

Abstract
Unlike other essential trace elements (e.g., zinc and iron) it is the toxicity of manganese (Mn) that is more common in human populations than its deficiency. Data suggest alterations in dopamine biology may drive the effects associated with Mn neurotoxicity, though recently γ-aminobutyric acid (GABA) has been implicated. In addition, iron deficiency (ID), a common nutritional problem, may cause disturbances in neurochemistry by facilitating accumulation of Mn in the brain. Previous data from our lab have shown decreased brain tissue levels of GABA as well as decreased 3H-GABA uptake in synaptosomes as a result of Mn exposure and ID. These results indicate a possible increase in the concentration of extracellular GABA due to alterations in expression of GABA transport and receptor proteins. In this study weanling-male Sprague-Dawley rats were randomly placed into one of four dietary treatment groups: control (CN; 35 mg Fe/kg diet), iron-deficient (ID; 6 mg Fe/kg diet), CN with Mn supplementation (via the drinking water; 1 g Mn/L) (CNMn), and ID with Mn supplementation (IDMn). Using in vivo microdialysis, an increase in extracellular GABA concentrations in the striatum was observed in response to Mn exposure and ID although correlational analysis reveals that extracellular GABA is related more to extracellular iron levels and not Mn. A diverse effect of Mn exposure and ID was observed in the regions examined via Western blot and RT-PCR analysis, with effects on mRNA and protein expression of GAT-1, GABAA, and GABAB differing between and within the regions examined. For example, Mn exposure reduced GAT-1 protein expression by approximately 50% in the substantia nigra, while increasing mRNA expression approximately four-fold, while in the caudate putamen mRNA expression was decreased with no effect on protein expression. These data suggest that Mn exposure results in an increase in extracellular GABA concentrations via altered expression of transport and receptor proteins, which may be the basis of the neurological characteristics of manganism.
Keywords: manganese, GABA, rat, microdialysis, brain
Introduction
Manganese (Mn), an essential nutrient functioning as a critical cofactor for many key enzymes in the body (Hurley and Keen, 1987), is required for proper immune function, regulation of metabolism, reproduction, digestion, bone growth, and blood clotting (see review by Aschner et al., 2005). However, exposure to high concentrations of Mn is known to result in neurotoxicity. Manganese neurotoxicity, or manganism, initially characterized by a psychiatric disorder resembling schizophrenia, shares neuropathologies with several clinical disorders, in particular Parkinson’s disease (Pal et al., 1999). Manganese neurotoxicity is associated with the accumulation of Mn in iron-rich, dopaminergic regions of the brain, specifically areas of the basal ganglia (Aschner et al., 2005). Manganism is most often associated with occupational exposure of welders, miners, and steel workers to chronic high levels of airborne particulate Mn (Pal et al., 1999; Mergler et al., 1994), though cases from exposure to contaminated drinking water have been reported as well (Wasserman et al., 2006; Kondakis et al., 1989).
Iron deficiency (ID) has become a clear risk factor for Mn accumulation (Davis et al., 1992; Erikson et al., 2002; Finley, 1999; Kwik-Uribe et al., 2000). A common nutritional problem throughout the world, ID is associated with cognitive and behavioral problems (Beard, 2001), potentially disturbing neurochemistry by facilitating accumulation of Mn in the brain, as iron (Fe) status is known to affect absorption of Mn, regardless of Mn concentrations in the body (Chandra and Shukla, 1976; Shukla et al., 1976). Manganese most likely competes with Fe for transport via divalent metal transporter 1 (DMT-1) (Gunshin et al., 1997), a transporter of various divalent metals. DMT-1 mRNA expression is known to increase in cases of ID (Gunshin et al., 2001; Burdo et al., 1999), and has recently become of interest as a potential transport protein for Mn (Garrick et al., 2003; Roth and Garrick, 2003). While most research has focused on the effects of Mn neurotoxicity on the metabolism of dopamine (DA) due to locomotor effects, alterations in the metabolism of other neurotransmitters, such as γ-aminobutyric acid (GABA), have been noted (Garcia et al., 2006; 2007; Li, 1998).
GABA is the most abundant inhibitory neurotransmitter in the adult brain (Beleboni et al., 2004; Olsen and DeLorey, 1999) and is found in the medium spiny neurons of the striatum, mediating the dopaminergic activity in this region (Ade et al., 2008). Cortical glutamate afferents project into the striatum where, in concert with GABA and DA, motor behaviors are controlled (Carlsson and Carlsson, 1990). Li (1998) found alterations in GABA metabolism during ID, though no absolute changes in GABA concentration were detected. Gwiazda et al. (2002) found a significant 16% increase in striatal GABA concentrations resulting from cumulative low-dose Mn exposure (14.4 mg Mn/kg body weight i.p./week, over five weeks) in a pre-Parkinsonian rat model. More recent studies have shown a significant increase in tissue levels of GABA in rats exposed to a high Mn diet at post-natal day 21 (Garcia et al., 2006), but not at earlier time points (Garcia et al., 2007), as well as a marginally significant (p<0.1) decrease in pallidal GABA concentrations in monkeys exposed to MnSO4, with a 20% decrease in caudate and putamen of Mn exposed monkeys compared to controls (Struve et al., 2007). Erikson et al. (2002) observed an inverse correlation between Mn and GABA concentration in the caudate putamen of ID male rats. However, none of these studies examined extracellular levels of GABA, which would reflect what is happening at the synaptic level and can be associated with behaviors more relevantly than tissue levels. While various microdialysis studies have observed the effect of Mn on extracellular DA concentrations (Vidal et al., 2005) or the relationship between GABA and DA (Expósito et al., 1999; Galindo et al., 1999), the effect of dietary Mn exposure on extracellular GABA concentrations has not been examined.
A recent study by this lab found a significant decrease in 3H-GABA uptake correlating with increased Mn concentrations and ID in rat striatal synaptosomes (Anderson et al., 2007a), and we hypothesized that this was due to altered expression of GABA transporter (GAT-1). It is known that ID affects dopamine transporter (DAT) number and function (Erikson et al., 2000), and that it is a member of the solute carrier-6 (SLC-6) transporter family like the GAT-1 protein (Gether et al., 2006). The DAT has also been shown to respond to Mn and to play a role in Mn transport during toxicity (Anderson et al., 2007b; Erikson et al., 2005; Ingersoll et al., 1999). Therefore, we hypothesize that increased Mn and/or decreased Fe in the brain will cause decreased GAT-1 expression leading to elevated extracellular GABA and overall altered GABA biology, including GABA receptors, in the basal ganglia.
In the brain, the inhibitory actions of GABA are mediated by ionotropic GABAA receptors and metabotropic GABAB receptors. While GABAA receptors occur post-synaptically, GABAB receptors are present pre-synaptically as autoreceptors, post-synaptically, and extra-synaptically as heteroreceptors, taking on a more neuromodulatory role. Upregulation of peripheral type benzodiazepine receptors in globus pallidus has been shown in rats exposed to Mn (Hazell et al., 2003) and in cultured primary rat astrocytes (Hazell et al., 1999). Additionally, Mn is known to affect GABA by dose-dependently increasing binding at GABAB receptors (Kerr and Ong, 1995). Alterations in neurotransmitter receptor expression can affect expression of transport proteins of the respective neurotransmitter via regulation by autoreceptors. This is well characterized in mice lacking functional D2 receptors, which consequently experience a significant downregulation of the DAT (Dickinson et al., 1999). Similar effects on GAT-1 could be possible through action at GABAB autoreceptors (Zahniser and Doolen, 2001).
Based upon previous data from our lab showing decreased tissue levels of GABA (Erikson et al., 2002) and decreased 3H-GABA uptake (Anderson et al., 2007a) as a result of dietary Mn exposure and ID, coupled with the fact that behaviors associated with Mn neurotoxicity suggest altered GABA metabolism, we hypothesize that dietary Mn exposure could potentially lead to an increase in the concentration of extracellular GABA, most likely as a result of altered GABA transporter and/or receptor expression. To test this hypothesis, we developed this study: 1.) to establish alterations in the neuromodulatory extracellular concentrations of GABA in response to dietary Mn exposure and ID; 2.) to examine alterations in the protein and mRNA expression of the transport and receptor proteins of GABA resulting from dietary Mn exposure and ID.
