

Abstract
During normal lung development and in lung diseases structural cells in the lungs adapt to permit changes in lung function. Fibroblasts, myofibroblasts, smooth muscle, epithelial cells, and various progenitor cells can all undergo phenotypic modulation. In the pulmonary vasculature occlusive vascular lesions that occur in severe pulmonary arterial hypertension are multifocal, polyclonal lesions containing cells presumed to have undergone phenotypic transition resulting in altered proliferation, cell lifespan or contractility. Dynamic changes in gene expression and protein composition that underlie processes responsible for such cellular plasticity are not fully defined. Advances in molecular biology have shown that multiple classes of ribonucleic acid (RNA) collaborate to establish the set of proteins expressed in a cell. Both coding Messenger Ribonucleic acid (mRNA) and small noncoding RNAs (miRNA) act via multiple parallel signaling pathways to regulate transcription, mRNA processing, mRNA stability, translation and possibly protein lifespan. Rapid progress has been made in describing dynamic features of miRNA expression and miRNA function in some vascular tissues. However posttranscriptional gene silencing by microRNA-mediated mRNA degradation and translational blockade is not as well defined in the pulmonary vasculature. Recent progress in defining miRNAs that modulate vascular cell phenotypes is reviewed to illustrate both functional and therapeutic significance of small noncoding RNAs in pulmonary arterial hypertension.
Keywords: hypertension, Krueppel-like factor 4, myocardin, smooth muscle, translation, vascular remodeling, vascular injury
INTRODUCTION
During development of the airways and the pulmonary circulation, multiple cell types adapt to changing chemical and physical signals to establish appropriate lung structure and function. Progenitor cells of various vascular cell types proliferate, migrate and reorient themselves to form nascent vessels and airways that eventually mature to the adult phenotype. During disease development there are also adaptive changes in function and structure, some of which are maladaptive and some of which oppose the disease process. Functional adaptations are manifest in the pulmonary circulation in pulmonary hypertension syndromes as hypercontractility.
Structural adaptations are manifest as vascular pruning, medial thickening, leukocyte invasion and development of a variety of occlusive lesions. The initial triggers or stimuli for these remodeling events are under intense investigation, as are the pathways and proteins altered during pulmonary diseases. Structural studies suggest multiple cell types including lung fibroblasts, myofibroblasts, smooth muscle, epithelial, endothelial and progenitor cells all undergo varying degrees of phenotypic modulation during disease development. Blood vessel remodeling events include matrix remodeling, secretion of numerous cell signaling molecules, cell and tissue hypertrophy, and hyperplasia. In all hollow organs, including pulmonary blood vessels, smooth muscle cells undergo dynamic changes in gene expression and protein composition to adapt to changes in the local environment. When such changes are long-lasting, they are described as being due to “cellular plasticity.” The set of proteins expressed is determined by multiple parallel signal transduction pathways that ultimately regulate one or more events in transcription, translation, mRNA half-life and protein degradation. Transcriptional controls have been studied extensively in vascular cells, but epigenetic mechanisms contributing to smooth muscle phenotype are not as well defined. Dynamic changes in methylation of CpG sites in key promoters, histone modifications and microRNA-induced gene silencing are subjects of intense study in cardiovascular physiology. These phenomena are not nearly as well defined in smooth muscle cells as they are in cardiac muscle cancer cells and the immune system.[1–3]
The goal of this review is to summarize emerging knowledge of the microRNA (miRNA) class of small, noncoding RNAs in vascular smooth muscle cell phenotypes in normal tissue and in pulmonary diseases. We will focus on miRNAs with validated targets that are relevant to smooth muscle contractility and vascular development. The reader interested in miRNAs in endothelial cells and stem cells is referred to several excellent recent reviews of the subject and a recent review of miRNAs in pulmonary hypertension.[4–8] Our narrow approach to the topic in this review is justified by the compelling need to identify novel, druggable targets for modifying vascular remodeling. There is certainly continuing need to define sets of miRNAs that control both conserved (proliferation and cell survival) and unique (smooth muscle contractile protein expression) processes in all vascular cells and all cells in the lung. There is also an appealing opportunity to capitalize on current knowledge of miRNA-induced gene silencing in developing novel therapeutic approaches to pulmonary hypertension. To that end, we describe examples of RNAi-based therapy of animal models of cardiovascular and respiratory diseases. These studies provide an exciting proof of principle for RNAi therapy of lung diseases including pulmonary hypertension.
Smooth muscle cell phenotypes
Smooth muscle cells in vitro are highly plastic cells that are easily manipulated by altering culture conditions to favor a more contractile phenotype or a proliferative, secretory and migrating phenotype. Contractile characteristics are promoted by culturing at high density and in reduced serum concentrations in the presence of soluble factors including insulin, retinoic acid, and transforming growth factor beta 1 (TGF-β1). Some soluble factors are clearly derived from or promoted by endothelial cells, which, in coculture, promote differentiated, contractile pulmonary artery smooth muscle cells.[9] Contractile smooth muscle cells are defined as cells expressing smooth muscle-restricted contractile and cytoskeletal proteins that contract in response to physiological agonists (e.g., norepinephrine, serotonin, histamine, enthothelin-1). There are several well-defined smooth muscle-restricted contractile proteins including myosin II heavy chain, α and γ smooth-muscle actins, h-caldesmon, h1-calponin, smooth muscle tropomyosins, SM22 (transgelin) and smoothelin.[10,11] The contractile proteins are typically downregulated by conditions promoting proliferation. In culture proliferation, cell migration and secretion of mediators of inflammation can be induced by serum-containing medium with the trophic growth factors epidermal growth factor and fibroblast growth factor. The gene expression profile of proliferative and migratory vascular smooth muscle cells is not as well defined as the contractile phenotype. It is frequently used in the context of cultured cells that proliferate in serum-containing medium, express a chemotactic response to platelet derived growth factor (PDGF), and secrete a variety of proteins. Secreted proteins include type I collagen, cytokines, chemo-kines and growth factors. Growth, migration and proliferation in vitro are thought to recapitulate organogenesis during fetal and neonatal development.
