

Abstract
Injuries to articular cartilage are one of the most challenging issues of musculoskeletal medicine due to the poor intrinsic ability of this tissue for repair. Despite progress in orthopaedic surgery, the lack of efficient modalities of treatment for large chondral defects has prompted research on tissue engineering combining chondrogenic cells, scaffold materials and environmental factors. The aim of this review is to focus on the recent advances made in exploiting the potentials of cell therapy for cartilage engineering. These include: 1) defining the best cell candidates between chondrocytes or multipotent progenitor cells, such as multipotent mesenchymal stromal cells (MSC), in terms of readily available sources for isolation, expansion and repair potential; 2) engineering biocompatible and biodegradable natural or artificial matrix scaffolds as cell carriers, chondrogenic factors releasing factories and supports for defect filling, 3) identifying more specific growth factors and the appropriate scheme of application that will promote both chondrogenic differentiation and then maintain the differentiated phenotype overtime and 4) evaluating the optimal combinations that will answer to the functional demand placed upon cartilage tissue replacement in animal models and in clinics. Finally, some of the major obstacles generally encountered in cartilage engineering are discussed as well as future trends to overcome these limiting issues for clinical applications.
Keywords: Animals, Biocompatible Materials, therapeutic use, Cartilage, metabolism, pathology, Cartilage Diseases, pathology, therapy, Cell Differentiation, Chondrocytes, metabolism, pathology, transplantation, Guided Tissue Regeneration, Hematopoietic Stem Cell Mobilization, Humans, Intercellular Signaling Peptides and Proteins, metabolism, Mesenchymal Stem Cell Transplantation, Stem Cell Niche, Tissue Engineering, Tissue Scaffolds
INTRODUCTION
Structure and function of articular cartilage
Articular cartilage is a highly specialized tissue that reduces joint friction and protects the bone ends from the shear forces associated with high mechanical load. The articular cartilage consists of chondrocytes and few progenitor cells [1] which are organized in various layers, from the fibrotic to the mature and hypertrophic mineralizing layer of chondrocytes in direct contact with the sub-chondral bone. The extracellular matrix (ECM) of chondrocytes is distinct from that of other connective tissues. This ECM is composed of a network of fibrillar collagens that give the tissue its shape, strength and tensile force and, proteoglycans that give resistance to compression [2]. It contains the large aggregating proteoglycan aggrecan which is attached to hyaluronic acid polymers via a link protein and predominantly, the collagens type II (80–90% of total collagens), IX and XI. Once the cartilage is formed in the adult, the turn-over of ECM protein replacement is low with a collagen and proteoglycan half-live of 100 and 3–24 years, respectively [3]. This, low rate of matrix remodelling partly explains why chondrocytes are relatively inactive metabolically although they can respond to various stimuli to maintain normal homeostasis.
Poor intrinsic capacity of cartilage for repair
After injury due to traumas or osteo-articular diseases, the articular cartilage is frequently damaged resulting in fibrillation and subsequent degradation which can also lay down into the sub-chondral bone. This is the result of the limited capacity of cartilage for repair due to the absence of vasculature that cannot provide the progenitor cells from the blood or the bone marrow to enter the tissue. Accordingly, the resident articular progenitor cells or chondrocytes entrapped within the surrounding matrix do not migrate into the lesions to secrete a reparative matrix. Consequently, the limited repair capacity and the absence of pharmacological agents have prompted researchers and surgeons to develop surgical methods to restore cartilage surfaces using tissue grafts or cell-based therapies (for review, see [4]). However, none of the current cartilage repair approaches allowed the generation of long term hyaline cartilage replacement tissue.
Limitations of current surgical methods for cartilage repair
Among the possible explanations for the limited results described with the current methods of cartilage repair is the lack of integration of the chondrocytes within the existing cartilage. This is likely to be due to the insufficient capacity of implanted cells to secrete the cartilaginous matrix and to recapitulate the complex events resulting in the zonal organization of the cartilaginous tissue [5]. The lack of integration may also be due to the incomplete differentiation of chondroprogenitor cells or instability of the chondrocytic phenotype. The implantation of undifferentiated cells has already been applied in humans and although a significantly improved patient outcome was observed after one to five years, the defects were filled with a fibrocartilaginous tissue [6]. Finally, leakage of the cell suspension may be the cause of loss or decreased viability of the implanted cells as currently reported in autologous chondrocyte transplantation [7]. In summary, cell-based therapies have proved their feasibility but showed no superiority over other surgical methods on the long-term highlighting a crucial need for optimizing various combinations of cell types, scaffolds and/or chondrogenic factors (Fig. 1).
Figure 1.
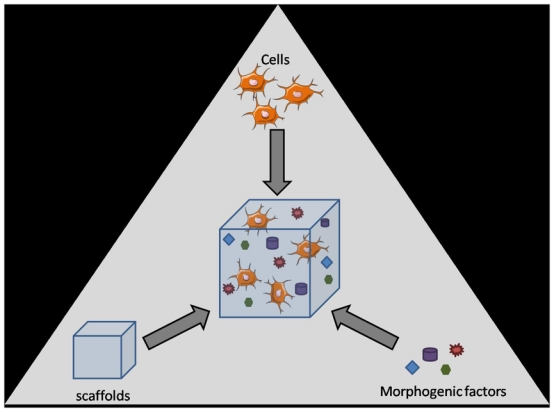
Schematic representation of the componants required for cartilage tissue engineering. Cells (stem cells with chondrogenic potential or chondrocytes), morphogenic factors and scaffolds (natural or synthetic) are combined in vitro to form an engineered scaffold suitable for implantation. Illustration with permission from Servier (http://www.servier.fr/smart/TermsOfUse.asp )
CELL SOURCES FOR CARTILAGE REPAIR
Chondrocytes
Chondrocytes, the resident cells of cartilage, produce the components of the ECM and represent the cells of choice for engineering articular cartilage. Adult chondrocytes have been isolated from various sources like articular cartilage, nasal septum, ribs or ear cartilage [8, 9]. However, ear cartilage is an elastic cartilage, which exhibit different mechanical properties as compared to the hyaline cartilage found in joint and nasal septuml. Isogai et al. have shown that chondrocytes give rise to cartilage tissue having the characteristics of its original tissue [8]. A chondrocyte from ear cartilage will thus give rise to an elastic cartilage. With a view to an application to the repair of articular cartilage, it therefor seems more appropriate to use hyaline cartilage as a source of chondrocytes. A comparison between different sources of hyaline chondrocytes (nasal, costal, and articular) has shown the superiority of costal and nasal chondrocytes on articular chondrocytes in term of quantity of cartilage formed after transplantation in subcutaneous sites [8]. One of the main limits related to the use of chondrocytes, is their instability in monolayer culture resulting in the loss of their phenotype. Indeed, chondrocytes lose the expression of the chondrocytic markers which are collagen II and aggrecan mainly, but also SZP (superficial zone protein) [10]. This loss of the chondrocytic phenotype is accompanied by a phenotypic shift towards a fibroblastic one. This fibroblastic phenotype is characterized by an increased expression of collagen I and the adoption of the spindle-shape characteristic of fibroblasts [11]. This process of dedifferentiation is however reversible. Indeed, if dedifferentiated chondrocytes are placed in a three-dimensional environment, they retrieve their differentiated phenotype [12, 13]. Chondrocytes from osteoarthritic (OA) cartilage have also been considered. However, OA chondrocytes undergo metabolic alterations, which can lead to a low response to inductive environmental factors [14, 15]. Although chondrocytes derived from OA patients appear less appropriate for articular cartilage repair, it has been reported that OA chondrocytes may be able to recover a normal chondrocytic phenotype after in vitro three-dimensional (3D) culture [16]. However, additional studies are required to clearly decipher whether OA chondrocytes could be manipulated in vitro to be suitable for cell therapy of cartilage.
Mesenchymal stem cells
Multipotent mesenchymal stromal cells or mesenchymal stem cells (MSC) are an attractive source of cells for cartilage engineering due to their easy access and high capacity of in vitro expansion. They are mainly isolated from bone marrow or adipose tissue but have been isolated from a number of other tissues including synovium, periosteum, umbilical cord vein or placenta [17]. MSC were first characterized by their clonogenic potential determined by the capacity to form Fibroblast Colony Forming Units (CFU-F). In the bone marrow, the frequency of CFU-F is in the range of 1 cell in 104–105 mononuclear cells [18, 19]. They are now characterized by their capacity to adhere to plastic, their phenotype (CD73+, CD90+, CD105+, CD14− ou CD11b−, CD19− ou CD79α−, CD34−, CD45− and HLA-DR−) and their trilineage differentiation potential [20]. MSC also exhibit the potential to differentiate into myocytes, tendinocytes, ligamentocytes [21], cardiomyocytes [22], neuronal cells [23, 24] and other cell types [25]. More recently, these cells have been described as immunoregulatory cells since they were shown to escape the immune recognition and to inhibit the host defence mechanisms (for review, see [26]). The interest of various other progenitor cells, such as multipotent adult progenitor cells (MAPC) or marrow-isolated adult multilineage inducible (MIAMI) cells, for cartilage differentiation has been shown in vitro but has still to be validated in vivo [27, 28].
DIFFERENTIATION FACTORS REQUIRED FOR CARTILAGE ENGINEERING
A number of growth and differentiation factors that regulate cartilage development and homeostasis of mature articular cartilage have been identified. The most characterized factors which stimulate the anabolic activity in cartilage include Transforming Growth Factor (TGF)-β, Bone Morphogenetic Protein (BMP), Fibroblast Growth Factors (FGF), Insulin Growth factor (IGF)-1, Hedgehog (hh) and Wingless (Wnt) proteins.
Transforming Growth Factor-β family
The TGF-β superfamily of polypeptides includes TGF-β, BMP, activins and inhibins. Because TGF-β and BMP are the best characterized peptides and their role in osteogenesis and chondrogenesis is well documented [3], we will focus on these molecules in the following paragraphs. The TGF-β family includes 5 members (TGF-β1–5) which are produced by many different cell types. However, the concentration of TGF-β is approximately 100-fold greater in bone than in other tissues [29]. Articular cartilage itself contains large amounts of latent TGF-β. Although present in tiny quantities in normal physiological conditions, active TGF-β1, 2 and 3 are generally considered to be potent stimulators of proteoglycans and type II collagen synthesis in primary chondrocytes (for review[3, 29]. In vitro, TGF-β1, 2 and 3 were also shown to induce the chondrogenic differentiation of MSC [30–32]. Recently, rabbit MSC encapsulated with thermo-reversible hydrogel releasing heparin-bind TGF-β3 were shown to differentiate toward chondrocytes [33]. In vivo, a porous gelatin-chondroitin-hyaluronate scaffold in combination with a controlled release of TGF-β1 could induce the chondral differentiation of MSCs to form ectopic cartilage [34]. The same group also reported that implantation of rabbit MSC in a full-thickness defect resulted in better chondrocyte morphology, integration, continuous subchondral bone, and much thicker newly formed cartilage layer when compared to control group [34]. However, several studies have shown that injection of TGF-β or TGF-β-expressing adenoviruses results in side effects in the joints, such as osteophyte formation, swelling and synovial hyperplasia [35–37].
