

Abstract
The acoustic startle response is a protective response, elicited by a sudden and intense acoustic stimulus. Facial and skeletal muscles are activated within a few milliseconds, leading to a whole body flinch in rodents1. Although startle responses are reflexive responses that can be reliably elicited, they are not stereotypic. They can be modulated by emotions such as fear (fear potentiated startle) and joy (joy attenuated startle), by non-associative learning processes such as habituation and sensitization, and by other sensory stimuli through sensory gating processes (prepulse inhibition), turning startle responses into an excellent tool for assessing emotions, learning, and sensory gating, for review see 2, 3. The primary pathway mediating startle responses is very short and well described, qualifying startle also as an excellent model for studying the underlying mechanisms for behavioural plasticity on a cellular/molecular level3.
We here describe a method for assessing short-term habituation, long-term habituation and prepulse inhibition of acoustic startle responses in rodents. Habituation describes the decrease of the startle response magnitude upon repeated presentation of the same stimulus. Habituation within a testing session is called short-term habituation (STH) and is reversible upon a period of several minutes without stimulation. Habituation between testing sessions is called long-term habituation (LTH)4. Habituation is stimulus specific5. Prepulse inhibition is the attenuation of a startle response by a preceding non-startling sensory stimulus6. The interval between prepulse and startle stimulus can vary from 6 to up to 2000 ms. The prepulse can be any modality, however, acoustic prepulses are the most commonly used.
Habituation is a form of non-associative learning. It can also be viewed as a form of sensory filtering, since it reduces the organisms' response to a non-threatening stimulus. Prepulse inhibition (PPI) was originally developed in human neuropsychiatric research as an operational measure for sensory gating7. PPI deficits may represent the interface of "psychosis and cognition" as they seem to predict cognitive impairment8-10. Both habituation and PPI are disrupted in patients suffering from schizophrenia11, and PPI disruptions have shown to be, at least in some cases, amenable to treatment with mostly atypical antipsychotics12, 13. However, other mental and neurodegenerative diseases are also accompanied by disruption in habituation and/or PPI, such as autism spectrum disorders (slower habituation), obsessive compulsive disorder, Tourette's syndrome, Huntington's disease, Parkinson's disease, and Alzheimer's Disease (PPI)11, 14, 15 Dopamine induced PPI deficits are a commonly used animal model for the screening of antipsychotic drugs16, but PPI deficits can also be induced by many other psychomimetic drugs, environmental modifications and surgical procedures.
Protocol
1. Protocol Design
Calibration: Before a set of experiments, calibrate the loudspeakers. This is important so that loudspeaker display the exact volume that was set by the experimenter. Also calibrate the sensitivity of the transducer platform of the startle boxes according to the supplier's manual. The transducer converts the vertical movement of the platform into a voltage signal. Make sure that there are no ongoing experiments when calibrating the system, and that all boxes are calibrated the same way.
i/o function: If new strains of mice or rats are measured, an input/output function should be established. After an acclimation period of 5-10 minutes with a constant background white noise of 65 to 68 dB (see below), startle stimuli (20 ms white noise) should be displayed every 20 sec, starting at around 70-75 dB. Startle stimulus intensity will be increased between each stimulus by 2-5 dB until reaching 120-130 dB, resulting in 10-30 trials with startle stimuli (see figure 1).
Protocol structure: Habituation and prepulse inhibition can be measured within one protocol. The protocol is divided into an acclimation period, a block I (habituation), immediately followed by a block II (PPI, figure 2). Before measuring prepulse inhibition, animals should always undergo startle habituation, so that startle attenuations due to habituation do not interfere with PPI measurements.
Acclimation period: Each time an animal is tested, it first undergoes an acclimation phase in order to adapt to the animal holder, startle box and background noise. During a 5-10 minutes acclimation period, the constant background noise of 65-68dB white noise (depending on the noise of the environment) is displayed, but no startle stimuli. During this phase the animal will calm down, stop to explore the environment and stop moving around.
Block I habituation: For short-term habituation (STH), between 30 -100 startle stimuli should be applied on the background. Startle stimuli are commonly white noises of 20 ms duration and very steep rise times (0, if possible). The intensity is ideally at the volume where the i/o function reached the plateau of maximum startle response, commonly at 105 to 115 dB. The intervals between single trials should be either always 20 sec or randomized between 10 and 30 sec (see discussion and figure 3).
Block II prepulse inhibition: In order to measure PPI, trials with a startle pulse alone and trials with a prepulse are pseudorandomized in block II. Background noise and startle stimulus are the same as in block I. The prepulse is a white noise of 4 ms duration and also steep rise time. Two parameters can be varied: the interstimulus interval between prepulse and startle pulse and the intensity of the prepulse (see discussion). We propose to commonly use two different prepulse intensities (75 and 85 dB) and two different interstimulus intervals (30 ms and 100 ms). Thus there are four different prepulse-pulse trials plus the startle pulse alone trials to be pseudorandomized and displayed 10 times each, which totals 50 trials. Inter-trial intervals can be either 20 sec or randomized between 10 and 30 sec (see discussion). In some cases it might be beneficial to add a sixth type of trials, which is a prepulse alone trial (see discussion and figure 4).