There are differing views of the two “phenotypes” of vascular smooth muscle. One view holds that switching from proliferating/migrating cells to contractile cells is a stable, mutually exclusive condition—a binary phenomenon.[11] An alternate view is that the phenotype of smooth muscle cells is graded with cells in a tissue having a mosaic pattern of contractile protein gene expression.[10,12,13] In either case, gene expression programs in smooth muscle appear to be highly adaptable depending on tissue type, culture conditions and disease processes.
Current progress in epigenetic mechanisms controlling gene expression strongly suggests part of the adaptability of vascular smooth muscle and other vascular wall cells is due to dynamic changes in gene expression. One topic of great interest is the influence miRNAs might have on networks of target genes that are important in disease progression. Defining the miRNAs and their targets in pulmonary vascular smooth muscle phenotype switching should add novel therapeutic targets for anti-remodeling drugs. Progress on this topic will have high impact on translational research aimed at developing novel treatments of pulmonary hypertension.
MiRNA biogenesis and miRNA-induced silencing
MiRNA biogenesis and the mechanisms of miRNA-induced gene silencing have been well described, and the basic steps appear to be highly conserved among various cell types. The current consensus on biogenesis is illustrated in Figure 1, and the interested reader is directed to recent reviews of the topic for more detailed description of the process.[14,15] Many miRNA genes are hosted within other genes distributed throughout mammalian genomes. They are often located in introns and sometimes in exons, and some are in intergenic regions rather than within a host gene. Clusters of coexpressed, polycistronic primary transcripts are common and many miRNA genes have multiple copies in the human genome. Hypoxia-regulated miRNAs are a good example of coexpressed clusters of miRNAs relevant to pulmonary vascular diseases.[16] MiRNAs hosted by protein-coding genes are under control of Pol II promoters and familiar transcription factor families that control expression of mRNAs. However, some miRNA genes have independent promoters. A few miRNA genes are transcribed by Pol III, which transcribes tRNA, 5S rRNA and small nuclear RNA genes. Primary miRNA transcripts are capped and polyadenylated then cropped to a ~70 nucleotide precursor (Pre-miRNA) by the nuclear ribonuclease Drosha. After export from the nucleus a cytoplasmic RNase (Dicer) cleaves the loop structure of the pre-miRNA yielding a mature 21~24 nucleotide miRNA duplex (Fig. 1). These processing steps are necessary for proper smooth muscle development based on studies of smooth-muscle restricted knockout of Dicer in mice. Knocking out Dicer is known to inhibit blood vessel maturation and intestinal tract development.[17,18] MiRNA processing is also under control of functionally important extracellular signals in vascular smooth muscle. For example, TGF-β family proteins have profound effects on processing of miR-21 in human pulmonary artery cells in culture.[19]
Figure 1.
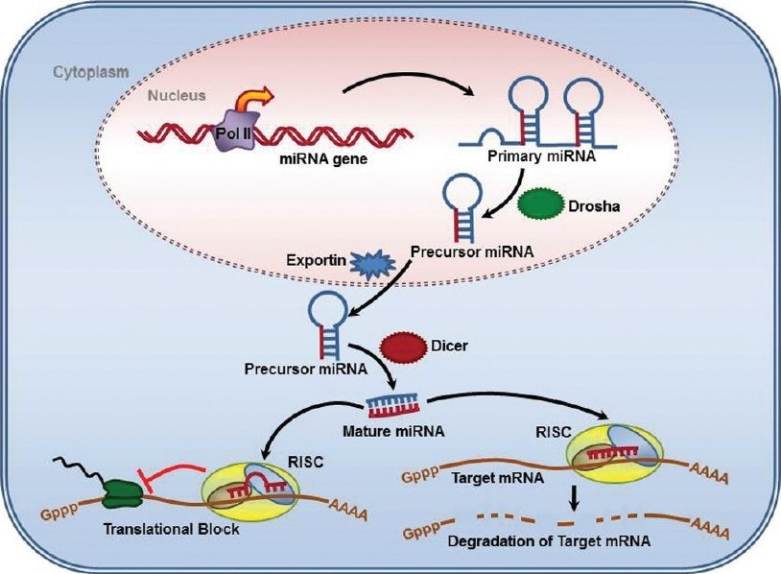
MicroRNA biogenesis and RNA-induced gene silencing. Transcription of primary micro RNA (Pri-miRNA) from miRNA genes is followed by cleavage to precursor mRNA (Pre-miRNA) by the Drosha nuclear RNase III. The Pre-miRNA is then exported to the cytoplasm by exportin via nuclear pore. In the cytoplasm, Pre-miRNA is further processed by RNase activity of Dicer to the mature micro RNA duplex. The duplex loads onto Argonaut ribonucleases in the RISC complex and separates. One of the mature miRNA strands (red strand) mediates small interfering RNA silencing by degrading the target mRNA or interfering with translation. The outcome of RISC formation varies with the degree of complementarity of the seed sequence of miRNA and 3’ untranslated regions (UTR) of the target mRNA.
Gene silencing mediated by the mature miRNAs then occurs by two somewhat different mechanisms that both require the mature miRNA to complex with several proteins including Argonaut family members Ago-1 and Ago-2. The mature dsRNA duplex loads into RNA-induced silencing complexes (RISCs) that mediate posttranscriptional silencing by reducing mRNA stability or by translational block depending on the degree of complementarity of the miRNA seed sequence (nucleotides 2-8) with the target sequence (Fig. 1). MiRNA tends to be cleaved by Ago-2 when complementarity is perfect, although this is not universally true. The transcript can then be further modified by uridinylation and decapping, and then completely degraded by exonuclease cleavage. When complementarity is imperfect a variety of miRNA/mRNA/RISC structures can form that block initiation, cause premature termination, and induce dissociation of ribosomes. MiRNA degradation then occurs following deadenylation, decapping and exonuclease action. Translation of sets of target proteins is thereby reduced.
For the purposes of this review we are interested in miRNA-mediated gene silencing, but the reader should be aware that instances of miRNA-mediated translation enhancement have also been reported.[20] The remainder of the review focuses on particular miRNAs that target genes important in vascular smooth muscle development, contraction and lifespan. The aim is to show how miRNA-induced silencing could alter smooth muscle progenitor differentiation, smooth muscle restricted contractile protein expression, smooth muscle proliferation, and proinflammatory mediator synthesis. Each of these processes participate in vessel wall remodeling that contributes to pathogenesis of pulmonary vascular diseases.
