

Summary
Purpose
KCNJ10 encodes subunits of inward rectifying potassium (Kir) channel Kir4.1 found predominantly in glial cells within the brain. Genetic inactivation of these channels in glia impairs extracellular K+ and glutamate clearance and produces a seizure phenotype. In both mice and humans, polymorphisms and mutations in the KCNJ10 gene have been associated with seizure susceptibility. The purpose of the present study was to determine whether there are differences in Kir channel activity and potassium and glutamate buffering capabilities between astrocytes from seizure resistant C57BL/6 (B6) and seizure susceptible DBA/2 (D2) mice that are consistent with an altered K+ channel activity as a result of genetic polymorphism of KCNJ10.
Methods
Using cultured astrocytes and hippocampal brain slices together with whole-cell patch-clamp, we determined the electrophysiological properties, particularly K+ conductances, of B6 and D2 mouse astrocytes. Using a colorimetric assay, we determined glutamate clearance capacity by B6 and D2 astrocytes.
Results
Barium-sensitive Kir currents elicited from B6 astrocytes are substantially larger than those elicited from D2 astrocytes. In addition, potassium and glutamate buffering by D2 cortical astrocytes is impaired, relative to buffering by B6 astrocytes.
Discussion
In summary, the activity of Kir4.1 channels differs between seizure susceptible D2 and seizure resistant B6 mice. Reduced activity of Kir4.1 channels in astrocytes of D2 mice is associated with deficits in potassium and glutamate buffering. These deficits may, in part, explain the relatively low seizure threshold of D2 mice.
Keywords: astrocyte, seizure, potassium buffering, glutamate clearance, Kir4.1
Introduction
Potassium and glutamate homeostases play important roles in regulating neuronal excitability. Increased extracellular concentrations of K+ and glutamate result in hyperexcitability of neurons and abnormal synaptic transmission and their accumulation has been associated with seizure activity (Rothstein et al., 1996; Janigro et al., 1997; Tanaka et al., 1997; D’Ambrosio et al., 1998; Djukic et al., 2007). Astrocytes are primarily responsible for buffering K+ and glutamate. Inward rectifying K+ (Kir) channels, in particular those generated by Kir4.1 subunits, are directly involved in extracellular K+ buffering (Neusch et al., 2006; Kucheryavykh et al., 2007; Djukic et al., 2007; Olsen et al., 2007) and are also involved in glutamate clearance (Kucheryavykh et al., 2007; Djukic et al., 2007).
Kir4.1, first cloned by Takumi and colleagues (1995), was found to generate currents that were described as ATP-dependent inward rectifier potassium channels, and expressed predominantly in glial cells (Poopalasundaram et al., 2000; Schröder et al., 2002; Li et al, 2001; Higashi et al., 2001). In mice in which this Kir subunit was genetically inactivated, K+ conductance was reduced (Kofuji et al., 2000; Djukic et al., 2007) and mice displayed a seizure phenotype (Djukic et al., 2007).
In both mice and humans, polymorphisms or mutations in KCNJ10, the gene encoding Kir4.1 subunits, have been associated with seizures. Initially, quantitative trait loci (QTL) mapping in C57BL/6J (B6) and DBA/2J (D2) mice identified a distal chromosome 1 locus that has a major effect on maximal electroshock seizure threshold (MEST) (Ferraro et al., 2001), a classical measure of seizure susceptibility. The QTL was fine-mapped to a 6 Mb critical interval (Ferraro et al., 2004) and the creation of transgenic mouse lines supported Kcnj10 (Kir4.1) as a quantitative trait gene at this locus (Ferraro et al., 2007). An amino acid substitution at position 262 of Kir4.1 (threonine in B6, serine in D2) is postulated to contribute to the genetic mechanism involved in the control of seizure susceptibility in mice (Ferraro et al., 2004). In humans, a nearby amino acid substitution in Kir4.1, Arg271Cys has been shown to be associated with common forms of epilepsy in two independent studies such that control individuals are significantly more likely to harbor the minor Cys allele (Buono et al., 2004; Lenzen et al., 2005). Interestingly, neither the mouse nor human Kir4.1 variants described above alter the biophysical properties of inward rectifying potassium channels (Shang et al., 2005). Very recently, rare mutations that substantially reduce channel conductance have been identified as the cause of severe clinical syndromes that include epilepsy (Scholl et al., 2009; Bockenhauer et al., 2009).