Long-term habituation: In order to measure LTH, the entire protocol is run on at least five subsequent days. Alternatively, only the acclimation phase and block I could be run, however, in order to see LTH, block I should contain at least startle 100 stimuli. The presentation of 30 stimuli per day leads to very little or no LTH in most animals, especially in mice. Runs should be on approximately the same time of each day, since startle response amplitudes fluctuate with the diurnal cycle.
2. Handling and Acclimation of Animals
There are big differences in the handling and acclimation of rats versus mice. Mice will be placed into the appropriate animal holder (they should not be restrained) for 2-5 minutes with background noise but no startle stimuli (acclimation phase of the program). This procedure should be repeated 3-5 times, once or twice a day, until defecation and urination in the mouse holder ceases or considerably decreases. Animal holders should be always replaced or cleaned after an animal is removed.
Rats should be handled for at least three sessions17. At the end of the third handling sessions they are placed into an appropriate animal holder (no restrain) and exposed to background noise for several minutes. After removing them, they can be rewarded with sunflower seeds in order to form positive associations with the testing procedure. This procedure is repeated two more times, gradually expanding the acclimation time, before the entire protocol is run.
For testing sessions, animals are placed into all chambers, doors closed and the protocol with acclimation phase, block I and block II is run. If there are different groups of animals (injections, genotypes), they should be mixed or randomized over the different runs and the different boxes. If an animal is repeatedly tested (e.g. with different treatments), it should be re-tested in the same box. For repeated PPI testing in rats, we also recommend to run an entire protocol before the actual data collection takes place. PPI often improves between the first and the second testing session (PPI learning), and stays consistent thereafter. It will also eliminate a big portion of LTH.
3. Data Analysis
Short-term habituation: For short-term habituation analysis, all startle responses of block I are plotted for each animal. If animals within a group have similar startle response amplitudes, values can be averaged between animals. In most cases, however, absolute startle amplitudes differ considerably between animals and startle levels are not normally distributed. In this case it is more viable to normalize the data of each animal to its first, or the average of the first two, startle responses in block I (animals sometimes fall asleep during the acclimation phase resulting in a low first startle response and a high second startle). The normalized data can then be averaged across all animals in order to plot the course of habituation. For a quantitative assessment of the amount of habituation, a score can be calculated for each animal, e.g. the average of the last 10 startle responses divided by the average of the first two responses (figure 6).
Prepulse inhibition: For analyzing prepulse inhibition, the data of block II has to be sorted according to the type of trial (e.g. by exporting all relevant data columns into excel and sort by prepulse intensity and duration of ISI). The ten traces per trial type are then averaged, and the resulting values for the prepulse-pulse trials are divided by the startle pulse alone value and multiplied by 100. This reveals the amount of remaining startle (in percent of baseline startle) under different prepulse conditions for each animal. Baseline startle (pulse alone) is 100%. These values can then be averaged across animals of a group and be plotted (figure 7a). Alternatively, the amount of PPI can be plotted by subtracting the remaining startle response from 100% (figure 7b). Please be aware: when you compare PPI in different groups of animals, you should always also report whether there is a difference in baseline startle amplitudes, by e.g. comparing the absolute startle amplitudes of the startle pulse alone trials (or startle amplitudes in block I).
Long-term habituation: In order to analyze LTH the first two responses of block I of each day are averaged and plotted over the minimum of five consecutive testing sessions. This eliminates the possibility that differences in STH affect the result of LTH analysis. If it has been established that a treatment/gene does not affect STH, alternatively all responses in block I can be simply averaged for each day and be plotted. LTH can be quantified by calculation of a habituation score where the last days' value is divided by the first days' value and multiplied by 100, so that the percentage of initial startle level remaining after LTH is displayed. Habituation scores can then be averaged across animals (figure 8).
4. Representative Results:
i/o function: Rodents typically begin to startle from a volume of 85-90 dB on (with 20 ms duration, white noise). The startle response increases with increasing volume and normally reaches a maximum at 100-110 dB. If animals deviate considerably from these values, animals might have disrupted hearing abilities or motor abilities. Typical i/o functions are displayed in figure 5.
Short-term habituation: Well handled rats normally habituate to around 60% of their initial startle response; however, there are huge individual differences and also strain differences. The strongest habituation effect occurs normally within the first several stimuli. Mice do generally habituate less than rats (typically to about 80%), but strain differences can be very large. A typical habituation course is shown in figure 6.
Prepulse inhibition: Most rats show PPI of around 90% with an optimal prepulse (85dB, 4 ms, white noise). PPI is very robust and individual differences are relatively small with these experimental settings. Lower volume prepulses yield less PPI and more variability (even within an animal), but also seems to be more vulnerable to pharmacological or genetic manipulations. Different PPI results are plotted in figure 7.
Long-term habituation: Long-term habituation can be observed over several testing sessions. LTH is very robust in rats. In mice, it often requires the presentation of a lot of startle stimuli in each session in order to observe LTH. Typical LTH results can be seen in figure 8.