MiRNAs and vascular smooth muscle plasticity
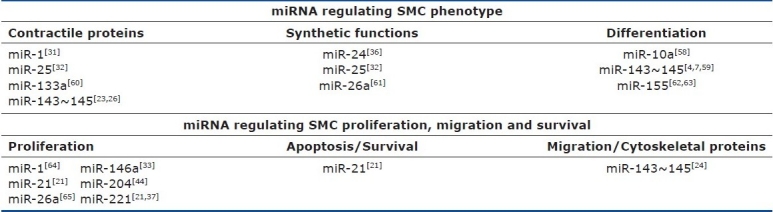
The role of miRNA-mediated gene silencing in vascular smooth muscles was first described in 2007. The number of studies describing miRNAs expressed vascular tissues under a variety of conditions is growing exponentially. There are now reports of miRNAs necessary for normal vascular development as well as miRNAs that are altered in vascular diseases including vascular damage, atherosclerosis and pulmonary hypertension. These miRNAs are sorted into functional groups in Table 1 to illustrate the miRNAs known to contribute to smooth muscle cell fate.
Table 1.
MicroRNAs regulating smooth muscle cell fate
The earliest reports linking miRNAs and vascular smooth muscle remodeling were studies of miRNAs upregulated during injury. Zhang and coworkers discovered miR-21 levels increased following carotid artery injury.[21] They then went on to establish that miR-21 promotes vascular smooth muscle proliferation by silencing expression of phosphatase and tensin homolog (PTEN) and increasing expression of B-cell leukemia/lymphoma 2 (Bcl2). Paradoxically, miR-21 was subsequently shown by Davis et al.[19] to promote contractile protein expression induced by TGF-β family proteins in cultured pulmonary artery vascular smooth muscle by silencing programmed cell death 4 (PDCD4). TGF-β family proteins enhanced processing of the miR-21 primary transcript to the mature miRNA, and increased miR-21 was found to enhance smooth-muscle restricted contractile protein expression.[19] These findings are important because they were the first example of growth-factor regulation of miRNA processing in smooth muscle, and they showed that one miRNA (miR-21) under different conditions promotes either contractile or proliferative phenotypes. It remains to be seen whether other miRNAs also exert dual effects on smooth muscle phenotype, but the early studies point to the complexity and potential duplicity of miRNA targets and physiological effects.
The initial studies of miR-21 in vascular remodeling were quickly followed by a series of landmark studies of the miR-143~145 cluster. Neointimal lesion formation is associated with downregulation of miR-145 as well as downregulation of contractile protein expression and increased proliferation of neointimal cells.[22] Downregulation of miR-143~miR-145 and downregulation of contractile protein expression was then shown to be cause-and-effect in a series of loss of function studies by Cordes et al.[23] Studies in cultured vascular smooth muscle cells and knockout mice have defined a pathway for reciprocal control of Kruppel-like factor 4 (KLF4) and myocardin expression by miR-145 as shown in Figure 2. Knockout mouse studies have corroborated the initial cell culture studies and have verified that the miR-143~145 cluster is a dominant regulator of smooth muscle differentiation. The miR143~145 cluster enhances contractile protein expression required for contractility and proper blood pressure regulation.[24–26] It also has a profound effect: to promote differentiation of stem cells to smooth muscle cells.[4] MiR-145 can directly silence expression of KLF4 and can indirectly upregulate myocardin expression (Fig. 2), which contributes to TGF-β1 enhancement of serum response factor (SRF)-dependent contractile protein expression.[27] SRF regulates a loosely coordinated set of smooth muscle contractile, cytoskeletal and matrix protein genes with CArG boxes in the 5’ untranslated region.[24,28,29]
Figure 2.
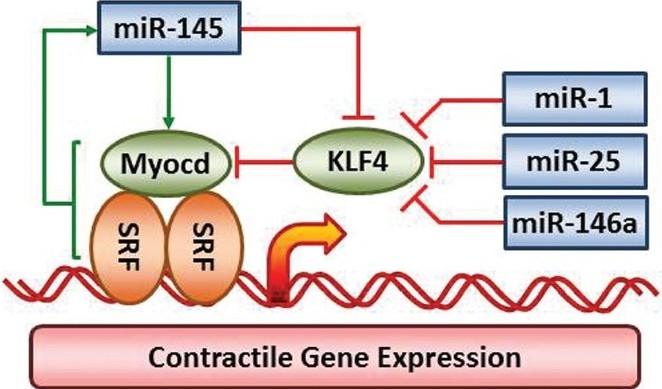
MicroRNAs regulating smooth muscle restricted contractile protein expression. Multiple miRNAs modulate the key transcriptional co-regulators myocardin and KLF4, which are positive and negative regulators of SRF-dependent smooth muscle gene expression. Current evidence shows miR-1, miR-25, miR-133a, miR-146a and miR-145 all modulate expression of either KLF4 or myocardin to influence contractile protein expression. The red lines indicate silencing of protein expression or inhibition of miRNA expression by pathway components. The green arrows indicate activation or upregulation of the pathway component.
In addition to regulating contractile protein expression, miR-143~145 also influences expression of proteins involved in matrix remodeling and cell migration. Downregulation of miR-143~145 upregulates formation of podosomes and upregulates expression of PDGF receptor, protein kinase C (PKC) epsilon and the actin bundling protein fascin.[30] Podosomes are discrete sites of matrix remodeling necessary for invasive migration of vascular smooth muscle cells during vascular wall remodeling. Whether podosome formation is necessary for development of arteriopathy in pulmonary vascular diseases is unknown, but seems plausible given the extensive structural remodeling that occurs in humans and in animal models of pulmonary hypertension.