Bone morphogenetic proteins
BMPs constitute a large sub-class of polypeptides whose role is essential for chondrogenesis during skeletal development. Indeed, mutations in BMP-5 and BMP-14 genes result in brachypodism in mice and chondrodysplasia in humans. A number of mice deficient for BMP are nonviable but in BMPR-IB, ActR-IA, BMP-7 or BMP-14 deficient mice, severe appendicular skeletal defects have been observed suggesting that they play important synergistic or overlapping roles in cartilage and bone formation in vivo [38]. A number of BMP, including BMP-2, -4, -6, -7, -13, -14 can stimulate the chondrogenic differentiation of MSC [39–42] and enhance the synthesis of collagen type II and aggrecan by chondrocytes in vitro [43]. In vivo, healing of full-thickness cartilage defects in the rabbit was improved when combining microfracture and recombinant BMP-7 [44]. Similarly, the implantation of a type I collagen sponge containing BMP-2-expressing naked plasmid DNA implanted in full-thickness cartilage defects stimulates the transfection of MSC subjacent to the defect and cartilage repair [34]. The use of ex vivo retrovirally transduced muscle-derived stem cells isolated from mouse skeletal muscle to express BMP-4 enhanced chondrogenesis and significantly improved articular cartilage repair in rats [45]. A more recent study indicated that repair of chondral lesions in the knee joints of miniature pigs by periosteal precursor cells is facilitated in deeper hypoxic zones of cartilage repair tissue and is stimulated by BMP-2, and to a lesser extend IGF-1, which enhance HIF-1α activity [46]. However, when implanted in ectopic localizations, BMPs led to bone formation via endochondral ossification which should be avoided for articular cartilage engineering [47]. This observation points to the notion that optimal regulation of BMPs may enhance their efficacy in a regulated tissue engineering strategy. Indeed, Huard’s team showed that Noggin delivery can inhibit heterotopic ossification caused by BMP-4, demineralized bone matrix, and trauma in calvaria defect model [48, 49] suggesting that this strategy may be also useful for inhibing endochondral ossification induced by hypertrophic cartilage. Although these BMP have shown great potential in animal models, no clinical studies have been conducted to validate their potential to enhance cartilage repair in humans.
Wingless family
The Wnt family of secreted ligands contains more than 20 members in vertebrates that are characterized by conserved cysteine residues. These proteins exhibit unique expression patterns and distinct functions in development [50]. The Wnt family members signal through the canonical β-catenin-dependent pathway or β-catenin-independent pathways. Various Wnt members are involved both in early and late skeletal development and play a role in the control of chondrogenesis and hypertrophy. 9 Wnt genes are expressed in the postnatal growth plate, including Wnt-4, -5a, -5b, -10b and -11 at higher levels, and Wnt-2b, -7b, -9a and -10a at very low levels [51]. Wnt-1, Wnt-4, Wnt-7a, Wnt-8 block chondrogenic differentiation but display different effects on hypertrophy. On the contrary, Wnt-5a and Wnt-5b promote chondrogenesis [52, 53]. Wnt-5a together with Wnt-5b were shown to regulate the proliferation of chondrocytes and their maturation into hypertrophic chondrocytes in both the embryonic and postnatal growth plate and Wnt-11 does not affect chondrogenic differentiation. The role of Wnt-3a on chondrogenesis is more controversial. Importantly, we recently identified Wnt-6 as a new factor able to induce the chondrogenic differentiation of primary MSCs while inhibiting both osteogenic and adipogenic differentiation (pers.com.). The key genes identified in both embryonic cartilage development and postnatal endochondral bone formation includes the Wnt-5a/Fz-5 receptor complex and Wnt-7a [54]. It is generally reported that Wnt members responsible for the induction of the osteogenic differentiation activate the β-catenin-dependent pathway. Indeed, in vitro inactivation of β-catenin in MSC causes chondrocyte differentiation under conditions allowing only osteoblasts to form [55]. Consistently, in vitro loss- and gain-of-function analyses reveal that β -catenin activity is necessary and sufficient to repress the differentiation of mesenchymal cells into Runx2- and Sox9-positive skeletal precursors [56]. However, β-catenin was recently shown to be required for both osteogenesis and chondrogenesis in adult mature tissues [57]. Overall, it appears that the Wnt network has dual roles in cartilage, as has been described in other tissues: it is an important regulator of chondrocyte development, but deregulated signaling is detrimental to mature tissues and may lead to disease.
Fibroblast growth factor family
In vertebrates, the FGF family comprises twenty-two structurally related proteins with a molecular mass from 17 to 34 kDA that bind one of four tyrosine kinase FGF receptors (FGFR) [58]. The importance of FGF signalling in skeletal development is highlighted by the number of dysplasias, mainly dwarfing chondrodysplasias and craniosynostosis syndromes, attributed to specific mutation in the genes encoding the FGFR-1, -2 and -3 [59]. During limb development, FGFR-2, Sox9 and collagen type II are among the earliest genes upregulated in condensing mesenchyme. Mice lacking FGFR-3 develop skeletal overgrowth while mice overexpressing an activated form of FGFR-3 develop skeletal dwarfism. These data have shown that signalling through FGFR-3 negatively regulates chondrocyte proliferation through a STAT1 pathway and differentiation through the MAPK pathway. Genetic studies have also identified defects in chondrogenesis in mice lacking FGF18 whereas no apparent defects in chondrogenesis occurs in mice lacking FGF-2, -5, -6, -7, -8 and -17. Additionally, the skeletons of FGF-9−/− mice are sightly smaller than the wild type littermates [60].
In the adult cells, the chondrogenic effect of FGF and FGFR has been confirmed by very few studies. Indeed, forced expression of FGFR-3 in the murine C3H10T1/2 MSC line was shown sufficient for chondrogenic differentiation [61]. Accordingly, FGF-18 which was shown to be a selective ligand for FGFR-3 in limb bud mesenchymal cells, suppress their proliferation while promoting differentiation to produce cartilage matrix [62]. In cultured chicken chondrocytes, FGF-9, another ligand for FGFR3, rapidly induces the upregulation and secretion of the matrix phosphoprotein osteopontin, known to be associated with chondrocyte and osteoblast differentiation. Unexpectedly, FGF-9-induced osteopontin was accompanied by inhibition of differentiation and increased proliferation of the treated chondrocytes [63]. In adult chondrocytes, FGF-2 is mainly mitogenic although recent studies have shown that it can inhibit the anabolic ativities of other growth factors, such as IGF-1. MSCs expanded in FGF-2-supplemented medium proliferated more rapidly than control MSCs and FGF-2 treatment enhanced subsequent chondrogenic differentiation in a 3-dimensional culture [64, 65]. In addition, in a rabbit model, FGF-2 can stimulate articular cartilage restoration in temporomandibular osteoarthritic defects, although the effective concentration range of FGF-2 would have to be determined [66]. Implantation of a fibrin sealant incorporating FGF-2 successfully induced healing of a mechanically induced defect with hyaline cartilage and concomitant repair of the subchondral bone [67]. One study reports the use of FGF-18 in a setting of rapidly progressive osteoarthritis in rats. The data showed that FGF-18 induced an increase in chondrophyte size and remodeling of the subchondral bone suggesting that it can stimulate repair of damaged cartilage [68]. The important issues coming from the contradictory results reported to date include a better characterization of the signalling pathways activated by FGFs, a better understanding of the interplay between these pathways as well as understanding the contribution of additional factors, such as HS, in regulating FGF activity in cartilage and bone development.
Insulin-like growth factor family
The IGF family is composed of two ligands (IGF-1 and IGF-2), two cell surface receptors (IGF1R and IGF2R), at least six different IGF binding proteins (IGFBP-1 to IGFBP-6), and multiple IGFBP proteases, which regulate IGF activity in several tissues. IGF-1 is the most studied form with respect to cartilage repair. In embryonic development, mice with IGF-1−/− mutations display severe growth retardation, have developmental defects in various organs while IGF-2−/− are 60% smaller than their wild-type littermates but grow normally after birth. Accordingly, mice nullizygous for the IGF1R gene demonstrate severe fetal growth retardation [69]. In humans, a reported natural deletion of exons 4 and 5 of the IGF-1 gene results in severe pre- and postnatal growth and developmental deficits, combined with mental retardation [70].
IGF-1 and IGF1R are expressed by chondrocytes, osteoblasts, and osteoclasts. IGF-1 is considered an essential mediator of cartilage homeostasis through its capacity to stimulate proteoglycan synthesis and, promote chondrocyte survival and proliferation [71, 72]. IGF-1 also induces the differentiation of MSC towards the chondrocytic phenotype as shown by the upregulation of the specific markers [33, 73]. In a critical size full-thickness cartilage defect horse model, defects filled with fibrin clots loaded with IGF-1 repaired better than empty defects and contained mainly chondrocytes with predominantly collagen type II rich matrix [74]. In the same model, the combined use of chondrocytes and IGF-1 tend to improve the overall continuity and consistency of the repair tissue [74]. However, an age-related or OA-associated decline in the responsiveness of chondrocytes to IGF-1 appears to be due at least in part to over-expression of IGFBPs. The titration of FGFs by IGFBPs may account for the variable results reported to date with IGF-1 treatment for in vivo cartilage repair studies. Improvement will require optimizing the dose, injection regimen and/or combination with other growth factors.
Hedgehog family
In mammals, the Hh family comprises 3 members of highly conserved proteins: Sonic hedgehog (Shh), Indian hedgehog (Ihh), and Desert hedgehog (Dhh). In concert with other signaling molecules, Ihh has been found to function as a central regulator of endochondral ossification, coordinating chondrocyte proliferation, differentiation, and ossification of the perichondrium. Expression of Ihh induces the upregulation of a second secreted factor, parathyroid hormone-related protein (PTHrP), which is expressed in distal chondrocytes of the skeletal elements. Mice overexpressing Ihh or a constitutively activated allele of Smo displayed an increased chondrocyte proliferation [75]. Furthermore, as Gli3 is the major mediator of Shh signaling during limb patterning, it is not surprising that mutations in the human Gli3 gene cause a variety of inherited skeletal patterning defects [76]. The requirement of Hh signalling varies in different developmental processes: osteoblast differentiation in the perichondrium and chondrocyte differentiation in the sclerotome are induced by transient exposures to Ihh or Shh, respectively, whereas the onset of columnar and hypertrophic chondrocyte differentiation depends on continuous Ihh signaling to maintain the Ihh-PTHrP interactions. Furthermore, hedgehog signaling has been shown to interact with several other signaling pathways, including those of FGFs, Wnts, and BMPs.
Although hedgedhog signalling has been mainly described in growth plate chondrocytes, the role of Shh during chondrogenesis has been shown. Indeed, in vitro, Shh-treated MSCs showed expression of cartilage markers aggrecan, Sox9, CEP-68, and collagen type II and type X within 3 weeks [77]. In addition, Hh pathway dramatically impaired adipogenesis of MSC, with reduced lipid accumulation, a decrease in adipocyte-specific markers, and acquisition of insulin-resistant phenotype stimulation [78]. In vivo, few studies are available and report the role of Shh on bone regeneration [79].
NATURAL AND SYNTHETIC SCAFFOLDS FOR CARTILAGE ENGINEERING
A number of matrices have been tested in vitro and in vivo in preclinical and clinical studies. These matrices can be classified according to their nature (proteic, polysaccharidic, synthetic and natural) or to their form (mass, mass porous, foam, viscous liquid and hydrogel). The ideal properties of a matrix are biocompatibility to prevent the inflammatory reactions to protect host tissue; three-dimensional shape allowing proliferation and cellular differentiation and porosity permitting migration of cells and diffusion of molecules, nutrients and oxygen. The matrix must also allow cell adhesion to facilitate the implantation of cells in the lesion and maintainance in the implant. It can also be bioactive and allow the homogeneous and controlled release of growth factors or morphogens. Finally, the matrix has to adhere to the host tissue; maintain its mechanical integrity in order to avoid its flow after implantation and be degradable to integrate the physiological processes of tissue remodeling. The matrix must be applicable by mini-invasive surgery thus if possible injectable. Main matrices used in tissue engineering of the cartilage are referred in table 1. Mixed matrices can be also obtained either by an association of two or several matrices, or by modifying them chemically or structurally [80].