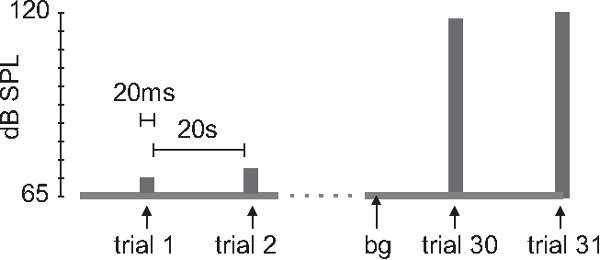 Figure 1.
Stimulus protocol for i/o function. After an acclimation period of 5-10 min. with 65 dB sound pressure level (SPL) background noise and no startle stimuli (not shown), 20 ms white noise stimuli are presented every 20 sec. The intensity gradually increases from 75 to 130 dB in 5 dB increments (bg = background noise).
Figure 1.
Stimulus protocol for i/o function. After an acclimation period of 5-10 min. with 65 dB sound pressure level (SPL) background noise and no startle stimuli (not shown), 20 ms white noise stimuli are presented every 20 sec. The intensity gradually increases from 75 to 130 dB in 5 dB increments (bg = background noise).
 Figure 2.
Protocol structure for combined habituation and PPI measurement. During the whole protocol, a constant background noise of 65 dB is applied. There is an acclimation period of 5-10 min. without any further stimulation. Immediately thereafter, habituation is tested by 30-100 startle stimuli (block I, see figure 3). This is immediately followed by PPI testing (block II, see figure 4).
Figure 2.
Protocol structure for combined habituation and PPI measurement. During the whole protocol, a constant background noise of 65 dB is applied. There is an acclimation period of 5-10 min. without any further stimulation. Immediately thereafter, habituation is tested by 30-100 startle stimuli (block I, see figure 3). This is immediately followed by PPI testing (block II, see figure 4).
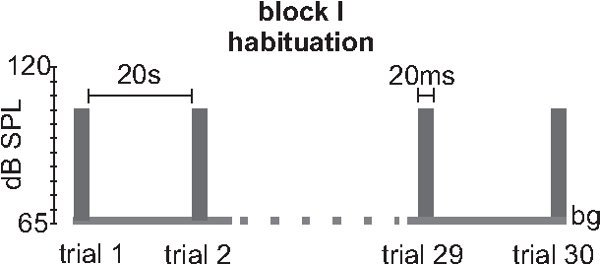 Figure 3.
Stimulus protocol for measuring habituation (block I). An example for a typical block I for testing short-term habituation is shown. It consists of 30 100 identical trials where a 20 ms 105 dB white noise with a 0 rise time is presented with an inter-trial interval (ITI) of 20 sec. Variations of this protocol may include higher startle stimulus intensities or variable ITIs
Figure 3.
Stimulus protocol for measuring habituation (block I). An example for a typical block I for testing short-term habituation is shown. It consists of 30 100 identical trials where a 20 ms 105 dB white noise with a 0 rise time is presented with an inter-trial interval (ITI) of 20 sec. Variations of this protocol may include higher startle stimulus intensities or variable ITIs
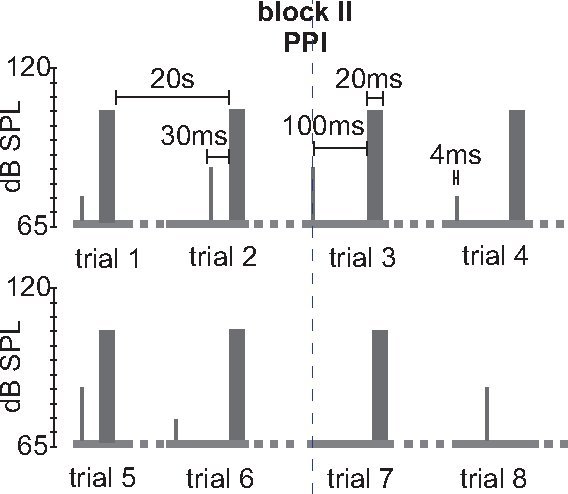 Figure 4.
Stimulus protocol for measuring PPI (block II). An example for a typical part of a block II for testing PPI is shown. Block II consists of 5-6 different trial types that are presented 10 times each in a pseudorandomized order. Here, two different prepulse intensities (75 dB and 85 dB) and two different interstimulus intervals (ISIs, 30 and 100 ms) are tested. Startle stimulus alone trials and prepulse alone trials are interspersed. This block would have 6x10= 60 trials. Prepulses are 4 ms white noise pulses with 0 rise time. Variations of this protocol would consist in variable ITIs, higher startle stimulus intensities, different prepulse intensities and/or durations, and different ISIs between prepulse and pulse.
Figure 4.
Stimulus protocol for measuring PPI (block II). An example for a typical part of a block II for testing PPI is shown. Block II consists of 5-6 different trial types that are presented 10 times each in a pseudorandomized order. Here, two different prepulse intensities (75 dB and 85 dB) and two different interstimulus intervals (ISIs, 30 and 100 ms) are tested. Startle stimulus alone trials and prepulse alone trials are interspersed. This block would have 6x10= 60 trials. Prepulses are 4 ms white noise pulses with 0 rise time. Variations of this protocol would consist in variable ITIs, higher startle stimulus intensities, different prepulse intensities and/or durations, and different ISIs between prepulse and pulse.