In addition to the miR-143~145 cluster, other miRNAs can modulate smooth muscle gene expression by altering KLF4 expression (Fig. 2). KLF4 is a direct target of miR-1 in stem cell differentiation,[31] miR-25 in airway smooth muscle,[32] and miR-146a in vascular smooth muscle.[33] Regulation of KLF4 by miR-146a involves a feedback loop in which miR-146a silences KLF4 which competes with KLF5 to reduce transcription of the miR-146a gene. Neointima formation is thereby enhanced by smooth muscle cell proliferation and migration due in part to increased KLF4 expression. The proximal signals that activate the miR-146a-KLF4/KLF5 pathway are not defined in vascular smooth muscle. However, in airway smooth muscle expression of primary-miR-146a expression is activated by nuclear factor kappa beta signaling and primary-miR-146a processing is regulated by MEK-1/2 and JNK-1/2.[34] Mature miR-146a is also induced by stretch in C2C12 myoblast cells.[35] Defining the trigger and upstream transduction pathways in vascular smooth muscle might identify high-value targets for anti-remodeling therapy in PAH.
The peptide growth factors PDGF and TGF-β1 are known to regulate smooth muscle phenotype and to mediate vascular development and remodeling in PAH. PDGF-BB promotes the proliferative/migratory/secretory phenotype in culture and is necessary for proper formation of new blood vessels during development. In contrast, TGF-β family proteins often enhance the contractile phenotype via Smad-dependent signaling. Although they can produce opposing effects on smooth muscle phenotype, both proteins signal changes in primary miRNA transcription and processing. Recent evidence points to signaling convergence of these factors that explain functional antagonism in smooth muscles. PDGF-BB induces expression of miR-24 which directly silences expression of Tribbles-like protein 3 (Trb3) and indirectly decreases Smad1 levels.[36] Overexpression of miR-24 reduces Smad2 and Smad3 expression and reduces TGF-β-mediated activation of Smad2. Therefore, miR-24 is a point of antagonistic signaling convergence for PDGF-BB and the TGF-β family members in vascular smooth muscle. This suggests that miR-24 might be an interesting target to alter vascular remodeling. However, the timing and effect of any intervention with a miR-24 antagonist is difficult to predict given the complex interplay between BMP and TGF-β family members during pathogenesis of pulmonary hypertension. Empirical tests in animal models of pulmonary hypertension at various stages of disease development are needed to establish an effective therapeutic strategy.
The PDGF signaling pathway in vascular smooth muscle also induces expression of miR-221 which might also contribute to neointimal proliferation.[37] MiR- 221 is upregulated in a variety of cancers, and miR-221 silences expression of p27Kip1 during skeletal muscle differentiation.[38] In cultured human pulmonary artery smooth muscle, silencing p27Kip1 by miR-221 overexpression promotes proliferation.[37] In a separate study of rat aorta, smooth muscle miR-221 and miR-222 expression was induced with PDGF, which also decreased p27Kip1 and p57Kip2 expression.[39] MiR-221 and miR-222, like miR-21, are good examples of miRNAs that are conserved in many cells and have consistent effects on cell cycle control proteins in vascular smooth muscle cells. It will be important to determine whether modifying the highly conserved process of cell cycle transit with RNAi-based therapy can be an effective anti-remodeling strategy. The timing of such treatments during development of pulmonary vascular remodeling will be important as was suggested above for modifying growth factor signaling with miR-24 antagonists.
MiRNAs and pulmonary hypertension
Studies of miRNAs in vascular remodeling during development of pulmonary hypertension must consider the multifocal, multicellular nature of the changes in vascular structures. Vascular lesions involving multiple cell types are observed in humans and in animal models of pulmonary arterial hypertension.[40] It seems likely that phenotype modulation of endothelial cells, smooth muscle cells, fibroblasts and both resident and immigrating progenitor cell types occurs.[41] Therefore understanding miRNA expression patterns in each cell type as a function of disease development and degree of severity is vital for designing novel therapeutic strategies.
To this point we have focused on miRNAs in vascular smooth muscle cells; but there is also a significant literature on miRNAs in endothelial cells and various types of pluripotent cells that is highly relevant. Comprehensive discussion of this issue is beyond the scope of this review, but some key observations are worth making. A number of miRNAs described in smooth muscle cells (e.g., miR-21 and miR-221 in Figure 3) have conserved functions in differentiation, proliferation and survival of endothelial cells and other vascular mural cells.[6] The initial studies of miRNAs in the cardiovascular system cited above suggest some likely targets for RNAi-based antagonism of remodeling—e.g., miR-21, miR-145 and miR-221.[42] However, until recently it was not clear which miRNAs were altered during development of pulmonary arterial hypertension (PAH). MiRNAs that promote arterial muscularization, that increase cell survival or proliferation and promote endothelial to mesenchymal transitions are clearly of great interest.
Figure 3.
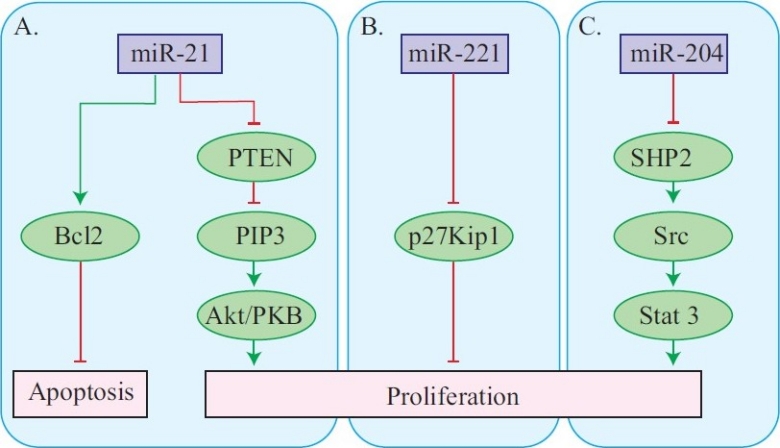
Signal transduction pathways implicated in pathogenesis of vascular remodeling relevant to pulmonary hypertension. Panel A: miR-21 may increase vascular smooth muscle cell number by targeting proteins that regulate cell proliferation (PTEN) and apoptosis (Bcl2). Changes in miR-21 expression have been observed in lung tissues and in vascular smooth muscle in animal models of pulmonary hypertension. Panel B: miR221 promotes vascular smooth muscle cell proliferation by silencing the cell cycle inhibitor p27Kip1. Panel C: miR-204 downregulated in pulmonary hypertension in animals and in human leukocytes can indirectly promote cell proliferation. Derepression of SHP2 expression activates a Src/Stat3 cascade that promotes vascular smooth muscle proliferation.
