

Abstract
Damnacanthal, an anthraquinone compound, is isolated from the roots of Morinda citrifolia L. (noni), which has been used for traditional therapy in several chronic diseases including cancer. Although noni has been consumed for a long time in Asian and Polynesian countries, the molecular mechanisms by which it exerts several benefits are starting to emerge. In this report, we examined systematic approaches on the cancer suppressing capability of damnacanthal in colorectal tumorigenesis. Damnacanthal exhibits cell growth arrest as well as caspase activity induction in colorectal cancer cells. We also examined several potential target proteins and found that the pro-apoptotic protein Nonsteroidal anti-inflammatory activated gene-1 (NAG-1) is highly induced. Subsequently, we have found that damnacanthal also enhances transcription factor C/EBPβ, which controls NAG-1 transcriptional activity. Blocking of C/EBPβ by shRNA results in the reduction of NAG-1 expression as well as caspase activity in the presence of damnacanthal. Taken together, these results indicate that damnacanthal increases anti-tumorigenic activity in human colorectal cancer cells, and C/EBPβ plays a role in damnacanthal-induced NAG-1 expression.
Keywords: Damnacanthal, Noni, NAG-1, GDF15, C/EBPβ, Colorectal cancer
1. Introduction
Cancer is a major public health problem in the United States with an estimated 1.5 million new cancer cases expected in 2010 . It seems that there is still no satisfactory improvement in the success of cancer treatment although anti-cancer agents are constantly being introduced. Prevention research is a promising approach to reduce cancer incidence. In particular, chemoprevention using herbal medicine has been emerging due to its anti-cancer activity. Several medicinal plants and herbs, and their various phytochemical constituents including phenolics, flavonoids, carotenoids, and alkaloids confer protective effects against a wide range of cancers . Morinda citrifolia L. (Rubiaceae), commonly called noni, is one of the traditional folk medicinal plants that has a long history of use in Polynesia. Noni has been reported to have a broad range of health benefits for cancer, infection, diabetes, asthma, cough, hypertension, pain, ulcers, wounds, hemorrhoids and rheumatoid arthritis . Several mechanisms have been proposed to understand how noni manifests its anti-cancer effect, such as the inhibition of angiogenesis , prevention of carcinogen-DNA adduct formation or activation of immune responses. Subsequent studies using various animal models showed that noni has anti-tumorigenic activity in vivo .
Damnacanthal, an anthraquinone compound, was isolated from the roots of Morinda citrifolia L. and identified as a potent inhibitor of p56lck tyrosine kinase activity through high-volume screening of natural product extracts . Damnacanthal also exhibits cytotoxic activity against breast cancer cell lines as well as small cell lung cancer cell lines . In addition to its anti-cancer activity, damnacanthal exhibits anti-fungal activity against Candida albicans, and anti-tuberculosis activity against Mycobacterium tuberculosis . However, the molecular targets by which damnacanthal acts against several pathophysiological diseases have not been elucidated.
Non-steroidal anti-inflammatory drug (NSAID)-activated gene-1 (NAG-1, also known as GDF15) is a member of the transforming growth factor β superfamily, which has pro-apoptotic and anti-tumorigenic activities . It is believed that NAG-1 may serve as a molecular target of many phytochemicals . Transgenic mice (NAG-1Tg+/BL6, C57/BL6 background) expressing human NAG-1 have been developed by our group , and we have reported that NAG-1Tg+/BL6 mice are resistant to chemically- and genetically-induced intestinal polyp formation as well as lung tumors and inflammation . These data indicate that NAG-1 plays a pivotal role in colon and lung tumorigenesis in vivo. It also has been reported that several transcription factors contribute to NAG-1 expression at the transcription level, including EGR-1 , p53 , C/EBPβ , and ATF3 .
In the present study, we have shown that damnacanthal exhibits anti-tumorigenic activity as assessed by several in vitro assays. We report a novel finding of C/EBPβ induction by damnacanthal, followed by NAG-1 induction in human colorectal cancer cells. This sequential pathway seems to be mediated by the ERK pathway.
2. Materials and methods
2.1. Materials
Human colorectal cancer cell lines (HCT-116, SW480 and LoVo) were purchased from American Type Culture Collection (Manassas, VA). The cell culture media were purchased from Hyclone (Logan, UT) and Mediatech (Herndon, VA). NAG-1 antibody was previously described , and cyclin D1, p53, C/EBPβ and actin antibodies were purchased from Santa Cruz Biotechnology (Santa Cruz, CA). The p21 antibody was obtained from Cell Signaling Technology (Beverly, MA). All chemicals were purchased from Fisher Scientific (Pittsburgh, PA), unless otherwise specified.
2.2. Damnacanthal
Damnacanthal was extracted from the root of Morinda citrifolia L. in Thailand, as previously reported . The isolated pure compound was verified by various methods: 1) comparison of Rf value with reference standard checked by thin layer chromatography (TLC), 2) measuring the melting point of samples compared to the previous report, 3) investigating mass spectrum data by the mass spectroscopy with LC system coupled to a MicrOTOF mass spectrometer (Bruker Daltonics, Bremen, Germany) and determining 1H NMR and 13C NMR spectra determination by an INOVA-500 nuclear magnetic resonance spectroscopy.
2.3. Cell culture
HCT-116 cells were cultured in McCoy’s 5A medium. SW480 was maintained in RPMI1640 medium, and LoVo cells were maintained in Ham’s F-12 medium. All culture media were supplemented with 10% fetal bovine serum (Hyclone), 50 units/ml penicillin, and 50 μg/ml streptomycin.
2.4. Human NAG-1 promoters and expression vectors
The NAG-1 promoters and internal deletion constructs were described previously . The C/EBPβ, C/EBPα, C/EBPδ, RARα expression vectors were also described previously , as were the wild type and dominant negative constructs of ERK2 .
2.5. Cell proliferation analysis
The effect of damnacanthal on cell proliferation in HCT-116 and SW480 cells was investigated using the CellTiter 96 Aqueous One Solution Cell Proliferation Assay (Promega, Madison, WI). Cells were seeded at the concentration of 1000 cells/well for HCT-116 and 2000 cells/well for SW480 in 96-well tissue culture plates in four replicates. The cells were then treated with 100 nM, 1, 10, and 100 μM of damnacanthal in the presence of serum. At 1, 2, and 4 days after treatment, 20 μl of CellTiter 96 Aqueous One Solution was added to each well, and the plate was incubated for 1 h at 37 °C. Absorbance at 490 nm was recorded in an enzyme-linked immunosorbent assay (ELISA) plate reader (Bio-Tek Instruments, Winooski, VT).
