Ang II encode secreted and growth factors (Wnt11), adhesion molecules (Icam1), receptors (Grik2, Crlf1, Fgfr1), transcription factors (Zfp42, Etv4, Etv5), chemotactic factors (Ccl20), phosphatases (Dusp8), signaling molecules (Vsnl1, Rhof), regulators of signal transduction (Spry1) and cell cycle (Ect2, Myb), consistent with a role of Ang II in proliferation of UB tip cells (Yosypiv et al., 2008). Genes downregulated by Ang II include matrix metalloproteases (Mmp 7 and 9), receptors (Gpr123), ion transporters (Clc9a2) and secreted and growth factors (Wnt9b, Fgf12). We hypothesize that these genes together mediate the effects of Ang II on UB branching and kidney development.
Table 1.
Change in gene expression patterns in isolated intact ureteric buds cultured with angiotensin (Ang ) II or GDNF.
| Gene symbol | Gene name | Fold change | |
|---|---|---|---|
| Ang II | GDNF | ||
| Upregulated: | |||
| Dusp8 | Dual specificity phosphatase 8 | 6.5 | - |
| Dusp6 | Dual specificity phosphatase 6 | - | 2.9 |
| Grik2 | Glutamate receptor, ionotropic, kainate 2 | 6.1 | - |
| Zfp42 | Zinc finger protein 42 | 5.4 | - |
| Ccl20 | Chemokine (C-C motif) ligand 20 | 5.1 | - |
| Rhof | Ras homolog gene family, member f | 4.7 | - |
| Icam1 | Intercellular adhesion molecule | - | 4.1 |
| Crlf1 | Cytokine receptor-like factor 1 | 3.0 | 4.4 |
| Fgfr1 | Fibroblast growth factor receptor 1 | 2.5 | - |
| Etv4 | Ets variant gene 4 | 2.4 | 2.9 |
| Etv5 | Ets variant gene 5 | 2.0 | 5.5 |
| Wnt11 | Wingless-related MMTV integration site 11 |
2.0 | - |
| Ect2 | Ect2 oncogene | 2.0 | - |
| Myb | Myeloblastosis oncogene | 2.0 | 3.8 |
| Vsnl1 | Visinin like 1 | 2.0 | - |
| Spry1 | Sprouty 1 | 1.6 | 3.3 |
| Ret | Rearranged during transfection | - | 3.3 |
| Cxcr4 | Chemokine receptor 4 | - | 10.0 |
| Ccnd1 | Cyclin D1 | - | 2.9 |
| Ccnb1 | Cyclin B1 | - | 2.6 |
| Arg2 | Arginase type 2 | - | 3.5 |
| Downregulated: | |||
| Mmp9 | Matrix metalloprotease 9 | 6.1 | - |
| Gpr123 | G protein-coupled receptor 123 | 5.2 | - |
| Mmp7 | Matrix metalloprotease 7 | 5.1 | - |
| Clc9a2 | Solute-carrier 9a | 4.7 | - |
| Fgf12 | Fibroblast growth factor 12 | 4.2 | - |
| Wnt9b | Wingless-type MMTV integration site 9B | 2.0 | - |
| Dkk1 | Dickkopf homolog 1 (Xenopus laevis) | - | 1.9 |
| Vamp2 | Vesicle-associated membrane protein 2 | - | 1.9 |
| Igfbp7 | Insulin-like growth factor binding protein 7 | - | 2.0 |
| Slc40a1 | Solute carrier family 40 member 1 | - | 2.1 |
| Pth1r | Parathyroid hormone 1 receptor | - | 2.1 |
| Hspb1 | Heat shock protein 1 | - | 2.2 |
- No change. Data on the effect of GDNF are from Lu et al., 2009
Given that GDNF is essential positive regulator of UB branching (Costantini and Kopan, 2010) and that treatment of intact embryonic metanephroi ex vivo with exogenous Ang II induces GDNF expression in the mesenchyme (Yosypiv et al., 2008), we next determined direct (GDNF/Ret-independent) and indirect (GDNF/Ret-dependent) targets of Ang II in the ureteric cell lineage. To do so, we compared transcriptomes of Ang II- and GDNF-stimulated iUBs (Lu et al., 2009) (Table 1). While Ang II or GDNF caused similar changes in the expression of a number of genes, a subset of genes were differentially regulated by Ang II or GDNF. The list of genes upregulated by Ang II contained a number of GDNF targets (Etv4, Etv5, Vsnl1, Crlf1). Genes induced by Ang II, but not by GDNF, include Zfp42, Rhof and Ccl20 (Table 1). These finding indicate that a part of iUB transcriptome is regulated by Ang II independent of GDNF. We validated the results of our global gene expression analysis using quantitative real time PCR (qPCR) and in situ hybridization. qPCR demonstrated changes in mRNA expression consistent with results obtained by microarray (Fig. 2B). Even though our array assays did not reveal significant changes in Ret expression, the more sensitive qPCR assays indicated upregulation of Ret expression by Ang II (Fig. 2B).