In the earliest published study of miRNAs in pulmonary hypertension, Caruso et al.[43] surveyed miRNA expression in total lung extracts from two rat models (chronic hypoxia and monocrotaline model) of PAH. MiR-21 was downregulated in the monocrotaline model and in lung samples from humans with PAH. In the same study miR-451 was upregulated in both models but no differences were detected in lung samples from control and PAH human subjects. This initial report showed that there are disease-related changes in miRNAs associated with development of PAH in both animals and humans. A subsequent study by Courboulin et al.[44] found miR-204 was also downregulated in humans with PAH and in rat models of PAH. Delivery of a miR-204 mimic to rat lungs reduced the severity of the disease, providing an exciting proof of principle for rescuing vascular smooth muscle phenotype in vivo with RNAi-based therapy. Courboulin et al.[44] also showed downregulation of expression of miR-204 in mononuclear cells from in blood of PAH patients. This raises the possibility that changes in miRNAs in plasma and leukocytes might be useful biomarkers of PAH pathogenesis.
In the animal models of PAH Courboulin et al.[44] identified potential targets for miR-204. They found Stat3 activation was increased upon attenuation of miR-204 expression and that miR-204 directly regulates SHP2 by targeting its 3’UTR. They developed the signaling model shown in Figure 3 where decreased miR-204 increases SHP2, which by activating Src increases Stat3 activation. Stat3 is hypothesized to promote smooth muscle proliferation and pulmonary vessel wall thickening. This landmark study provides solid proof of principle that “rescue” of low miRNA expression can prevent progression of established PAH.[44] Rapid advances are being made in RNAi therapy of several vascular diseases in which target miRNAs have been identified, some target proteins and processes have been identified, and some demonstration of effective drug delivery has been presented.[45–47]
The study of miR-204 in PAH[44] is unique because roles for miR-204 in myogenesis and in other vascular diseases have not been described before. MiR-204 may have a particular set of functions in the pulmonary circulation that differ qualitatively or quantitatively from its function in other vascular beds. It seems reasonable to speculate that miR-204 is one of several miRNAs that promote differentiation of vascular smooth muscle, and that downregulation of miR-204 might occur during pathogenic vascular remodeling in atherosclerosis and restenosis. However, there are no reports of a strong association or cause-effect in these other vascular diseases. Nevertheless, targets of miR-204 have been validated in other cell types. Some of the target proteins have important roles in smooth muscle cell physiology and vascular diseases: TGF-β receptor 2,[48] epidermal growth factor (EGF) receptor signaling,[49] forkhead box C1 (FOXC1),[50] and runt-related transcription factor 2 (Runx2).[51] It is not clear whether these other targets of miR-204 are also contributing to arteriopathy in pulmonary hypertension, but these are target proteins worthy of further study.
CONCLUSIONS AND FUTURE DIRECTIONS
MiRNA expression surveys have yielded several candidate molecules that could contribute to disease development involving remodeling of smooth muscles. The results of unbiased expression surveys and hypothesis-driven biochemical and functional validation studies have provided important insights into disease mechanisms and potential targets of new treatments of vascular remodeling (Table 1). Some conserved miRNAs have been described in vascular smooth muscle tissues and cells in culture that are known from the cancer literature. MiRNAs controlling cell proliferation and cell survival (miR-21, miR-221 and miR-222) are altered in a variety of diseases. Some miRNAs appear to serve important roles in smooth muscle differentiation unique to this cell type. Novel findings relevant to miRNAs in differentiation were led by investigations of vascular remodeling in disease models and in humans. The best example is the prominent role of the miR-143~145 cluster in regulating KLF4 and myocardin in smooth muscle differentiation. Further investigation is needed to define how miRNAs such as miR-145 can control a set of highly smooth-muscle restricted genes and yet in other settings act as tumor suppressors and regulators of pluripotency.
A number of important questions are raised by the recent surge in knowledge about miRNAs in vascular diseases. One issue is related to the general question of the significance of dynamic changes in epigenetic mechanisms of gene expression in pulmonary vascular diseases. Do pulmonary vascular smooth muscle cells respond to environmental inputs by altering epigenetic factors such as DNA methylation patterns in CpG regions of promoters? Are there diagnostic or prognostic modifications of histones that prime particular genes for expression that is subsequently modulated by the miR-143~145 cluster? Will new assays of miRNA expression and miRNA processing during various stages of disease development illuminate some new biomarkers or new candidates for inhibiting pathological vascular remodeling? Can RNA mimics and antagonists be effective anti-remodeling drugs in vivo?
There are reasons for optimism that RNAi therapy might be a useful anti-remodeling approach. One of the earliest examples of RNAi “therapy” in animals was intranasal delivery of antisense oligonucleotides against a respiratory syncytial virus protein to the lungs of mice inhibited virus replication.[52,53] RNAi therapy can also be scaled up for use in primates. A locked nucleic acid modified miR-122 when administered IV to green monkeys inhibits cholesterol synthesis.[54] Recently RNAi therapies targeting smooth muscle remodeling have also been shown to effective in animal models. Pulmonary hypertension and asthma in animal models are both responsive to lung-restricted delivery of RNAi drugs that rescue (miR-204),[44] or antagonize (miR-145)[55] miRNAs altered by the disease. There is also hope that atherosclerotic plaque stability might be susceptible to manipulation via systemic delivery of RNAi-based drugs.[46,56,57] To address the question of effective RNAi-based anti-remodeling therapy, novel RNAi-based drugs and novel delivery methods must be developed in animal models and then rapidly moved to first-in-human trials. It is now clear that chemically stabilized antisense ribonucleotides and modified miRNAs can be effective “therapy” in animal models of vascular and respiratory diseases. With ongoing development in RNAi drug design and delivery, these approaches should soon be applied to humans.[65]
Footnotes
Source of Support: Nil
Conflict of Interest: None declared.