2.6. Cell cycle analysis
Cells were plated at 4×105 cells/well in 6-well plates, incubated for 24 h, and then treated with 1, 10, and 50 μM of damnacanthal in the presence of serum for 72 h. The cells (attached and floating cells) were then harvested, washed with PBS, fixed by the slow addition of 1 ml cold 70% ethanol, and stored at −80 °C. The fixed cells were pelleted, wash with 50%, and 20% ethanol, followed by PBS, and stained with 0.5 ml of 70 μM propidium iodide containing 1 mg/ml RNase in PBS for 15 min at room temperature. Twenty thousand cells were examined by flow cytometry, using a Becton Dickinson Fluorescence-activated cell sorter (FACS) equipped with CellQuest software, by gating on an area versus width dot plot to exclude cell debris and cell aggregates.
2.7. Caspase 3/7 enzyme activity
Enzyme activity of caspase 3/7 was analyzed using the Caspase-Glo 3/7 Assay kit (Promega) according to the manufacturer’s protocol. The cells were harvested with radioimmunoprecipitation assay buffer (RIPA buffer) containing protease inhibitors, and the same volume of Caspase-Glo 3/7 reagent was added to the cell lysates (30 μg protein) in 96-well plates and incubated at room temperature in the dark for 1 h. The luminescence was measured using a FLX800 microplate reader (BioTek).
2.8. Clonogenic assay
The effect of damnacanthal on HCT-116 cell colony formation was investigated using clonogenic assay. HCT-116 cells were plated (in triplicates) in 12-well plates at 300 cells per well. After 24 h, cells were then treated with DMSO, 1, 10 and 50 μM of damnacanthal in the presence of serum. Plates were incubated for 2 weeks at 37°C. Cell colonies were fixed with 0.4 ml of 100% methanol for 5 min and stained with 0.4 ml of Giemsa solution (Acros organics, Fairlawn, NJ) for 5 min. The colonies were counted by using Colony Version 1.1 program (Fuji Photo Film Co. Stamford, CT).
2.9. Soft agar cloning assay
Soft agar assays were performed to compare the clonogenic potential of HCT-116 cells in semisolid medium. HCT-116 cells at the concentration of 3000 cells per well were resuspended in 2 ml of 0.4% agar in McCoy’s 5A medium in the presence of DMSO, 1, 10 or 50 μM of damnacanthal and plated on top of 2 ml of 0.8% agar in six-well plates. Plates were incubated for 2 to 3 weeks at 37°C. Cell colonies were visualized by staining with 0.5 ml of p-iodonitrotetrazolium violet (Sigma, Germany). The colonies were counted by using Colony Version 1.1 program (Fuji Photo Film Co.).
2.10. Cell migration assay
HCT-116 cells were plated in 12-well plates at a density of approximately 2.5×105 cells/well and grown to confluency. Cell monolayer was scratched with a sterile pipette tip, and the cells were treated with DMSO 1, 10, and 50 μM of damnacanthal. Wound size was monitored at various time points by observation under microscope, and the degree of cell migration was quantified by the ratio of gap distance at 48 h to that at 0 h.
2.11. Western blot analysis
Cells were grown to 60-80% confluence in 6-cm plates followed by 24 h treatment of various concentrations of damnacanthal or DMSO as vehicle control in the absence of serum. Total cell lysates were isolated using RIPA buffer (1xPBS, 1%NP-40, 0.5% sodium deoxycholate, 0.1% SDS) supplemented with protease inhibitors (1 mM phenylmethylsulfonyl fluoride, 5 μg/ml aprotinin and 5 μg/ml leupeptin) and phosphatase inhibitors (1 mM Na3VO4 and 1 mM NaF). The protein concentration was determined by the BCA protein assay (Pierce, Rockford, IL) using bovine serum albumin as the standard. Thirty or fifty micrograms of protein were separated on sodium dodecyl sulfate-polyacrylamide gel and transferred for 90 min onto a nitrocellulose membrane (Pall Life Sciences, Pensacola, FL). The blots were blocked for 1 h with 5% skim milk in Tris buffered saline (TBS) containing 0.05% Tween-20 (TBS-T), and probed with a specific primary antiserum in TBS/Tween 0.05% (TBS-T) and 5% non-fat dry milk at 4°C overnight. After three washes with TBS-T, the blots were incubated with horseradish peroxidase-conjugated secondary antibody for 1 h and washed several times. Proteins were detected by the enhanced chemiluminescence system (Thermo scientific, Rockford, IL).
2.12. Plasmid transient transfections
Transient transfections were performed using PolyJet (Signagen Laboratories, Ijamsville, MD) according to the manufacturer’s protocol. Briefly, HCT-116 were plated in 12-well plates at the concentration of 2×105 cells per well. The next day, plasmid mixtures containing 0.75 μg of the reporter gene and 0.075 μg of pRL-null vector were co-transfected for 24 h. For the co-transfection experiment, 0.375 μg of NAG-1 promoter and 0.375 μg of expression vectors were co-transfected with 0.075 μg of pRL-null vector. The transfected cells were then incubated with DMSO or 10 μM of damnacanthal for 24 h. For the ERK kinase inhibitor experiment, 0.75 μg of NAG-1 promoter and 0.075 μg of pRL-null vector were co-transfected for 24 h, and the transfected cells were pretreated with 1 or 5 μM of U0126 (ERK inhibitor) for 30 min. The pretreated cells were then incubated with DMSO or 20 μM of damnacanthal for 24 h. Cells were harvested in 1× luciferase lysis buffer (Promega), and luciferase activity was measured and normalized to the pRL-null luciferase activity using a dual luciferase assay kit (Promega).
2.13. Chromatin immunoprecipitation assay
The chromatin immunoprecipitation assay was performed as described previously .
2.14. RNA interference
Interference for C/EBPβ was performed as described previously . Briefly, 2.5 μg of control shRNA or C/EBPβ shRNA was transfected using PolyJet (Signagen Laboratories) for 48 h. The transfected cells were treated with either DMSO or 20 μM damnacanthal in serum-free media for 24 h, and the cells were then harvested using RIPA buffer.
2.15. Statistical analysis
Statistical analysis was performed with the Student’s t test, with statistical significance set at * P<0.05; ** P<0.01; *** P<0.001.