Table 1.
Main scaffolds used for the 3-dimensional culture and transplantation of chondrogenic cells in cartilage tissue engineering.
| Proteic | Collagen [6, 83, 87, 88] |
| Fibrin [89, 93–95] | |
| Laminin (Matrigel ™) [148, 149] | |
| Gelatin [113, 150] | |
| Polysaccharidic | Agarose [98–100] |
| Alginate [103, 105, 106] | |
| Hyaluronic acid* [32] | |
| Chitosan [115] | |
| Cellulose [118, 121, 151] | |
| Synthetic | Poly lactic acid [152, 153] |
| Poly glycolic acid [127] | |
| Carbon fibres [129] | |
| Dacron (polyethylene terephtalate) Teflon (polytetrafluoroethylene) [130, 131] | |
| Polyestherurethane [154] | |
| Polybutyric acid [155] | |
| Polyethylemethacrylate [156] | |
Natural matrices
Collagen matrices
Collagen-based matrices or collagen sponges are among the mostly used matrices for cartilage engineering. Collagen is naturally degraded by collagenases and serines proteases. Its degradation is controlled locally by the cells present in the tissue [81]. These collagen matrices implanted alone improve the spontaneous repair process of osteochondral defects in the rabbit [82]. They are however generally associated with chondrocytes [83] or MSC [84]}. MSC seeded in type I and III collagen gels and implanted in osteochondral defects allowed the formation of cartilage and subchondral bone which was mechanically inferior to healthy articular cartilage and showed signs of degeneration after 24 weeks [85]. Moreover, the use of type I collagen from bovine origin may induce the production of antibodies by the host [86]. Collagen gels like Atelocollagene® (Koken Co Ltd, Tokyo, Japan) are favorable for the culture of chondrocytes and the synthesis of the ECM. These gels have been used in vitro as three-dimensional support for the culture of autologous human chondrocytes and in humans. This last study showed encouraging results, since 93% of the patients exhibited hyaline cartilage containing type II collagen with a biomechanical response similar between repaired tissue and healthy cartilage [87]. Another multicentric clinical study also showed positive preliminary results after implantation of grafts of collagen gel containing autologous chondrocytes and three-dimensional culture in vitro [88]. Collagen gels containing MSC formed hyaline-like tissue in cartilaginous defects after 7 months and after 1 year, patients had recovered a normal activity [6]. Despite immunoreactivity associated to its bovine origin, collagen gels therefore could appear as suitable matrices for cartilage tissue engineering.
Fibrin Glue
The fibrin adhesive is obtained by polymerization of fibrinogen in the presence of thrombin [89]. The fibrinogen is a physiologic liquid present in blood which is activated to polymerization in vascular lesions or in pathological situations [90]. Physiologically, fibrin promotes the spontaneous repair activity of articular cartilage but also has a pro-inflammatory activity. Fibrin induces its own degradation by the ECM components into non toxic endpoint components. Several studies have reported that the use of fibrin glue and chondrocytes improve cartilage repair in vivo [91–94]. In horse, fibrin glue containing either chondrocytes or MSC allowed the formation of a new cartilaginous tissue containing high proteoglycan and type II collagen contents [95, 96]. After 30 days, the defects filled with fibrin containing MSC exhibited a higher arthroscopic score compared with fibrin alone but it was no more significant after 6 months [96]. Nevertheless, due to its lack of mechanical stability, the use of fibrin glue is restricted. In human, its use is limited to seal off the periosteal flap in the autologous chondrocyte implantation (ACI) technique [97].
Agarose
Agarose is a polysaccharide which contains residues of L and D-galactose and is isolated from Chinese algae. Agarose has been mostly used in vitro as a matrix for the 3D culture of chondrocytes. It has also been used to help the chondrogenic differentiation of stem cells [98]. Implantation of agarose containing chondrocytes or MSC in osteochondral defects allows the formation of a repaired tissue containing collagens and proteoglycanes [99]. However, when implanted alone, agarose inhibits the process of spontaneous repair [100]. Moreover, due to its weak degradation, agarose has been poorly studied in vivo.
Alginate
Alginate is a linear polysaccharide purified from brown algae. It can undergo a reversible gelation in aqueous medium by crosslinking of bivalent cations and its dissolution can be obtained very quickly by using a calcium chelating agent. Alginate has been widely used in vitro as a matrix for the three-dimensional culture of chondrocytes because it allows the maintenance of the chondrocytic phenotype and the synthesis of ECM proteins [101–103]. In nude mice, the subcutaneous implantation of alginate beads containing MSC differentiated in vitro towards a chondrocytic phenotype allowed the production of cartilaginous protein [104]. In vivo, the results were disappointing since alginate alone inhibits spontaneous repair [105] and when associated with chondrocytes, it did not repair osteochondral defects partly due to severe immunological reactions [106, 107]. Nevertheless, a hybrid agarose-alginate hydrogel, Cartipatch® (Tissue Bank of France, Lyon, France), was tested for in vivo implantation of autologous chondrocytes in human. After two years, all patients clinically improved and 8 out of 13 patients had hyaline cartilage in biopsies from neotissue [108].
Hyaluronic acid
Hyaluronic acid (HA) is a component of the cartilaginous ECM which forms macromolecules of important size. HA is degraded naturally by hyaluronidases [81] but its degradation products are able to induce chondrolysis [109]. HA-based matrices increase the synthesis of ECM by chondrocytes in vitro and in vivo [110]. Nevertheless, under an unmodified form, HA is not suitable for cartilage repair [111] and needs crosslinking to enhance its biocompatibility [112]. HA may also be associated to other matrices. As an example, a tripolymer of gelatine, chondroitin and HA sulphate allowed the maintenance of the chondrocytic phenotype and type II collagen synthesis in vitro [113]. Hyalograft© (Fidia Advanced Biopolymers, Abano Terme, Italy), a tissue engineered graft composed of autologous chondrocytes and HYAFF 11 (Fidia Advanced Biopolymers, Abano Terme, Italy) showed improvement of cartilage function in 91.5% of patients [114].
Chitosan
Chitosan is a natural linear polymer pertaining to the glycosaminoglycan (GAG) family. Chitosan is mainly found in cuticules of arthropods, the endosquelette of cephalopods and in mushrooms. Chitosan can be associated with chondroitin sulphate to form hydrogels [90] and can be degraded by lysosomes [81]. Matrices containing chitosan are biocompatible and are widely used for cellular encapsulation, drug release and cell culture [115]. Several in vitro studies indicate that matrices containing chitosan are able to improve cartilage repair, promote chondrogenic activity of human chondrocytes and synthesis of ECM proteins [90]; when used alone or in association with various other polymers, like alginate or hyaluronic acid [116]. In vivo, chitosan-based matrices were reported to induce the formation of a hyaline-like repair tissue in articular cartilage defects [117]. To our knowledge, chitosan-based matrices have not yet been evaluated in human.
Cellulose
Cellulose is a semicrystalline polymer of glucose. The cellulose is found in plants and is the most widely spread natural polymer. Cellulose is degradable by enzymes like cellulases [118]. The biocompatibility of cellulose and its derivatives is well established [119, 120]. In vitro, the use of a cellulose polymer allowed the proliferation of chondrocytes and showed good biocompatibility [118]. In addition, we report that injectable hydroxypropylmethylcellulose hydrogel may be used for articular cartilage repair [121]. To date, however, few in vivo studies have been performed in this field of tissue engineering.
Artificial matrices
Polylactic acid and polyglycolic acid
Polylactic acid (PLA) and polyglycolic acid (PGA) are derived from alpha hydroxypolyesters. PLA and PGA are degraded either by hydrolysis, or specific cleavage of oligopeptides [81]. Their degradation products are however partially cytotoxic [80] and these polymers induce important immunological reactions [122, 123]. Originally, they were developed to form resorbable suture wire (vicryl™) and medical devices (screw, plates). Since twenty years, they are tested alone or mixed with other matrices for cartilage tissue engineering [124, 125]. Various forms of these polymers, from the fine fibrillary layer to the sponge, have been developed. PGA polymers provide the best in vitro results, with a cellular density near of that found in vivo and a continuous production of type II collagen [126]. In vivo studies were mainly performed in the rabbit model [127]. In human, BioSeed® (BioTissue Technologies, Freiburg, Germany) containing autologous articular chondrocytes was reported to induce the formation of a hyaline cartilage and to improve significantly clinical scores [128].
Carbon fibers
Carbon fibers are inert and therefore did not induce specific biological answer. They were used, without success, to fill rabbit cartilage defect in order to improve the spontaneous repair [129]. The neotissue was fibrous and exhibited only weak mechanical properties. Despite these unsatisfactory results, carbon fibers have been applied in human with very variable results [90].
Dacron and Teflon
Dacron (polyethylene terephthalate) and Teflon (polytetrafluoroethylene) have been used to improve spontaneous repair of articular cartilage in rabbit. Results have reported the formation of a repair tissue, which was either a vascularized fibrous tissue or a fibrocartilage [130, 131]. Due to an increased rigidity of joint after resurfacing with Teflon [132] and immunological reaction observed when these matrices was used as suture wires, these matrices seem not adequate for cartilage tissue engineering.
Hydrogel
The search for a minimally-invasive surgery has justified the development of injectable matrices for cartilage tissue engineering. These injectable matrices have to be able to solidify, once implanted, to gain the desired shape and present the mechanical properties of the tissue to repair [80]. Hydrogels are three-dimensional polymeric networks that are able to absorb and retain large volume of water. Viscous polymers from various origins (see table 1) can be transformed in hydrogel by modifying their environnement. Crosslinking of hydrogels can be initiated by physical stimuli like pH, temperature or ionic environment or chemical crosslink through crossslinking agent, photopolymerization or enzymatic reaction [81]. Hydrogels generally present good biocompatibility. Moreover, cells, growth factors or bioactives components can be homogeneously incorporated. Their high water content allowed rapid diffusion of nutrients and metabolites [133]. Hydrogels are produced from natural or synthetic polymers (see table 2). Collagen gels represent the main protein–based polymer used for hydrogel production [134]. Among polysaccharide- based polymers, HA [114], alginate [135], chitosan [136] or cellulose derivatives [94, 137] have been used with satisfactory results. Among synthetic polymers, polyvinyl alcohol, polyethylene glycols (PEG) and poly(lactide-coglycolide) represent the mostly used. Hydrogels therefore appeared as appealing materials for cartilage tissue engineering.
Table 2.
Main matrices used to prepare hydrogels for cartilage tissue engineering
CELL THERAPIES IN CLINICS
Therapies currently used or available in clinics are bone marrow stimulation techniques (subchondral drilling [138], abrasion, microfracture [139]), multiple osteochondral graft like mosaicplasty [140] and autologous chondrocyte implantation (ACI) [141]. Clinical studies report an improvement in clinical outcome measures compared with preoperative assessment but no superiority of one technique over the others [142].