Because members of the Pea3 family of ETS transcription factors, ETS variant (Etv) 4 and Etv5 are key transcription factors that mediate the effects of GDNF/Ret on UB morphogensis (Lu et al., 2009), we examined the spatial expression levels of Etv4/Etv5 by in situ hybridization. Consistent with qPCR findings, expression of Etv4 and Etv5 was increased in the UB tips in E12.5 metanephroi treated with Ang II (Fig. 2C). Together, these results demonstrate that Ang II regulates the expression of a number of genes required for UB branching as well as of novel genes whose role in UB development is currently unknown independent of the mesenchyme.
3. Discussion
The present study demonstrates that angiotensin (Ang) II stimulates in vitro branching morphogenesis of the isolated intact ureteric bud (iUB) and induces migration of UB cells. Expression of a number of known UB morphogenetic program genes as well as novel genes whose role in UB development is currently unknown is altered by Ang II signaling independent of the influence of the mesenchyme.
The embryonic kidney expresses all the components of the renin-angiotensin system (RAS). Angiotensinogen (AGT), Ang II AT1R and AT2R are expressed in the UB and the mesenchyme as early as on E12 in the mouse (Iosipiv and Schroeder, 2003; Song et al., 2010a). Renin-expressing cells are present in the stroma on E12 (Lopez et al., 2001). Angiotensin-converting enzyme (ACE) is detected in the embryonic kidney in rodents and humans (Jung et al., 1993; Yosypiv et al., 1994). Collectively, the fetal metanephros has the capacity to both generate Ang II and to transmit its actions.
We recently reported that Ang II, acting via the AT1R or AT2R, stimulates UB branching morphogenesis in the whole intact metanephric kidneys cultured ex vivo and induces tubulogenesis in immortalized UB cells grown in collagen matrix gels in vitro (Iosipiv and Schroeder, 2003; Yosypiv et al., 2006). In the present study, using the iUB model, we tested the hypothesis that Ang II can stimulate UB morphogenesis by directly acting on the UB, examined the role of cell movement in Ang II-induced UB branching and identified genes that mediate the effects of Ang II on UB morphogenesis independent of the mesenchyme. Consistent with our results in immortalized UB cells (Iosipiv and Schroeder, 2003), Ang II stimulated UB branching in the iUB. In light of the presence of AT1R/AT2R mRNA in the iUB, these results support the hypothesis that Ang II, generated locally in the mesenchyme, acts in a paracrine fashion on the adjacent AT1R/AT2R-expressing UBs to regulate branching in vivo.
One mechanism by which Ang II can stimulate UB branching is by inducing migration of the UB cells. In this regard, treatment with glial cell-derived neurotrophic factor (GDNF), a ligand for Ret, stimulates directional migration of Ret9-transfected Madin-Darby canine kidney (MDCK) cells towards GDNF source in vitro (Tang et al., 2002). Recent in vivo studies demonstrate that movement of Ret-expressing Wolffian duct (WD) cells accounts for the UB outgrowth from the WD (Chi et al., 2009). The present study demonstrates that Ang II induces migration of cultured UB tip cells in vitro. We speculate that Ang II-induced UB cell movement may contribute to directional bud elongation and UB branching.
Expression of a variety of novel genes was altered by Ang II in the iUB culture. Most highly upregulated genes include Dusp8, Grik2, Zfp42, Ccl20, Rhof, Crlf1, Etv4, Etv5 and Vsnl1. Dusp8 is a phosphatase which preferentially inactivates p38 and JNK MAP kinases. Although JNK is not detected in the developing kidney, inhibition of p38 retards growth of embryonic metanephroi grown ex vivo (Awazu et al, 2002). Grik2 is a member of glutamate receptor ion channels which mediate excitatory transmission in the mammalian brain (Traynelis et al., 2010). The Zfp42 (Rex-1) gene encodes a zinc finger family transcription factor which plays an important role in stem cell self-renewal and differentiation (Thompson and Gudas, 2002). Ccl20 is a chemokine which mediates renal recruitment of T lymphocytes in glomerulonephritis and renal cell carcinoma (Turner et al., 2010; Middel et al., 2010). Rhof is a member of the Rho-GTPase family which controls the formation of actin stress fibers, fundamental components of the actin cytoskeleton that produce contractile force, in MDCK cells (Fan et al., 2010). Since contraction at the apical cell surface is important during outpouching of the prospective UB tips (Meyer et al., 2004), Ang II-induced upregulation of Rhof1 expression in the UB may act to promote expansion of the ampulla. Interestingly, Ang II increased expression of ETS transcription factors Etv4 and Etv5 compared with control. Etv4/Etv5 are key transcription factors which function downstream of Ret to regulate UB morphogenesis. Specifically, Etv4−/−;Etv5+/− WD cells have limited ability to contribute to the UB tip domain in vivo (Kuure et al., 2011). Thus, it is conceivable that Ang II may stimulate UB cell movement, in part, via upregulation of Etv4/Etv5. Vsnl1, which encodes a calcium-sensing protein and is expressed exclusively in the UB tip domain, is upregulated by both Ang II and GDNF (Ola et al., 2011). Because UB tip cells contain elevated intracellular calcium levels (Ola et al., 2011) and activation of calcium signaling induces actin cytoskeleton reorganization in MDCK cells (Miranda et al., 2010), Ang II-induced upregulation of Vsnl1 expression in the UB may act to promote expansion of the ampulla by creating wedge-shaped cells through an apical purse-string mechanism (Meyer et al., 2004).