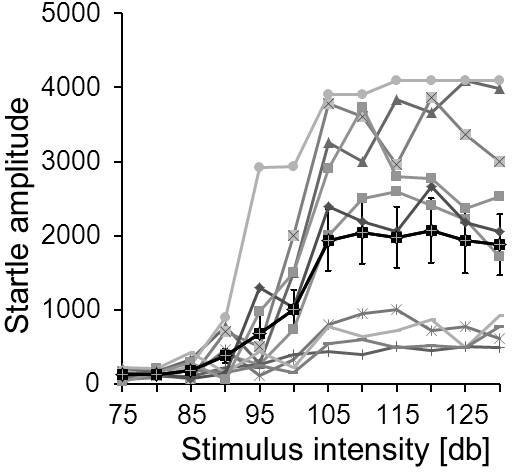 Figure 5.
Example for an i/o function. The input/output curves of 11 individual mice of the same strain are displayed in grey. In this case, the individual startle amplitudes vary considerably (startle responses are in arbitrary units). The solid black line shows the average startle amplitudes and standard errors at different startle stimulus intensities. These mice reached their maximum startle response at around 105 dB.
Figure 5.
Example for an i/o function. The input/output curves of 11 individual mice of the same strain are displayed in grey. In this case, the individual startle amplitudes vary considerably (startle responses are in arbitrary units). The solid black line shows the average startle amplitudes and standard errors at different startle stimulus intensities. These mice reached their maximum startle response at around 105 dB.
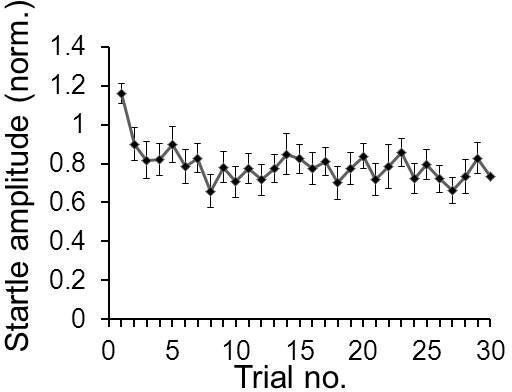 Figure 6.
Example for short-term habituation data. A typical average short-term habituation curve of 20 mice is shown. Startle amplitudes of each mouse in response to 30 startle stimuli were normalized to the average of its first two startle responses in trials 1 & 2. The normalized data was then averaged across mice and the standard error was calculated.
Figure 6.
Example for short-term habituation data. A typical average short-term habituation curve of 20 mice is shown. Startle amplitudes of each mouse in response to 30 startle stimuli were normalized to the average of its first two startle responses in trials 1 & 2. The normalized data was then averaged across mice and the standard error was calculated.
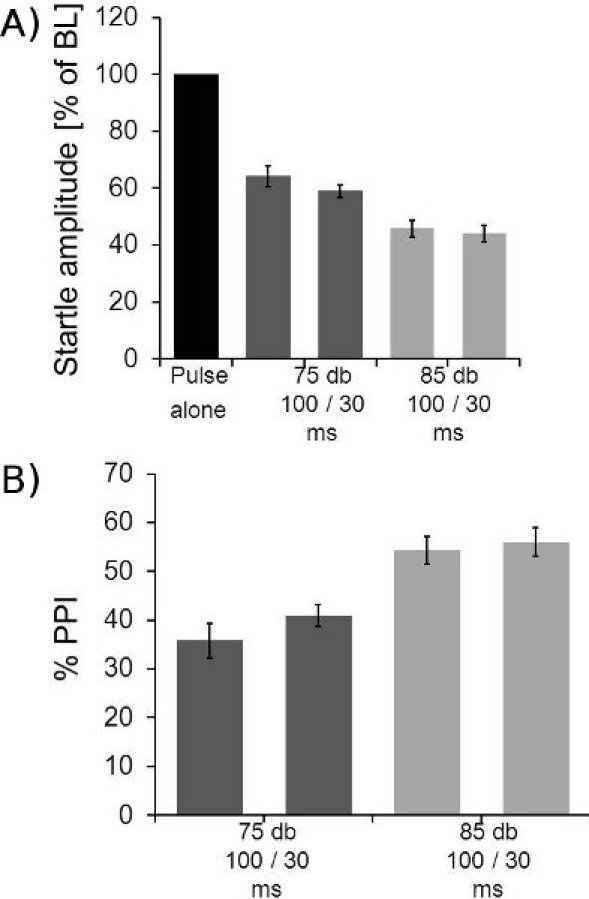 Figure 7.
Example for PPI data. A: Averaged PPI data of 8 mice is shown. The 10 startle alone trials of block II were averaged for each mouse and the averages of the other trial types expressed as the percentage of the stimulus alone startle amplitudes. The figure shows the startle response amplitudes under different prepulse conditions. Two different ISIs (30 and 100 ms) and two different prepulse intensities (75 and 85 dB) were measured. B: Same data as in A, but plotted as amount of PPI in percent of baseline startle. Data shown above was subtracted from 100. These mice showed a maximum PPI of around 50%. Please note that the same protocol yield PPI in most rat strains of around 90%.