Materials and Methods
Animals
Male 21-day-old Sprague-Dawley rats (Harlan Sprague-Dawley, Indianapolis, IN) (n=24 for the microdialysis studies; n=24 for protein and mRNA analysis) were randomly divided into four dietary treatment groups as in previous studies (Anderson et al., 2007a; 2007b): control (CN; 35 mg Fe/kg, 10 mg Mn/kg diet & d.i. water); control Mn-exposed (CNMn; control diet & 1 g Mn (as MnCl2)/L d.i. water); iron-deficient (ID; 4 mg Fe/kg, 10 mg Mn/kg diet & d.i. water); and iron-deficient/Mn exposed (IDMn; ID diet & 1 g Mn/L d.i. water). Diets were obtained from Bio-Serv (Frenchtown, NJ) and certified for metal content. Rats had free access to food and water 24 h/day, and the lights were turned off between 1800 and 600 h. Room temperature was maintained at 25 ± 1° C. The University of North Carolina at Greensboro Animal Care and Use Committee approved all of the animal procedures.
Stereotaxic Surgery
After five weeks of dietary treatment and one week prior to microdialysis experiments, rats were anesthetized with ketamine-HCl (80 mg/kg) and xylazine (12 mg/kg) and maintained on a heating pad at 37° C. The heads of the rats were shaved and wiped with a 5% povidone-iodine solution to reduce risk of infection. Sterile instruments and gloves were used throughout the surgical procedure. The rats were secured in the stereotaxic frame and an incision was made perpendicular to the bregma. A guide cannula (CMA/12, CMA Microdialysis, Acton, MA) was implanted into the striatum using the following coordinates: 2.4 mm lateral to midline, 7.5 mm anterior to the lambda. The cannula was lowered to a depth of 2.5 mm, positioning it in the medial area of the striatum (Paxinos and Watson, 1998). Anchoring screws were utilized to maintain the position of the cannula before being cemented into place using dental adhesive. Animals were given 0.9% sterile saline (0.5 ml/kg body weight, i.p.) to reduce fluid lost while under anesthesia and to aid in recovery time. Animals were returned to shoebox cages with Tek-Fresh bedding (Harlan, Indianapolis, IN) and monitored daily until microdialysis experiments began.
Microdialysis
During week six of the dietary protocol, a microdialysis probe (CMA/12 Elite, CMA Microdialysis, Acton, MA) was inserted into the guide cannula and the rat perfused with artificial CSF (155 mM Na+, 0.83 mM Mg2+, 2.9 mM K+, 132.76 mM Cl−, 1.1 mM Ca+, pH 7.4) for one hour at a flow rate of 1 μl/min. After perfusion, the flow rate was adjusted to 0.5 μl/min and 30 minute fractions were collected in microtubes for a total of two hours (4 samples per rat) in a refrigerated fraction collector (CMA Microdialysis, Acton MA). Samples were stored at −80° C until analysis of the dialysate fraction. To quantify levels of GABA from the microdialysate, fractions were analyzed for GABA content using capillary electrophoresis with laser induced fluorescence detection (CE-LIF) (Biorad Biofocus 2000, Hercules, CA, with 488 nm diode laser/590 nm emission filter). Rats were then returned to their home cage and, the following day, were sacrificed, brains removed, and probe placement verified post mortem. Brains were dissected into five regions (caudate putamen, globus pallidus, substantia nigra, hippocampus, and cerebellum) for metal, protein, and mRNA analyses (see below). Regions were selected based on the known heterogeneous accumulation of metals in response to alterations in dietary Fe and Mn levels (Anderson et al., 2007a, 2007b; Erikson et al., 2002) and the density of GABAergic neurons.
CE-LIF analysis
A protocol by Chen et al. (2001) allowing for detection of amino acids and biogenic amines at nanomolar concentrations was modified to accommodate the needs of this study. The advantages of applying CE analysis to neuroactive compounds include minimal required sample volumes, speed of analysis, and high separation efficiency (Powell and Ewing, 2005). Briefly, on the day of sample analysis, 5 μl of microdialysate sample were derivatized at 40°C by the addition to 100 nmol ATTO-TAG™ FQ fluorogenic reagent (Molecular Probes, Eugene, OR) and 10 μL of a 10 mM borate (Fisher, Fair Lawn, NJ)/25 mM KCN (Fluka) solution (pH 9.18). The total sample volume was adjusted to 20 μl using HPLC grade methanol (G.J. Chemical Company, Newark, NJ). After a minimum reaction time of 90 min., 1 μl of an FQ derivatized homoserine (Sigma, St.Louis, MO) internal standard solution was added to the derivatized microdialysate sample and analyzed. CE-LIF conditions leading to high efficiency peaks for GABA samples were 10 kV for 10 min with sample injections at 10 psi/sec. Uncoated silica capillary (Polymicro, Arizona) with an i.d. of 25 μm, o.d. of 361 μm, and effective/total lengths of 25.4/30.0 cm was used. The run buffer was 15 mM sodium borate (Fisher), pH 9.0, with 45 mM sodium dodecyl sulfate (Pierce, Rockford, IL), 5 mM sodium cholate (Anatrace, Maumee, OH), and 4% (v/v) 2-propanol (Fisher). Three replicates were analyzed for each sample, with a calibration curve for GABA constructed each day of sample analysis using three points with a concentration range of 50 nM to 1 μM GABA. GABA (Sigma) and homoserine standard solutions used for construction of calibration curves were prepared in artificial cerebral spinal fluid with the same composition as that used in the microdialysis studies. A representative limit of detection for this method is 6.9 nM GABA (based on 3σ, where σ represents the standard deviation of the background) with a linear dynamic range of 3.64 decades. The ratio of GABA peak height to homoserine peak height for each sample was used to determine the concentration of GABA based on the calibration curve response.
Metal analysis
Tissue Mn and Fe concentrations were measured with graphite furnace atomic absorption spectroscopy (Varian AA240, Varian, Inc., USA). Blood samples were collected at the end of the experiment into heparinized tubes, cooled to 4° C, and centrifuged in a clinical centrifuge for 15 minutes to separate cells from plasma. Plasma was frozen at −80° C until analyzed for Mn and Fe. Equal volumes of plasma and 0.5% Triton-X were vortexed for 30 s before being centrifuged at 12,000 × g for 10 minutes. The supernatant was removed and an aliquot of 50 μl brought to 1 ml total volume in 2% nitric acid and analyzed for Mn and Fe content. Brain regions were digested in ultra-pure nitric acid (1:10 w/v dilution) for 48–72 hours in a sand bath (60° C). An aliquot of 50 μl of digested tissue was brought to 1 ml total volume with 2% nitric acid and analyzed for Mn and Fe. Microdialysate samples were pooled from individual animals and diluted (1:2 dilution in 2% nitric acid) to a final volume of 60 μl prior to metal analysis. Bovine liver (NBS Standard Reference Material, USDC, Washington, DC) (10 μg Mn/g; 184 μg Fe/g) was digested in ultrapure nitric acid and used as an internal standard for analysis (final concentration 5 μg Mn/L; 92 μg Fe/L).
Protein extraction
Protein was extracted from the brain tissue samples for Western blot analysis from animals that did not undergo surgery to ensure no changes in expression due to surgery. Tissue samples were sonicated in 500 μl of RIPA lysis buffer (1% Nonidet 40, 1% SDS, 0.5% sodium deoxycholate, 1 mM NaF, 2 mM β-glycerolphosphate, 1 mM sodium orthovanadate, and 1X protease inhibitor cocktail (Sigma, St. Louis, MO) in 1X PBS) on ice until completely homogenized. Homogenates were incubated on ice for 20 minutes before being centrifuged at 12,000 × g for 20 minutes at 4°C. Supernatant was then transferred to new tubes and total protein concentration determined by BCA assay (Pierce, Rockford, IL) before proceeding with Western analysis.
Western blot analysis
Western blot analysis was conducted to examine the effects of the dietary treatment on expression of GAT-1, GABAA, and GABAB proteins in vivo. Protein samples (20 μg) were combined with 4X LDS sample buffer (Invitrogen, Carlsbad, CA) containing 5% β-mercaptoethanol and heated at 70°C in a heat block for 10 minutes. Samples were then loaded onto a 4–12% Bis-Tris pre-cast mini gel (Invitrogen, Carlsbad, CA) and electrophoretically separated under denaturing conditions in 1X MOPS buffer containing 1% antioxidant (Invitrogen, Carlsbad, CA). Proteins were transferred to a PVDF membrane (Millipore, Billerica, MA) before blocking with 5% BSA. Membranes were probed overnight at 4°C with primary antibody (mouse monoclonal anti-β-actin, Santa Cruz Biotech, Santa Cruz, CA) (rabbit polyclonal anti-GABAB(R2) subunit; rabbit polyclonal anti-GAT-1, C-terminus; rabbit polyclonal anti-GABAA(α1); Chemicon, Temecula, CA) for the protein of interest in 5% BSA. Membranes were rinsed in 1X TBST (10 mM Tris, pH 7.4, 150 mM NaCl, 0.05% Tween 20) and probed for 2 hours at room temperature with an HRP-conjugated secondary antibody (goat anti-rabbit, Chemicon, Temecula, CA) (goat anti-mouse, Santa Cruz Biotech, Santa Cruz, CA) in 5% BSA. Membranes were then rinsed several times in 1X TBST before incubation in ECL solution (Perkin Elmer, Waltham, MA) and exposure to radiographic film (Pierce, Rockford, IL). Membranes were also probed for β-actin to verify equal loading and for image analysis. This housekeeping protein was chosen for comparison over GAPDH, which has been shown to be affected by Mn concentrations (Hazell, 2002). Films were analyzed using image analysis software (Image J, NIH, Bethesda, MD), with the amount of the target protein from each sample standardized to the amount of β-actin from the sample.