After a first generation of ACI, a second generation has recently been developed where autologous chondrocytes are associated with a matrix which provides a three-dimensional environment and support at the implantation site [143]. Various concepts are presently accessible in clinics: MACI® (Verigen, Leverkusen, Germany); Maix® (Matricel, Hezoenrath, Germany); Chondro-gide® (Geistlich Biomaterials, Wolhusen, Switzerland); Atelocollagen® (Koken Co Ltd, Tokyo, Japan), Hyalograft® C (Fidia Advanced Biopolymers, Abano Terme, Italy), Bio-Seed®-C (BioTissue Technologies, Freiburg, Germany). Despite good clinical results, these above mentioned concepts suffer a major limitation related to the fact that matrices require a surgical incision into the joint to be implanted. To address the issue of less invasive transplantation technique, the development of injectable biomaterials as hydrogels suitable for minimally invasive transplantation of chondrogenic cells is the focus of current research. The second limitation of the ACI and matrix-induced ACI is the use of chondrocytes harvested from articular joints which implies a limited quantity of autologous chondrocytes [144]. Therefore, cellular therapies using MSC, an easily accessible source of autologous cells, have been proposed. Only a few case studies have been reported to date. Transplantation of bone marrowderived MSCs seeded on type I collagen membranes in articular cartilage defects resulted in increased arthroscopic and histologic grading scores [145]. Centeno et al. have injected bone marrow-derived MSCs percutaneously in the knee of a patient with radiographic signs of degenerative joint disease [146]. 24 weeks later, they observed a significant cartilage growth and the pain decreased. Kuroda et al. have also transplanted bone marrow MSCs, incorporated in a collagen gel, in an athlete with a grade IV cartilage defect [6]. After seven months, the athlete has regained its normal activity and histologic analysis reveals the formation of hyaline cartilage within the defect. The third study is a three case report where undifferentiated bone marrow-derived MSCs were associated with collagen sheet [147]. After 1 year, the three patients presented an increase IKDC score (International Knee Documentation Committee) and in 2 patients, a fibrocartilage-like repaired tissue totally recovered the defect [6]. Although the results suggest that autologous MSC transplantation may be an effective approach to promote the repair of articular cartilage defects, improvement is needed to achieve hyaline cartilage formation. Currently, only one Phase I clinical trial in cartilage tissue engineering using MSCs is underway. This clinical trial is sponsored by the Royan Institute and Tehran University of Medical Sciences and concerns the use of bone marrow MSCs mixed with type I collagen scaffolds and implanted in patients with knee cartilage defect or osteoarthritis.
CONCLUSIONS AND FUTURE PERSPECTIVES
In conclusion, cell-based strategies not only have proved the feasibility of such approaches for cartilage repair but also have provided good clinical results. However, these protocols are still far from generating a tissue that is comparable to native cartilage with respect to quality and stability. The use of chondrogenic progenitors, in particular MSCs, will undoubtedly be of high potential for such application. Nevertheless, more sophisticated approaches combining deliverable bioactive factors together with a chondro-conductive scaffold will be required (Fig. 2). Although some growth factors have been proposed, none are capable to specifically induce the desired lineage and a timely regulated combination of factors is likely to be required for the obtention of a functional and stable chondrocyte phenotype. These will rely on the understanding on the complex molecular events involved in chondrogenesis induction and maintenance of the chondrocyte phenotype taking place during embryogenesis that will have to be reproduced in adult tissue repair.
Figure 2.
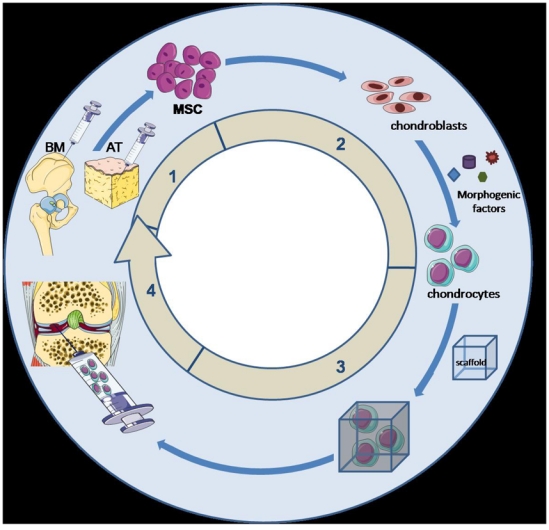
Sequence of events from stem cell isolation to engineered scaffold implantation. The first step is the isolation of mesenchymal stem cell (MSC) from bone marrow (BM) or adipose tissue (AT) and ex vivo expansion. This step is followed by the second and third steps of MSC differentiation towards chondrocytes by the addition of morphogenic factors and inclusion into a scaffold, preferentially an injectable biomaterial that allows an easy and minimally invasive injection of bioengineered scaffold into the cartilage defect as shown in step 4. Illustration with permission from Servier (http://www.servier.fr/smart/TermsOfUse.asp )
Acknowledgments
This work was supported by grants from the “Fondation de l’Avenir pour la Recherche Médicale Appliquée”, the “Fondation pour la Recherche Médicale”, the “Arthritis-Fondation Courtin”, the “Société Française de Rhumatologie”, Graftys SARL and ANR “Scartifold” and ANR “Chondrograft”.
References
- 1.Alsalameh S, Amin R, Gemba T, Lotz M. Identification of mesenchymal progenitor cells in normal and osteoarthritic human articular cartilage. Arthritis Rheum. 2004;50:1522–32. doi: 10.1002/art.20269. [DOI] [PubMed] [Google Scholar]
- 2.Poole AR, Kojima T, Yasuda T, Mwale F, Kobayashi M, Laverty S. Composition and structure of articular cartilage: a template for tissue repair. Clin Orthop Relat Res. 2001:S26–33. doi: 10.1097/00003086-200110001-00004. [DOI] [PubMed] [Google Scholar]
- 3.Goldring MB. Update on the biology of the chondrocyte and new approaches to treating cartilage diseases. Best Pract Res Clin Rheumatol. 2006;20:1003–25. doi: 10.1016/j.berh.2006.06.003. [DOI] [PubMed] [Google Scholar]
- 4.Smith GD, Knutsen G, Richardson JB. A clinical review of cartilage repair techniques. J Bone Joint Surg Br. 2005;87:445–9. doi: 10.1302/0301-620X.87B4.15971. [DOI] [PubMed] [Google Scholar]
- 5.Klein TJ, Schumacher BL, Schmidt TA, et al. Tissue engineering of stratified articular cartilage from chondrocyte subpopulations. Osteoarthritis Cartilage. 2003;11:595–602. doi: 10.1016/s1063-4584(03)00090-6. [DOI] [PubMed] [Google Scholar]
- 6.Kuroda R, Ishida K, Matsumoto T, et al. Treatment of a full-thickness articular cartilage defect in the femoral condyle of an athlete with autologous bone-marrow stromal cells. Osteoarthritis Cartilage. 2007;15:226–31. doi: 10.1016/j.joca.2006.08.008. [DOI] [PubMed] [Google Scholar]
- 7.Wood JJ, Malek MA, Frassica FJ, et al. Autologous cultured chondrocytes: adverse events reported to the United States Food and Drug Administration. J Bone Joint Surg Am. 2006;88:503–7. doi: 10.2106/JBJS.E.00103. [DOI] [PubMed] [Google Scholar]
- 8.Isogai N, Kusuhara H, Ikada Y, et al. Comparison of different chondrocytes for use in tissue engineering of cartilage model structures. Tissue Eng. 2006;12:691–703. doi: 10.1089/ten.2006.12.691. [DOI] [PubMed] [Google Scholar]
- 9.Kafienah W, Jakob M, Demarteau O, et al. Three-dimensional tissue engineering of hyaline cartilage: comparison of adult nasal and articular chondrocytes. Tissue Eng. 2002;8:817–26. doi: 10.1089/10763270260424178. [DOI] [PubMed] [Google Scholar]
- 10.Darling EM, Athanasiou KA. Rapid phenotypic changes in passaged articular chondrocyte subpopulations. J Orthop Res. 2005;23:425–32. doi: 10.1016/j.orthres.2004.08.008. [DOI] [PubMed] [Google Scholar]
- 11.Schnabel M, Marlovits S, Eckhoff G, et al. Dedifferentiation-associated changes in morphology and gene expression in primary human articular chondrocytes in cell culture. Osteoarthritis Cartilage. 2002;10:62–70. doi: 10.1053/joca.2001.0482. [DOI] [PubMed] [Google Scholar]
- 12.Domm C, Schunke M, Christesen K, Kurz B. Redifferentiation of dedifferentiated bovine articular chondrocytes in alginate culture under low oxygen tension. Osteoarthritis Cartilage. 2002;10:13–22. doi: 10.1053/joca.2001.0477. [DOI] [PubMed] [Google Scholar]
- 13.Malda J, van Blitterswijk CA, Grojec M, Martens DE, Tramper J, Riesle J. Expansion of bovine chondrocytes on microcarriers enhances redifferentiation. Tissue Eng. 2003;9:939–48. doi: 10.1089/107632703322495583. [DOI] [PubMed] [Google Scholar]
- 14.Fukui N, Purple CR, Sandell LJ. Cell biology of osteoarthritis: the chondrocyte’s response to injury. Curr Rheumatol Rep. 2001;3:496–505. doi: 10.1007/s11926-001-0064-8. [DOI] [PubMed] [Google Scholar]
- 15.Sandell LJ, Aigner T. Articular cartilage and changes in arthritis. An introduction: cell biology of osteoarthritis. Arthritis Res. 2001;3:107–13. doi: 10.1186/ar148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Tallheden T, Bengtsson C, Brantsing C, et al. Proliferation and differentiation potential of chondrocytes from osteoarthritic patients. Arthritis Res Ther. 2005;7:R560–568. doi: 10.1186/ar1709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Chen Y, Shao JZ, Xiang LX, Dong XJ, Zhang GR. Mesenchymal stem cells: a promising candidate in regenerative medicine. Int J Biochem Cell Biol. 2008;40:815–20. doi: 10.1016/j.biocel.2008.01.007. [DOI] [PubMed] [Google Scholar]
- 18.Friedenstein AJ, Chailakhjan RK, Lalykina KS. The development of fibroblast colonies in monolayer cultures of guinea-pig bone marrow and spleen cells. Cell Tissue Kinet. 1970;3:393–403. doi: 10.1111/j.1365-2184.1970.tb00347.x. [DOI] [PubMed] [Google Scholar]
- 19.Gronthos S, Simmons PJ. The biology and application of human bone marrow stromal cell precursors. J Hematother. 1996;5:15–23. doi: 10.1089/scd.1.1996.5.15. [DOI] [PubMed] [Google Scholar]