Figure 7.
Example for PPI data. A: Averaged PPI data of 8 mice is shown. The 10 startle alone trials of block II were averaged for each mouse and the averages of the other trial types expressed as the percentage of the stimulus alone startle amplitudes. The figure shows the startle response amplitudes under different prepulse conditions. Two different ISIs (30 and 100 ms) and two different prepulse intensities (75 and 85 dB) were measured. B: Same data as in A, but plotted as amount of PPI in percent of baseline startle. Data shown above was subtracted from 100. These mice showed a maximum PPI of around 50%. Please note that the same protocol yield PPI in most rat strains of around 90%.
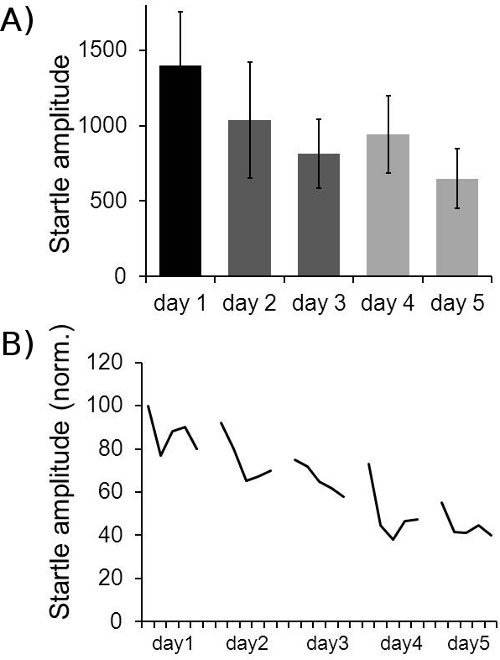 Figure 8.
Example for long-term habituation data. A: Averaged LTH data for 18 mice is shown. The first two startle responses in block I of each day were averaged across all mice. The relatively large standard error bars are mainly caused by differences in absolute startle amplitude between individual mice. B: Normalized startle amplitudes of 18 mice over five days. In order to reduce noise, groups of 6 consecutive startle responses in block I (30 stimuli) were always averaged per animal, resulting in five values for block I for each animal per day. These were normalized for each animal to the first value of the first day (100%). The average over all 18 animals is displayed. It shows STH within each day, as well as LTH across five days.
Figure 8.
Example for long-term habituation data. A: Averaged LTH data for 18 mice is shown. The first two startle responses in block I of each day were averaged across all mice. The relatively large standard error bars are mainly caused by differences in absolute startle amplitude between individual mice. B: Normalized startle amplitudes of 18 mice over five days. In order to reduce noise, groups of 6 consecutive startle responses in block I (30 stimuli) were always averaged per animal, resulting in five values for block I for each animal per day. These were normalized for each animal to the first value of the first day (100%). The average over all 18 animals is displayed. It shows STH within each day, as well as LTH across five days.
Discussion
Variations of the testing protocol
Modulation of startle responses have been studied for many decades in both humans and animals. A huge variety of different protocols have been used in the past. The current protocol is a relatively short and easy to perform test that works well in rodents, however, depending on the focus of interest and previous work on the respective questions, it might be useful to vary this protocol in order to obtain data that is comparable to previous relevant studies. A common variation includes the addition of more prepulse intensities ranging from 3 db above background noise to 20 db above noise. Also, the habituation block can be split into a short block of 5-10 stimuli before the PPI block, plus a third block of 5-10 stimuli after the PPI block18-20. A thorough study of the existing literature before designing a testing protocol is therefore essential.
Differences between species and strains
Startle response amplitudes and the amount of habituation differ considerably between single animals of the same specie and strain, whereas PPI seems to be relatively consistent. Mice do generally move more (voluntarily) during testing, which might be one reason why their data generally has a higher variability than rat data. Mice do also not habituate as well as rats. Differences between individual mouse or rat strains can be huge21-24, and it might be necessary to adapt stimulus parameter to the startle behavior of a specific strain in order to get optimal results. It should be avoided to use the same equipment to test both mice and rats. If it is inevitable, equipment should be thoroughly cleaned with ethanol.
Gain factors
Sometimes there are huge differences in individual startle responses within a group. In order to measure PPI and habituation, the baseline or first startle responses should be ideally covering the most part of the dynamic range of the measuring system. Overshoots are detrimental, since they lead to a systemic error, typically underestimating the amount of habituation or PPI. If startle responses are too small, however, modulations may be occluded by noise. Startle systems allow for the adjustment of a gain factor that amplifies the platform signal. Gain factors can be adjusted by displaying two or three startle stimuli during the last acclimation session (gain =1), however, one should keep in mind that they change the absolute startle response amplitude and therefore do not allow for a comparison of absolute startle amplitudes anymore. In order to avoid this drawback, the three startle responses that are used for gain factor adjustments could be used for determining the baseline startle magnitude. Alternatively, gain factors could be adjusted only after block I, so that the block II startle responses cover most of the dynamic range, while block I can be used for determining the baseline startle response.