3. Results
3.1. Damnacanthal inhibits cell proliferation and induces caspase activity in colorectal cancer cell
We first investigated the anti-proliferative effects of damnacanthal (Dam) on human colorectal carcinoma, HCT-116 and SW480 cells. Cells were treated with 100 nM, 1, 10, 100 μM Dam for 1, 2, and 4 days. Treating HCT-116 cells with Dam resulted in a significant reduction of cell proliferation at 1 μM (P < 0.05), 10 μM (P < 0.01), and 100 μM (P < 0.01) after 4 days’ treatment, compared to DMSO-treated cells. SW480 cells also showed a significant reduction of cell proliferation at 10 μM (P < 0.05), and 100 μM (P < 0.001) after 4 days’ treatment with Dam (Fig. 1A). These data suggest that Dam has a cytotoxic effect on colorectal cancer cells, both in p53 wild type (HCT-116) and p53-mutated (SW480) cells. Based on preliminary data in which we observed a strong growth inhibitory effect of Dam in human colorectal cancer cells, we selected doses of 1, 10, and 50 μM to determine the possible inhibitory effect of Dam on cell cycle progression and apoptosis. As shown in Fig. 1B, treatment of HCT-116 cells with Dam resulted in a significant enrichment in the number of cells in the S/G1 and G2/G1 phases at concentration of 50 μM, compared to DMSO-treated cells. To observe whether Dam affects apoptosis in human colorectal carcinoma cells, we treated HCT-116 cells with DMSO or 10 μM Dam, and caspase 3/7 activity was measured. As shown in Fig. 1C, 10 μM Dam treatment significantly increased caspase 3/7 activity (3.2 fold, P < 0.05), compared to that of DMSO. These results support that damnacanthal induces cell cycle arrest and caspase activity in HCT-116 cells.
Fig. 1.
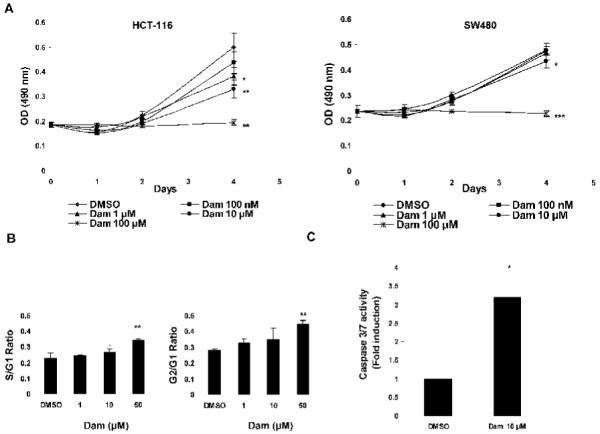
Damnacanthal inhibits cell proliferation and induces caspase activity in colorectal cancer cells. (A) HCT-116 (left panel) and SW480 (right panel) cells were treated with DMSO or various concentrations of Dam for 4 days. Cell growth was measured using the CellTiter96 Aqueous One Solution Cell Proliferation Assay. Values are expressed as mean ± SD of four replicates. *P<0.05, **P<0.01, ***P<0.001 versus DMSO-treated cells. (B) Flow cytometric analysis of DMSO or Dam-treated HCT-116 cells was analyzed. Cell were treated with DMSO or 1, 10 and 50 μM of Dam for 72 h and analyzed for cell cycle progression as described in Materials and methods. S/G1 and G2/G1 ratios are expressed as mean ± SD of three replicates. *P<0.05, **P<0.01 versus DMSO-treated cells. (C) Caspase 3/7 enzymatic activity was measured as described in Materials and methods. HCT-116 cells were treated with DMSO or 10 μM of Dam for 24 h. Y-axis represents fold induction over DMSO-treated cells. Values are expressed as mean ± SD of three replicates. *P<0.05 versus DMSO-treated cells.
3.2. Anti-tumorigenic activity of Damnacanthal
Anti-tumorigenic activity of Dam was investigated using several assays including cloning efficiency, soft-agar, and cell migration assays. As shown in Fig. 2A, Dam treatment for 2 weeks showed significant decreasing colony number in HCT-116 cells in a concentration-dependent manner, at concentrations of 10 μM (18 colonies, P < 0.05) and 50 μM (5 colonies, P < 0.01), compared to that of DMSO (43.33 colonies). We also investigated anchorage-independent cell growth by the soft agar cloning assay because the ability to form colonies in soft agar is reflective of tumorigenicity. Dam-treated cells showed a dramatic inhibition of clonogenic capacity at 10 μM (286.33 colonies, P < 0.01) and 50 μM (105.67 colonies, P < 0.001), compared to DMSO treatment (576 colonies) (Fig. 2B). Furthermore, cell migration was investigated. HCT-116 cells were plated in a 12-well plate, grown to confluency, and were scratched and treated with DMSO, 1, 10 and 50 μM of Dam as described in Materials and methods. The migration ratio was documented by photography at 0 and 48 h. As shown in Fig. 2C, Dam-treated cells significantly inhibited the migration of HCT-116 cells at 1 μM (0.41, P < 0.01), 10 μM (0.46, P < 0.001) and 50 μM (0.78, P < 0.001), compared to DMSO treatment (0.26). These results indicate that damnacanthal exhibits anti-tumorigenic activity as assessed by several in vitro assays.
Fig. 2.
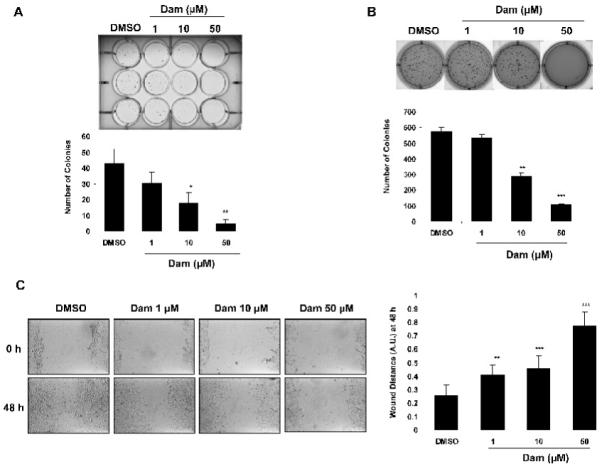
Anti-tumorigenic activity of Damnacanthal. (A) Clonogenic assay. HCT-116 cells were treated with DMSO or 1, 10 and 50 μM of Dam for 2 weeks, and cell colonies were stained and counted. The data represent mean ± SD of three replicates. *P<0.05, **P<0.01 versus DMSO-treated cells. (B) Soft agar cloning assay. HCT-116 cells were grown in 0.4% soft agar in the presence of DMSO, 1, 10 or 50 μM of Dam for 2 weeks, and then cell colonies were stained and counted. Values are expressed as mean ± SD of three replicates. **P<0.01, ***P<0.001 versus DMSO-treated cells. (C) Cell migration assay. HCT-116 cells were plated, scratched, and treated with DMSO, 1, 10 and 50 μM of Dam. Wound size was monitored at 0 and 48 h by observation under an optical microscope (left panel) at 100X magnification. The degree of cell migration was quantified by the ratio of gap distance at 48 h to that at 0 h (right panel). Values are expressed as mean ± SD of three replicates. **P<0.01, ***P<0.001 versus DMSO-treated cells.