Genes downregulated by Ang II in iUB culture include matrix metalloproteases (MMP) MMP7 and MMP9. MMPs promote UB branching by degrading components of the extracellular matrix and releasing growh factors. Given that MMP9 stimulates UB branching (Arnould et al., 2009), the significance of Ang II-induced downregulation of MMP9 mRNA expression in the iUB model is not clear. Since MMP9-null metanephroi exhibit decreased levels of activated form of the stem cell factor (SCF) receptor, c-kit, expressed in the mesenchyme (Arnould et al., 2009), it is conceivable that observed decline in MMP9 expression in iUB culture may result from disrupted cross-talk between the mesenchyme and the UB. The role of MMP7 in kidney development is unknown. Of interest, MMP7 expression can be induced in the adult kidney by noncanonical WNT signaling (Jovanovic et al., 2008). Because noncanonical Wnt9b signaling is important in oriented cell division and elongation of UB-derived collecting ducts (Karner et al., 2009), decreased MMP7 levels may reflect reduced ratio of noncanonical to canonical Wnt signaling to promote UB branching (Bridgewater et al., 2008). Ingenuity Pathway Analysis of Ang II targets in the iUB revealed upregulation of the p85 and p110 subunits of the phosphatidylinositol 3-kinase (PI3K) pathway (Fig. 3). Given that pharmacologic inhibition of PI3K pathway abrogates Ang II-induced UB branching in whole embryonic metanephroi grown ex vivo (Song et al., 2010c), it is conceivable that Ang II-induced UB cell migration depends on activation of PI3K signaling.
Figure 3.
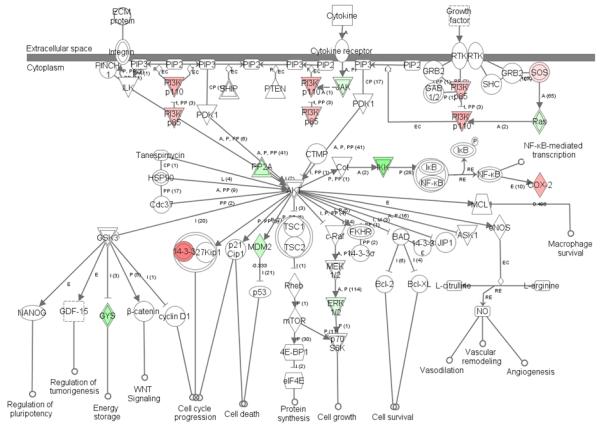
Ingenuity pathway analysis of phosphatidylinositol 3-kinase (PI3K) signaling pathway affected by angiotensin II. Pink color indicates upregulated genes; green color indicates downregulated genes.
In summary, the present study demonstrates that Ang II stimulates in vitro branching morphogenesis of the isolated ureteric bud and directional migration of UB cells. Expression of the number of genes in the UB lineage is altered by Ang II signaling independent of the influence of the mesenchyme. A part of iUB transcriptome is regulated by Ang II independent of GDNF. We hypothesize that these genes together mediate the effects of Ang II on UB branching. As RAS gene mutations in humans result in CAKUT, understanding the molecular mechanisms behind these renal birth defects would aid in developing tools that may direct preventional strategies, predict functional outcomes or guide interventions for these children.