REFERENCES
- 1.McDonald OG, Owens GK. Programming smooth muscle plasticity with chromatin dynamics. Circ Res. 2007;100:1428–41. doi: 10.1161/01.RES.0000266448.30370.a0. [DOI] [PubMed] [Google Scholar]
- 2.Majesky MW, Dong XR, Regan JN, Hoglund VJ. Vascular smooth muscle progenitor cells: Building and repairing blood vessels. Circ Res. 2011;108:365–77. doi: 10.1161/CIRCRESAHA.110.223800. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Clifford RL, Coward WR, Knox AJ, John AE. Transcriptional regulation of inflammatory genes associated with severe asthma. Curr Pharm Des. 2011;17:653–66. doi: 10.2174/138161211795429000. [DOI] [PubMed] [Google Scholar]
- 4.Xu N, Papagiannakopoulos T, Pan G, Thomson JA, Kosik KS. MicroRNA-145 regulates OCT4, SOX2, and KLF4 and represses pluripotency in human embryonic stem cells. Cell. 2009;137:647–58. doi: 10.1016/j.cell.2009.02.038. [DOI] [PubMed] [Google Scholar]
- 5.Bonauer A, Boon RA, Dimmeler S. Vascular microRNAs. Curr Drug Targets. 2010;11:943–9. doi: 10.2174/138945010791591313. [DOI] [PubMed] [Google Scholar]
- 6.Ohtani K, Dimmeler S. Control of cardiovascular differentiation by microRNAs. Basic Res Cardiol. 2011;106:5–11. doi: 10.1007/s00395-010-0139-7. [DOI] [PubMed] [Google Scholar]
- 7.Mallanna SK, Rizzino A. Emerging roles of microRNAs in the control of embryonic stem cells and the generation of induced pluripotent stem cells. Dev Biol. 2010;344:16–25. doi: 10.1016/j.ydbio.2010.05.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Paulin R, Courboulin A, Barrier M, Bonnet S. From oncoproteins/tumor suppressors to microRNAs, the newest therapeutic targets for pulmonary arterial hypertension. J Mol Med (Berl) 2011;89:1089–101. doi: 10.1007/s00109-011-0788-5. [DOI] [PubMed] [Google Scholar]
- 9.Gairhe S, Bauer NR, Gebb SA, McMurtry IF. Myoendothelial gap junctional signaling induces differentiation of pulmonary arterial smooth muscle cell. Am J Physiol Lung Cell Mol Physiol. 2011 doi: 10.1152/ajplung.00091.2011. [DOI] [PubMed] [Google Scholar]
- 10.Owens GK, Kumar MS, Wamhoff BR. Molecular regulation of vascular smooth muscle cell differentiation in development and disease. Physiol Rev. 2004;84:767–801. doi: 10.1152/physrev.00041.2003. [DOI] [PubMed] [Google Scholar]
- 11.Larsson E, McLean SE, Mecham RP, Lindahl P, Nelander S. Do two mutually exclusive gene modules define the phenotypic diversity of mammalian smooth muscle? Mol Genet Genomics. 2008;280:127–37. doi: 10.1007/s00438-008-0349-y. [DOI] [PubMed] [Google Scholar]
- 12.Majesky MW. Developmental basis of vascular smooth muscle diversity. Arterioscler Thromb Vasc Biol. 2007;27:1248–58. doi: 10.1161/ATVBAHA.107.141069. [DOI] [PubMed] [Google Scholar]
- 13.Eddinger TJ, Meer DP. Myosin II isoforms in smooth muscle: Heterogeneity and function. Am J Physiol Cell Physiol. 2007;293:C493–508. doi: 10.1152/ajpcell.00131.2007. [DOI] [PubMed] [Google Scholar]
- 14.Ku G, McManus MT. Behind the scenes of a small RNA gene-silencing pathway. Hum Gene Ther. 2008;19:17–26. doi: 10.1089/hum.2007.1226. [DOI] [PubMed] [Google Scholar]
- 15.Farazi TA, Spitzer JI, Morozov P, Tuschl T. miRNAs in human cancer. J Pathol. 2011;223:102–15. doi: 10.1002/path.2806. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Kulshreshtha R, Ferracin M, Wojcik SE, Garzon R, Alder H, Agosto-Perez FJ, et al. A microRNA signature of hypoxia. Mol Cell Biol. 2007;27:1859–67. doi: 10.1128/MCB.01395-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Albinsson S, Skoura A, Yu J, DiLorenzo A, Fernández-Hernando C, Offermanns S, et al. Smooth muscle miRNAs are critical for post-natal regulation of blood pressure and vascular function. PLoS One. 2011;6:e18869. doi: 10.1371/journal.pone.0018869. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Park C, Yan W, Ward SM, Hwang SJ, Wu Q, Hatton WJ, et al. MicroRNAs dynamically remodel gastrointestinal smooth muscle cells. PLoS One. 2011;6:e18628. doi: 10.1371/journal.pone.0018628. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Davis BN, Hilyard AC, Lagna G, Hata A. SMAD proteins control DROSHA-mediated microRNA maturation. Nature. 2008;454:56–61. doi: 10.1038/nature07086. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Vasudevan S, Tong Y, Steitz JA. Switching from repression to activation: microRNAs can up-regulate translation. Science. 2007;318:1931–4. doi: 10.1126/science.1149460. [DOI] [PubMed] [Google Scholar]
- 21.Ji R, Cheng Y, Yue J, Yang J, Liu X, Chen H, et al. MicroRNA expression signature and antisense-mediated depletion reveal an essential role of MicroRNA in vascular neointimal lesion formation. Circ Res. 2007;100:1579–88. doi: 10.1161/CIRCRESAHA.106.141986. [DOI] [PubMed] [Google Scholar]