3.3. Damnacanthal induces NAG-1 expression in HCT-116 cells
To elucidate the molecular mechanism by which Dam affects anti-tumorigenesis in colorectal cancer cells, HCT-116 and SW480 cells were treated with 0.1, 1, 10 μM of Dam for 24 h. As shown in Fig. 3A and B, NAG-1 was induced in HCT-116 cells in a dose- and time-dependent manner. Cyclin D1 expression was reduced at 10 μM of Dam, whereas p21 and p53 did not alter their expression in the presence of Dam. PARP cleavage was seen at 10 μM Dam treatment only in HCT-116 cells, where NAG-1 is induced. In contrast, NAG-1 was not increased in SW480 cells nor was PARP cleavage increased. Dam treatment also increased NAG-1 expression in a time-dependent manner, wherein we were beginning to see NAG-1 induction at 2 h, and saw it highly increased at 24 h (Fig. 2B). NAG-1 induction was not only in HCT-116 cells, but also in LoVo (p53 wild type) cells (Fig. 3C).
Fig. 3.
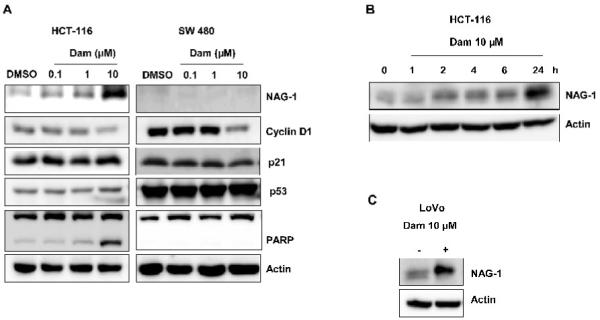
Damnacanthal induces NAG-1 expression in HCT-116 cells. (A) HCT-116 (left panel) and SW480 (right panel) cells were treated with DMSO, 0.1, 1 and 10 μM of Dam for 24 h, and Western blot was performed for NAG-1, cyclin D1, p21, p53, PARP and actin. (B) HCT-116 cells were treated with 10 μM of Dam. At the indicated times, the cell lysates were harvested to perform Western blot analysis for NAG-1 and actin. (C) LoVo cells were treated with DMSO or 10 μM of Dam for 48 h, and Western blot was performed for NAG-1 and actin.
3.4. C/EBPβ mediates damnacanthal-induced NAG-1 expression
Because phytochemicals increase several transcriptional factors that play an important role in NAG-1 expression , we next investigated whether damnacanthal activated NAG-1 at the transcriptional level. We first performed a deletion assay using the four serial deletion constructs of human NAG-1 promoter (pNAG-1-3500/+41, pNAG-1-1086/+41, pNAG-1-474/+41 and pNAG-1-133/+41) to observe whether Dam affects transcriptional regulation of the NAG-1 gene. NAG-1 promoters were transfected into HCT-116 cells and treated with 10 μM of Dam for 24 h. As shown in Fig. 4A, Dam treatment resulted in the stimulation of NAG-1 promoter activity in all the constructs tested, implying that the −133 and +41 region of the promoter is responsible for damnacanthal’s effect.
Fig. 4.
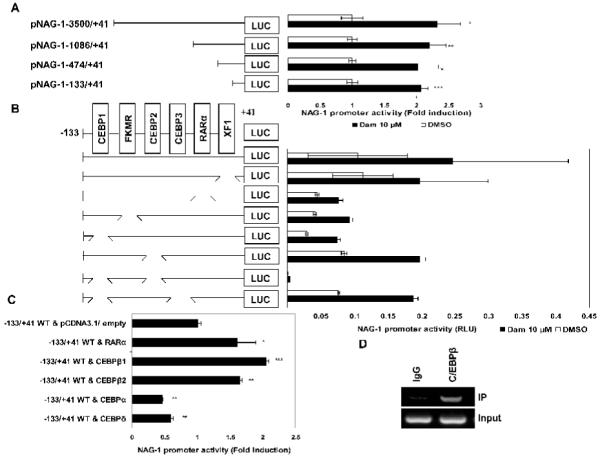
Identification of the promoter region responsible for damnacanthal-induced NAG-1 transactivation. (A) Each indicated construct of the NAG-1 promoter and pRL-null vector were transiently transfected into HCT-116 cells and then treated with DMSO or 10 μM of Dam for 24 h. The promoter activity was measured as a ratio of firefly luciferase signal/renilla luciferase signal. Fold induction refers to ratio of luciferase (LUC) activity of Dam-treated cells compared to DMSO-treated cells of each construct. The results are the mean ± SD of three replicates. *P<0.05, **P<0.01, ***P<0.001 versus DMSO-treated cells. (B) HCT-116 cells were transfected with each internal deletion construct of the NAG-1 promoter (pNAG-1-133/+41) and then treated with 10 μM of Dam for 24 h. The x-axis shows relative luciferase unit of each construct. RLU, relative luciferase unit. (C) The NAG-1 promoter (pNAG-1-133/+41) was co-transfected with indicated expression vector as described in Materials and methods and then treated with 10 μM of Dam for 24 h. The x-axis shows fold induction over empty vector. *P<0.05, **P<0.01, ***P<0.001 versus control vector-treated cells. (D) Chromatin immunoprecipitation assay was performed using a DNA-protein complex treated with 20 μM of Dam for 24 h as described in Materials and methods. The sequence of the NAG-1 promoter region (−131/+137) was amplified by PCR primer pairs as described previously. The input represents PCR products obtained from 1% aliquots of chromatin pellets escaping immunoprecipitation.
It has been reported that there are several cis-acting elements in this region, including C/EBPs, RARα, XF1 and FKMR . To confirm the responsible site for the transactivation of NAG-1 gene by Dam, we utilized internal deletion clones for C/EBPs, RARα, XF1 and FKMR. HCT-116 cells were transfected with wild-type or internal-deletion constructs, and treated with 10 μM of Dam for 24 h. As shown in Fig. 4B, none of the internal deletion clones, including C/EBP, FKMR, RARα and XF1, showed significant differences in Dam-induced promoter activity, compared to wild type promoter. Interestingly, transfection with double deletion clone lacking two potential C/EBP binding sites (CEBP1 and CEBP2) abolished the basal and Dam-induced promoter activity. These data suggest that these two regions on the C/EBP binding site are important for NAG-1 promoter activation by damnacanthal treatment.
To find the trans-acting elements responsible for Dam-induced NAG-1 transactivation, we tested whether overexpression of different types of C/EBP or RARα affect NAG-1 transactivation. We cotransfected expression vectors for C/EBPβ1, C/EBPβ2, C/EBPα1, C/EBPδ and RARα with the NAG-1 promoter constructs (pNAG-1 −133/+41), and promoter activity was determined. As shown in Fig 4C, C/EBPβ1, C/EBPβ2 and RARα expression showed a significant increase of damnacanthal-induced promoter activity, whereas C/EBPα and C/EBPδ expression slightly decreased Dam-induced promoter activity (Fig. 4C). To confirm the binding of C/EBPβ to this site, the chromatin immunoprecipitation assay was performed using C/EBPβ antibody. As shown in Fig. 4D, C/EBPβ binds to the −133/+41 region of the NAG-1 promoter, whereas rabbit control IgG did not pull down any protein to this region. Taken together, these results indicate that two C/EBPβ binding sites (CEBP1 and CEBP2) are pivotal cis-acting element to confer damnacanthal-induced NAG-1 expression.