- 20.Dominici M, Le Blanc K, Mueller I, et al. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy. 2006;8:315–7. doi: 10.1080/14653240600855905. [DOI] [PubMed] [Google Scholar]
- 21.Pittenger M, Vanguri P, Simonetti D, Young R. Adult mesenchymal stem cells: potential for muscle and tendon regeneration and use in gene therapy. J Musculoskelet Neuronal Interact. 2002;2:309–20. [PubMed] [Google Scholar]
- 22.Makino S, Fukuda K, Miyoshi S, et al. Cardiomyocytes can be generated from marrow stromal cells in vitro. J Clin Invest. 1999;103:697–705. doi: 10.1172/JCI5298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Phinney DG, Isakova I. Plasticity and therapeutic potential of mesenchymal stem cells in the nervous system. Curr Pharm Des. 2005;11:1255–65. doi: 10.2174/1381612053507495. [DOI] [PubMed] [Google Scholar]
- 24.Tropel P, Platet N, Platel JC, et al. Functional neuronal differentiation of bone marrow-derived mesenchymal stem cells. Stem Cells. 2006;24:2868–76. doi: 10.1634/stemcells.2005-0636. [DOI] [PubMed] [Google Scholar]
- 25.Bhatia R, Hare JM. Mesenchymal stem cells: future source for reparative medicine. Congest Heart Fail. 2005;11:87–91. doi: 10.1111/j.1527-5299.2005.03618.x. quiz 92–83. [DOI] [PubMed] [Google Scholar]
- 26.Djouad F, Charbonnier LM, Bouffi C, et al. Mesenchymal stem cells inhibit the differentiation of dendritic cells through an interleukin-6-dependent mechanism. Stem Cells. 2007;25:2025–32. doi: 10.1634/stemcells.2006-0548. [DOI] [PubMed] [Google Scholar]
- 27.Reyes M, Lund T, Lenvik T, Aguiar D, Koodie L, Verfaillie CM. Purification and ex vivo expansion of postnatal human marrow mesodermal progenitor cells. Blood. 2001;98:2615–25. doi: 10.1182/blood.v98.9.2615. [DOI] [PubMed] [Google Scholar]
- 28.D’Ippolito G, Diabira S, Howard GA, Menei P, Roos BA, Schiller PC. Marrow-isolated adult multilineage inducible (MIAMI) cells, a unique population of postnatal young and old human cells with extensive expansion and differentiation potential. J Cell Sci. 2004;117:2971–81. doi: 10.1242/jcs.01103. [DOI] [PubMed] [Google Scholar]
- 29.Grimaud E, Heymann D, Redini F. Recent advances in TGF-beta effects on chondrocyte metabolism. Potential therapeutic roles of TGF-beta in cartilage disorders. Cytokine Growth Factor Rev. 2002;13:241–57. doi: 10.1016/s1359-6101(02)00004-7. [DOI] [PubMed] [Google Scholar]
- 30.Mackay AM, Beck SC, Murphy JM, Barry FP, Chichester CO, Pittenger MF. Chondrogenic differentiation of cultured human mesenchymal stem cells from marrow. Tissue Eng. 1998;4:415–28. doi: 10.1089/ten.1998.4.415. [DOI] [PubMed] [Google Scholar]
- 31.Johnstone B, Hering TM, Caplan AI, Goldberg VM, Yoo JU. In vitro chondrogenesis of bone marrow-derived mesenchymal progenitor cells. Exp Cell Res. 1998;238:265–72. doi: 10.1006/excr.1997.3858. [DOI] [PubMed] [Google Scholar]
- 32.Facchini A, Lisignoli G, Cristino S, et al. Human chondrocytes and mesenchymal stem cells grown onto engineered scaffold. Biorheology. 2006;43:471–80. [PubMed] [Google Scholar]
- 33.Choi JJ, Yoo SA, Park SJ, et al. Mesenchymal stem cells overexpressing interleukin-10 attenuate collagen-induced arthritis in mice. Clin Exp Immunol. 2008;153:269–76. doi: 10.1111/j.1365-2249.2008.03683.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Di Cesare PE, Frenkel SR, Carlson CS, Fang C, Liu C. Regional gene therapy for full-thickness articular cartilage lesions using naked DNA with a collagen matrix. J Orthop Res. 2006;24:1118–27. doi: 10.1002/jor.20143. [DOI] [PubMed] [Google Scholar]
- 35.Glansbeek HL, van Beuningen HM, Vitters EL, van der Kraan PM, van den Berg WB. Stimulation of articular cartilage repair in established arthritis by local administration of transforming growth factor-beta into murine knee joints. Lab Invest. 1998;78:133–42. [PubMed] [Google Scholar]
- 36.van Beuningen HM, van der Kraan PM, Arntz OJ, van den Berg WB. Transforming growth factor-beta 1 stimulates articular chondrocyte proteoglycan synthesis and induces osteophyte formation in the murine knee joint. Lab Invest. 1994;71:279–90. [PubMed] [Google Scholar]
- 37.Blaney Davidson EN, Vitters EL, van Beuningen HM, van de Loo FA, van den Berg WB, van der Kraan PM. Resemblance of osteophytes in experimental osteoarthritis to transforming growth factor beta-induced osteophytes: limited role of bone morphogenetic protein in early osteoarthritic osteophyte formation. Arthritis Rheum. 2007;56:4065–73. doi: 10.1002/art.23034. [DOI] [PubMed] [Google Scholar]
- 38.Chen D, Zhao M, Mundy GR. Bone morphogenetic proteins. Growth Factors. 2004;22:233–41. doi: 10.1080/08977190412331279890. [DOI] [PubMed] [Google Scholar]
- 39.Majumdar MK, Wang E, Morris EA. BMP-2 and BMP-9 promotes chondrogenic differentiation of human multipotential mesenchymal cells and overcomes the inhibitory effect of IL-1. J Cell Physiol. 2001;189:275–84. doi: 10.1002/jcp.10025. [DOI] [PubMed] [Google Scholar]
- 40.Gruber R, Mayer C, Bobacz K, et al. Effects of cartilage-derived morphogenetic proteins and osteogenic protein-1 on osteochondrogenic differentiation of periosteum-derived cells. Endocrinology. 2001;142:2087–94. doi: 10.1210/endo.142.5.8163. [DOI] [PubMed] [Google Scholar]
- 41.Jorgensen C, Gordeladze J, Noel D. Tissue engineering through autologous mesenchymal stem cells. Curr Opin Biotechnol. 2004;15:406–10. doi: 10.1016/j.copbio.2004.08.003. [DOI] [PubMed] [Google Scholar]
- 42.Sekiya I, Larson BL, Vuoristo JT, Reger RL, Prockop DJ. Comparison of effect of BMP-2, -4, and -6 on in vitro cartilage formation of human adult stem cells from bone marrow stroma. Cell Tissue Res. 2005;320:269–76. doi: 10.1007/s00441-004-1075-3. [DOI] [PubMed] [Google Scholar]
- 43.Grunder T, Gaissmaier C, Fritz J, et al. Bone morphogenetic protein (BMP)-2 enhances the expression of type II collagen and aggrecan in chondrocytes embedded in alginate beads. Osteoarthritis Cartilage. 2004;12:559–67. doi: 10.1016/j.joca.2004.04.001. [DOI] [PubMed] [Google Scholar]
- 44.Kuo AC, Rodrigo JJ, Reddi AH, Curtiss S, Grotkopp E, Chiu M. Microfracture and bone morphogenetic protein 7 (BMP-7) synergistically stimulate articular cartilage repair. Osteoarthritis Cartilage. 2006;14:1126–35. doi: 10.1016/j.joca.2006.04.004. [DOI] [PubMed] [Google Scholar]
- 45.Kuroda R, Usas A, Kubo S, et al. Cartilage repair using bone morphogenetic protein 4 and muscle-derived stem cells. Arthritis Rheum. 2006;54:433–42. doi: 10.1002/art.21632. [DOI] [PubMed] [Google Scholar]
- 46.Gelse K, Muhle C, Franke O, et al. Cell-based resurfacing of large cartilage defects: long-term evaluation of grafts from autologous transgene-activated periosteal cells in a porcine model of osteoarthritis. Arthritis Rheum. 2008;58:475–88. doi: 10.1002/art.23124. [DOI] [PubMed] [Google Scholar]
- 47.Noel D, Gazit D, Bouquet C, et al. Short-term BMP-2 expression is sufficient for in vivo osteochondral differentiation of mesenchymal stem cells. Stem Cells. 2004;22:74–85. doi: 10.1634/stemcells.22-1-74. [DOI] [PubMed] [Google Scholar]
- 48.Hannallah D, Peng H, Young B, Usas A, Gearhart B, Huard J. Retroviral delivery of Noggin inhibits the formation of heterotopic ossification induced by BMP-4, demineralized bone matrix, and trauma in an animal model. J Bone Joint Surg Am. 2004;86-A:80–91. doi: 10.2106/00004623-200401000-00013. [DOI] [PubMed] [Google Scholar]
- 49.Jonsson H, Allen P, Peng SL. Inflammatory arthritis requires Foxo3a to prevent Fas ligand-induced neutrophil apoptosis. Nat Med. 2005;11:666–71. doi: 10.1038/nm1248. [DOI] [PubMed] [Google Scholar]
- 50.Kikuchi A, Yamamoto H, Kishida S. Multiplicity of the interactions of Wnt proteins and their receptors. Cell Signal. 2007;19:659–71. doi: 10.1016/j.cellsig.2006.11.001. [DOI] [PubMed] [Google Scholar]
- 51.Macsai CE, Foster BK, Xian CJ. Roles of Wnt signalling in bone growth, remodelling, skeletal disorders and fracture repair. J Cell Physiol. 2008;215:578–87. doi: 10.1002/jcp.21342. [DOI] [PubMed] [Google Scholar]
- 52.Church V, Nohno T, Linker C, Marcelle C, Francis-West P. Wnt regulation of chondrocyte differentiation. J Cell Sci. 2002;115:4809–18. doi: 10.1242/jcs.00152. [DOI] [PubMed] [Google Scholar]
- 53.Rudnicki JA, Brown AM. Inhibition of chondrogenesis by Wnt gene expression in vivo and in vitro. Dev Biol. 1997;185:104–18. doi: 10.1006/dbio.1997.8536. [DOI] [PubMed] [Google Scholar]
- 54.Andrade AC, Nilsson O, Barnes KM, Baron J. Wnt gene expression in the post-natal growth plate: regulation with chondrocyte differentiation. Bone. 2007;40:1361–9. doi: 10.1016/j.bone.2007.01.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Day TF, Guo X, Garrett-Beal L, Yang Y. Wnt/beta-catenin signaling in mesenchymal progenitors controls osteoblast and chondrocyte differentiation during vertebrate skeletogenesis. Dev Cell. 2005;8:739–50. doi: 10.1016/j.devcel.2005.03.016. [DOI] [PubMed] [Google Scholar]
- 56.Hill TP, Spater D, Taketo MM, Birchmeier W, Hartmann C. Canonical Wnt/beta-catenin signaling prevents osteoblasts from differentiating into chondrocytes. Dev Cell. 2005;8:727–38. doi: 10.1016/j.devcel.2005.02.013. [DOI] [PubMed] [Google Scholar]
- 57.Chen Y, Whetstone HC, Youn A, et al. Beta-catenin signaling pathway is crucial for bone morphogenetic protein 2 to induce new bone formation. J Biol Chem. 2007;282:526–33. doi: 10.1074/jbc.M602700200. [DOI] [PubMed] [Google Scholar]