RNA isolation and cDNA synthesis
Total RNA was isolated from brain regions for quantitative PCR analysis from animals that did not undergo surgery to ensure no changes in mRNA expression as a result of surgery. Tissue samples were stored in 1 ml of RNAlater® solution (Ambion Inc., Austin, TX) and stored at −20° C until analysis. Total RNA was isolated utilizing the ToTALLY RNA™ kit (Ambion Inc., Austin, TX) following manufacturer’s instructions. RNA concentration and purity were determined by spectrophotometric analysis before carrying out cDNA synthesis. Synthesis of cDNA from total RNA was performed using the High Capacity cDNA Reverse Transcriptase Kit (Applied Biosystems, Foster City, CA) following manufacturer’s instructions.
Quantitative PCR
Quantitative real-time PCR analysis was utilized to determine expression of GAT-1, GABAA, and GABAB. Triplicate aliquots of cDNA were analyzed on 96-well plates using expression assays for the genes of interest obtained from Applied Biosystems (Foster City, CA). Values of cDNA expression were normalized relative to the expression of β-actin analyzed from the same sample on the same plate and reported as percent of control.
Statistical analysis
Statistical analyses were conducted using SPSS v14. Data were examined for normality of distribution using a one-sample Kolmogorov-Smirnov test and presence of outliers by boxplot analysis. Data were analyzed using analysis of variance, with Dunnett’s post-hoc analysis conducted to assess difference from controls when p<0.05. Pearson’s correlational analyses were conducted to examine relationships between extracellular concentrations of GABA, Mn, and Fe.
Results
Microdialysis analysis
GABA concentrations were increased in CNMn and ID versus CN (Figure 1A). For analysis, we excluded the IDMn group because only two animals survived surgery. Though the difference in GABA concentrations was not statistically significant (p=0.06), the observation does denote an obvious biological response to the dietary protocol. Concentrations of Mn in dialysate samples were significantly increased (p<0.001) in CNMn and ID versus CN (Figure 1B). No correlation was found between the extracellular concentrations of GABA and Mn (data not shown). Concentrations of Fe in dialysate samples were significantly decreased (p=0.023) in CNMn and ID versus CN (Figure 1C). An inverse correlation was observed between the extracellular concentrations of GABA and Fe, with GABA concentrations decreasing with increased Fe (Figure 1C, inset), though the correlation was not statistically significant (p=0.08). Being that there is a trend towards significance (p<0.10), it does suggest an effect of Fe levels on extracellular GABA concentrations. The statistical significance of this experiment is most likely affected by the small sample size. Only two animals from the IDMn group survived the surgery, with microdialysis samples being collected. Of the other three treatment groups, samples were collected from four out of the six animals. This is not uncommon given the 50% success rate associated with this procedure (Nelson et al., 1997), due either to animals succumbing to surgery or removing the cannula before sample collection.
Figure 1.
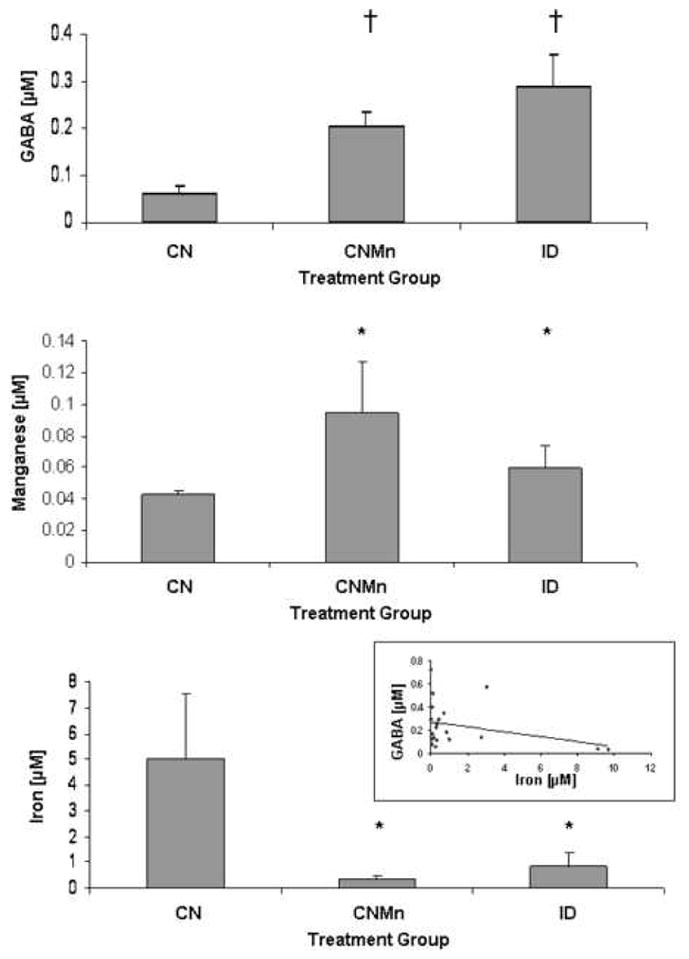
Microdialysate analysis. Mean concentrations ± SEM are shown for (A) extracellular GABA, (B) manganese, and (C) iron in microdialysate samples from the striatum after six weeks of dietary treatment. Inset: Correlational analysis of extracellular Fe and GABA concentrations (R= −0.32; p=0.08). *p<0.001; †p<0.10
Plasma manganese and iron
Plasma Mn concentrations were significantly increased in those animals receiving Mn supplementation versus those animals receiving deionized water alone (Figure 2A) (p=0.02). A significant decrease in plasma Fe concentrations was observed in animals receiving the ID diet versus the CN diet (Figure 2B) (p=0.007).
Figure 2.
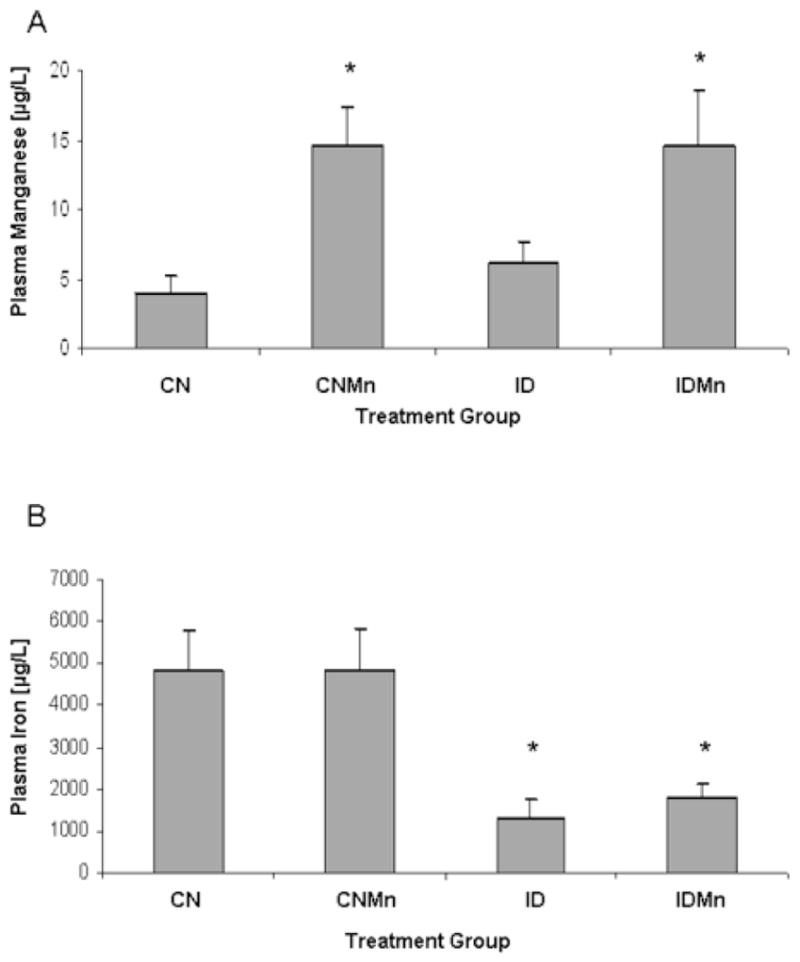
Plasma metal concentrations at six weeks. (A) Plasma manganese concentrations were significantly increased in those animals receiving manganese supplementation versus those animals receiving deionized water alone (p=0.02). (B) A significant decrease in plasma iron concentration was observed in animals receiving the ID diet versus the CN diet (p=0.007).