The purpose of the present study was to determine whether astrocytes derived from B6 and D2 mice differ with regard to Kir channel activity and their capabilities for buffering potassium and glutamate, and whether any such differences might be consistent with differences in their respective seizure susceptibility.
Methods
Animals
In vitro slice experiments utilized male mice from the strains B6 and D2 purchased from Charles River Laboratories (Willmington, MA). Experiments with cultured astrocytes utilized both male and female mice. Mice were maintained on a 12 hour light/dark schedule and had access to food and water ad libitum. All experiments were approved by the Institutional Animal Care and Use Committee.
Electrophysiology Recording from Cultured Astrocytes
Primary cultures of astrocytes were prepared from neocortex of 1–2 day old mice as previously described (Kucheryavykh et al., 2007). Membrane currents were measured with the single electrode whole-cell patch-clamp technique. Two Narishige hydraulic micromanipulators (Narishige, MMW-203, Japan) were used for (1) voltage-clamp recording, and (2) positioning a micropipette with 30 – 50 μm tip diameter for application of test solutions. A five valve system for fast drug application (MS Concentration Clamp; VS-2001, Vibraspec, PA), controlled by a second computer, was connected to the outlets.
Electrodes from hard glass (GC-150-10 glass tubing, Clark Electromedical Instruments, England) were pulled in four steps using a Sutter P-97 puller (Novato, CA). After filling with intracellular solution containing (in mM): 141 KCl, 1 MgCl2, 1 CaCl2, 10 EGTA, 10 HEPES, 3 Na2ATP, 0.25 spermine HCl, pH adjusted to 7.2 with NaOH/HCl, they had resistances of 4–6 MΩ; after cell penetration, the access resistance was 10–15 MΩ, compensated by at least 75%. The extracellular solution contained (in mM): 138 NaCl, 2 CaCl2, 1.9 MgCl2, and 10 HEPES; KCl varied from 1 to 30 mM (substituted by NaCl to adjust osmolarity to 308 mOsm). High frequencies (>1 kHz) were cut off, using an Axopatch-200B amplifier and a CV-203BU headstage, and digitized at 5 kHz through a DigiData 1200A interface (Axon Instruments, CA). The pClamp 9 software (Axon Instruments, CA) was used for data acquisition and analysis.
The membrane potentials were determined immediately after attainment of whole-cell mode, and cells were then subsequently held under voltage-clamp at this potential. Applications of solutions with increased K+ were used to assess potassium uptake ability of the cell as the shift in inward current following shift in [K+]o from 3 mM to 10 or 30 mM (Skatchkov et al., 1999).
Electrophysiological Recording from Astrocytes in Brain Slices
Male B6 and D2 mice (30 – 72 days postnatal) were decapitated and horizontal slices (250 μM) containing the hippocampal area were prepared using a vibratome (VT1000S, Leica, Germany). Slices were cut in ice-cold oxygenated artificial cerebrospinal fluid (ACSF) containing (in mM): 127 NaCl; 2.5 KCl; 1.25 NaH2PO4; 25 NaHCO3; 2 CaCl2; 1 MgCl2; and 25 D-glucose. The solution was saturated with 95%O2–5%CO2 to achieve pH 7.4. For the test application, external KCl (10 mM) or KCl (10 mM) with BaCl2 (100 μM) was added to ACSF.
Membrane currents were measured with the single electrode whole-cell patch-clamp technique. Borosilicate glass patch pipettes (O.D. 1.5 mm, I.D. 1,0 mm; WPI, Sarasota, FL) were pulled to a final resistance of 8–10 MΩ for astrocyte recordings in four steps using a Sutter P-97 puller (Novato, CA). The intracellular solution contained (in mM): 130 K-gluc, 10 Na- gluc, 4 NaCl, 4 phosphocreatine, 0.3 GTP-Na, 4 Mg–ATP, 10 HEPES, and the pH was adjusted to 7.2 adjusted with KOH. Cells in the hippocampal CA1 area were visualized using an Olympus infrared microscope equipped with DIC (BX51WI Olympus, Japan). Two motorized micromanipulators (MX7500 with MC-1000 drive, Siskiyou, OR) were used for voltage-clamp and current-clamp recording. A two-channel MultiClamp 700A patch-clap amplifier with DigiData 1322A interface (Axon Instruments, CA) was used for recording and stimulation. The pClamp 9 software (Axon Instruments, CA) was used for data acquisition and analysis.