Habituation versus sensitization
Habituation decreases the startle response amplitudes. This is opposed by a sensitization, which leads to an increase of startle responses upon repeated presentation25. Habituation and sensitization are two independent processes affecting the same behavior26. In order to measure habituation, sensitization should be minimized. Animals sensitize if a stimulus is aversive, thus too loud startle stimuli should be avoided for habituation measurements, for review see27. Stress, anxiety and fear do also increase startle responses28,oppose habituation and affect PPI18. Animals should therefore be well handled and acclimatized to the startle testing apparatus. Also, animal holders that are too small and physically restrain the animals are counterproductive, since they induce stress in the animals29.
Fixed versus randomized ITI
Common startle protocols use either a fixed inter-trial interval (ITI) typically of 20 or 30 sec or a variable interval that pseudorandomizes on values between 15 and 30 sec. The advantage of a randomized ITI lies in the fact that the animal cannot predict the time point of the next stimulation. It has been shown that e.g. attention to the prepulse augments its efficacy in suppressing startle responses13, 30. Measuring PPI with a fixed ITI may therefore also probe for attention processes. ITIs below 15 sec should be avoided in order to prevent effects caused by muscle fatigue and refractory periods of muscle responses.
Intensity and duration of prepulse
We use a very short prepulse of 4 ms duration in this protocol. Many other studies use a 20 ms prepulse. In order to be able to vary the interstimulus intervals (ISIs) and to measure also very short intervals, this short prepulse was introduced. The efficacy of the prepulse seems to be attenuated by its short duration as compared to a 20 ms prepulse of the same volume. We therefore use relatively loud prepulses of 75 and 85 dB. Whereas a 85 dB startle stimulus (20 ms) can be above threshold, a 85 dB prepulse (4ms) does normally not elicit startle responses. However, it is important to evaluate whether there are no startle responses elicited by the prepulse itself that would cause muscle fatigue and refractory states during the startle stimulus. Some treatments that disrupt PPI have shown to enhance prepulse sensivity31 (indicating the PPI disruption is not due to a loss of acoustic sensitivity), however, this could not be found in schizophrenic patients32Evaluations of the prepulse sensitivity can be done either by analyzing the platform data in the period between prepulse or startle pulse or by including prepulse alone trials in block II.
Different ISI versus different prepulse intensities
PPI in humans was originally measured at an ISI of 100 ms, where its effect is at its maximum7. In rats and mice PPI is at its maximum at 30-50 ms ISI, probably due to the smaller size of the brains33. In recent years it has become apparent that different transmitter and transmitter receptors are engaged in a serial manner in order to exert the fast but long-lasting inhibition of startle3, 34. Depending on the system affected, drugs or genetic manipulations might therefore affect PPI only at specific ISIs. We therefore recommend varying the ISI between 30 ms and 100 ms. This also allows recent studies to be compared to former studies that used 100 ms ISI only. The 85dB prepulse leads to a very robust maximum PPI of around 90%. Please be aware that this PPI cannot necessarily be augmented without running into a ceiling effect. PPI induced this way also seems to be rather robust, however, it is significantly disrupted e.g. by 1 mg/kg amphetamine. We recommend using a second prepulse of 75 dB which leads to 50-60% PPI only. This PPI can be augmented (e.g. by 1 mg/kg s.c. nicotine), and seems to be more vulnerable to genetic and pharmacological manipulation in general, however, it also seems to be more variable and inconsistent even within a subject. Former studies have used a huge variety of prepulse intensities and have often shown effects of treatments on PPI with specific prepulse intensities and no affect on PPI with other prepulse intensities. Thorough studies of the existing literature is therefore essential before choosing prepulse intensities and interstimulus intervals.
Combination with injections systemic/stereotaxic
Habituation and PPI testing is often performed in combination with systemic or stereotaxic injections. It is evident that in these experiments animals of a control group receive control vehicle injections. The injection procedure itself, however, might be very stressful for an animal, leading to a higher anxiety level and a potentiation and/or sensitization of the startle response (see above). It is therefore recommended to control for the effect of the injection procedure itself as well. If habituation is studied, prior injections might be a major obstacle. In order to alleviate the animal's anxiety, animals should be returned to their home cage for as long as possible before tested (without the drug wearing off). Injections should also be administered by an experienced person, in order to minimize the impact of the procedure on the animal. If stereotaxic injections are made through chronically implanted cannulae, the surgeon who implants the cannulae should avoid rupturing the rats' eardrums with the pointed ear bars. This might lead to hearing deficits. Blunt ear bars or ear cuffs that do not rupture eardrums are available for all stereotaxic devices. When rats are handled after surgery, the dust caps or dummies should be manipulated each time, so that the animals get used to it.
Acoustic startle as a hearing test
Finally it should be noted that i/o functions of acoustic startle and PPI can serve as a simple hearing test for rats and mice 35-37. Hearing deficits shift an i/o function to the right. Once PPI is established for a rat or mouse strain, animals can also be tested with variable prepulse intensities. If an animal is deaf or cannot hear the prepulse as loud as a control animal, it will display no or less PPI than control animals. On the other hand, an observed PPI deficit could always be caused by a hearing deficit, thus an i/o startle test or comparisons of baseline startle responses are crucial controls.