4. Experimental Procedures
4.1. Intact isolated UB culture
Intact UBs (iUBs) were isolated from embryonic (E) day E11.5 metanephroi of Hoxb7-GFP+ mice which express green fluorescent protein (GFP) exclusively in the UB. The day when the vaginal plug was observed was considered to be E0.5. Metanephroi were incubated with trypsin/DNAse-I for 15 min at 37° C and iUBs were then mechanically dissected from the loosened mesenchyme. Absence of mesenchymal contamination was determined by the lack of GDNF expression in the iUBs. iUBs were suspended in 100 μl of Matrigel (BD Biosciences) and grown for 48 hours at 37° C on transwell filters (Corning Costar, 0.5 μm) located on top of DMEM/F12 medium (Gibco BRL) containing medium conditioned by cells derived from metanephric mesenchyme (BSN) (Shah et al., 2009), GDNF (120 ng/ml, R&D Systems, Minneapolis, MN) and FGF1 (100 ng/ml) (control, n=8) or BSN/GDNF/FGF1 and Ang II (10−5 M, Sigma, n=8). Addition of GDNF and FGF1 was required to maintain viability of iUBs grown in Matrigel. The media were changed every 24 hours. Images were acquired at time of iUB isolation and after 48 hours of culture directly from the plates via an Olympus IX70 inverted phase-contrast microscope and Olympus MagnaFire FW camera, processed with Adobe PhotoShop 7.0, and the number of UB tips was counted. All samples were blind-coded for counting.
4.2. Transwell Migration Assay
UB cells were initially obtained from microdissected ureteric buds of an E11.5 mouse transgenic for simian virus 40 (SV40) large T antigen (Immorto-mouse, Charles River) (Barasch et al., 1996). UB cells were used at passage numbers 10 to 15. Quiescent UB cells were trypsinized, resuspended in serum-free DMEM/F12 medium, plated at 1 × 105 cells/ml on top of transwell filters (24 well plates, 8-μm-pore size, Corning, Acton, MA) the bottom side of which was precoated with 2 μg/ml fibronectin, and incubated overnight at 37° C and 5% CO2. Ang II (10−5 M) or media (control) were added to the bottom of the well. Following overnight culture, the cells on the upper surface of the filter were removed with a cotton swab. Cells on the bottom of the well were fixed with paraformaldehyde (PFA 4% in PBS; Electron Microscopy Sciences, Hatfield, PA) for 15 min at room temperature and stained with hematoxylin/eosin, and the membrane was mounted on a glass slide. The mean number of cells that migrated through each membrane was determined by counting the number of cells in 5 high-power fields (x400) in each well (n=4 wells/treatment group) utilizing an Olympus IX70 inverted phase-contrast microscope. Experiment was repeated twice. All samples were blind-coded for counting.
4.3. Reverse-transcription polymerase chain reaction (RT-PCR)
Semiquantitative RT-PCR was utilized to assess potential contamination of iUB preparation by the mesenchyme and to determine whether iUBs express AT1R and AT2R mRNA. RNA was extracted using the TRIzol reagent (Invitrogen). 3 μg RNA was reverse-transcribed. cDNA was amplified using the Perkin Elmer Gene Amplification system 2400 (Cetus Instruments, Norwalk, CT) from 25% of RT mixture using gene-specific primers: AT1R: sense- 5′-GCATCATCTTTGTGGTGGG-3′; antisense- 5′-GAAGAAAAGCACAATCGCC-3′; AT2R: sense-5′-ATTCCTGTTCTCTACTAC-3′; antisense- 5′-GTAACACGTTGCTATGAA-3′; Ret: sense-5′-GGCATTAAAGCAGGCTACGGCA-3; antisense- 5′GAGGAATAACTGATTGGGAA-3′; GDNF: sense- 5′-GCCCTTCGCGCT-GACCAGTGAC-3′; antisense-5′-GATGG-TGGCGATAGTGGGATA-3′.
4.4. Agilent microarray and pathway analysis
iUBs were isolated from E11.5 CD1 mice metanephroi (Charles River Laboratories, New York, NY) and grown on air-fluid interface on polycarbonate transwell filters (Corning Costar, 0.5 μm) inserted into 6-well plates containing DMEM/F12 medium (Gibco BRL) alone or in the presence of Ang II (10−5 M) for 6 hours at 37° C and 5% CO2. At this concentration, Ang II stimulates UB branching in whole intact E11.5 metanephroi grown ex vivo and in UB cells grown in collagen matrix gels in vitro (Iosipiv and Schroeder et al., 2003; Yosypiv et al., 2006). Following incubation, iUBs were homogenized in TRIzol and stored at −80 °C. RNA was isolated from batches of 30–40 pooled ureteric buds, quantified and checked for integrity using a NanoDrop 2000 bioanalyzer (Thermo scientific). Two pools of iUBs cultured with Ang II (10−5 M) and two pools without Ang II were used for hybridization to Agilent mouse GE4X44K gene expression microarray. Hybridization, scanning and analysis were done by a core facility of the Tulane Cancer Center. Microarray data analysis was performed in R (versions 2.9-2.10) (R Development Core Team, 2009) using independently developed quality-control scripts to identify array artifacts and normalize for spatial variation. Loess and intensity quantile normalization routines from the Bioconductor limma package (Smyth et al., 2005) were applied to adjust for dye bias and variation among arrays, respectively. A linear model was then fit to the normalized data using the “lmFit” function in the limma package to estimate the average log2 ratios and associated standard errors for each probe. The Benjamini-Hochberg correction for multiple testing was applied to the set of p-values generated for the probe coefficients, and probes with adjusted p-values of <0.05 were identified as being significantly differentially expressed. Molecular pathway analyses were performed by Ingenuity Pathway Analysis version 7.1 (Redwood City, CA). All experiments involving mice were approved by Tulane Institutional Animal Care and Use Committee.