Colorimetric Assay to Assess Glutamate Clearance by Astrocytes
To evaluate the glutamate clearance capacity, astrocyte cultures were grown in 24 well dishes. Glutamate remaining in the medium was determined as we have previously described (Kucheryavykh et al., 2007; 2009) using the protocol of Abe et al. (2000). Briefly, medium in each well of the dish was replaced with 250 μl of serum free media containing 400 μM glutamate. After 60 min, the medium was removed and 50 μl of culture supernatant was transferred to 96-well culture plates, and mixed with 50 μl of substrate mixture (20 U/ml glutamate dehydrogenase, 2.5 mg/ml β-nicotineamide adenine dinucleotide (NAD), 0.25 mg/ml 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium (MTT), 100 μM 1-Methoxyphenazine methosulfate (MPMS), 0.1% (v/v) Triton X-100 and 0.2 M Tris- HCl (pH)8.2). Glutamate dehydrogenation was allowed to proceed for 10 min at 37°C and then stopped by adding 100 μl of a solution containing 50% dimethylformamide and 20% sodium dodecyl sulfate (pH 4.7). The production of MTT formazan was assessed by measurement of absorbance at 550 nm using a microplate reader. A standard curve was constructed in each assay using cell-free culture medium containing known concentrations of glutamate. The concentration of extracellular glutamate in the samples was estimated from the standard curve. As a control for each experiment, serum-free medium containing 400 μM glutamate was added to empty wells of a 24 well dish (no astrocytes) and processed together with the astrocytes, i.e. 60 minutes in the incubator until sample collection. In our hands, the lower limit of sensitivity of this assay is approximately 75μM glutamate. The concentration of glutamate remaining in the medium is a reflection of glutamate taken up by the astrocytes, as well as that released from the astrocytes.
Chemicals
TBOA ((3S)-3-[[3-[[4-(Trifluoromethyl) benzoyl]amino] phenyl]methoxy]-L-aspartic acid) was purchased from Tocris Bioscience (Bristol, UK). All other chemicals were purchased from Sigma Chemical Company (St. Louis, MO).
Statistical Analysis
The Student’s t-test for independent samples was used to analyze electrophysiological data, whereas one-way ANOVA followed by Tukey’s (Multiple Comparison Test) was used to analyze the glutamate clearance experiments.
Results
Because genetic polymorphisms in Kcnj10 (gene encoding for Kir4.1 channels) have been implicated in seizure susceptibility differences between C57BL/6J (B6) and DBA/2J (D2) mice, we examined the Kir channel activity in cultured cortical astrocytes from B6 and D2 mice to see if it differed between the two mouse strains. Astrocytes were perfused with a physiological salt solution containing 3 mM K+ and dialyzed with a pipette solution containing 150 mM K+. Under these conditions, application of 100 ms steps to potentials between −100 mV and +100 mV from a holding potential equal to the resting membrane potential evoked both inward and outward currents that were substantially larger in B2 astrocytes than those elicited in D2 astrocytes (Figure 1). Upon application of the Kir channel blocker, Ba2+ (200 μM), currents in astrocytes from B6 mice were inhibited, particularly in the inward direction (Figure 1 A, B1, B2), but there was little effect on the current-voltage curve of D2 astrocytes (Figure 1 C, D1, D2). Taken together, the smaller current elicited from D2 astrocytes and the lack of response to barium application indicate that functional (membrane) Kir channel activity is substantially reduced in astrocytes of D2 mice as compared with activity in B6 astrocytes.
Fig. 1.

A and C: I–V curves from astrocytes of B6 and D2 mice. Whole-cell current-voltage relations of astrocytes from B6 mice (A; n=10) and D2 mice (C; n=19) in the absence (circles) and presence (inverted triangles) of 200 μM barium. Currents were recorded in response to 20 mV steps from −100 mV to +100 mV from Vm. In these experiments, the cell was held at the steady state potential (Vh=Vm). The mean Vm values and S.E.M. are −51.5 ± 4.1 and −45.9 ± 3.2 for B6 and D2 astrocytes, respectively. These values are not statistically different. B and D: Representative current traces from astrocytes of B6 and D2 mice in response to 20 mV steps from 100 mV to +100 mV from Vm. B shows representative current traces from a B6 astrocyte in the absence (B1) and presence (B2) of 200 μM barium. D shows representative current traces from a D2 astrocyte in the absence (D1) and presence (D2) of 200 μM barium.