Disclosures
Production of this article was partially-funded by Med Associates, manufacturers of the instrument used in this article.
Acknowledgments
This work was funded by the Ontario Mental Health Foundation, the Natural Sciences and Engineering Research Council Canada, and Med Associates Inc.
References
- Davis M. The mammalian startle response. In: Eaton RC, editor. Neural Mechanisms of startle. New York: Plenum Press; 1984. [Google Scholar]
- Koch M. The neurobiology of startle. Prog Neurobiol. 1999;59:107–128. doi: 10.1016/s0301-0082(98)00098-7. [DOI] [PubMed] [Google Scholar]
- Fendt M, Li L, Yeomans JS. Brain stem circuits mediating prepulse inhibition of the startle reflex. Psychopharmacology (Berl) 2001;156:216–224. doi: 10.1007/s002130100794. [DOI] [PubMed] [Google Scholar]
- Davis M, Wagner AR. Habituation of startle response under incremental sequence of stimulus intensities. J Comp Physiol Psychol. 1969;67:486–492. doi: 10.1037/h0027308. [DOI] [PubMed] [Google Scholar]
- Pilz PK, Carl TD, Plappert CF. Habituation of the acoustic and the tactile startle responses in mice: two independent sensory processes. Behav Neurosci. 2004;118:975–983. doi: 10.1037/0735-7044.118.5.975. [DOI] [PubMed] [Google Scholar]
- Swerdlow NR, Geyer MA, Braff DL. Neural circuit regulation of prepulse inhibition of startle in the rat: current knowledge and future challenges. Psychopharmacology (Berl) 2001;156:194–215. doi: 10.1007/s002130100799. [DOI] [PubMed] [Google Scholar]
- Braff DL, Grillon C, Geyer MA. Gating and habituation of the startle reflex in schizophrenic patients. Arch Gen Psychiatry. 1992;49:206–2015. doi: 10.1001/archpsyc.1992.01820030038005. [DOI] [PubMed] [Google Scholar]
- van den Buuse M. Modeling the positive symptoms of schizophrenia in genetically modified mice: pharmacology and methodology aspects. Schizophr Bull. 2010;36:246–270. doi: 10.1093/schbul/sbp132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Geyer MA. Are cross-species measures of sensorimotor gating useful for the discovery of procognitive cotreatments for schizophrenia? Dialogues Clin Neurosci. 2006;8:9–16. doi: 10.31887/DCNS.2006.8.1/mgeyer. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fenton WS, Stover EL, Insel TR. Breaking the log-jam in treatment development for cognition in schizophrenia: NIMH perspective. Psychopharmacology (Berl) 2003;169:365–366. doi: 10.1007/s00213-003-1564-1. [DOI] [PubMed] [Google Scholar]
- Braff DL, Geyer MA, Swerdlow NR. Human studies of prepulse inhibition of startle: normal subjects, patient groups, and pharmacological studies. Psychopharmacology (Berl) 2001;156:234–258. doi: 10.1007/s002130100810. [DOI] [PubMed] [Google Scholar]
- Kumari V, Soni W, Sharma T. Normalization of information processing deficits in schizophrenia with clozapine. Am J Psychiatry. 1999;156:1046–1051. doi: 10.1176/ajp.156.7.1046. [DOI] [PubMed] [Google Scholar]
- Weike AI, Bauer U, Hamm AO. Effective neuroleptic medication removes prepulse inhibition deficits in schizophrenia patients. Biol Psychiatry. 2000;47:61–70. doi: 10.1016/s0006-3223(99)00229-2. [DOI] [PubMed] [Google Scholar]
- Swerdlow NR. Impaired prepulse inhibition of acoustic and tactile startle response in patients with Huntington's disease. J Neurol Neurosurg Psychiatry. 1995;58:192–200. doi: 10.1136/jnnp.58.2.192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Castellanos FX. Sensorimotor gating in boys with Tourette's syndrome and ADHD: preliminary results. Biol Psychiatry. 1996;39:33–41. doi: 10.1016/0006-3223(95)00101-8. [DOI] [PubMed] [Google Scholar]
- Swerdlow NR. Forebrain D1 function and sensorimotor gating in rats: effects of D1 blockade, frontal lesions and dopamine denervation. Neurosci Lett. 2006;402:40–45. doi: 10.1016/j.neulet.2006.03.060. [DOI] [PubMed] [Google Scholar]
- Cannizzaro C. Prenatal exposure to diazepam and alprazolam, but not to zolpidem, affects behavioural stress reactivity in handling-naive and handling-habituated adult male rat progeny. Brain Res. 2002;953:170–180. doi: 10.1016/s0006-8993(02)03282-1. [DOI] [PubMed] [Google Scholar]
- Gururajan A, Taylor DA, Malone DT. Effect of cannabidiol in a MK-801-rodent model of aspects of Schizophrenia. Behav Brain Res. 2011;222:299–308. doi: 10.1016/j.bbr.2011.03.053. [DOI] [PubMed] [Google Scholar]