3.5. Effect of kinase inhibitors on NAG-1 expression
Transcriptional activity of C/EBPβ is regulated by its translation or post-translational modulation . To examine whether Dam increases C/EBPβ expression, HCT-116 cells were treated with DMSO, 0.1, 1, 10, or 20 μM of Dam for 24 h and Western blot was performed using C/EBPβ specific antibody. As shown in Fig. 5A, C/EBPβ protein increased in HCT-116 cells in a dose-dependent manner. To explore the upstream molecular mechanism involved in Dam-induced NAG-1 and C/EBPβ expression, we examined several signaling pathways that are affected by damnacanthal. We screened several kinase-specific inhibitors at a concentration that does not deviate from their selectivity. HCT-116 cells were pretreated with kinase inhibitors for 30 min, prior to incubation with 20 μM of Dam. As shown in Fig. 5B, the Dam-induced NAG-1 expression was strongly suppressed in the presence of U0126 (ERK inhibitor) and RO318220 (PKC inhibitor). In addition, we found that C/EBPβ was also suppressed by U0126 and RO318220. These results suggest roles of the ERK and PKC protein kinase pathways in Dam-induced NAG-1 and C/EBPβ expression. The construct pNAG-1-133/+41 was further transfected into HCT-116 cells and treated with 1 or 5 μM of U0126 (ERK inhibitor) for 30 min prior to incubation with 20 μM of Dam. As shown in Fig. 5C, NAG-1 promoter activity is diminished in the presence of U0126 compound in a dose-dependent manner. This result demonstrates that damnacanthal affects ERK activity, followed by the induction of NAG-1 expression at the transcriptional level.
Fig. 5.
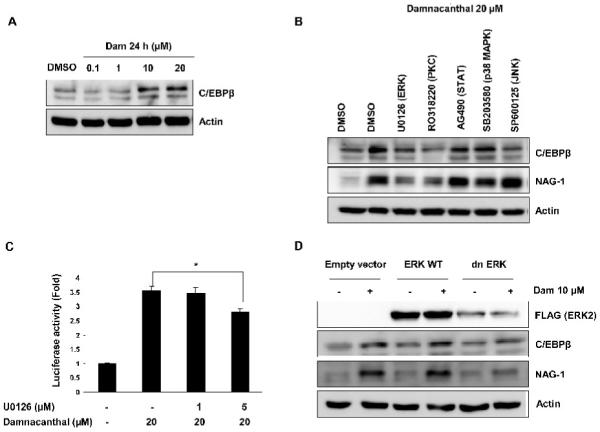
ERK and PKC mediate C/EBPβ and damnacanthal-induced NAG-1 expression. (A) HCT-116 cells were treated with DMSO, 0.1, 1, 10 and 20 μM of Dam for 24 h, and Western blot was performed for C/EBPβ and actin as described in Materials and methods. (B) HCT-116 cells were pretreated with different kinase inhibitors (U0126, 5 μM; RO318220, 2.5 μM; AG490, 50 μM; SB203580, 15 μM; SP 600125, 30 μM) for 30 min and treated with 20 μM of Dam for 24 h. Western blot was performed for C/EBPβ, NAG-1 and actin. (C) The NAG-1 promoter (pNAG-1-133/+41) and pRL-null vector were transfected for 24 h, as described in Materials and methods, pretreated with U0126 (1 or 5 μM) and then treated with DMSO or 20 μM of Dam for 24 h. The y-axis shows fold induction over DMSO-treated cells. *P<0.05 versus Dam 20 μM-treated cells. (D) HCT-116 cells were transfected with pcDNA 3.1 empty vector, ERK2 wild type or a dominant negative ERK2 as described in Materials and methods, and treated with DMSO or 10 μM of Dam for 24 h. Western blot was performed for FLAG, C/EBPβ , NAG-1 and actin.
To confirm the possible regulatory effect of ERK on Dam-induced NAG-1 expression, HCT-116 cells were transfected with empty vector, ERK2 wild type, or dominant-negative ERK2, and treated with 10 μM of Dam for 24 h. As shown in Fig. 5D, transfection with dominant-negative ERK suppressed Dam-induced NAG-1 expression, compared to control empty vector and ERK2 wild type transfected cells with Dam treatment. Similar results were obtained in C/EBPβ expression. These results indicated that damnacanthal affects ERK2 activity, followed by the induction of C/EBPβ and NAG-1 expression in HCT-116 cells.
3.6. C/EBPβ and NAG-1 are involved in damnacanthal-induced apoptosis
To explore whether C/EBPβ expression contributes to Dam-induced apoptosis, we performed knock-down of the endogeneous C/EBPβ gene by RNA interference and analyzed apoptosis-related caspase 3/7 activity. HCT-116 cells were transfected with shRNA for C/EBPβ. As shown in Fig. 6A, transfection with C/EBPβ shRNA clearly suppressed Dam-induced C/EBPβ. Moreover, we also explored NAG-1 expression in C/EBPβ shRNA- transfected cells. In the presence of 20 μM of Dam, NAG-1 expression was decreased in C/EBPβ shRNA-transfected cells, compared to control shRNA transfected cells with Dam treatment. Treatment of C/EBPβ shRNA-transfected cells with Dam decreased caspase 3/7 activity (1.5 fold, P < 0.001), compared to control shRNA-transfected cells with Dam treatment (3.3 fold) (Fig. 6B), indicating an important role of C/EBPβ in damnacanthal-induced caspase activity.
Fig. 6.
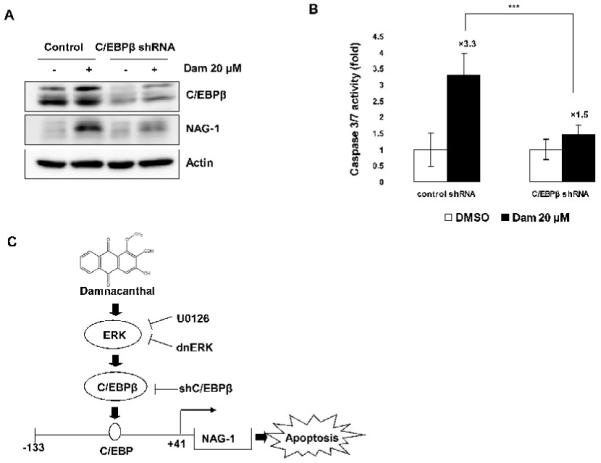
Knockdown of C/EBPβ alters damnacanthal-induced NAG-1 expression and apoptosis. (A) HCT-116 cells were transfected with control or C/EBPβ shRNA for 48 h and then treated with DMSO or 20 μM of Dam for 24 h. Western analysis was performed for C/EBPβ, NAG-1, and actin. (B) HCT-116 cells were transfected with control or C/EBPβ shRNA for 48 h and then treated with DMSO or 20 μM of Dam for 24 h. Caspase 3/7 enzymatic activity was measured as described in Materials and methods. Values are expressed as mean ± SD of three replicates. ***P<0.001 versus control shRNA-transfected Dam treatment cells. (C) Proposed mechanism by which Dam induces NAG-1 transcription in human colorectal cancer cells. Dam activates C/EBPβ through the ERK kinase pathway. Enhanced expression of C/EBPβ increases the binding affinity of C/EBPβ onto the NAG-1 promoter and activates transcription of the NAG-1 gene. Up-regulation of the NAG-1 gene results in an increase of apoptosis in colorectal cancer cells.