We next assessed the ability of cultured astrocytes to buffer potassium by measuring membrane currents in response to extracellular [K+] steps (Skatchkov et al., 1999; Zhou and Kimelberg, 2000). The astrocytes were used for studies 3 weeks after plating. To measure the currents evoked by different physiological and pathological [K+]o, extracellular solutions with 1, 10 and 30 mM K+ (normal bath = 3 mM K+) were applied (‘K+ steps’; duration of step = 1 – 1.5 sec). Figure 2 summarizes the current response to K+ steps. The inward currents in response to 10 and 30 mM K+ steps in D2 astrocytes were less than 20% of currents elicited from B6 astrocytes, indicating that the ability of D2 astrocytes to take up K+ was impaired substantially.
Fig. 2.

Summary of the current responses of B6 and D2 astrocytes to changes in [K+]o. The astrocytes were bathed in solution containing 3 mM K+ and this solution was changed to 1, 10 or 30 mM K+ and the current response measured using the whole-cell voltage clamp technique. (n=12 and 13 for B6 and D2, respectively). * indicates significant difference from B6 group (p<0.01; Student’s t-Test for independent samples). In these experiments, the cell was held at the steady state potential (Vh=Vm). The mean Vm values and S.E.M. are −55.8 ± 3.3 and −48.8 ± 4.6 for B6 and D2 astrocytes, respectively. These values are not statistically different.
We next examined the activity of Kir channels in astrocytes in brain slices. These studies were done for two reasons. The first was to demonstrate that the differences in Kir channel activity and potassium uptake by astrocytes of B6 and D2 mice were not due to anomalies resulting from the use of cultured astrocytes. The second was to be able to correlate the findings using mice of similar age as those used for seizure susceptibility studies (Ferraro et al., 1998). It has been shown that Kir4.1 channel expression is stable in glia by two weeks (Seifert et al., 2009; Schopf et al., 2004). Therefore, the present experiment utilized 30 – 72 day old mice in which Kir4.1 channel expression was at maximal levels and could, therefore, be directly compared with the 56–63 day old mice used by Ferraro et al. (1998).
Using whole cell voltage clamp to record from astrocytes in stratum radiatum area of hippocampal slices from B6 and D2 mouse, we measured inward K+ current in response to switching the external solution from one containing 2.5 mM K+ to one containing 10 mM K+ (Fig. 3A), in the presence and absence of 100 μM Ba2+. At this concentration, Ba2+ is a relatively selective blocker of Kir channels. We found that there was significantly less barium sensitive Kir current in D2 astrocytes (Fig. 3B), consistent with the data obtained using cultured astrocytes.
Fig. 3.

A. Representative whole cell currents recorded from B6 and D2 astrocytes in hippocampal brain slices. Inward currents were obtained by changing extracellular K+ from 2.5 mM to 10 mM in the presence or absence of 100 μM Ba2+. The cells were held at the steady state potential (Vh=Vm). The scales bars are equal for all current traces in Part A. B. Summary of the relative barium sensitive Kir currents measured in B6 and D2 astrocytes (n= 9 and 14, respectively). The data are expressed as % of control current that is barium sensitive where control is the maximal current measured by switching extracellular [K+] from 2.5 to 10 mM. * indicates significant difference from B6 group (p<0.01; Student’s t-Test for independent samples). The mean Vm values and S.E.M. are −82.6 ± 1.1 and −85.6 ± 2.2 for B6 and D2 astrocytes, respectively. These values are not statistically different.