4.5. Quantitative real-time reverse-transcription polymerase chain reaction (RT-PCR)
SYBR Green quantitative real-time RT-PCR was conducted on RNA obtained from E11.5 iUBs treated or not with Ang II for 6 hours as described above for microarray analysis in the Mx3000P equipment (Stratagene, La Jolla, CA) using MxPro QPCR software (Stratagene) (Yosypiv et al., 2008). The quantity of each target mRNA expression was normalized by that of GAPDH mRNA expression. Three RNA samples per treatment group were analyzed in triplicates in each run. PCR reaction was performed twice.
4.6. In situ hybridization
Wild-type CD1 mice embryos were dissected aseptically from the surrounding tissues on E12.5 and grown on air-fluid interface on polycarbonate transwell filters (Corning Costar, 0.5 μm) inserted into 6-well plates containing DMEM/F12 medium (Gibco BRL) alone, in the presence of Ang II (10−5 M; Sigma) for 24 hours at 37° C and 5% CO2 as previously described (Yosypiv et al., 2008) and then processed for in situ hybridization. Preparation of RNA probes and whole-mount in situ hybridization were performed according to protocols (http://www.hhmi.ucla.edu/-derobertis/protocol_page/-mouse.PDF) established in the De Robertis laboratory. 5 embryonic kidneys per treatment group per probe were examined. The metanephroi were photographed using an Olympus model SC35 camera mounted on an Olympus model BH-2 microscope, and digital images were captured using Adobe Photoshop software.
4.7. Statistical analysis
Data are presented as Mean±SEM. Differences among the treatment groups in the number of UB tips and mRNA levels in media vs. Ang II were analyzed by Student’s t test. A p value of <0.05 was considered statistically significant.
Supplementary Material
Supplemental Figure 4. Effect of media on branching of iUBs. iUBs were dissected on E11.5 and grown in collagen matrix gels in the presence of GDNF, FGF1 and media (control) or GDNF, FGF1 for 48 hours. iUB tips are marked with arrows.
Supplemental Figure 5. Effect of angiotensin (Ang) II on branching of iUBs. iUBs were dissected on E11.5 and grown in collagen matrix gels in the presence of GDNF, FGF1 and Ang II (10−5 M) for 48 hours. iUB tips are marked with arrows.
HIGHLIGHTS.
Ang II stimulates in vitro branching morphogenesis of the isolated intact ureteric bud (iUB) and directional migration of UB cells.
Expression of the number of genes in the iUB is altered by Ang II signaling independent of the influence of the mesenchyme.
A part of iUB transcriptome is regulated by Ang II independent of GDNF.