Brain manganese and iron
In all brain regions, Mn concentration was significantly increased in the Mn-exposure group versus those animals that did not receive Mn supplementation (Figure 3A) (p<0.05). A heterogeneous response was observed in the brain in regard to Fe levels, with a general decrease in Fe concentration in those animals receiving the ID diet versus animals receiving the CN diet, though the only significant decrease was seen in the globus pallidus (Figure 3B) (p<0.05).
Figure 3.
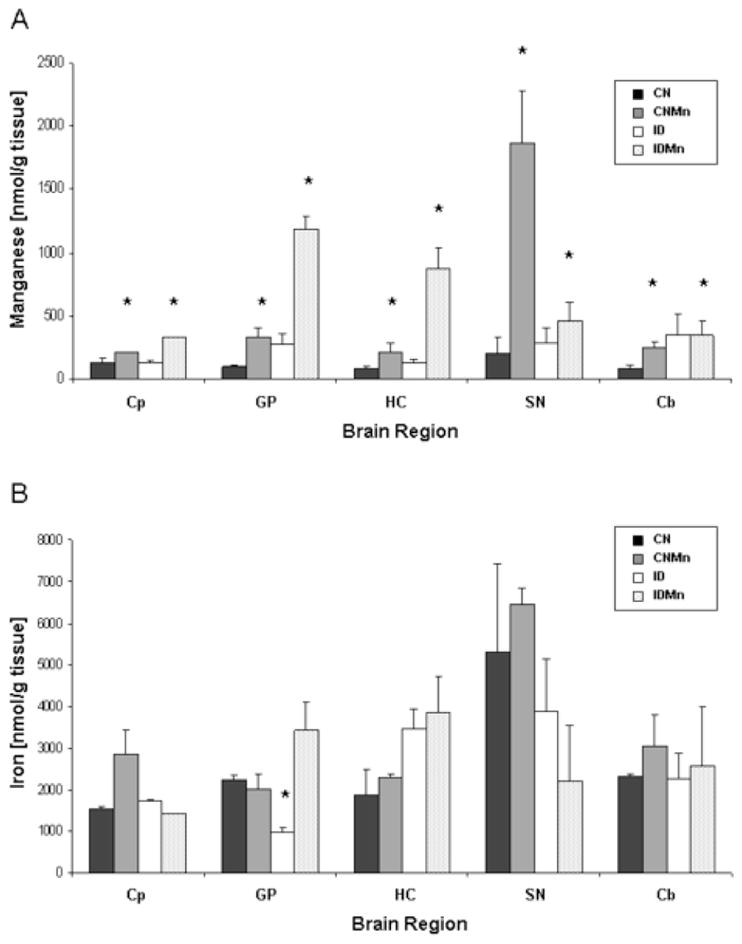
Brain metal concentrations at six weeks. Mean concentrations ± SEM are shown for manganese (A) and iron (B) for caudate putamen (Cp), globus pallidus (GP), hippocampus (HC), substantia nigra (SN), and cerebellum (Cb). CN is represented in black, CNMn in gray, ID in white, and IDMn in dotted area. *p<0.05 according to Dunnet’s post-hoc analysis
Western blot analysis
Elevated brain Mn and ID caused an approximate 50% decrease in GAT-1 protein expression in both the hippocampus and substantia nigra (Figure 4A). In the hippocampus, GAT-1 protein expression was significantly decreased in CNMn (41%) and ID (47%) versus CN (p<0.05). Similarly, in the substantia nigra, there was a significant decrease in expression of GAT-1 in CNMn (68%), ID (48%), and IDMn (44%) versus CN (p=0.03). No significant change in GAT-1 protein expression was observed in the globus pallidus, caudate putamen, or cerebellum. The dietary treatment had no effect on β-actin protein levels. Representative blots for GAT-1 and β-actin for each region are shown in Figure 4C, with each band representing an individual animal.
Figure 4.
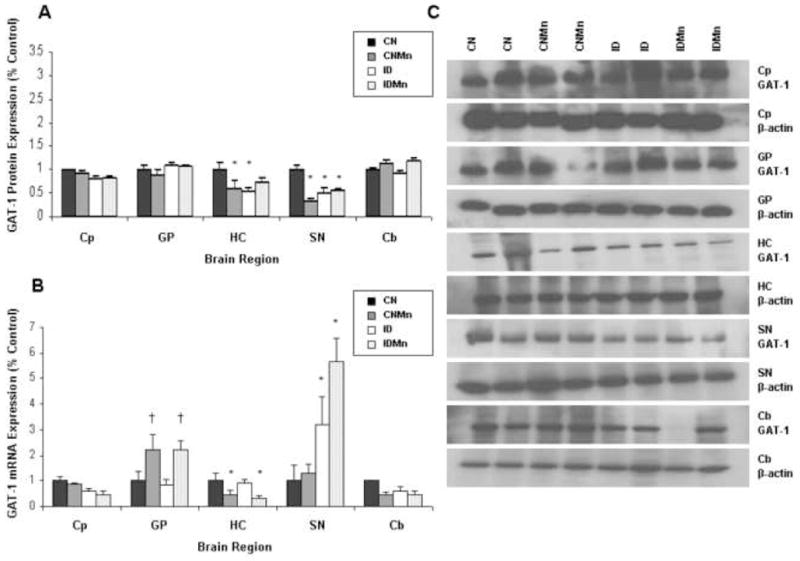
Effect of dietary treatment on GAT-1 protein and mRNA expression. Mean expression as percentage of control ± SEM for GAT-1 (A) protein and (B) mRNA relative to β-actin are shown for caudate putamen (Cp), globus pallidus (GP), hippocampus (HC), substantia nigra (SN), and cerebellum (Cb). CN is represented in black, CNMn in gray, ID in white, and IDMn in dotted area. (C) Representative blots for GAT-1 and β-actin for each region are shown, with each band representing an individual animal. *p<0.05 according to Dunnet’s post-hoc analysis
Increased Mn and ID were associated with a significant increase in protein expression of GABAA in globus pallidus (CNMn 75%; ID 300%; IDMn 300%; p=0.012) and substantia nigra (ID 200%; IDMn 200%; p=0.045) versus CN (Figure 5A). GABAA expression was significantly increased in hippocampus in CNMn (72%) versus CN (p=0.03) (Figure 5A). No significant difference in GABAA protein expression was observed in caudate putamen or cerebellum. Representative blots for GABAA and β-actin for each region are shown in Figure 5C, with each band representing an individual animal.
Figure 5.
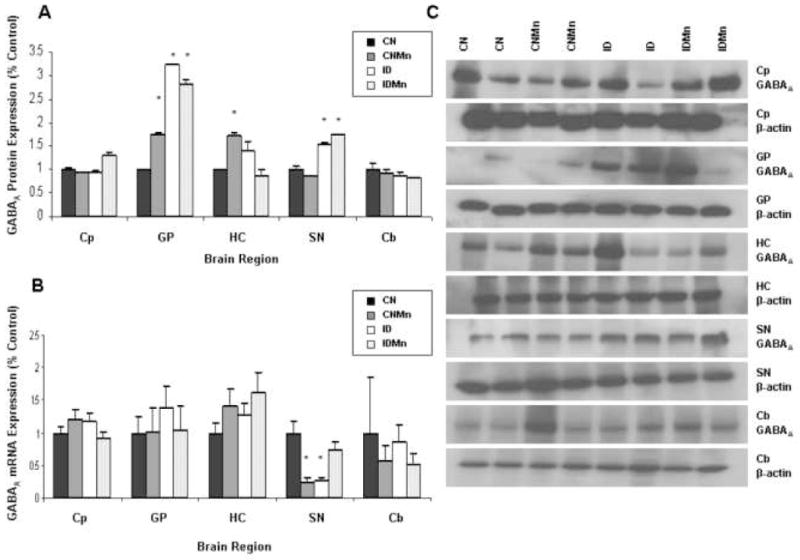
Effect of dietary treatment on GABAA protein and mRNA expression. Mean expression as percentage of control ± SEM for GABAA (A) protein and (B) mRNA relative to β-actin are shown for caudate putamen (Cp), globus pallidus (GP), hippocampus (HC), substantia nigra (SN), and cerebellum (Cb). CN is represented in black, CNMn in gray, ID in white, and IDMn in dotted area. (C) Representative blots for GABAA and β-actin for each region are shown, with each band representing an individual animal. *p<0.05 according to Dunnet’s post-hoc analysis
Iron deficiency and Mn exposure were associated with a decrease in GABAB protein in globus pallidus (IDMn 46%; p=0.03), substantia nigra (ID 41%; IDMn 54%; p=0.02), and hippocampus (ID 70%; IDMn 55%; p=0.027) (Figure 6A). No significant change in GABAB protein expression was observed in caudate putamen or cerebellum. Representative blots for GABAB and β-actin for each region are shown in Figure 6C, with each band representing an individual animal.