- 58.Ornitz DM, Itoh N. Fibroblast growth factors. Genome Biol. 2001;2:REVIEWS3005. doi: 10.1186/gb-2001-2-3-reviews3005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Liu Z, Xu J, Colvin JS, Ornitz DM. Coordination of chondrogenesis and osteogenesis by fibroblast growth factor 18. Genes Dev. 2002;16:859–69. doi: 10.1101/gad.965602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Ornitz DM. FGF signaling in the developing endochondral skeleton. Cytokine Growth Factor Rev. 2005;16:205–13. doi: 10.1016/j.cytogfr.2005.02.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Hoffmann A, Czichos S, Kaps C, et al. The T-box transcription factor Brachyury mediates cartilage development in mesenchymal stem cell line C3H10T1/2. J Cell Sci. 2002;115:769–81. doi: 10.1242/jcs.115.4.769. [DOI] [PubMed] [Google Scholar]
- 62.Davidson D, Blanc A, Filion D, et al. Fibroblast growth factor (FGF) 18 signals through FGF receptor 3 to promote chondrogenesis. J Biol Chem. 2005;280:20509–15. doi: 10.1074/jbc.M410148200. [DOI] [PubMed] [Google Scholar]
- 63.Weizmann S, Tong A, Reich A, Genina O, Yayon A, Monsonego-Ornan E. FGF upregulates osteopontin in epiphyseal growth plate chondrocytes: implications for endochondral ossification. Matrix Biol. 2005;24:520–9. doi: 10.1016/j.matbio.2005.07.003. [DOI] [PubMed] [Google Scholar]
- 64.Solchaga LA, Penick K, Porter JD, Goldberg VM, Caplan AI, Welter JF. FGF-2 enhances the mitotic and chondrogenic potentials of human adult bone marrow-derived mesenchymal stem cells. J Cell Physiol. 2005;203:398–409. doi: 10.1002/jcp.20238. [DOI] [PubMed] [Google Scholar]
- 65.Stewart AA, Byron CR, Pondenis H, Stewart MC. Effect of fibroblast growth factor-2 on equine mesenchymal stem cell monolayer expansion and chondrogenesis. Am J Vet Res. 2007;68:941–5. doi: 10.2460/ajvr.68.9.941. [DOI] [PubMed] [Google Scholar]
- 66.Takafuji H, Suzuki T, Okubo Y, Fujimura K, Bessho K. Regeneration of articular cartilage defects in the temporomandibular joint of rabbits by fibroblast growth factor-2: a pilot study. Int J Oral Maxillofac Surg. 2007;36:934–7. doi: 10.1016/j.ijom.2007.06.007. [DOI] [PubMed] [Google Scholar]
- 67.Ishii I, Mizuta H, Sei A, Hirose J, Kudo S, Hiraki Y. Healing of full-thickness defects of the articular cartilage in rabbits using fibroblast growth factor-2 and a fibrin sealant. J Bone Joint Surg Br. 2007;89:693–700. doi: 10.1302/0301-620X.89B5.18450. [DOI] [PubMed] [Google Scholar]
- 68.Moore EE, Bendele AM, Thompson DL, et al. Fibroblast growth factor-18 stimulates chondrogenesis and cartilage repair in a rat model of injury-induced osteoarthritis. Osteoarthritis Cartilage. 2005;13:623–31. doi: 10.1016/j.joca.2005.03.003. [DOI] [PubMed] [Google Scholar]
- 69.Dupont J, Holzenberger M. Biology of insulin-like growth factors in development. Birth Defects Res C Embryo Today. 2003;69:257–71. doi: 10.1002/bdrc.10022. [DOI] [PubMed] [Google Scholar]
- 70.Woods KA, Camacho-Hubner C, Savage MO, Clark AJ. Intrauterine growth retardation and postnatal growth failure associated with deletion of the insulin-like growth factor I gene. N Engl J Med. 1996;335:1363–7. doi: 10.1056/NEJM199610313351805. [DOI] [PubMed] [Google Scholar]
- 71.Davies LC, Blain EJ, Gilbert SJ, Caterson B, Duance VC. The Potential of IGF-1 and TGFbeta1 for Promoting “Adult” Articular Cartilage Repair: An In Vitro Study. Tissue Eng Part A. 2008 doi: 10.1089/ten.tea.2007.0211. [DOI] [PubMed] [Google Scholar]
- 72.Yoon DM, Fisher JP. Effects of Exogenous IGF-1 Delivery on the Early Expression of IGF-1 Signaling Molecules by Alginate Embedded Chondrocytes. Tissue Eng Part A. 2008;14:1263–73. doi: 10.1089/ten.tea.2007.0172. [DOI] [PubMed] [Google Scholar]
- 73.Uebersax L, Merkle HP, Meinel L. Insulin-like growth factor I releasing silk fibroin scaffolds induce chondrogenic differentiation of human mesenchymal stem cells. J Control Release. 2008;127:12–21. doi: 10.1016/j.jconrel.2007.11.006. [DOI] [PubMed] [Google Scholar]
- 74.Schmidt MB, Chen EH, Lynch SE. A review of the effects of insulin-like growth factor and platelet derived growth factor on in vivo cartilage healing and repair. Osteoarthritis Cartilage. 2006;14:403–12. doi: 10.1016/j.joca.2005.10.011. [DOI] [PubMed] [Google Scholar]
- 75.Bianco P, Riminucci M, Gronthos S, Robey PG. Bone marrow stromal stem cells: nature, biology, and potential applications. Stem Cells. 2001;19:180–92. doi: 10.1634/stemcells.19-3-180. [DOI] [PubMed] [Google Scholar]
- 76.Ehlen HW, Buelens LA, Vortkamp A. Hedgehog signaling in skeletal development. Birth Defects Res C Embryo Today. 2006;78:267–79. doi: 10.1002/bdrc.20076. [DOI] [PubMed] [Google Scholar]
- 77.Warzecha J, Gottig S, Bruning C, Lindhorst E, Arabmothlagh M, Kurth A. Sonic hedgehog protein promotes proliferation and chondrogenic differentiation of bone marrow-derived mesenchymal stem cells in vitro. J Orthop Sci. 2006;11:491–6. doi: 10.1007/s00776-006-1058-1. [DOI] [PubMed] [Google Scholar]
- 78.Fontaine C, Cousin W, Plaisant M, Dani C, Peraldi P. Hedgehog signaling alters adipocyte maturation of human mesenchymal stem cells. Stem Cells. 2008;26:1037–46. doi: 10.1634/stemcells.2007-0974. [DOI] [PubMed] [Google Scholar]
- 79.Edwards PC, Ruggiero S, Fantasia J, et al. Sonic hedgehog gene-enhanced tissue engineering for bone regeneration. Gene Ther. 2005;12:75–86. doi: 10.1038/sj.gt.3302386. [DOI] [PubMed] [Google Scholar]
- 80.Nesic D, Whiteside R, Brittberg M, Wendt D, Martin I, Mainil-Varlet P. Cartilage tissue engineering for degenerative joint disease. Adv Drug Deliv Rev. 2006;58:300–22. doi: 10.1016/j.addr.2006.01.012. [DOI] [PubMed] [Google Scholar]
- 81.Drury JL, Mooney DJ. Hydrogels for tissue engineering: scaffold design variables and applications. Biomaterials. 2003;24:4337–51. doi: 10.1016/s0142-9612(03)00340-5. [DOI] [PubMed] [Google Scholar]
- 82.Lubiatowski P, Kruczynski J, Gradys A, Trzeciak T, Jaroszewski J. Articular cartilage repair by means of biodegradable scaffolds. Transplant Proc. 2006;38:320–2. doi: 10.1016/j.transproceed.2005.12.012. [DOI] [PubMed] [Google Scholar]
- 83.Wakitani S, Goto T, Young RG, Mansour JM, Goldberg VM, Caplan AI. Repair of large full-thickness articular cartilage defects with allograft articular chondrocytes embedded in a collagen gel. Tissue Eng. 1998;4:429–44. doi: 10.1089/ten.1998.4.429. [DOI] [PubMed] [Google Scholar]
- 84.Lee CR, Grodzinsky AJ, Hsu HP, Spector M. Effects of a cultured autologous chondrocyte-seeded type II collagen scaffold on the healing of a chondral defect in a canine model. J Orthop Res. 2003;21:272–81. doi: 10.1016/S0736-0266(02)00153-5. [DOI] [PubMed] [Google Scholar]
- 85.Wakitani S, Goto T, Pineda SJ, et al. Mesenchymal cell-based repair of large, full-thickness defects of articular cartilage. J Bone Joint Surg Am. 1994;76:579–92. doi: 10.2106/00004623-199404000-00013. [DOI] [PubMed] [Google Scholar]
- 86.McCoy JP, Jr, Schade WJ, Siegle RJ, Waldinger TP, Vanderveen EE, Swanson NA. Characterization of the humoral immune response to bovine collagen implants. Arch Dermatol. 1985;121:990–4. [PubMed] [Google Scholar]
- 87.Ochi M, Uchio Y, Kawasaki K, Wakitani S, Iwasa J. Transplantation of cartilage-like tissue made by tissue engineering in the treatment of cartilage defects of the knee. J Bone Joint Surg Br. 2002;84:571–8. doi: 10.1302/0301-620x.84b4.11947. [DOI] [PubMed] [Google Scholar]
- 88.Schneider U, Andereya S. First results of a prospective randomized clinical trial on traditional chondrocyte transplantation vs CaReS-Technology. Z Orthop Ihre Grenzgeb. 2003;141:496–7. [PubMed] [Google Scholar]
- 89.Sims CD, Butler PE, Cao YL, et al. Tissue engineered neocartilage using plasma derived polymer substrates and chondrocytes. Plast Reconstr Surg. 1998;101:1580–5. doi: 10.1097/00006534-199805000-00022. [DOI] [PubMed] [Google Scholar]
- 90.Hunziker EB. Articular cartilage repair: basic science and clinical progress. A review of the current status and prospects. Osteoarthritis Cartilage. 2002;10:432–63. doi: 10.1053/joca.2002.0801. [DOI] [PubMed] [Google Scholar]
- 91.Fussenegger M, Meinhart J, Hobling W, Kullich W, Funk S, Bernatzky G. Stabilized autologous fibrin-chondrocyte constructs for cartilage repair in vivo. Ann Plast Surg. 2003;51:493–8. doi: 10.1097/01.sap.0000067726.32731.E1. [DOI] [PubMed] [Google Scholar]
- 92.Peretti GM, Randolph MA, Villa MT, Buragas MS, Yaremchuk MJ. Cell-based tissue-engineered allogeneic implant for cartilage repair. Tissue Eng. 2000;6:567–76. doi: 10.1089/107632700750022206. [DOI] [PubMed] [Google Scholar]
- 93.van Susante JL, Buma P, Schuman L, Homminga GN, van den Berg WB, Veth RP. Resurfacing potential of heterologous chondrocytes suspended in fibrin glue in large full-thickness defects of femoral articular cartilage: an experimental study in the goat. Biomaterials. 1999;20:1167–75. doi: 10.1016/s0142-9612(97)00190-7. [DOI] [PubMed] [Google Scholar]
- 94.Vinatier C, Gauthier O, Masson M, et al. Nasal chondrocytes and fibrin sealant for cartilage tissue engineering. J Biomed Mater Res A. 2009;89:176–85. doi: 10.1002/jbm.a.31988. [DOI] [PubMed] [Google Scholar]
- 95.Hendrickson DA, Nixon AJ, Grande DA, et al. Chondrocyte-fibrin matrix transplants for resurfacing extensive articular cartilage defects. J Orthop Res. 1994;12:485–97. doi: 10.1002/jor.1100120405. [DOI] [PubMed] [Google Scholar]
- 96.Wilke MM, Nydam DV, Nixon AJ. Enhanced early chondrogenesis in articular defects following arthroscopic mesenchymal stem cell implantation in an equine model. J Orthop Res. 2007;25:913–25. doi: 10.1002/jor.20382. [DOI] [PubMed] [Google Scholar]