Aknowledgements
This work was supported by NIH Grants P20 RR17659 and DK-71699 (Ihor Yosypiv). We thank Michelle Lacey (Tulane Cancer Center) for the assistance with microarray data analysis, Dr. Frank Costantini (Columbia University Medical Center) for providing Hoxb7-GFP+ mice, Dr. Sanjay Nigam (University of California) for BSN-conditioned media and Dr. Carlton Bates (University of Pittsburgh) for providing the probes for in situ hybridization. Microarray data are available from GEO under accession code GSE24932.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Arnould C, Lelièvre-Pégorier M, Ronco P. MMP9 limits apoptosis and stimulates branching morphogenesis during kidney development. J Am Soc Nephrol. 2009;20:2171–2180. doi: 10.1681/ASN.2009030312. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Awazu M, Omori S, Hida M. MAP kinase in renal development. Nephrol Dial Transplant. 2002;17:5–7. doi: 10.1093/ndt/17.suppl_9.5. [DOI] [PubMed] [Google Scholar]
- 3.Barasch J, Pressler L, Connor J, Malik A. A ureteric bud cell line induces nephrogenesis in two steps by distinct signals. Am J Physiol. 1996;271:F50–F61. doi: 10.1152/ajprenal.1996.271.1.F50. [DOI] [PubMed] [Google Scholar]
- 4.Bridgewater D, Cox B, Cain J, Lau A, Athaide V, Gill PS, Kuure S, Sainio K, Rosenblum ND. Canonical WNT/beta-catenin signaling is required for ureteric branching. Dev Biol. 2008;317:83–94. doi: 10.1016/j.ydbio.2008.02.010. [DOI] [PubMed] [Google Scholar]
- 5.Chi X, Michos O, Shakya R, Riccio P, Enomoto H, Licht JD, Asai N, Takahashi M, Ohgami N, Kato M, Mendelsohn C, Costantini F. Ret-dependent cell rearrangements in the Wolffian duct epithelium initiate ureteric bud morphogenesis. Dev Cell. 2009;17:199–209. doi: 10.1016/j.devcel.2009.07.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Costantini F, Kopan R. Patterning a complex organ: branching morphogenesis and nephron segmentation in kidney development. Dev Cell. 2010;18:698–712. doi: 10.1016/j.devcel.2010.04.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Ekblom P. Developmentally regulated conversion of mesenchyme to epithelium. FASEB J. 1989;3:2141–2150. doi: 10.1096/fasebj.3.10.2666230. [DOI] [PubMed] [Google Scholar]
- 8.Esther C, Jr., Howard T, Marino E, Goddard JM, Capecchi MR, Bernstein KE. Mice lacking angiotensin-converting enzyme have low blood pressure, renal pathology, and reduced male fertility. Lab Invest. 1996;7:953–965. [PubMed] [Google Scholar]
- 9.Fan L, Pellegrin S, Scott A, Mellor H. The small GTPase Rif is an alternative trigger for the formation of actin stress fibers in epithelial cells. J Cell Sci. 2010;123:1247–1252. doi: 10.1242/jcs.061754. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Gribouval O, Gonzales M, Neuhaus T, Aziza J, Bieth E, Laurent N, Bouton JM, Feuillet F, Makni S, Amar H. Ben, Laube G, Delezoide AL, Bouvier R, Dijoud F, Ollagnon-Roman E, Roume J, Joubert M, Antignac C, Gubler MC. Mutations in genes in the renin-angiotensin system are associated with autosomal recessive renal tubular dysgenesis. Nat Genet. 2005;37:964–968. doi: 10.1038/ng1623. [DOI] [PubMed] [Google Scholar]
- 11.Grobstein C. Inductive epithelio-mesenchymal interaction in cultured organ rudiments of the mouse metanephros. Science. 1953;118:52–55. doi: 10.1126/science.118.3054.52. [DOI] [PubMed] [Google Scholar]
- 12.Iosipiv I, Schroeder M. A role for angiotensin II AT1 receptors in ureteric bud cell branching. Am J Physiol. 2003;285:F199–F207. doi: 10.1152/ajprenal.00401.2002. [DOI] [PubMed] [Google Scholar]
- 13.Jovanovic V, Dugast A, Heslan J, Ashton-Chess J, Giral M, Degauque N, Moreau A, Pallier A, Chiffoleau E, Lair D, Usal C, Smit H, Vanhove B, Soulillou JP, Brouard S. Implication of matrix metalloproteinase 7 and the noncanonical wingless-type signaling pathway in a model of kidney allograft tolerance induced by the administration of anti-donor class II antibodies. J Immunol. 2008;180:1317–1325. doi: 10.4049/jimmunol.180.3.1317. [DOI] [PubMed] [Google Scholar]