Figure 6.

Effect of dietary treatment on GABAB protein and mRNA expression. Mean expression as percentage of control ± SEM for GABAB (A) protein and (B) mRNA relative to β-actin are shown for caudate putamen (Cp), globus pallidus (GP), hippocampus (HC), substantia nigra (SN), and cerebellum (Cb). CN is represented in black, CNMn in gray, ID in white, and IDMn in dotted area. (C) Representative blots for GABAB and β-actin for each region are shown, with each band representing an individual animal. *p<0.05 according to Dunnet’s post-hoc analysis
Quantitative PCR analysis
Increased Mn significantly decreased mRNA expression of GAT-1 in the hippocampus (CNMn 52%; IDMn 67%; p=0.023) and caudate putamen (IDMn 54%; p=0.020) (Figure 4B). In the substantia nigra, mRNA expression of GAT-1 was significantly increased three-fold in ID and over five-fold in IDMn versus CN (p=0.003) (Figure 4B). A two-fold increase in GAT-1 expression was observed in the globus pallidus in the Mn-exposed groups versus CN (Figure 4B). This increase, though not statistically significant (p=0.080), does indicate a physiological response to the presence of Mn. The dietary treatment had no effect on β-actin mRNA levels.
In substantia nigra, GABAA mRNA expression was significantly decreased in CNMn (67%) and ID (64%) versus CN (p=0.006) (Figure 5B). No significant changes in mRNA expression of GABAA were observed in any of the other regions analyzed (Figure 5B). Expression of GABAB receptor mRNA was significantly decreased in the hippocampus (CNMn 59%; ID 45%; IDMn 43%; p=0.04), substantia nigra (CNMn 92%; ID 90%; IDMn 94%; p<0.001) and caudate putamen (CNMn 55%; IDMn 57%; p=0.003) versus CN (Figure 6B). No significant changes in mRNA expression were observed in globus pallidus, substantia nigra, or cerebellum.
Discussion
We report for the first time that increased dietary Mn (via drinking water; 100 times more concentrated than in the diet) causes elevated extracellular GABA and altered expression of GABA receptors and transporters, at both the protein and mRNA level. Extracellular GABA concentrations increased in response to Mn exposure and the ID diet (Figure 1A). Correlational analyses revealed an inverse correlation between extracellular GABA and Fe concentrations (Figure 1C, inset) with no correlation between extracellular GABA and Mn, suggesting GABA concentrations are altered as a result of the lowered extracellular Fe caused by Mn exposure and the ID diet. In addition, dietary Mn exposure was shown to alter expression of GABA receptor and transport proteins and mRNA in vivo, with a varied effect observed across the brain regions examined (Figures 4, 5, and 6). These data suggest that altered tissue levels of GABA (Erikson et al., 2002) and decreased uptake of GABA (Anderson et al., 2007a) due to Mn exposure may be the result of altered expression of its transport protein resulting in elevated extracellular GABA concentrations.
As in earlier studies (Anderson et al., 2007a; 2007b), our dietary protocol led to perturbations in systemic levels of Mn and Fe. The addition of Mn in the drinking water significantly increased the levels of Mn in the plasma (Figure 2A), while the ID diet lowered the plasma concentration of Fe (Figure 2B). In the brain, Mn accumulated in a diverse fashion across the regions examined, with increased levels of Mn found in those animals receiving Mn supplementation via the drinking water (Figure 3A). In general, Fe was depleted in those animals receiving the ID diet; however, the only statistical difference in Fe levels in the brain was observed in the globus pallidus (Figure 3B) similar to a previous study from our lab (Erikson et al., 2004). This varied response in Fe levels was observed in our previous studies, at both four weeks (Anderson et al., 2007b) and six weeks (Anderson et al., 2007a) of dietary treatment, with Fe levels varying between and within regions. Additionally, when examining the Fe:Mn ratio in the brain, a reduction in brain Fe levels in these animals is apparent, with the ID diet causing a significant reduction in the Fe:Mn ratio in most regions (data not shown).
Extracellular GABA concentrations in the caudate putamen were increased in response to Mn exposure and ID (Figure 1A); and while this difference was not statistically significant (p=0.06), it does indicate a significant physiological response to changes in regional metal levels (e.g., high Mn=3 fold increase; low Fe=4 fold increase compared to CN). An important question arose from these findings; is the elevated GABA due to increased extracellular Mn or decreased extracellular Fe? To answer this question, we measured both Mn and Fe concentrations in the microdialysis samples and correlated them with the GABA concentrations. Manganese levels were significantly increased in those animals receiving Mn supplementation and the ID diet (Figure 1B); however, no correlation was found between extracellular Mn and GABA concentrations, contrary to our original hypothesis. Concentrations of extracellular Fe were significantly decreased in those animals receiving the ID diet as well as in the CNMn animals versus CN (Figure 1C). This finding is interesting given that plasma Fe levels in the CNMn animals were normal. However, the Fe found in the plasma represents a more labile pool than that found in the CSF. An inverse correlation between extracellular GABA and Fe concentrations was also observed, with GABA concentration increasing with decreased Fe, suggesting that decreased Fe (due to both dietary deficiency and Mn-exposure) affects the levels of extracellular GABA. Iron deficiency affects GABA synthesis via glutamate decarboxylase (GAD) (Li, 1998) and a reduction in GABA brain tissue concentrations has been shown in rats fed a marginally ID diet for eight weeks (Shukla et al., 1989). Low tissue neurotransmitter levels may be the result of decreased uptake of the respective neurotransmitter (Gainetdinov et al., 1998), increasing the extracellular concentrations, a scenario which might be occurring in the current study.
The observed increase in extracellular GABA concentrations could potentially mediate the locomotor effects seen in Mn neurotoxicity, such as hyperkinesia, ataxia, and dystonia. GABAergic neurons in the striatum receive dopaminergic terminals from the substantia nigra (Smith and Bolam, 1990), in turn modulating the dopaminergic functioning in the striatum (Galindo et al., 1999). Increased extracellular levels of GABA in the striatum could affect the activity of the GABA projection neurons to the substantia nigra (Koós et al., 1999), leading to dysregulation of striatal DA release via the nigrostriatal pathway and increased inhibition, causing hypokinetic activity, a hallmark of Mn neurotoxicity (Figure 7). Normandin et al. (2004) observed decreases in locomotor activity in Mn-exposed young adult rats, while motor deficits were also observed in a study utilizing a pre-Parkinsonian rat model and cumulative low-dose Mn exposure (Gwiazda et al., 2002). In both of these studies, DA was not altered, suggesting that changes in GABA may precede and facilitate changes in DA during manganism.
Figure 7.