- 22.Cheng Y, Liu X, Yang J, Lin Y, Xu DZ, Lu Q, et al. MicroRNA-145, a novel smooth muscle cell phenotypic marker and modulator, controls vascular neointimal lesion formation. Circ Res. 2009;105:158–66. doi: 10.1161/CIRCRESAHA.109.197517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Cordes KR, Sheehy NT, White MP, Berry EC, Morton SU, Muth AN, et al. miR-145 and miR-143 regulate smooth muscle cell fate and plasticity. Nature. 2009;460:705–10. doi: 10.1038/nature08195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Xin M, Small EM, Sutherland LB, Qi X, McAnally J, Plato CF, et al. MicroRNAs miR-143 and miR-145 modulate cytoskeletal dynamics and responsiveness of smooth muscle cells to injury. Genes Dev. 2009;23:2166–78. doi: 10.1101/gad.1842409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Elia L, Quintavalle M, Zhang J, Contu R, Cossu L, Latronico MV, et al. The knockout of miR-143 and -145 alters smooth muscle cell maintenance and vascular homeostasis in mice: Correlates with human disease. Cell Death Differ. 2009;16:1590–8. doi: 10.1038/cdd.2009.153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Boettger T, Beetz N, Kostin S, Schneider J, Krüger M, Hein L, et al. Acquisition of the contractile phenotype by murine arterial smooth muscle cells depends on the Mir143/145 gene cluster. J Clin Invest. 2009;119:2634–47. doi: 10.1172/JCI38864. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Davis-Dusenbery BN, Chan MC, Reno KE, Weisman AS, Layne MD, Lagna G, et al. down-regulation of Kruppel-like factor-4 (KLF4) by microRNA-143/145 is critical for modulation of vascular smooth muscle cell phenotype by transforming growth factor-beta and bone morphogenetic protein 4. 2011;286:28097-110. J Biol Chem. 2011;286:28097–110. doi: 10.1074/jbc.M111.236950. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Park C, Hennig GW, Sanders KM, Cho JH, Hatton WJ, Redelman D, et al. Serum response factor-dependent MicroRNAs regulate gastrointestinal smooth muscle cell phenotypes. Gastroenterology. 2011;141:164–75. doi: 10.1053/j.gastro.2011.03.058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Long X, Miano JM. TGF{beta}1 utilizes distinct pathways for the transcriptional activation of microRNA 143/145 in human coronary artery smooth muscle cells. J Biol Chem. 2011;286:30119–29. doi: 10.1074/jbc.M111.258814. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Quintavalle M, Elia L, Condorelli G, Courtneidge SA. MicroRNA control of podosome formation in vascular smooth muscle cells in vivo and in vitro. J Cell Biol. 2010;189:13–22. doi: 10.1083/jcb.200912096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Xie C, Huang H, Sun X, Guo Y, Hamblin M, Ritchie RP, et al. MicroRNA-1 regulates smooth muscle cell differentiation by repressing Kruppel-like factor 4. Stem Cells Dev. 2011;20:205–10. doi: 10.1089/scd.2010.0283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Kuhn AR, Schlauch K, Lao R, Halayko AJ, Gerthoffer WT, Singer CA. MicroRNA expression in human airway smooth muscle cells: Role of miR-25 in regulation of airway smooth muscle phenotype. Am J Respir Cell Mol Biol. 2010;42:506–13. doi: 10.1165/rcmb.2009-0123OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Sun SG, Zheng B, Han M, Fang XM, Li HX, Miao SB, et al. miR-146a and Kruppel-like factor 4 form a feedback loop to participate in vascular smooth muscle cell proliferation. EMBO Rep. 2011;12:56–62. doi: 10.1038/embor.2010.172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Larner-Svensson HM, Williams AE, Tsitsiou E, Perry MM, Jiang X, Chung KF, et al. Pharmacological studies of the mechanism and function of interleukin-1beta-induced miRNA-146a expression in primary human airway smooth muscle. Respir Res. 2010;11:68. doi: 10.1186/1465-9921-11-68. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Kuang W, Tan J, Duan Y, Duan J, Wang W, Jin F, et al. Cyclic stretch induced miR-146a upregulation delays C2C12 myogenic differentiation through inhibition of Numb. Biochem Biophys Res Commun. 2009;378:259–63. doi: 10.1016/j.bbrc.2008.11.041. [DOI] [PubMed] [Google Scholar]
- 36.Chan MC, Hilyard AC, Wu C, Davis BN, Hill NS, Lal A, et al. Molecular basis for antagonism between PDGF and the TGFbeta family of signalling pathways by control of miR-24 expression. EMBO J. 2010;29:559–73. doi: 10.1038/emboj.2009.370. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Davis BN, Hilyard AC, Nguyen PH, Lagna G, Hata A. Induction of microRNA-221 by platelet-derived growth factor signaling is critical for modulation of vascular smooth muscle phenotype. J Biol Chem. 2009;284:3728–3738. doi: 10.1074/jbc.M808788200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Cardinali B, Castellani L, Fasanaro P, Basso A, Alemà S, Martelli F, et al. Microrna-221 and microrna-222 modulate differentiation and maturation of skeletal muscle cells. PLoS One. 2009;4:e7607. doi: 10.1371/journal.pone.0007607. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Liu X, Cheng Y, Zhang S, Lin Y, Yang J, Zhang C. A necessary role of miR-221 and miR-222 in vascular smooth muscle cell proliferation and neointimal hyperplasia. Circ Res. 2009;104:476–87. doi: 10.1161/CIRCRESAHA.108.185363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Abe K, Toba M, Alzoubi A, Ito M, Fagan KA, Cool CD, et al. Formation of plexiform lesions in experimental severe pulmonary arterial hypertension. Circulation. 2010;121:2747–54. doi: 10.1161/CIRCULATIONAHA.109.927681. [DOI] [PubMed] [Google Scholar]
- 41.Toshner M, Morrell NW. Endothelial progenitor cells in pulmonary hypertension - dawn of cell-based therapy. Int J Clin Pract. 2010;165(Supp):7–12. doi: 10.1111/j.1742-1241.2009.02232.x. [DOI] [PubMed] [Google Scholar]
- 42.Zhang C. MicroRNA-145 in vascular smooth muscle cell biology: A new therapeutic target for vascular disease. Cell Cycle. 2009;8(21):3469–3473. doi: 10.4161/cc.8.21.9837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Caruso P, MacLean MR, Khanin R, McClure J, Soon E, Southgate M, et al. Dynamic changes in lung microRNA profiles during the development of pulmonary hypertension due to chronic hypoxia and monocrotaline. Arterioscler Thromb Vasc Biol. 2010;30:716–23. doi: 10.1161/ATVBAHA.109.202028. [DOI] [PubMed] [Google Scholar]