Because decreased expression of Kir4.1 channel subunits has been shown to cause decreased glutamate transport by astrocytes (Kucheryavykh et al., 2007; Djukic et al., 2007), we next examined the glutamate uptake capability of cultured astrocytes from B6 and D2 mice. In preliminary experiments (not shown), we identified a time-dependent reduction in glutamate in the medium over 120 min of incubation, but no change in dishes that do not contain astrocytes. Based on these preliminary experiments, we assessed glutamate concentration remaining in the medium, 60 minutes after incubation with 250 μl of serum-free culture medium containing 400 μM glutamate with the presence or absence of the glutamate transporter inhibitor TBOA (200 μM). Glutamate concentrations in the medium of B6 and D2 astrocytes were reduced by 53% and 37%, respectively (Figure 4). This indicates that glutamate clearance by D2 astrocytes was less efficient than that by B6 astrocytes. In addition, glutamate clearance by both B6 and D2 astrocytes was eliminated by addition of TBOA (Figure 4). This suggests that the differences in glutamate clearance by B6 and D2 astrocytes are due to differences in glutamate transporter function and not glutamate release mechanisms. Furthermore, the differences in glutamate clearance are not due to increased astrocytic cell death in D2 astrocytes upon application of glutamate. Using the Invitrogen Live/Dead Viability/Cytotoxicity kit, we observed virtually 100% viability of both B6 and D2 astrocytes after 60 min incubation in 400μM glutamate (data not shown).
Fig. 4.
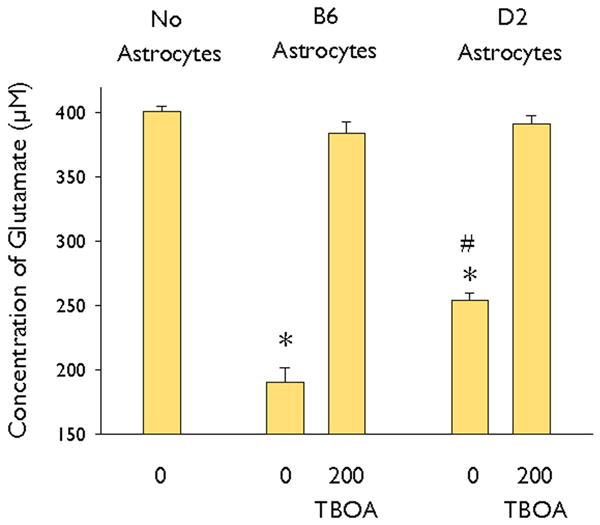
The effect of TBOA on glutamate clearance by B6 and D2 astrocytes. The concentration of glutamate was determined using a colorimetric assay 60 min after addition of 400 μM glutamate in medium and compared with the concentration of glutamate measured in the absence of astrocytes. B6 astrocytes cleared significantly more glutamate in 60 mins than D2 astrocytes. TBOA (200μM) completely blocked glutamate clearance by both B6 and D2 astrocytes. * indicates significant difference from control (no astrocytes) and # indicates a significant difference between the B6 and D2 astrocytes (p<0.01; ANOVA followed by Tukey’s test) with n = 12 per group.
Discussion
Although it was long accepted that dysfunction of neurons was the sole cause of seizures and epilepsy, it is now becoming increasingly apparent that changes in astrocyte function can also contribute to epileptogenesis. Reactive astrocytes can release glutamate and trigger seizures (Tian et al., 2005). Metabolic inhibition of astrocytes can cause swelling and release of glutamate which causes neuronal hyperexcitability (Broberg et al., 2008). Even more directly related to the current study, mutations of Kir4.1 channel subunits that result in decreased Kir activity have been found in patients with epilepsy and seizure phenotypes (Scholl et al., 2009; Bockenhauer et al., 2009). Kir4.1 subunits underlie the major Kir conductance in Müller cells and astrocytes (Kofuji et al., 2000; Neusch et al., 2006; Olsen et al. 2007; Seifert et al., 2009) and have a predominant role in K+ clearance (Neusch et al., 2006; Kucheryavykh et al., 2007; Djukic et al., 2007). In addition, knock-down of Kir4.1 impairs glutamate clearance by astrocytes (Kucheryavykh et al., 2007; Djukic et al., 2007). The present study demonstrates that Kir channel activity is lower in astrocytes from seizure susceptible D2 mice as compared with seizure resistant B6 mice. Furthermore, K+ and glutamate buffering capabilities of D2 astrocytes are impaired relative to B6. Imbalances in K+ and glutamate extrusion to and clearance from the extracellular space have been associated with abnormal neuronal excitability and function (Zuckermann and Glaser, 1968; Dichter et al., 1972; Yaari et al., 1986; Traynelis and Dingledine, 1988; Janigro et al., 1997; Rothstein et al., 1996; Tanaka et al., 1997; Campbell and Hablitz, 2008;). Taken together, these findings suggest that lower Kir channel activity in D2 mice is a major contributor to the differences in seizure susceptibility. Genetic analysis reveals a single major variant in Kir4.1 between B6 and D2 strains of mice, namely the Thr262Ser amino acid variation.