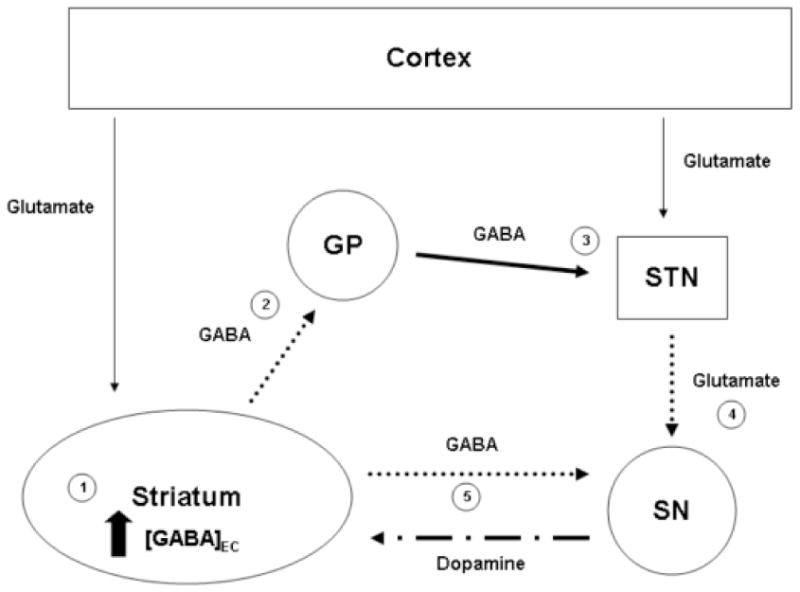
GABA biology during Mn overload. This simple schematic of the basal ganglia represents the potential consequences of the increased extracellular GABA concentrations in the striatum due to alterations of Mn and Fe status observed in the current study. (1) Increased extracellular GABA concentrations in the striatum would reduce the activity of the GABA striatopallidal projection neurons (2) (dotted line). This reduction in activity would (3) increase the GABAergic inhibitory firing from the globus pallidus (GP) to the subthalamic nucleus (STN) (heavy black line), in turn (4) decreasing the excitatory glutamatergic firing from this region to the substantia nigra (SN) (dotted line). (5) Decreased glutamatergic excitation in the substantia nigra, along with decreased GABAergic inhibition from the striatonigral projection neurons (dotted line) and decreased protein expression of GAT-1 and GABAB, would lead to a dysregulation of dopaminergic firing to the striatum (alternating line). This decrease in GABAergic firing to the substantia nigra may also contribute to the dopaminergic alterations observed during ID (Nelson et al., 1997; Erikson et al., 2000; 2001).
A major component of our hypothesis is that increased extracellular GABA due to Mn-exposure is caused by alterations in GAT-1. While protein expression of GAT-1 was not decreased in caudate putamen, mRNA expression was decreased (Figure 4B), indicating modulation of GAT-1 and a possible transition occurring in that region. Expression of GAT-1 mRNA was significantly decreased in hippocampus as well, while a two-fold increase was observed in the globus pallidus and a three and five-fold increase observed in substantia nigra (Figure 4B). While the two-fold increase in GAT-1 mRNA expression in globus pallidus was not statistically significant (p=0.08), it does indicate a physiological response to the dietary protocol within this region, perhaps as a mechanism to maintain adequate levels of GAT-1 protein. It should be noted that the current results were obtained after six weeks of dietary treatment. However, this represents only a snapshot of the modification occurring in the globus pallidus over the course of the treatment period as a result of increased Mn. Protein levels of GAT-1 in the globus pallidus may have been attenuated earlier than six weeks, with the increases in mRNA observed at six weeks the result of a compensatory mechanism. In our previous study, decreases in 3H-GABA uptake were observed after four weeks of dietary treatment in relation to Fe levels(Anderson et al., 2007a), indicating the likelihood of perturbations in GAT-1 at an earlier time. The increase seen in mRNA expression of GAT-1 in substantia nigra as a result of Mn exposure could potentially be a physiological response to compensate for decreases in GAT-1 protein in that region by increasing transcription. These changes in GAT-1 expression in the substantia nigra and globus pallidus could affect extracellular concentrations of GABA in the caudate putamen via nigrostriatal projections (Figure 7).
Decreased protein expression of GAT-1 in hippocampus and substantia nigra may also be the result of protein degradation and recycling via protein kinase C (PKC). Protein kinase C is known to regulate GAT-1 via phosphorylation (Mandela and Ordway, 2006), causing internalization of these proteins, degradation, and recycling (Gadea and López-Colomé, 2001). An acute Mn treatment (300 μM) has been shown to activate caspase-3 leading to activation of PKCδ in N27 mesenchephalic cells (Latchoumycandane et al., 2005). A similar response was seen in those cells incubated for 72 hours with a lower dose of Mn (50 μM), replicating chronic exposure. This 300 μM dose represents a physiologically relevant dose similar to that achieved in the current study. Additionally, expression of GAT-1 may be inhibited through effects on pre-synaptic GABAB autoreceptors, which can play a part in regulating expression of GAT-1 (Zahniser and Doolen, 2001). Decreases in these GABAB autoreceptors could result in a general decrease of GAT-1 density in that region.
As anticipated, GABAB protein expression was significantly decreased in globus pallidus, substantia nigra, and hippocampus in those animals receiving the ID diet and Mn supplementation (Figure 6A). Additionally, expression of GABAB mRNA was attenuated in the hippocampus, substantia nigra, and caudate putamen of the same animals (Figure 6B). While Mn and Fe are not known to have specific actions on the transcription of GABAB, Mn has been shown to bind to various forms of DNA structure, leading to conformational changes that might potentially affect the efficiency of gene transcription (Kennedy and Bryant, 1986). Protein expression of GABAB may also be attenuated through post-translational processes, such as recycling and degradation, mediated by regulatory kinases that may be affected by excess Mn. Decreases in both mRNA and protein expression of GABAB in hippocampus might indicate neuronal loss in that region as a result of Mn exposure (Sloot et al., 1994) as Mn has been shown to activate caspase-3 (Latchoumycandane et al., 2005), a mediator of apoptosis. A similar scenario might be the case in the substantia nigra.
Decreases in GABAB expression due to Mn exposure and increased extracellular GABA may result in marked changes in behavior, particularly related to anxiety-like behaviors. GABAB deficient mice have ‘panic’ attacks and are more anxious in anxiety related behavioral screens (Mombereau et al., 2005). Similarly, GABAB knockout mice display spontaneous seizures, hyperalgesia, hyperlocomotion, and memory impairments (Emson, 2007). Although both manganism and ID share many of these characteristics (Pal et al., 1999, Aschner et al., 2005) with these mice models, it should be noted that changes in GABAA receptors might play a role.
The change in mRNA expression in substantia nigra might represent interactions of Mn with DNA, causing a decrease in gene transcription efficiency (Hazell et al., 2003). The lack of changes in GABAA mRNA expression in the other regions suggests that the alterations observed in GABAA protein expression result from effects on post-translational processes. One possible mechanism for the increase seen in GABAA protein expression could be through increased trafficking of the receptor to the plasma membrane via phosphorylation of GABAA subunits by Akt (Wang et al., 2003) and since treatment of microglial cells with 500 μM Mn has been shown to activate Akt (Bae et al., 2006).
The changes in protein and mRNA expression observed in the hippocampus and substantia nigra are particularly interesting given these two regions were observed to have the most impact from our dietary protocol. The hippocampus plays a vital role in memory and learning, with the majority of the neurons present in the hippocampus GABAergic (Gadea and López-Colomé, 2001). Manganese has also been shown to generate lesions in the hippocampus (Sloot et al., 1994) and impaired memory and learning scores have been observed in children exposed to high levels of manganese from the drinking water (Wasserman et al., 2006). The dramatic changes in expression of protein and mRNA levels in the substantia nigra, a region association with Parkinson’s disease and a target region for Mn accumulation (Aschner et al., 2005), lend credence to the hypothesis of a role for GABA in the effects on DA in this region during manganism. Recall that GABA neurons project from the striatum to the substantia nigra, inhibiting the dopaminergic output from the substantia nigra (Figure 7). The differences in protein and mRNA expression observed in this study present a potential mechanism(s) to explain behaviors associated with Mn neurotoxicity.
The novel results of this study illustrate the complex mechanisms involved in Mn neurotoxicity. The alterations observed in protein and mRNA expression of GABA receptor and transport proteins were not universal, with a diverse effect of Mn exposure on these two indices not only from region to region, but, in some cases, within the same region. Future studies to tease out the pathways involved in these shifts in expression are warranted. These alterations in receptor and transporter expression are important in light of the effects observed in animal models lacking GAT-1 (tremor and anxiety) and GABAB (anxiety and nervousness) (Emson, 2007; Chiu et al., 2005; Mombereau et al., 2005), which are similar to symptoms of Mn neurotoxicity. Given the current findings, the increased levels of extracellular GABA in response to Mn exposure and ID are most likely due to decreased uptake of GABA rather than an increase in GABA release. Continuing studies in our lab will pharmacologically probe the effect of ID and Mn exposure on these extracellular GABA levels in order to fully understand the neurobiological consequences of Mn toxicity, especially as they relate to dietary exposures.
Acknowledgments
This research was supported by NIEHS Grant #ES013791-01 (KME) and Wake Forest University (CLC and TLW).