- 14.Jung F, Bouyounes B, Barrio R, Tang SS, Diamant D, Ingelfinger JR. Angiotensin converting enzyme in renal ontogeny: hypothesis for multiple roles. Pediatr Nephrol. 1993;7:834–840. doi: 10.1007/BF01213370. [DOI] [PubMed] [Google Scholar]
- 15.Karner C, Chirumamilla R, Aoki S, Igarashi P, Wallingford JB, Carroll TJ. Wnt9b signaling regulates planar cell polarity and kidney tubule morphogenesis. Nat Genet. 2009;41:793–799. doi: 10.1038/ng.400. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Kobori H, Ozawa Y, Suzaki Y, Prieto-Carrasquero MC, Nishiyama A, Shoji T, Cohen EP, Navar LG. Young Scholars Award Lecture: Intratubular angiotensinogen in hypertension and kidney diseases. Am J Hypertens. 2006;19:541–550. doi: 10.1016/j.amjhyper.2005.11.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Kuure S, Chi X, Lu B, Costantini F. The transcription factors Etv4 and Etv5 mediate formation of the ureteric bud tip domain during kidney development. Development. 2011;137:1975–1979. doi: 10.1242/dev.051656. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Lopez M, Pentz E, Robert B, Abrahamson DR, Gomez RA. Embryonic origin and lineage of juxtaglomerular cells. Am J Physiol. 2001;281:F345–F356. doi: 10.1152/ajprenal.2001.281.2.F345. [DOI] [PubMed] [Google Scholar]
- 19.Lu B, Cebrian C, Chi X, Kuure S, Kuo R, Bates CM, Arber S, Hassell J, MacNeil L, Hoshi M, Jain S, Asai N, Takahashi M, Schmidt-Ott KM, Barasch J, D’Agati V, Costantini F. Etv4 and Etv5 are required downstream of GDNF and Ret for kidney branching morphogenesis. Nat Genet. 2009;41:1295–1302. doi: 10.1038/ng.476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Meyer T, Schwesinger C, Bush K, Stuart RO, Rose DW, Shah MM, Vaughn DA, Steer DL, Nigam SK. Spatiotemporal regulation of morphogenetic molecules during in vitro branching of the isolated ureteric bud: toward a model of branching through budding in the developing kidney. Dev Biol. 2004;275:44–67. doi: 10.1016/j.ydbio.2004.07.022. [DOI] [PubMed] [Google Scholar]
- 21.Middel P, Brauneck S, Meyer W, Radzun HJ. Chemokine-mediated distribution of dendritic cell subsets in renal cell carcinoma. BMC Cancer. 2010;10:578–590. doi: 10.1186/1471-2407-10-578. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Miranda L, Carpentier S, Platek A, Hussain N, Gueuning MA, Vertommen D, Ozkan Y, Sid B, Hue L, Courtoy PJ, Rider MH, Horman S. AMP-activated protein kinase induces actin cytoskeleton reorganization in epithelial cells. Biochem Biophys Res Commun. 2010;396:656–661. doi: 10.1016/j.bbrc.2010.04.151. [DOI] [PubMed] [Google Scholar]
- 23.Miyazaki Y, Tsuchida S, Fogo A, Ichikawa I. The renal lesions that develop in neonatal mice during angiotensin inhibition mimic obstructive nephropathy. Kidney Int. 1999;55:1683–1695. doi: 10.1046/j.1523-1755.1999.00420.x. [DOI] [PubMed] [Google Scholar]
- 24.Nagata M, Tanimoto K, Fukamizu A, Kon Y, Sugiyama F, Yagami K, Murakami K, Watanabe T. Nephrogenesis and renovascular development in angiotensinogen-deficient mice. Lab Invest. 1996;75:745–753. [PubMed] [Google Scholar]
- 25.Niimura F, Labosky P, Kakuchi J, Okubo S, Yoshida H, Oikawa T, Ichiki T, Naftilan AJ, Fogo A, Inagami T. Gene targeting in mice reveals a requirement for angiotensin in the development and maintenance of kidney morphology and growth factor regulation. J Clin Invest. 1995;96:2947–2954. doi: 10.1172/JCI118366. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Nishimura H, Yerkes E, Hohenfellner K, Miyazaki Y, Ma J, Hunley TE, Yoshida H, Ichiki T, Threadgill D, Phillips JA, 3rd, Hogan BM, Fogo A, Brock JW, 3rd, Inagami T, Ichikawa I. Role of the angiotensin type 2 receptor gene in congenital anomalies of the kidney and urinary tract, CAKUT, of mice and men. Mol Cell. 1999;3:1–10. doi: 10.1016/s1097-2765(00)80169-0. [DOI] [PubMed] [Google Scholar]
- 27.North American Pediatric Renal Trials and Collaborative Studies NAPRTCS Annual report. 2010.