There are two major ways that a missense variation could alter Kir channel activity. The first is by decreasing K+ conductance through the channel. Surprisingly, when these variants (Kir4.1 Thr262 and and Kir4.1 Ser262) were expressed in Xenopus oocytes (either as homomeric Kir4.1 channels or heteromeric Kir4.1/Kir5.1 channels) and compared, there was no difference in overall current in oocytes, response to intracellular acidification, inhibition by extracellular barium or single channel properties (Shang et al., 2005). Furthermore, we have found that there are no differences in the rectification properties of either the threonine or serine variant of Kir4.1 when transiently expressed in tsA201 cells (Eaton et al., 2009). Taken together, these data suggest that the Thr262Ser variation of Kir4.1 channels does not alter the overall biophysical properties and K+ conductance through the channel.
An alternative way to reduce Kir channel activity is by decreasing the overall cell surface expression of Kir4.1 channels in astrocytes. Reduction in Kir4.1 channels is consistent with what is seen in some epilepsies, such as Ammon’s horn sclerosis (Schröder et al., 2000; Hinterkeuser et al., 2000). Furthermore, reduction of Kir4.1 channels in astrocytic plasma membrane has been shown to reduce K+ and glutamate clearance by astrocytes (Kucheryavykh et al., 2007; Djukic et al., 2007).
Some of the polymorphisms associated with seizure susceptibility that occur in the KCNJ10 gene, in both humans and mice, cause alterations of amino acids predicted to be located in a cytoplasmic domain of the ion channel (Shang et al., 2005) and potentially involved in channel trafficking. Surface expression of mammalian Kir channels is tightly regulated by their cytoplasmic domains with specific amino acid sequences in both the N-terminal and C-terminal regions of Kir4.1 subunits controlling intracellular protein trafficking (Stockklausner and Klöcker, 2003). It has been reported that short amino acid sequences in the C-terminus control trafficking between the endoplasmic reticulum and the Golgi complex whereas the proximal N-terminus controls the ability of the protein to be transported out of the Golgi and to reach the cell membrane. The lack of appropriate sequence information in these regions results in accumulation of the protein intracellularly and a significant decrease in cell surface expression (Stockklausner and Klocker, 2003). Furthermore, interactions with co-expressed proteins have been shown to be involved in transport of Kir4.1 to the cell membrane as well as in the normal clustered pattern of channel distribution (Noël et al., 2005; Connors et al., 2004; Connors and Kofuji, 2002). Thus, it is possible that the common mouse (Thr272Ser) and human (Arg271Cys) amino acid substitutions in Kir4.1 influence seizure susceptibility by virtue of their ability to affect the kinetics of intracellular Kir4.1 transport and/or membrane targeting, but this will require further studies of subunit trafficking in native tissues.
In summary, the activity of Kir4.1 channels differs between seizure susceptible D2 and seizure resistant B6 mice. Reduced activity of Kir4.1 channels in astrocytes of D2 mice is associated with deficits in potassium and glutamate buffering. These deficits may, in part, explain the relatively low seizure threshold of D2 mice.
Acknowledgments
The authors thank Paola López Pieraldi and Natalia Skachkova for their superior technical assistance. This publication was made possible by NIH grant numbers S11-NS48201 (MJE), U54 NS039408 (SNS), S06 GM050695 (SNS, MJE), G12 RR03035 (for facilities at UCC) and R01 NS40554 (TNF) from NINDS, NIGMS and NCRR. Its contents are solely the responsibility of the authors and do not necessarily represent the official views of NIH. The Coatesville Veteran’s Affairs Medical Center animal facilities and resources were used to support some of this work. The contents do not reflect the official views of the Department of Veteran Affairs or any other US government agency. We confirm that we have read the Journal’s position on issues involved in ethical publication and affirm that this report is consistent with those guidelines.