- 44.Courboulin A, Paulin R, Giguère NJ, Saksouk N, Perreault T, Meloche J, et al. Role for miR-204 in human pulmonary arterial hypertension. J Exp Med. 2011;208:535–48. doi: 10.1084/jem.20101812. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Albinsson S, Sessa WC. Can microRNAs control vascular smooth muscle phenotypic modulation and the response to injury? Physiol Genomics. 2011;43:529–33. doi: 10.1152/physiolgenomics.00146.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Zhang C. MicroRNA and vascular smooth muscle cell phenotype: New therapy for atherosclerosis? Genome Med. 2009;1:85. doi: 10.1186/gm85. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Wang S, Olson EN. AngiomiRs--key regulators of angiogenesis. Curr Opin Genet Dev. 2009;19:205–11. doi: 10.1016/j.gde.2009.04.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Wang FE, Zhang C, Maminishkis A, Dong L, Zhi C, Li R, et al. MicroRNA-204/211 alters epithelial physiology. FASEB J. 2010;24:1552–71. doi: 10.1096/fj.08-125856. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Lee Y, Yang X, Huang Y, Fan H, Zhang Q, Wu Y, et al. Network modeling identifies molecular functions targeted by miR-204 to suppress head and neck tumor metastasis. PLoS Comput Biol. 2010;6:e1000730. doi: 10.1371/journal.pcbi.1000730. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Chung TK, Lau TS, Cheung TH, Yim SF, Lo KW, Siu NS, et al. Dysregulation of microRNA-204 mediates migration and invasion of endometrial cancer by regulating FOXC1. Int J Cancer. 2011 Mar 11; doi: 10.1002/ijc.26060. doi: 101002/ijc26060 [Epub ahead of print] [DOI] [PubMed] [Google Scholar]
- 51.Zhang Y, Xie RL, Croce CM, Stein JL, Lian JB, van Wijnen AJ, et al. A program of microRNAs controls osteogenic lineage progression by targeting transcription factor Runx2. Proc Natl Acad Sci U S A. 2011;108:9863–8. doi: 10.1073/pnas.1018493108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Bitko V, Musiyenko A, Shulyayeva O, Barik S. Inhibition of respiratory viruses by nasally administered siRNA. Nat Med. 2005;11:50–5. doi: 10.1038/nm1164. [DOI] [PubMed] [Google Scholar]
- 53.Bitko V, Barik S. Nasal delivery of siRNA. Methods Mol Biol. 2008;442:75–82. doi: 10.1007/978-1-59745-191-8_6. [DOI] [PubMed] [Google Scholar]
- 54.Elmén J, Lindow M, Schütz S, Lawrence M, Petri A, Obad S, et al. LNA-mediated microRNA silencing in non-human primates. Nature. 2008;452:896–9. doi: 10.1038/nature06783. [DOI] [PubMed] [Google Scholar]
- 55.Collison A, Herbert C, Siegle JS, Mattes J, Foster PS, Kumar RK. Altered expression of microRNA in the airway wall in chronic asthma: miR-126 as a potential therapeutic target. BMC Pulm Med. 2011;11:29. doi: 10.1186/1471-2466-11-29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.O’Sullivan JF, Martin K, Caplice NM. Microribonucleic acids for prevention of plaque rupture and in-stent restenosis: “a finger in the dam”. J Am Coll Cardiol. 2011;57:383–9. doi: 10.1016/j.jacc.2010.09.029. [DOI] [PubMed] [Google Scholar]
- 57.Jamaluddin MS, Weakley SM, Zhang L, Kougias P, Lin PH, Yao Q, et al. miRNAs: roles and clinical applications in vascular disease. Expert Rev Mol Diagn. 2011;11:79–89. doi: 10.1586/erm.10.103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Huang H, Xie C, Sun X, Ritchie RP, Zhang J, Chen YE. miR-10a contributes to retinoid acid-induced smooth muscle cell differentiation. J Biol Chem. 2010;285:9383–9. doi: 10.1074/jbc.M109.095612. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Sachdeva M, Zhu S, Wu F, Wu H, Walia V, Kumar S, et al. p53 represses c-Myc through induction of the tumor suppressor miR-145. Proc Natl Acad Sci U S A. 2009;106:3207–12. doi: 10.1073/pnas.0808042106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Liu N, Olson EN. MicroRNA regulatory networks in cardiovascular development. Dev Cell. 2010;18:510–25. doi: 10.1016/j.devcel.2010.03.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Mohamed JS, Lopez MA, Boriek AM. Mechanical stretch up-regulates microRNA-26a and induces human airway smooth muscle hypertrophy by suppressing glycogen synthase kinase-3beta. J Biol Chem. 2010;285:29336–47. doi: 10.1074/jbc.M110.101147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Zheng L, Xu CC, Chen WD, Shen WL, Ruan CC, Zhu LM, et al. MicroRNA-155 regulates angiotensin II typ.1 receptor expression and phenotypic differentiation in vascular adventitial fibroblasts. Biochem Biophys Res Commun. 2010;400:483–8. doi: 10.1016/j.bbrc.2010.08.067. [DOI] [PubMed] [Google Scholar]
- 63.Martin MM, Buckenberger JA, Jiang J, Malana GE, Nuovo GJ, Chotani M, et al. The human angiotensin II typ.1 receptor +1166 A/C polymorphism attenuates microrna-155 binding. J Biol Chem. 2007;282:24262–9. doi: 10.1074/jbc.M701050200. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 64.Chen J, Yin H, Jiang Y, Radhakrishnan SK, Huang ZP, Li J, et al. Induction of microRNA-1 by myocardin in smooth muscle cells inhibits cell proliferation. Arterioscler Thromb Vasc Biol. 2011;31:368–75. doi: 10.1161/ATVBAHA.110.218149. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Leeper NJ, Raiesdana A, Kojima Y, Chun HJ, Azuma J, Maegdefessel L, et al. MicroRNA-26a is a novel regulator of vascular smooth muscle cell function. J Cell Physiol. 2011;226:1035–43. doi: 10.1002/jcp.22422. [DOI] [PMC free article] [PubMed] [Google Scholar]