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Ade KK, Janssen MJ, Ortinski PI, Vicini S. Differential tonic GABA conductances in striatal medium spiny neurons. J Neurosci. 2008;28:1185–97. doi: 10.1523/JNEUROSCI.3908-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anderson JG, Cooney PT, Erikson KM. Brain manganese accumulation is inversely related to GABA uptake in male and female rats. Tox Sci. 2007a;95:188–95. doi: 10.1093/toxsci/kfl130. [DOI] [PubMed] [Google Scholar]
- Anderson JG, Cooney PT, Erikson KM. Inhibition of DAT function attenuates manganese accumulation in the globus pallidus. Environ Tox Pharmacol. 2007b;23:179–84. doi: 10.1016/j.etap.2006.08.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aschner M, Erikson KM, Dorman DC. Manganese dosimetry: species differences and implications for neurotoxicity. Crit Rev Tox. 2005;35:1–32. doi: 10.1080/10408440590905920. [DOI] [PubMed] [Google Scholar]
- Bae JH, Jang BC, Suh SI, Ha E, Baik HH, Kim SS, Lee MY, Shin DH. Manganese induces inducible nitric oxide synthase (iNOS) expression via activation of both MAP kinase and PI3K/Akt pathways in BV2 microglial cells. Neurosci Lett. 2006;398:151–4. doi: 10.1016/j.neulet.2005.12.067. [DOI] [PubMed] [Google Scholar]
- Beard JL. Iron biology in immune function, muscle metabolism, and neuronal functioning. J Nutr. 2001;131:568s–79s. doi: 10.1093/jn/131.2.568S. [DOI] [PubMed] [Google Scholar]
- Burdo JR, Martin J, Menzies SL, Dolan KG, Romano MA, Fletcher RJ, Garrick MD, Garrick LM, Connor JR. Cellular distribution of iron in the brain of the Belgrade rat. Neuroscience. 1999;93:1189–96. doi: 10.1016/s0306-4522(99)00207-9. [DOI] [PubMed] [Google Scholar]
- Carlsson M, Carlsson A. Interactions between glutamatergic and monoaminergic systems within the basal ganglia-implications for schizophrenia and Parkinson’s disease. Trends Neurosci. 1990;13:272–76. doi: 10.1016/0166-2236(90)90108-m. [DOI] [PubMed] [Google Scholar]
- Chandra SV, Shukla GS. Role of iron deficiency in inducing susceptibility to manganese toxicity. Arch Toxicol. 1976;35:319–23. doi: 10.1007/BF00570272. [DOI] [PubMed] [Google Scholar]
- Chen Z, Wu j, Baker GB, Parent M, Dovichi NJ. Application of capillary electrophoresis with laser-induced fluorescence detection to the determinations of biogenic amines and amino acids in brain microdialysate and homogenate samples. J Chromatography A. 2001;914:293–98. doi: 10.1016/s0021-9673(01)00539-8. [DOI] [PubMed] [Google Scholar]
- Chiu CS, Brickley S, Jensen K, Southwell A, McKinney S, Cull-Candy S, Mody I, Lester HA. GABA transporter deficiency causes tremor, ataxia, nervousness, and increased GABA-induced tonic conductance in cerebellum. J Neurosci. 2005;25:3234–45. doi: 10.1523/JNEUROSCI.3364-04.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davis C, Wolf T, Greger J. Varying levels of manganese and iron affect absorption and gut endogenous losses of manganese by rats. J Nutr. 1992;122:1300–08. doi: 10.1093/jn/122.6.1300. [DOI] [PubMed] [Google Scholar]
- Dickinson SD, Sabeti J, Larson GA, Giardina K, Rubinstein M, Kelly MA, Grandy DK, Low MJ, Gerhardt GA, Zahniser NR. Dopamine D2 receptor-deficient mice exhibit decreased dopamine transporter function but no changes in dopamine release in dorsal striatum. J Neurochem. 1999;72:148–56. doi: 10.1046/j.1471-4159.1999.0720148.x. [DOI] [PubMed] [Google Scholar]
- Emson PC. GABAB receptors: structure and function. Prog Brain Res. 2007;160:43–57. doi: 10.1016/S0079-6123(06)60004-6. [DOI] [PubMed] [Google Scholar]
- Erikson KM, John CE, Jones SR, Aschner M. Manganese accumulation in striatum of mice exposed to toxic doses is dependent upon a functional dopamine transporter. Environ Toxicol Pharmacol. 2005;20 :390–94. doi: 10.1016/j.etap.2005.03.009. [DOI] [PubMed] [Google Scholar]
- Erikson KM, Syversen T, Steinnes E, Aschner M. Globus pallidus: a target brain region for divalent metal accumulation associated with dietary iron deficiency. J Nutr Biochem. 2004;15:335–41. doi: 10.1016/j.jnutbio.2003.12.006. [DOI] [PubMed] [Google Scholar]
- Erikson KM, Shihabi ZK, Aschner JL, Aschner M. Manganese accumulates in iron-deficient rat brain regions in a heterogeneous fashion and is associated with neurochemical alterations. Biolog Trace Elem Res. 2002;87:143–56. doi: 10.1385/BTER:87:1-3:143. [DOI] [PubMed] [Google Scholar]
- Erikson KM, Jones BC, Hess EJ, Zhang Q, Beard JL. Iron deficiency decreases dopamine D1 and D2 receptors in rat brain. Pharmacol Biochem Behav. 2001;69:409–18. doi: 10.1016/s0091-3057(01)00563-9. [DOI] [PubMed] [Google Scholar]
- Erikson KM, Jones BC, Beard JL. Iron deficiency alters dopamine transporter functioning in rat striatum. J Nutr. 2000;130:2831–37. doi: 10.1093/jn/130.11.2831. [DOI] [PubMed] [Google Scholar]
- Expósito I, Del Arco A, Segovia G, Mora F. Endogenous dopamine increases extracellular concentrations of glutamate and GABA in striatum of the freely moving rat: involvement of D1 and D2 dopamine receptors. Neurochem Res. 1999;24:849–56. doi: 10.1023/a:1020901929419. [DOI] [PubMed] [Google Scholar]
- Finley JW. Manganese absorption and retention by young women is associated with serum ferritin concentration. Am J Clin Nutr. 1999;70:37–43. doi: 10.1093/ajcn/70.1.37. [DOI] [PubMed] [Google Scholar]
- Gadea A, López-Colomé AM. Glial transporters for glutamate, glycine, and GABA: II. GABA transporters. J Neurosci Res. 2001;63:461–68. doi: 10.1002/jnr.1040. [DOI] [PubMed] [Google Scholar]
- Gainetdinov RR, Jones SR, Fumagalli F, Wightman RM, Caron MG. Re-evaluation of the role of the dopamine transporter in dopamine system homeostasis. Brain Res Rev. 1998;26:148–53. doi: 10.1016/s0165-0173(97)00063-5. [DOI] [PubMed] [Google Scholar]
- Galindo A, Del Arco A, Mora F. Endogenous GABA potentiates the potassium-induced release of dopamine in striatum of the freely moving rat: a microdialysis study. Brain Res Bull. 1999;50:209–14. doi: 10.1016/s0361-9230(99)00199-9. [DOI] [PubMed] [Google Scholar]
- Garcia SJ, Gellein K, Syversen T, Aschner M. A manganese enhanced diet alters brain metals and transporters in the developing rat. Tox Sci. 2006;92:516–25. doi: 10.1093/toxsci/kfl017. [DOI] [PubMed] [Google Scholar]
- Garcia SJ, Gellein K, Syversen T, Aschner M. Iron deficient and manganese supplemented diets alter metals and transporters in the developing rat brain. Tox Sci. 2007;95:205–14. doi: 10.1093/toxsci/kfl139. [DOI] [PubMed] [Google Scholar]
- Garrick MD, Dolan KG, Horbinski C, Ghio AJ, Higgins D, Porubcin M, Moore EG, Hainsworth LN, Umbreit JN, Conrad ME, Feng L, Lis A, Roth JA, Singleton S, Garrick LM. DMT1: a mammalian transporter for multiple metals. BioMetals. 2003;16:41–54. doi: 10.1023/a:1020702213099. [DOI] [PubMed] [Google Scholar]
- Gether U, Andersen PH, Larsson EM, Schousboe A. Neurotransmitter transporters: molecular function of important drug targets. Trends Pharmacol Sci. 2006;27:375–83. doi: 10.1016/j.tips.2006.05.003. [DOI] [PubMed] [Google Scholar]
- Gunshin H, Allerson CR, Polycarpou-Schwarz M, Rofts A, Rogers JT, Kishi F, Hentze MW, Rouault TA, Andrews NC, Hediger MA. Iron-dependent regulation of the divalent metal ion transporter. FEBS Lett. 2001;509:309–16. doi: 10.1016/s0014-5793(01)03189-1. [DOI] [PubMed] [Google Scholar]