- Brosda J. Pharmacological and parametrical investigation of prepulse inhibition of startle and prepulse elicited reactions in Wistar rats. Pharmacol Biochem Behav. 2011;99:22–28. doi: 10.1016/j.pbb.2011.03.017. [DOI] [PubMed] [Google Scholar]
- Ballmaier M. Cannabinoid receptor antagonists counteract sensorimotor gating deficits in the phencyclidine model of psychosis. Neuropsychopharmacology. 2007;32:2098–2107. doi: 10.1038/sj.npp.1301344. [DOI] [PubMed] [Google Scholar]
- Glowa JR, Hansen CT. Differences in response to an acoustic startle stimulus among forty-six rat strains. Behav Genet. 1994;24:79–84. doi: 10.1007/BF01067931. [DOI] [PubMed] [Google Scholar]
- Bullock AE. Inbred mouse strains differ in the regulation of startle and prepulse inhibition of the startle response. Behav Neurosci. 1997;111:1353–1360. doi: 10.1037//0735-7044.111.6.1353. [DOI] [PubMed] [Google Scholar]
- Bast T. Effects of MK801 and neuroleptics on prepulse inhibition: re-examination in two strains of rats. Pharmacol Biochem Behav. 2000;67:647–658. doi: 10.1016/s0091-3057(00)00409-3. [DOI] [PubMed] [Google Scholar]
- Buuse Mvanden. Deficient prepulse inhibition of acoustic startle in Hooded-Wistar rats compared with Sprague-Dawley rats. Clin Exp Pharmacol Physiol. 2003;30:254–261. doi: 10.1046/j.1440-1681.2003.03823.x. [DOI] [PubMed] [Google Scholar]
- Davis M. Sensitization of the rat startle response by noise. J Comp Physiol Psychol. 1974;87:571–581. doi: 10.1037/h0036985. [DOI] [PubMed] [Google Scholar]
- Groves PM, Thompson RF. Habituation: a dual-process theory. Psychol Rev. 1970;77:419–450. doi: 10.1037/h0029810. [DOI] [PubMed] [Google Scholar]
- Grillon C, Baas J. A review of the modulation of the startle reflex by affective states and its application in psychiatry. Clin Neurophysiol. 2003;114:1557–1579. doi: 10.1016/s1388-2457(03)00202-5. [DOI] [PubMed] [Google Scholar]
- Davis M, Walker DL, Myers KM. Role of the amygdala in fear extinction measured with potentiated startle. Ann N Y Acad Sci. 2003;985:218–232. doi: 10.1111/j.1749-6632.2003.tb07084.x. [DOI] [PubMed] [Google Scholar]
- Pare WP, Glavin GB. Restraint stress in biomedical research: a review. Neurosci Biobehav Rev. 1986;10:339–370. doi: 10.1016/0149-7634(86)90017-5. [DOI] [PubMed] [Google Scholar]
- Li L. Top-down modulation of prepulse inhibition of the startle reflex in humans and rats. Neurosci Biobehav Rev. 2009;33:1157–1167. doi: 10.1016/j.neubiorev.2009.02.001. [DOI] [PubMed] [Google Scholar]
- Yee BK, Russig H, Feldon J. Apomorphine-induced prepulse inhibition disruption is associated with a paradoxical enhancement of prepulse stimulus reactivity. Neuropsychopharmacology. 2004;29:240–248. doi: 10.1038/sj.npp.1300323. [DOI] [PubMed] [Google Scholar]
- Csomor PA. Impaired prepulse inhibition and prepulse-elicited reactivity but intact reflex circuit excitability in unmedicated schizophrenia patients: a comparison with healthy subjects and medicated schizophrenia patients. Schizophr Bull. 2009;35:244–255. doi: 10.1093/schbul/sbm146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yeomans JS. GABA receptors and prepulse inhibition of acoustic startle in mice and rats. Eur J Neurosci. 2010;31:2053–2061. doi: 10.1111/j.1460-9568.2010.07236.x. [DOI] [PubMed] [Google Scholar]
- Jones CK, Shannon HE. Effects of scopolamine in comparison with apomorphine and phencyclidine on prepulse inhibition in rats. Eur J Pharmacol. 2000;391:105–112. doi: 10.1016/s0014-2999(00)00055-8. [DOI] [PubMed] [Google Scholar]
- Clark MG. Impaired processing of complex auditory stimuli in rats with induced cerebrocortical microgyria: An animal model of developmental language disabilities. J Cogn Neurosci. 2000;12:828–839. doi: 10.1162/089892900562435. [DOI] [PubMed] [Google Scholar]
- McClure MM. Rapid auditory processing and learning deficits in rats with P1 versus P7 neonatal hypoxic-ischemic injury. Behav Brain Res. 2006;172:114–121. doi: 10.1016/j.bbr.2006.05.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fitch RH. Use of a modified prepulse inhibition paradigm to assess complex auditory discrimination in rodents. Brain Res Bull. 2008;76:1–7. doi: 10.1016/j.brainresbull.2007.07.013. [DOI] [PMC free article] [PubMed] [Google Scholar]