4. Discussion
Cancer is second only to heart disease as the leading cause of death in the United States, and colorectal cancer is one of the most prevalent causes of cancer-related mortality in the Western world . Further development of therapeutic and preventive means of controlling this disease is clearly needed. Epidemiological studies have suggested that phytochemicals found in dietary plant products, including fruits, vegetables, beverages, herbs, and spices, play an important role in carcinogenesis, and a number of these compounds have been found to inhibit tumorigenesis in experimental animals and/or exhibit potent biological properties . One such phytochemical is damnacanthal, an anthraquinone and a naturally occurring component of noni root. In this study, the cell proliferation assay showed that damnacanthal treatment results in the inhibition of cell growth, compared to vehicle-treated cells in two different colorectal cancer cells (Fig. 1). Cell growth retardation properties of damnacanthal are more evident by the soft agar cloning, clonogenic, and cell migration assays (Fig. 2). All these results show that damnacanthal increased anti-tumorigenic activity by increased expression of C/EBPβ, followed by NAG-1 expression.
NAG-1 is a pro-apoptotic protein that exhibits anti-tumorigenic activity in culture cells as well as in animal models . NAG-1 transgenic mice (overexpression) formed fewer tumors in carcinogenic- and genetic-induced animal models . Interestingly, NAG-1 is induced by many phytochemicals, having anti-tumorigenic and anti-inflammatory activities . Therefore, NAG-1 may be an important protein that controls compound-induced anti-tumorigenesis as well as anti-inflammation. Indeed, not only phytochemicals but also drugs that show an anti-inflammation increase NAG-1 expression . Thus, NAG-1 may be a molecular target protein for these compounds that control cell growth and apoptosis.
Many transcription factors have been identified that control NAG-1 transcriptional regulation. These include EGR-1, p53, ATF3, and PPARγ . We observed transcriptional up-regulation of the NAG-1 by Dam and identified three putative C/EBP binding sites (CEBP1, CEBP2, and CEBP3) in the NAG-1 promoter (between −133 and +41). No single deletion of each C/EBP binding site changed Dam-mediated NAG-1 transactivation. Interestingly, double deletion of C/EBP binding sites (CEBP1 and CEBP2) completely inhibited basal as well as Dam-induced promoter activity, indicating the simultaneous significant role of these two sites. In addition, co-transfection with expression vectors and ChIP assay indicates that C/EBPβ mediates NAG-1 transactivation by direct binding to the NAG-1 promoter. Recently, we have also shown that C/EBPβ also plays an important role in capsaicin-induced NAG-1 expression with ATF3, C/EBPβ, and GSK-3β complexes . These complexes are facilitated by capsaicin via PKCδ activation. Although we have shown that Dam affects NAG-1 expression through a C/EBPβ-dependent mechanism, the mechanism by which Dam upregulates NAG-1 is different from that of capsaicin. First, Dam increases C/EBPβ expression, while capsaicin did not increase C/EBPβ expression. Rather capsaicin induces C/EBPβ activity by phosphorylation . Since the activity of C/EBPβ is also regulated by expression, overexpression of C/EBP by Dam may modulate cell growth and apoptosis by activating their target genes such as NAG-1. Our data support that NAG-1 is a Dam-regulating C/EBPβ target gene involved in apoptosis of human colorectal cancer cells. Secondly, Dam utilizes the ERK pathway to increase NAG-1 as well as C/EBPβ expression, while the effect of MEK inhibitor on capsaicin-induced NAG-1 expression is marginal . Indeed, it has been shown that Dam increases ERK activity by phosphorylation and ERK plays an important role in C/EBPβ expression in colorectal cancer cells . Thus, we propose that damnacanthal affects ERK activity, followed by C/EBPβ expression. These sequential pathways may ultimately affect NAG-1 induction (Fig. 6C). ERK also mediates NAG-1 expression by different anti-cancer compounds . Hiramatsu et al. screened tropical plant extracts for substances that induce normal morphology in K-rasts-NRK cells . As a result, they isolated damnacanthal from the chloroform extract of the root of noni and found that damnacanthal was an inhibitor of Ras function. Taken together with our data, it is suggested that damnacanthal alters several kinase pathways, leading to anti-tumorigenesis in cancer cells.
It has been known that various types of transcription factors, including those in the GATA family and ATF/CREB family, directly binding to the C/EBP site, regulating target genes . Although double deletion of a C/EBP binding sites or knockdown of C/EBPβ completely suppressed NAG-1 transactivation and protein expression (Fig. 4B, Fig. 6A), we do not exclude the possibility that other proteins modified by Dam may involve NAG-1 regulation by changing binding affinity to the C/EBP binding site. With respect to the biological activity of C/EBPβ in colorectal cancer, it has been known that C/EBPβ is a positive regulator of COX-2 . However, recent study showed a direct association between strong suppression of colon tumor growth and C/EBPβ overexpression in nude mice bearing CW-2 human colon cancer tumors . Thus, detailed studies are required for pro- or anti-tumorigenic activity of C/EBPβ in colorectal carcinoma.
In conclusion, damnacanthal, a bioactive compound from noni roots, activates ERK pathway and enhanced expression of C/EBPβ. Overexpression of C/EBPβ directly activates transcription and expression of NAG-1 and subsequently increases apoptosis in human colorectal cancer. These results will provide potential benefits of damnacanthal for further preclinical or clinical practice and damnacanthal may be a useful cancer prevention/therapeutic agent in human colorectal carcinoma.
Acknowledgements
We thank Misty Bailey for her critical reading of this manuscript. We also thank Nichelle Whitlock, Jae Hoon Bahn, and Jason Liggett for their technical assistance.
Grant: This work was supported by grants from National Institutes of Health (R01CA108975), and the University of Tennessee Center of Excellence in Livestock Diseases and Human Health to SJB. Financial support for TN by the Royal Golden Jubilee PhD Program (PHD/0159/2546), Thailand.