- 97.Peterson L, Brittberg M, Kiviranta I, Akerlund EL, Lindahl A. Autologous chondrocyte transplantation. Biomechanics and long-term durability. Am J Sports Med. 2002;30:2–12. doi: 10.1177/03635465020300011601. [DOI] [PubMed] [Google Scholar]
- 98.Awad HA, Wickham MQ, Leddy HA, Gimble JM, Guilak F. Chondrogenic differentiation of adipose-derived adult stem cells in agarose, alginate, and gelatin scaffolds. Biomaterials. 2004;25:3211–22. doi: 10.1016/j.biomaterials.2003.10.045. [DOI] [PubMed] [Google Scholar]
- 99.Diduch DR, Jordan LC, Mierisch CM, Balian G. Marrow stromal cells embedded in alginate for repair of osteochondral defects. Arthroscopy. 2000;16:571–7. doi: 10.1053/jars.2000.4827. [DOI] [PubMed] [Google Scholar]
- 100.Rahfoth B, Weisser J, Sternkopf F, Aigner T, von der Mark K, Brauer R. Transplantation of allograft chondrocytes embedded in agarose gel into cartilage defects of rabbits. Osteoarthritis Cartilage. 1998;6:50–65. doi: 10.1053/joca.1997.0092. [DOI] [PubMed] [Google Scholar]
- 101.Bonaventure J, Kadhom N, Cohen-Solal L, et al. Reexpression of cartilage-specific genes by dedifferentiated human articular chondrocytes cultured in alginate beads. Exp Cell Res. 1994;212:97–104. doi: 10.1006/excr.1994.1123. [DOI] [PubMed] [Google Scholar]
- 102.Carossino AM, Recenti R, Carossino R, et al. Methodological models for in vitro amplification and maintenance of human articular chondrocytes from elderly patients. Biogerontology. 2007 doi: 10.1007/s10522-007-9088-4. [DOI] [PubMed] [Google Scholar]
- 103.Almqvist KF, Wang L, Wang J, et al. Culture of chondrocytes in alginate surrounded by fibrin gel: characteristics of the cells over a period of eight weeks. Ann Rheum Dis. 2001;60:781–90. doi: 10.1136/ard.60.8.781. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Erickson GR, Gimble JM, Franklin DM, Rice HE, Awad H, Guilak F. Chondrogenic potential of adipose tissue-derived stromal cells in vitro and in vivo. Biochem Biophys Res Commun. 2002;290:763–9. doi: 10.1006/bbrc.2001.6270. [DOI] [PubMed] [Google Scholar]
- 105.Fragonas E, Valente M, Pozzi-Mucelli M, et al. Articular cartilage repair in rabbits by using suspensions of allogenic chondrocytes in alginate. Biomaterials. 2000;21:795–801. doi: 10.1016/s0142-9612(99)00241-0. [DOI] [PubMed] [Google Scholar]
- 106.Mierisch CM, Wilson HA, Turner MA, et al. Chondrocyte transplantation into articular cartilage defects with use of calcium alginate: the fate of the cells. J Bone Joint Surg Am. 2003;85-A:1757–67. doi: 10.2106/00004623-200309000-00015. [DOI] [PubMed] [Google Scholar]
- 107.Paige KT, Cima LG, Yaremchuk MJ, Schloo BL, Vacanti JP, Vacanti CA. De novo cartilage generation using calcium alginate-chondrocyte constructs. Plast Reconstr Surg. 1996;97:168–178. doi: 10.1097/00006534-199601000-00027. discussion 179–80. [DOI] [PubMed] [Google Scholar]
- 108.Selmi TA, Verdonk P, Chambat P, et al. Autologous chondrocyte implantation in a novel alginate-agarose hydrogel: outcome at two years. J Bone Joint Surg Br. 2008;90:597–604. doi: 10.1302/0301-620X.90B5.20360. [DOI] [PubMed] [Google Scholar]
- 109.Knudson W, Casey B, Nishida Y, Eger W, Kuettner KE, Knudson CB. Hyaluronan oligosaccharides perturb cartilage matrix homeostasis and induce chondrocytic chondrolysis. Arthritis Rheum. 2000;43:1165–74. doi: 10.1002/1529-0131(200005)43:5<1165::AID-ANR27>3.0.CO;2-H. [DOI] [PubMed] [Google Scholar]
- 110.Tognana E, Padera RF, Chen F, Vunjak-Novakovic G, Freed LE. Development and remodeling of engineered cartilage-explant composites in vitro and in vivo. Osteoarthritis Cartilage. 2005;13:896–905. doi: 10.1016/j.joca.2005.05.003. [DOI] [PubMed] [Google Scholar]
- 111.Goa KL, Benfield P. Hyaluronic acid. A review of its pharmacology and use as a surgical aid in ophthalmology, and its therapeutic potential in joint disease and wound healing. Drugs. 1994;47:536–66. doi: 10.2165/00003495-199447030-00009. [DOI] [PubMed] [Google Scholar]
- 112.Bulpitt P, Aeschlimann D. New strategy for chemical modification of hyaluronic acid: preparation of functionalized derivatives and their use in the formation of novel biocompatible hydrogels. J Biomed Mater Res. 1999;47:152–69. doi: 10.1002/(sici)1097-4636(199911)47:2<152::aid-jbm5>3.0.co;2-i. [DOI] [PubMed] [Google Scholar]
- 113.Chang CH, Liu HC, Lin CC, Chou CH, Lin FH. Gelatin-chondroitin-hyaluronan tri-copolymer scaffold for cartilage tissue engineering. Biomaterials. 2003;24:4853–8. doi: 10.1016/s0142-9612(03)00383-1. [DOI] [PubMed] [Google Scholar]
- 114.Marcacci M, Berruto M, Brocchetta D, et al. Articular cartilage engineering with Hyalograft C: 3-year clinical results. Clin Orthop Relat Res. 2005 Jun;(435):96–105. doi: 10.1097/01.blo.0000165737.87628.5b. [DOI] [PubMed] [Google Scholar]
- 115.Montembault A, Tahiri K, Korwin-Zmijowska C, Chevalier X, Corvol MT, Domard A. A material decoy of biological media based on chitosan physical hydrogels: application to cartilage tissue engineering. Biochimie. 2006;88:551–64. doi: 10.1016/j.biochi.2006.03.002. [DOI] [PubMed] [Google Scholar]
- 116.Di Martino A, Sittinger M, Risbud MV. Chitosan: a versatile biopolymer for orthopaedic tissue-engineering. Biomaterials. 2005;26:5983–90. doi: 10.1016/j.biomaterials.2005.03.016. [DOI] [PubMed] [Google Scholar]
- 117.Hoemann CD, Sun J, Legare A, McKee MD, Buschmann MD. Tissue engineering of cartilage using an injectable and adhesive chitosan-based cell-delivery vehicle. Osteoarthritis Cartilage. 2005;13:318–29. doi: 10.1016/j.joca.2004.12.001. [DOI] [PubMed] [Google Scholar]
- 118.Muller FA, Muller L, Hofmann I, Greil P, Wenzel MM, Staudenmaier R. Cellulose-based scaffold materials for cartilage tissue engineering. Biomaterials. 2006;27:3955–63. doi: 10.1016/j.biomaterials.2006.02.031. [DOI] [PubMed] [Google Scholar]
- 119.Martson M, Viljanto J, Laippala P, Saukko P. Connective tissue formation in subcutaneous cellulose sponge implants in the rat. The effect of the size and cellulose content of the implant. Eur Surg Res. 1998;30:419–25. doi: 10.1159/000008608. [DOI] [PubMed] [Google Scholar]
- 120.Andrews GP, Gorman SP, Jones DS. Rheological characterisation of primary and binary interactive bioadhesive gels composed of cellulose derivatives designed as ophthalmic viscosurgical devices. Biomaterials. 2005;26:571–80. doi: 10.1016/j.biomaterials.2004.02.062. [DOI] [PubMed] [Google Scholar]
- 121.Vinatier C, Gauthier O, Fatimi A, et al. An injectable cellulose-based hydrogel for the transfer of autologous nasal chondrocytes in articular cartilage defects. Biotechnol Bioeng. 2009;102:1259–67. doi: 10.1002/bit.22137. [DOI] [PubMed] [Google Scholar]
- 122.Sittinger M, Reitzel D, Dauner M, et al. Resorbable polyesters in cartilage engineering: affinity and biocompatibility of polymer fiber structures to chondrocytes. J Biomed Mater Res. 1996;33:57–63. doi: 10.1002/(SICI)1097-4636(199622)33:2<57::AID-JBM1>3.0.CO;2-K. [DOI] [PubMed] [Google Scholar]
- 123.Spain TL, Agrawal CM, Athanasiou KA. New technique to extend the useful life of a biodegradable cartilage implant. Tissue Eng. 1998;4:343–52. doi: 10.1089/ten.1998.4.343. [DOI] [PubMed] [Google Scholar]
- 124.Galois L, Freyria AM, Herbage D, Mainard D. Cartilage tissue engineering: state-of-the-art and future approaches. Pathol Biol (Paris) 2005;53:590–8. doi: 10.1016/j.patbio.2004.12.019. [DOI] [PubMed] [Google Scholar]
- 125.Wang X, Grogan SP, Rieser F, et al. Tissue engineering of biphasic cartilage constructs using various biodegradable scaffolds: an in vitro study. Biomaterials. 2004;25:3681–8. doi: 10.1016/j.biomaterials.2003.10.102. [DOI] [PubMed] [Google Scholar]
- 126.Freed LE, Vunjak-Novakovic G, Langer R. Cultivation of cell-polymer cartilage implants in bioreactors. J Cell Biochem. 1993;51:257–64. doi: 10.1002/jcb.240510304. [DOI] [PubMed] [Google Scholar]
- 127.Freed LE, Grande DA, Lingbin Z, Emmanual J, Marquis JC, Langer R. Joint resurfacing using allograft chondrocytes and synthetic biodegradable polymer scaffolds. J Biomed Mater Res. 1994;28:891–9. doi: 10.1002/jbm.820280808. [DOI] [PubMed] [Google Scholar]
- 128.Ossendorf C, Kaps C, Kreuz PC, Burmester GR, Sittinger M, Erggelet C. Treatment of posttraumatic and focal osteoarthritic cartilage defects of the knee with autologous polymer-based three-dimensional chondrocyte grafts: Two year clinical results. Arthritis Res Ther. 2007;9:R41. doi: 10.1186/ar2180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Carranza-Bencano A, Armas-Padron JR, Gili-Miner M, Lozano MA. Carbon fiber implants in osteochondral defects of the rabbit patella. Biomaterials. 2000;21:2171–6. doi: 10.1016/s0142-9612(00)00144-7. [DOI] [PubMed] [Google Scholar]
- 130.Messner K. Hydroxylapatite supported Dacron plugs for repair of isolated full-thickness osteochondral defects of the rabbit femoral condyle: mechanical and histological evaluations from 6–48 weeks. J Biomed Mater Res. 1993;27:1527–32. doi: 10.1002/jbm.820271209. [DOI] [PubMed] [Google Scholar]
- 131.Messner K. Durability of artificial implants for repair of osteochondral defects of the medial femoral condyle in rabbits. Biomaterials. 1994;15:657–64. doi: 10.1016/0142-9612(94)90163-5. [DOI] [PubMed] [Google Scholar]
- 132.Defrere J, Franckart A. Teflon/polyurethane arthroplasty of the knee: the first 2 years preliminary clinical experience in a new concept of artificial resurfacing of full thickness cartilage lesions of the knee. Acta Chir Belg. 1992;92:217–27. [PubMed] [Google Scholar]
- 133.Sontjens SH, Nettles DL, Carnahan MA, Setton LA, Grinstaff MW. Biodendrimer-based hydrogel scaffolds for cartilage tissue repair. Biomacromolecules. 2006;7:310–6. doi: 10.1021/bm050663e. [DOI] [PubMed] [Google Scholar]