- Gunshin H, Mackenzie B, Berger UV, Gunshin Y, Romero MF, Boron WF, Nussberger S, Gollan JL, Hediger MA. Cloning and characterization of a mammalian proton-coupled metal transporter. Nature. 1997;388:482–88. doi: 10.1038/41343. [DOI] [PubMed] [Google Scholar]
- Gwiazda RH, Lee D, Sheridan J, Smith DR. Low cumulative manganese exposure affects striatal GABA but not dopamine. Neurotoxicology. 2002;23:69–76. doi: 10.1016/s0161-813x(02)00002-5. [DOI] [PubMed] [Google Scholar]
- Hazell AS, Normandin L, Nguyen B, Kennedy G. Upregulation of ‘peripheral-type’ benzodiazepine receptors in the globus pallidus in a sub-acute rat model of manganese neurotoxicity. Neurosci Lett. 2003;349:13–6. doi: 10.1016/s0304-3940(03)00649-9. [DOI] [PubMed] [Google Scholar]
- Hazell AS. Astrocytes and manganese neurotoxicity. Neurochem Int. 2002;41:271–7. doi: 10.1016/s0197-0186(02)00013-x. [DOI] [PubMed] [Google Scholar]
- Hazell AS, Desjardins P, Butterworth RF. Chronic exposure of rat primary astrocyte cultures to manganese results in increased binding sites for the ‘peripheral-type’ benzodiazepine receptor ligand 3H-PK 11195. Neurosci Lett. 1999;271:5–8. doi: 10.1016/s0304-3940(99)00489-9. [DOI] [PubMed] [Google Scholar]
- Hurley LS, Keen CL. Manganese. In: Underwood E, Mertz W, editors. Trace Elements in Human Health and Animal Nutrition. Academic Press; New York, NY: 1987. pp. 185–223. [Google Scholar]
- Ingersoll RT, Montgomery EB, Jr, Aposhian HV. Central nervous system toxicity of manganese. II: cocaine or reserpine inhibit manganese concentration in the rat brain. Neurotoxicology. 1999;20:467–76. [PubMed] [Google Scholar]
- Kennedy SD, Bryant RG. Manganese-deoxyribonucleic acid binding modes. Nuclear magnetic relaxation dispersion results. Biophys J. 1986;50:669–76. doi: 10.1016/S0006-3495(86)83507-X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kerr DIB, Ong J. GABAB receptors. Pharmac Ther. 1995;67:187–246. doi: 10.1016/0163-7258(95)00016-a. [DOI] [PubMed] [Google Scholar]
- Kondakis XG, Makris N, Leotsinidis M, Prinou M, Papapetropoulos T. Possible health effects of high manganese concentration in drinking water. Arch Environ Health. 1989;44:175–78. doi: 10.1080/00039896.1989.9935883. [DOI] [PubMed] [Google Scholar]
- Koós T, Tepper JM. Inhibitory control of neostriatal projection neurons by GABAergic interneurons. Nat Neurosci. 1999;2:467–72. doi: 10.1038/8138. [DOI] [PubMed] [Google Scholar]
- Kwik-Uribe CL, Golub MS, Keen CL. Chronic marginal iron intakes during early development in mice alter brain iron concentrations and behavior despite postnatal iron supplementation. J Nutr. 2000;130:2040–48. doi: 10.1093/jn/130.8.2040. [DOI] [PubMed] [Google Scholar]
- Latchoumycandane C, Anantharam V, Kitazawa M, Yang Y, Kanthasamy A, Kanthasamy AG. Protein kinase Cδ is a key downstream mediator of manganese-induced apoptosis in dopaminergic neuronal cells. J Pharmacol Exper Therap. 2005;313:46–55. doi: 10.1124/jpet.104.078469. [DOI] [PubMed] [Google Scholar]
- Li D. Effects of iron deficiency on iron distribution and γ-aminobutyric acid (GABA) metabolism in young rat brain tissues. Hokkaido J Med Sci. 1998;73:215–25. [PubMed] [Google Scholar]
- Mandela P, Ordway GA. The norepinephrine transporter and its regulation. J Neurochem. 2006;97:310–33. doi: 10.1111/j.1471-4159.2006.03717.x. [DOI] [PubMed] [Google Scholar]
- Mergler D, Huel G, Bowler R, Iregren A, Belanger S, Baldwin M, Tardif R, Smargiassi A, Martin L. Nervous system dysfunction among workers with long-term exposure to manganese. Environ Res. 1994;64:151–80. doi: 10.1006/enrs.1994.1013. [DOI] [PubMed] [Google Scholar]
- Mombereau C, Kaupman K, Gassman M, Bettler B, van der Putten H, Cryan JF. Altered anxiety and depression-related behavior in mice lacking GABAB(2) receptor subunits. Neuroreport. 2005;16:307–10. doi: 10.1097/00001756-200502280-00021. [DOI] [PubMed] [Google Scholar]
- Nelson C, Erikson KM, Piñero DJ, Beard JL. In vivo dopamine metabolism is altered in iron deficient anemic rats. J Nutr. 1997;127:2282–88. doi: 10.1093/jn/127.12.2282. [DOI] [PubMed] [Google Scholar]
- Normandin L, Beaupre LA, Salehi F, StPierre A, Kennedy G, Mergler D, Butterworth RF, Philippe S, Zayed J. Manganese distribution in the brain and neurobehavioral changes following inhalation exposure of rats to three chemical forms of manganese. Neurotoxicology. 2004;25:433–41. doi: 10.1016/j.neuro.2003.10.001. [DOI] [PubMed] [Google Scholar]
- Pal PK, Samii A, Calne DB. Manganese neurotoxicity: a review of clinical features, imaging and pathology. Neurotoxicology. 1999;20:227–38. [PubMed] [Google Scholar]
- Paxinos G, Watson C. The rat brain in stereotaxic coordinates. Academic Press Inc; San Diego, CA: 1998. [Google Scholar]
- Powell PR, Ewing AG. Recent advances in the application of capillary electrophoresis to neuroscience. Anal Bioanal Chem. 2005;382:581–91. doi: 10.1007/s00216-005-3075-x. [DOI] [PubMed] [Google Scholar]
- Roth JA, Garrick MD. Iron interactions and other biological reactions mediating the physiological and toxic actions of manganese. Biochem Physiol. 2003;66:1–13. doi: 10.1016/s0006-2952(03)00145-x. [DOI] [PubMed] [Google Scholar]
- Shukla A, Agarwal KN, Shukla GS. Latent iron deficiency alters gamma-aminobutyric acid and glutamate metabolism in rat brain. Experientia. 1989;45:343–5. doi: 10.1007/BF01957472. [DOI] [PubMed] [Google Scholar]
- Shukla GS, Chandra SV, Seth PK. Effect of manganese on the levels of DNA, RNA, DNase and RNAse in cerebrum, cerebellum and rest of the brain regions of rat. Acta Pharmacol Toxicol (Copenhagen) 1976;39:562–69. doi: 10.1111/j.1600-0773.1976.tb03206.x. [DOI] [PubMed] [Google Scholar]
- Sloot WN, van der Sluijs-Gelling AJ, Gramsbergen JBP. Selective lesions by manganese and extensive damage by iron after injection into rat striatum or hippocampus. J Neurochem. 1994;62:205–16. doi: 10.1046/j.1471-4159.1994.62010205.x. [DOI] [PubMed] [Google Scholar]
- Smith AD, Bolam JP. The neural network of the basal ganglia as revealed by the study of synaptic connections of identified neurons. Trends Neurosci. 1990;13:259–65. doi: 10.1016/0166-2236(90)90106-k. [DOI] [PubMed] [Google Scholar]
- Struve MF, McManus BE, Wong BA, Dorman DC. Basal ganglia neurotransmitter concentrations in Rhesus monkeys following subchronic manganese sulfate inhalation. Amer J Indust Med. 2007;50:772–8. doi: 10.1002/ajim.20489. [DOI] [PubMed] [Google Scholar]
- Vidal L, Alfonso M, Campos F, Faro LRF, Cervantes RC, Durán R. Effects of manganese on extracellular levels of dopamine in rat striatum: an analysis in vivo by brain microdialysis. Neurochem Res. 2005;30:1147–54. doi: 10.1007/s11064-005-7775-6. [DOI] [PubMed] [Google Scholar]
- Wasserman GA, Liu X, Parvez F, Ahsan H, Levy D, Factor-Litvak P, Kline J, van Geen A, Slavkovich V, Lolacono NJ, Cheng Z, Zheng Y, Graziano JH. Water manganese exposure and children’s intellectual function in Araihazar, Bangladesh. Environ Health Persp. 2006;114:124–9. doi: 10.1289/ehp.8030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zahniser NR, Doolen S. Chronic and acute regulation of Na+/Cl− -dependent neurotransmitter transporters: drugs, substrates, presynaptic receptors, and signaling systems. Pharmacol Therap. 2001;92:21–55. doi: 10.1016/s0163-7258(01)00158-9. [DOI] [PubMed] [Google Scholar]