Abbreviations
- DMSO
dimethyl sulfoxide
- C/EBPβ
CCAAT/enhancer binding protein β
- ChIP
chromatin immunoprecipitation
- NAG-1
non-steroidal anti-inflammatory drug (NSAID)-activated gene-1
- RARα
retinoic acid receptor α
- ERK
extracellular-signal-regulated kinase
- dnERK
dominant-negative ERK
Footnotes
Conflict of Interest
No conflict of interest exists in the submission of this manuscript.
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- [1].Jemal A, Siegel R, Xu J, Ward E. Cancer Statistics. CA Cancer J Clin. 2010 doi: 10.3322/caac.20073. [DOI] [PubMed] [Google Scholar]
- [2].Rajamanickam S, Agarwal R. Natural products and colon cancer: current status and future prospects. Drug Dev Res. 2008;69:460–71. doi: 10.1002/ddr.20276. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [3].Wang M, et al. Morinda citrifolia (Noni):A literature review and recent advances in Noni research. Acta Pharmacol Sin. 2002;23:1127–41. [PubMed] [Google Scholar]
- [4].Lemmens R, Bunyapraphatsara N. Plant resources of South-East Asia. Backhuys Publishers; Leiden: 2003. [Google Scholar]
- [5].Hornick C, Myers A, Sadowska-Krowicka H, Anthony C, Woltering E. Inhibition of angiogenic initiation and disruption of newly established human vascular networks by juice from Morinda citrifolia (noni) Angiogenesis. 2003;6:143–9. doi: 10.1023/B:AGEN.0000011800.04507.4b. [DOI] [PubMed] [Google Scholar]
- [6].Wang MY, Su C. Cancer preventive effect of Morinda citrifolia (Noni) Ann N Y Acad Sci. 2001;952:161–8. doi: 10.1111/j.1749-6632.2001.tb02737.x. [DOI] [PubMed] [Google Scholar]
- [7].Palu AK, Kim AH, West BJ, Deng S, Jensen J, White L. The effects of Morinda citrifolia L. (noni) on the immune system: Its molecular mechanisms of action. Journal of Ethnopharmacology. 2008;115:502–6. doi: 10.1016/j.jep.2007.10.023. [DOI] [PubMed] [Google Scholar]
- [8].Li J, Stickel SL, Bouton-Verville H, Burgin KE, Yu X, Wong DK, et al. Fermented Noni exudate (fNE): a mediator between immune system and antitumor activity. Oncol Rep. 2008;20:1505–9. [PubMed] [Google Scholar]
- [9].Furusawa E, Hirazumi A, Story S, Jensen J. Antitumour potential of a polysaccharide-rich substance from the fruit juice of Morinda citrifolia (Noni) on sarcoma 180 ascites tumour in mice. Phytother Res. 2003;17:1158–64. doi: 10.1002/ptr.1307. [DOI] [PubMed] [Google Scholar]
- [10].Hirazumi A, Furusawa E. An immunomodulatory polysaccharide-rich substance from the fruit juice of Morinda citrifolia (noni) with antitumour activity. Phytother Res. 1999;13:380–7. doi: 10.1002/(sici)1099-1573(199908/09)13:5<380::aid-ptr463>3.0.co;2-m. [DOI] [PubMed] [Google Scholar]
- [11].Faltynek CRSJ, Mauvais P, Miller D, Wang S, et al. Damnacanthal is a highly potent, selective inhibitor of p56lck tyrosine kinase activity. Biochemistry. 1995;34:12404–10. doi: 10.1021/bi00038a038. [DOI] [PubMed] [Google Scholar]
- [12].Kanokmedhakul K, Kanokmedhakul S, Phatchana R. Biological activity of anthraquinones and triterpenoids from Prismatomeris fragrans. Journal of Ethnopharmacology. 2005;100:284–8. doi: 10.1016/j.jep.2005.03.018. [DOI] [PubMed] [Google Scholar]
- [13].Baek SJ, Kim KS, Nixon JB, Wilson LC, Eling TE. Cyclooxygenase inhibitors regulate the expression of a TGF-beta superfamily member that has proapoptotic and antitumorigenic activities. Mol Pharmacol. 2001;59:901–8. [PubMed] [Google Scholar]
- [14].Baek SJ, Eling TE. Changes in gene expression contribute to cancer prevention by COX inhibitors. Prog Lipid Res. 2006;45:1–16. doi: 10.1016/j.plipres.2005.10.001. [DOI] [PubMed] [Google Scholar]
- [15].Baek SJ, Okazaki R, Lee SH, Martinez J, Kim JS, Yamaguchi K, et al. Nonsteroidal anti-inflammatory drug-activated gene-1 over expression in transgenic mice suppresses intestinal neoplasia. Gastroenterology. 2006;131:1553–60. doi: 10.1053/j.gastro.2006.09.015. [DOI] [PubMed] [Google Scholar]
- [16].Cekanova M, Lee SH, Donnell RL, Sukhthankar M, Eling TE, Fischer SM, et al. Nonsteroidal anti-inflammatory drug-activated gene-1 expression inhibits urethane-induced pulmonary tumorigenesis in transgenic mice. Cancer Prev Res (Phila) 2009;2:450–8. doi: 10.1158/1940-6207.CAPR-09-0057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [17].Baek SJ, Kim JS, Moore SM, Lee SH, Martinez J, Eling TE. Cyclooxygenase inhibitors induce the expression of the tumor suppressor gene EGR-1, which results in the up-regulation of NAG-1, an antitumorigenic protein. Mol Pharmacol. 2005;67:356–64. doi: 10.1124/mol.104.005108. [DOI] [PubMed] [Google Scholar]
- [18].Baek SJ, Wilson LC, Eling TE. Resveratrol enhances the expression of non-steroidal anti-inflammatory drug-activated gene (NAG-1) by increasing the expression of p53. Carcinogenesis. 2002;23:425–34. doi: 10.1093/carcin/23.3.425. [DOI] [PubMed] [Google Scholar]
- [19].Lee SH, Krisanapun C, Baek SJ. NSAID-activated gene-1 as a molecular target for capsaicin-induced apoptosis through a novel molecular mechanism involving GSK3beta, C/EBPbeta and ATF3. Carcinogenesis. 2010;31:719–28. doi: 10.1093/carcin/bgq016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [20].Lee SH, Yamaguchi K, Kim JS, Eling TE, Safe S, Park Y, et al. Conjugated linoleic acid stimulates an anti-tumorigenic protein NAG-1 in an isomer specific manner. Carcinogenesis. 2006;27:972–81. doi: 10.1093/carcin/bgi268. [DOI] [PubMed] [Google Scholar]
- [21].Hiramatsu T, Imoto M, Koyano T, Umezawa K. Induction of normal phenotypes in ras-transformed cells by damnacanthal from Morinda citrifolia. Cancer Letters. 1993;73:161–6. doi: 10.1016/0304-3835(93)90259-c. [DOI] [PubMed] [Google Scholar]