Footnotes
Disclosure of Conflicts of Interest
None of the authors has any conflict of interest to disclose.
References
- Abe K, Abe Y, Saito H. Evaluation of L-glutamate clearance capacity of cultured rat cortical astrocytes. Biol Pharm Bull. 2000;23:204–207. doi: 10.1248/bpb.23.204. [DOI] [PubMed] [Google Scholar]
- Bockenhauer D, Feather S, Stanescu HC, Bandulik S, Zdebik AA, Reichold M, Tobin J, Lieberer E, Sterner C, Landoure G, Arora R, Sirimanna T, Thompson D, Cross JH, van’t Hoff W, Al Masri O, Tullus K, Yeung S, Anikster Y, Klootwijk E, Hubank M, Dillon MJ, Heitzmann D, Arcos-Burgos M, Knepper MA, Dobbie A, Gahl WA, Warth R, Sheridan E, Kleta R. Epilepsy, Ataxia, Sensorineural Deafness, Tubulopathy, and KCNJ10 Mutations. New Eng J Med. 2009;360:1960–1970. doi: 10.1056/NEJMoa0810276. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Broberg M, Pope KJ, Lewis T, Olsson T, Nilsson M, Willoughhby JO. Cell swelling precedes seizures induced by inhibition of astrocytic metabolism. Epilepsy Res. 2008;80:132–141. doi: 10.1016/j.eplepsyres.2008.03.012. [DOI] [PubMed] [Google Scholar]
- Buono RJ, Lohoff FW, Sander T, Sperling MR, O’Connor MJ, Dlugos DJ, Ryan SG, Golden GT, Zhao H, Scattergood TM, Berrettini WH, Ferraro TN. Association between variation in the human KCNJ10 potassium ion channel gene and seizure susceptibility. Epilepsy Res. 2004;58:175–183. doi: 10.1016/j.eplepsyres.2004.02.003. [DOI] [PubMed] [Google Scholar]
- Campbell SL, Hablitz JJ. Decreased glutamate transport enhances excitability in a rat model of cortical dysplasia. Neurobiol Dis. 2008;32:254–261. doi: 10.1016/j.nbd.2008.07.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Connors NC, Kofuji P. Dystrophin Dp71 is critical for the clustered localization of potassium channels in retinal glial cells. J Neurosci. 2002;22:4321–4327. doi: 10.1523/JNEUROSCI.22-11-04321.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Connors NC, Adams ME, Froehner SC, Kofuji P. The potassium channel Kir4.1 associates with the dystrophin-glycoprotein complex via alpha-syntrophin in glia. J Biol Chem. 2004;279:28387–28392. doi: 10.1074/jbc.M402604200. [DOI] [PubMed] [Google Scholar]
- D’Ambrosio R, Wenzel J, Schwartzkroin PA, McKhann GM, 2nd, Janigro D. Functional specialization and topographic segregation of hippocampal astrocytes. J Neurosci. 1998;18:4425–4438. doi: 10.1523/JNEUROSCI.18-12-04425.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dichter MA, Herman CJ, Selzer M. Silent cells during interictal discharges and seizures in hippocampal penicillin foci. Evidence for the role of extracellular K+ in the transition from the interictal state to seizures. Brain Res. 1972;48:173–183. doi: 10.1016/0006-8993(72)90177-1. [DOI] [PubMed] [Google Scholar]
- Djukic B, Casper KB, Philpot BD, Chin LS, McCarthy KD. Conditional knock-out of Kir4.1 leads to glial membrane depolarization, inhibition of potassium and glutamate uptake, and enhanced short-term synaptic potentiation. J Neurosci. 2007;27:11354–11365. doi: 10.1523/JNEUROSCI.0723-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eaton MJ, Kucheryavykh YV, Kucheryavykh LY, Inyushin M, Kurata H, Nichols CG, Buono RJ, Ferraro TN, Maldonado HM, Skatchkov SN. Neuroscience Meeting Planner. Chicago, IL: Society for Neuroscience; 2009. Astrocytic Kir4.1 channel variants and seizure susceptibility. Online. [Google Scholar]