- 134.Wallace DG, Rosenblatt J. Collagen gel systems for sustained delivery and tissue engineering. Adv Drug Deliv Rev. 2003;55:1631–49. doi: 10.1016/j.addr.2003.08.004. [DOI] [PubMed] [Google Scholar]
- 135.Temenoff JS, Mikos AG. Injectable biodegradable materials for orthopedic tissue engineering. Biomaterials. 2000;21:2405–12. doi: 10.1016/s0142-9612(00)00108-3. [DOI] [PubMed] [Google Scholar]
- 136.Kim IY, Seo SJ, Moon HS, et al. Chitosan and its derivatives for tissue engineering applications. Biotechnol Adv. 2008;26:1–21. doi: 10.1016/j.biotechadv.2007.07.009. [DOI] [PubMed] [Google Scholar]
- 137.Vinatier C, Magne D, Moreau A, et al. Engineering cartilage with human nasal chondrocytes and a silanized hydroxypropyl methylcellulose hydrogel. J Biomed Mater Res A. 2007;80:66–74. doi: 10.1002/jbm.a.30867. [DOI] [PubMed] [Google Scholar]
- 138.Insall J. The Pridie debridement operation for osteoarthritis of the knee. Clin Orthop Relat Res. 1974:61–7. [PubMed] [Google Scholar]
- 139.Steadman JR, Rodkey WG, Briggs KK, Rodrigo JJ. The microfracture technic in the management of complete cartilage defects in the knee joint. Orthopade. 1999;28:26–32. doi: 10.1007/s001320050318. [DOI] [PubMed] [Google Scholar]
- 140.Hangody L, Kish G, Karpati Z, Szerb I, Udvarhelyi I. Arthroscopic autogenous osteochondral mosaicplasty for the treatment of femoral condylar articular defects. A preliminary report. Knee Surg Sports Traumatol Arthrosc. 1997;5:262–7. doi: 10.1007/s001670050061. [DOI] [PubMed] [Google Scholar]
- 141.Brittberg M, Lindahl A, Nilsson A, Ohlsson C, Isaksson O, Peterson L. Treatment of deep cartilage defects in the knee with autologous chondrocyte transplantation. N Engl J Med. 1994;331:889–95. doi: 10.1056/NEJM199410063311401. [DOI] [PubMed] [Google Scholar]
- 142.Magnussen RA, Dunn WR, Carey JL, Spindler KP. Treatment of focal articular cartilage defects in the knee: a systematic review. Clin Orthop Relat Res. 2008;466:952–62. doi: 10.1007/s11999-007-0097-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Cherubino P, Grassi FA, Bulgheroni P, Ronga M. Autologous chondrocyte implantation using a bilayer collagen membrane: a preliminary report. J Orthop Surg (Hong Kong) 2003;11:10–5. doi: 10.1177/230949900301100104. [DOI] [PubMed] [Google Scholar]
- 144.Wakitani S, Kawaguchi A, Tokuhara Y, Takaoka K. Present status of and future direction for articular cartilage repair. J Bone Miner Metab. 2008;26:115–22. doi: 10.1007/s00774-007-0802-8. [DOI] [PubMed] [Google Scholar]
- 145.Wakitani S, Imoto K, Yamamoto T, Saito M, Murata N, Yoneda M. Human autologous culture expanded bone marrow mesenchymal cell transplantation for repair of cartilage defects in osteoarthritic knees. Osteoarthritis Cartilage. 2002;10:199–206. doi: 10.1053/joca.2001.0504. [DOI] [PubMed] [Google Scholar]
- 146.Centeno CJ, Busse D, Kisiday J, Keohan C, Freeman M, Karli D. Increased knee cartilage volume in degenerative joint disease using percutaneously implanted, autologous mesenchymal stem cells. Pain Physician. 2008;11:343–53. [PubMed] [Google Scholar]
- 147.Wakitani S, Nawata M, Tensho K, Okabe T, Machida H, Ohgushi H. Repair of articular cartilage defects in the patello-femoral joint with autologous bone marrow mesenchymal cell transplantation: three case reports involving nine defects in five knees. J Tissue Eng Regen Med. 2007;1:74–9. doi: 10.1002/term.8. [DOI] [PubMed] [Google Scholar]
- 148.Bradham DM, Passaniti A, Horton WE., Jr Mesenchymal cell chondrogenesis is stimulated by basement membrane matrix and inhibited by age-associated factors. Matrix Biol. 1995;14:561–71. doi: 10.1016/s0945-053x(05)80005-8. [DOI] [PubMed] [Google Scholar]
- 149.Bulic K. Articular chondrocytes interact with basement membrane Matrigel through laminin active binding sites. Acta Med Croatica. 1996;50:69–74. [PubMed] [Google Scholar]
- 150.Fan H, Hu Y, Li X, et al. Ectopic cartilage formation induced by mesenchymal stem cells on porous gelatin-chondroitin-hyaluronate scaffold containing microspheres loaded with TGF-beta1. Int J Artif Organs. 2006;29:602–11. doi: 10.1177/039139880602900610. [DOI] [PubMed] [Google Scholar]
- 151.Pulkkinen H, Tiitu V, Lammentausta E, et al. Cellulose sponge as a scaffold for cartilage tissue engineering. Biomed Mater Eng. 2006;16:S29–35. [PubMed] [Google Scholar]
- 152.Dounchis JS, Bae WC, Chen AC, Sah RL, Coutts RD, Amiel D. Cartilage repair with autogenic perichondrium cell and polylactic acid grafts. Clin Orthop Relat Res. 2000:248–64. doi: 10.1097/00003086-200008000-00033. [DOI] [PubMed] [Google Scholar]
- 153.Yan J, Qi N, Zhang Q. Rabbit articular chondrocytes seeded on collagen-chitosan-GAG scaffold for cartilage tissue engineering in vivo. Artif Cells Blood Substit Immobil Biotechnol. 2007;35:333–44. doi: 10.1080/10731190701460200. [DOI] [PubMed] [Google Scholar]
- 154.Grad S, Kupcsik L, Gorna K, Gogolewski S, Alini M. The use of biodegradable polyurethane scaffolds for cartilage tissue engineering: potential and limitations. Biomaterials. 2003;24:5163–71. doi: 10.1016/s0142-9612(03)00462-9. [DOI] [PubMed] [Google Scholar]
- 155.Kose GT, Korkusuz F, Ozkul A, et al. Tissue engineered cartilage on collagen and PHBV matrices. Biomaterials. 2005;26:5187–5197. doi: 10.1016/j.biomaterials.2005.01.037. [DOI] [PubMed] [Google Scholar]
- 156.Estrada LE, Dodge GR, Richardson DW, Farole A, Jimenez SA. Characterization of a biomaterial with cartilage-like properties expressing type X collagen generated in vitro using neonatal porcine articular and growth plate chondrocytes. Osteoarthritis Cartilage. 2001;9:169–77. doi: 10.1053/joca.2000.0373. [DOI] [PubMed] [Google Scholar]
- 157.Barbucci R, Lamponi S, Borzacchiello A, et al. Hyaluronic acid hydrogel in the treatment of osteoarthritis. Biomaterials. 2002;23:4503–13. doi: 10.1016/s0142-9612(02)00194-1. [DOI] [PubMed] [Google Scholar]
- 158.Liao E, Yaszemski M, Krebsbach P, Hollister S. Tissue-engineered cartilage constructs using composite hyaluronic acid/collagen I hydrogels and designed poly(propylene fumarate) scaffolds. Tissue Eng. 2007;13:537–50. doi: 10.1089/ten.2006.0117. [DOI] [PubMed] [Google Scholar]
- 159.Sharma B, Williams CG, Khan M, Manson P, Elisseeff JH. In vivo chondrogenesis of mesenchymal stem cells in a photopolymerized hydrogel. Plast Reconstr Surg. 2007;119:112–20. doi: 10.1097/01.prs.0000236896.22479.52. [DOI] [PubMed] [Google Scholar]
- 160.Alsberg E, Anderson KW, Albeiruti A, Rowley JA, Mooney DJ. Engineering growing tissues. Proc Natl Acad Sci U S A. 2002;99:12025–30. doi: 10.1073/pnas.192291499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Atala A, Cima LG, Kim W, et al. Injectable alginate seeded with chondrocytes as a potential treatment for vesicoureteral reflux. J Urol. 1993;150:745–7. doi: 10.1016/s0022-5347(17)35603-3. [DOI] [PubMed] [Google Scholar]
- 162.Elisseeff JH, Lee A, Kleinman HK, Yamada Y. Biological response of chondrocytes to hydrogels. Ann N Y Acad Sci. 2002;961:118–22. doi: 10.1111/j.1749-6632.2002.tb03062.x. [DOI] [PubMed] [Google Scholar]
- 163.Suh JK, Matthew HW. Application of chitosan-based polysaccharide biomaterials in cartilage tissue engineering: a review. Biomaterials. 2000;21:2589–98. doi: 10.1016/s0142-9612(00)00126-5. [DOI] [PubMed] [Google Scholar]
- 164.van Susante JLC, Pieper J, Buma P, et al. Linkage of chondroitin-sulfate to type I collagen scaffolds stimulates the bioactivity of seeded chondrocytes in vitro. Biomaterials. 2001;22:2359–69. doi: 10.1016/s0142-9612(00)00423-3. [DOI] [PubMed] [Google Scholar]
- 165.Taguchi T, Xu L, Kobayashi H, Taniguchi A, Kataoka K, Tanaka J. Encapsulation of chondrocytes in injectable alkali-treated collagen gels prepared using poly(ethylene glycol)-based 4-armed star polymer. Biomaterials. 2005;26:1247–52. doi: 10.1016/j.biomaterials.2004.04.029. [DOI] [PubMed] [Google Scholar]
- 166.Mercier NR, Costantino HR, Tracy MA, Bonassar LJ. A novel injectable approach for cartilage formation in vivo using PLG microspheres. Ann Biomed Eng. 2004;32:418–29. doi: 10.1023/b:abme.0000017547.84146.fd. [DOI] [PubMed] [Google Scholar]
- 167.Mercier NR, Costantino HR, Tracy MA, Bonassar LJ. Poly(lactide-co-glycolide) microspheres as a moldable scaffold for cartilage tissue engineering. Biomaterials. 2005;26:1945–52. doi: 10.1016/j.biomaterials.2004.06.030. [DOI] [PubMed] [Google Scholar]
- 168.Elisseeff J, Anseth K, Sims D, et al. Transdermal photopolymerization of poly(ethylene oxide)-based injectable hydrogels for tissue-engineered cartilage. Plast Reconstr Surg. 1999;104:1014–22. doi: 10.1097/00006534-199909040-00017. [DOI] [PubMed] [Google Scholar]
- 169.Elisseeff J, McIntosh W, Anseth K, Riley S, Ragan P, Langer R. Photoencapsulation of chondrocytes in poly(ethylene oxide)-based semi-interpenetrating networks. J Biomed Mater Res. 2000;51:164–71. doi: 10.1002/(sici)1097-4636(200008)51:2<164::aid-jbm4>3.0.co;2-w. [DOI] [PubMed] [Google Scholar]
- 170.Kim TK, Sharma B, Williams CG, et al. Experimental model for cartilage tissue engineering to regenerate the zonal organization of articular cartilage. Osteoarthritis Cartilage. 2003;11:653–64. doi: 10.1016/s1063-4584(03)00120-1. [DOI] [PubMed] [Google Scholar]
- 171.Burdick JA, Anseth KS. Photoencapsulation of osteoblasts in injectable RGD-modified PEG hydrogels for bone tissue engineering. Biomaterials. 2002;23:4315–23. doi: 10.1016/s0142-9612(02)00176-x. [DOI] [PubMed] [Google Scholar]