- 28.Ola R, Jakobson M, Kvist J, Perälä N, Kuure S, Braunewell KH, Bridgewater D, Rosenblum ND, Chilov D, Immonen T, Sainio K, Sariola H. The GDNF target Vsnl1 marks the ureteric tip. J Am Soc Nephrol. 2011;22:274–284. doi: 10.1681/ASN.2010030316. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Oliverio M, Kim H, Ito M, Le T, Audoly L, Best CF, Hiller S, Kluckman K, Maeda N, Smithies O, Coffman TM. Reduced growth, abnormal kidney structure, and type 2 (AT2) angiotensin receptor-mediated blood pressure regulation in mice lacking both AT1A and AT1B receptors for angiotensin II. Proc Natl Acad Sci USA. 1998;95:15496–15501. doi: 10.1073/pnas.95.26.15496. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Oshima K, Miyazaki Y, Brock J, Adams MC, Ichikawa I, Pope JC. Angiotensin type II receptor expression and ureteral budding. J Urol. 2001;166:1848–1852. [PubMed] [Google Scholar]
- 31.Qiao J, Sakurai H, Nigam S. Branching morphogenesis independent of mesenchymal-epithelial contact in the developing kidney. Proc Natl Acad Sci U S A. 1999;96:7330–7335. doi: 10.1073/pnas.96.13.7330. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.R Development Core Team. R. A language and environment for statistical computing. R Foundation for Statistical Computing; Vienna, Austria: 2009. ISBN 3-900051-07. [Google Scholar]
- 33.Saxen L. Organogenesis of the kidney. Cambridge University Press; Cambridge: 1987. [Google Scholar]
- 34.Shah M, Tee J, Meyer T, Meyer-Schwesinger C, Choi Y, Sweeney DE, Gallegos TF, Johkura K, Rosines E, Kouznetsova V, Rose DW, Bush KT, Sakurai H, Nigam SK. The instructive role of metanephric mesenchyme in ureteric bud patterning, sculpting, and maturation and its potential ability to buffer ureteric bud branching defects. Am J Physiol. 2009;297:F1330–F1341. doi: 10.1152/ajprenal.00125.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Smyth K, Gentleman R, Carey V, Dudoit S, Irizarry R, Huber W. Bioinformatics and Computational Biology Solutions using R and Bioconductor. Springer; New York: 2005. Limma: linear models for microarray data; pp. 397–420. [Google Scholar]
- 36.Song R, Spera M, Garrett C, El-Dahr SS, Yosypiv IV. Angiotensin II AT2 Receptor Regulates Ureteric Bud Morphogenesis. Am J Physiol. 2010a;298:F807–F817. doi: 10.1152/ajprenal.00147.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Song R, Van Buren T, Yosypiv I. Histone Deacetylases are Critical Regulators of the Renin-Angiotensin System During Ureteric Bud Branching Morphogenesis. Pediatric Research. 2010b;67:573–578. doi: 10.1203/PDR.0b013e3181da477c. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Song R, Spera M, Garrett C, Yosypiv IV. Angiotensin II-induced activation of c-Ret signaling is critical in ureteric bud branching morphogenesis. Mechanisms of Development. 2010c;127:21–27. doi: 10.1016/j.mod.2009.11.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Song R, Yosypiv I. Genetics of Congenital Anomalies of the Kidney and Urinary Tract. Pediatric Nephrology. 2011;26:353–364. doi: 10.1007/s00467-010-1629-4. [DOI] [PubMed] [Google Scholar]
- 40.Tang M, Cai Y, Tsai S, Wang YK, Dressler GR. Ureteric bud outgrowth in response to RET activation is mediated by phosphatidylinositol 3-kinase. Dev Biol. 2002;243:128–136. doi: 10.1006/dbio.2001.0557. [DOI] [PubMed] [Google Scholar]
- 41.Thompson J, Gudas L. Retinoic acid induces parietal endoderm but not primitive endoderm and visceral endoderm differentiation in F9 teratocarcinoma stem cells with a targeted deletion of the Rex-1 (Zfp-42) gene. Mol Cell Endocrinol. 2002;195:119–133. doi: 10.1016/s0303-7207(02)00180-6. [DOI] [PubMed] [Google Scholar]
- 42.Traynelis S, Wollmuth L, McBain C. Glutamate receptor ion channels: structure, regulation, and function. Pharmacol Rev. 2010;62:405–496. doi: 10.1124/pr.109.002451. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Turner J, Paust H, Steinmetz O. CCR6 recruits regulatory T cells and Th17 cells to the kidney in glomerulonephritis. J Am Soc Nephrol. 2010;21:974–985. doi: 10.1681/ASN.2009070741. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Yosypiv I, Schroeder M, El-Dahr S. Angiotensin II type 1 receptor-EGF receptor cross-talk regulates ureteric bud branching morphogenesis. J Am Soc Nephrol. 2006;17:1005–1014. doi: 10.1681/ASN.2005080803. [DOI] [PubMed] [Google Scholar]
- 45.Yosypiv I, Boh M, Spera M, El-Dahr SS. Downregulation of Spry-1, an inhibitor of GDNF/Ret, as a mechanism for angiotensin II-induced ureteric bud branching. Kidney International. 2008;74:1287–1293. doi: 10.1038/ki.2008.378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Yosipiv I, Dipp S, El-Dahr S. Ontogeny of somatic angiotensin-converting enzyme. Hypertension. 1994;23:369–374. doi: 10.1161/01.hyp.23.3.369. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplemental Figure 4. Effect of media on branching of iUBs. iUBs were dissected on E11.5 and grown in collagen matrix gels in the presence of GDNF, FGF1 and media (control) or GDNF, FGF1 for 48 hours. iUB tips are marked with arrows.
Supplemental Figure 5. Effect of angiotensin (Ang) II on branching of iUBs. iUBs were dissected on E11.5 and grown in collagen matrix gels in the presence of GDNF, FGF1 and Ang II (10−5 M) for 48 hours. iUB tips are marked with arrows.