- [22].Baek SJ, Horowitz JM, Eling TE. Molecular cloning and characterization of human nonsteroidal anti-inflammatory drug-activated gene promoter. Basal transcription is mediated by Sp1 and Sp3. J Biol Chem. 2001;276:33384–92. doi: 10.1074/jbc.M101814200. [DOI] [PubMed] [Google Scholar]
- [23].Lee SH, Bahn JH, Whitlock NC, Baek SJ. Activating transcription factor 2 (ATF2) controls tolfenamic acid-induced ATF3 expression via MAP kinase pathways. Oncogene. 29:5182–92. doi: 10.1038/onc.2010.251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [24].Auyeung KK, Ko JK. Coptis chinensis inhibits hepatocellular carcinoma cell growth through nonsteroidal anti-inflammatory drug-activated gene activation. Int J Mol Med. 2009;24:571–7. doi: 10.3892/ijmm_00000267. [DOI] [PubMed] [Google Scholar]
- [25].Jutooru I, Chadalapaka G, Chintharlapalli S, Papineni S, Safe S. Induction of apoptosis and nonsteroidal anti-inflammatory drug-activated gene 1 in pancreatic cancer cells by a glycyrrhetinic acid derivative. Mol Carcinog. 2009;48:692–702. doi: 10.1002/mc.20518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [26].Wu Z, Xie Y, Bucher NL, Farmer SR. Conditional ectopic expression of C/EBP beta in NIH-3T3 cells induces PPAR gamma and stimulates adipogenesis. Genes Dev. 1995;9:2350–63. doi: 10.1101/gad.9.19.2350. [DOI] [PubMed] [Google Scholar]
- [27].Park BH, Qiang L, Farmer SR. Phosphorylation of C/EBPbeta at a consensus extracellular signal-regulated kinase/glycogen synthase kinase 3 site is required for the induction of adiponectin gene expression during the differentiation of mouse fibroblasts into adipocytes. Mol Cell Biol. 2004;24:8671–80. doi: 10.1128/MCB.24.19.8671-8680.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [28].Potter JD, Slattery ML, Bostick RM, Gapstur SM. Colon cancer: a review of the epidemiology. Epidemiol Rev. 1993;15:499–545. doi: 10.1093/oxfordjournals.epirev.a036132. [DOI] [PubMed] [Google Scholar]
- [29].Wenzel U, Kuntz S, Brendel MD, Daniel H. Dietary flavone is a potent apoptosis inducer in human colon carcinoma cells. Cancer Res. 2000;60:3823–31. [PubMed] [Google Scholar]
- [30].Yamaguchi K, Lee SH, Eling TE, Baek SJ. A novel peroxisome proliferator-activated receptor gamma ligand, MCC-555, induces apoptosis via posttranscriptional regulation of NAG-1 in colorectal cancer cells. Mol Cancer Ther. 2006;5:1352–61. doi: 10.1158/1535-7163.MCT-05-0528. [DOI] [PubMed] [Google Scholar]
- [31].Hiwasa T, Arase Y, Chen Z, Kita K, Umezawa K, Ito H, et al. Stimulation of ultraviolet-induced apoptosis of human fibroblast UVr-1 cells by tyrosine kinase inhibitors. FEBS Letters. 1999;444:173–6. doi: 10.1016/s0014-5793(99)00057-5. [DOI] [PubMed] [Google Scholar]
- [32].Salmenpera P, Hamalainen S, Hukkanen M, Kankuri E. Interferon-gamma induces C/EBP beta expression and activity through MEK/ERK and p38 in T84 colon epithelial cells. Am J Physiol Cell Physiol. 2003;284:C1133–9. doi: 10.1152/ajpcell.00293.2002. [DOI] [PubMed] [Google Scholar]
- [33].Martinez JM, Sali T, Okazaki R, Anna C, Hollingshead M, Hose C, et al. Drug-induced expression of nonsteroidal anti-inflammatory drug-activated gene/macrophage inhibitory cytokine-1/prostate-derived factor, a putative tumor suppressor, inhibits tumor growth. J Pharmacol Exp Ther. 2006;318:899–906. doi: 10.1124/jpet.105.100081. [DOI] [PubMed] [Google Scholar]
- [34].Chen YL, Lin PC, Chen SP, Lin CC, Tsai NM, Cheng YL, et al. Activation of nonsteroidal anti-inflammatory drug-activated gene-1 via extracellular signal-regulated kinase 1/2 mitogen-activated protein kinase revealed a isochaihulactone-triggered apoptotic pathway in human lung cancer A549 cells. J Pharmacol Exp Ther. 2007;323:746–56. doi: 10.1124/jpet.107.126193. [DOI] [PubMed] [Google Scholar]
- [35].Blobel GA. CREB-binding protein and p300: molecular integrators of hematopoietic transcription. Blood. 2000;95:745–55. [PubMed] [Google Scholar]
- [36].Foti D, Iuliano R, Chiefari E, Brunetti A. A nucleoprotein complex containing Sp1, C/EBP beta, and HMGI-Y controls human insulin receptor gene transcription. Mol Cell Biol. 2003;23:2720–32. doi: 10.1128/MCB.23.8.2720-2732.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [37].Weber M, Sydlik C, Quirling M, Nothdurfter C, Zwergal A, Heiss P, et al. Transcriptional inhibition of interleukin-8 expression in tumor necrosis factor-tolerant cells: evidence for involvement of C/EBP beta. J Biol Chem. 2003;278:23586–93. doi: 10.1074/jbc.M211646200. [DOI] [PubMed] [Google Scholar]
- [38].Boruk M, Savory JG, Hache RJ. AF-2-dependent potentiation of CCAAT enhancer binding protein beta-mediated transcriptional activation by glucocorticoid receptor. Mol Endocrinol. 1998;12:1749–63. doi: 10.1210/mend.12.11.0191. [DOI] [PubMed] [Google Scholar]
- [39].van Hattem WA, Brosens LA, Marks SY, Milne AN, van Eeden S, Iacobuzio-Donahue CA, et al. Increased cyclooxygenase-2 expression in juvenile polyposis syndrome. Clin Gastroenterol Hepatol. 2009;7:93–7. doi: 10.1016/j.cgh.2008.07.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [40].Eisinger AL, Nadauld LD, Shelton DN, Peterson PW, Phelps RA, Chidester S, et al. The adenomatous polyposis coli tumor suppressor gene regulates expression of cyclooxygenase-2 by a mechanism that involves retinoic acid. J Biol Chem. 2006;281:20474–82. doi: 10.1074/jbc.M602859200. [DOI] [PubMed] [Google Scholar]
- [41].Sun L, Fu BB, Liu DG. Systemic delivery of full-length C/EBP beta/liposome complex suppresses growth of human colon cancer in nude mice. Cell Res. 2005;15:770–6. doi: 10.1038/sj.cr.7290346. [DOI] [PubMed] [Google Scholar]