- Engstrom FL, Woodbury DM. Seizure susceptibility in DBA and C57 mice: the effects of various convulsants. Epilepsia. 1988;29:389–395. doi: 10.1111/j.1528-1157.1988.tb03736.x. [DOI] [PubMed] [Google Scholar]
- Ferraro TN, Golden GT, Dahl JP, Smith GG, Schwebel CL, MacDonald R, Lohoff FW, Berrettini WH, Buono RJ. Analysis of a quantitative trait locus for seizure susceptibility in mice using bacterial artificial chromosome-mediated gene transfer. Epilepsia. 2007;48:1667–1677. doi: 10.1111/j.1528-1167.2007.01126.x. [DOI] [PubMed] [Google Scholar]
- Ferraro TN, Golden GT, Smith GG, Martin JF, Lohoff FW, Gieringer TA, Zamboni D, Schwebel CL, Press DM, Kratzer SO, Zhao H, Berrettini WH, Buono RJ. Fine mapping of a seizure susceptibility locus on mouse Chromosome 1: nomination of Kcnj10 as a causative gene. Mamm Genome. 2004;15:239–251. doi: 10.1007/s00335-003-2270-3. [DOI] [PubMed] [Google Scholar]
- Ferraro TN, Golden GT, Smith GG, Longman RL, Snyder RL, DeMuth D, Szpitzak I, Mulholland N, Eng E, Lohoff FW, Buono RJ, Berrettini WH. Quantitative genetic study of maximal electroshock seizure threshold in mice: evidence for a major seizure susceptibility locus on distal chromosome 1. Genomics. 2001;75:35–42. doi: 10.1006/geno.2001.6577. [DOI] [PubMed] [Google Scholar]
- Ferraro TN, Golden GT, Snyder R, Laibinis M, Smith GG, Buono RJ, Berrettini WH. Genetic influences on electrical seizure threshold. Brain Res. 1998;813:207–10. doi: 10.1016/s0006-8993(98)01013-0. [DOI] [PubMed] [Google Scholar]
- Higashi K, Fujita A, Inanobe A, Tanemoto M, Doi K, Kubo T, Kurachi Y. An inwardly rectifying K+ channel, Kir4.1, expressed in astrocytes surrounds synapses and blood vessels in brain. Am J Physiol Cell Physiol. 2001;281:C922–C931. doi: 10.1152/ajpcell.2001.281.3.C922. [DOI] [PubMed] [Google Scholar]
- Hinterkeuser S, Schroder W, Hager JG, Seifert G, Blumcke I, Elger CE, Schramm J, Steinhäuser C. Astrocytes in the hippocampus of patients with temporal lobe epilepsy display changes in potassium conductances. Eur J Neurosci. 2000;12:2087–2096. doi: 10.1046/j.1460-9568.2000.00104.x. [DOI] [PubMed] [Google Scholar]
- Janigro D, Gasparini S, D’Ambrosio R, McKhann GM, DiFrancesco D. Reduction of K+ uptake in glia prevents LTD maintenance and causes epileptiform activity. J Neurosci. 1997;17:2813–2824. doi: 10.1523/JNEUROSCI.17-08-02813.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kofuji P, Ceelen P, Zahs KR, Surbeck LW, Lester HA, Newman EA. Genetic inactivation of an inwardly rectifying potassium channel (Kir4.1 subunit) in mice:phenotypic impact in retina. J Neurosci. 2000;20:5733–5740. doi: 10.1523/JNEUROSCI.20-15-05733.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kucheryavykh LY, Kucheryavykh YV, Inyushin M, Shuba YM, Sanabria P, Cubano LA, Skatchkov SN, Eaton MJ. Ischemia increases TREK-2 channel expression in astrocytes: Relevance to glutamate clearance. Open Neurosci J. 2009 doi: 10.2174/1874082000903010040. Published online: 23-Jun-2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kucheryavykh YV, Kucheryavykh LY, Nichols CG, Maldonado HM, Baksi K, Reichenbach A, Skatchkov SN, Eaton MJ. Downregulation of Kir4.1 Inward Rectifying Potassium Channel Subunits by RNAi Impairs Potassium Transfer and Glutamate Uptake by Cortical Astrocytes. Glia. 2007;55:274–281. doi: 10.1002/glia.20455. [DOI] [PubMed] [Google Scholar]
- Lenzen KP, Heils A, Lorenz S, Hempelmann A, Höfels S, Lohoff FW, Schmitz B, Sander T. Supportive evidence for an allelic association of the human KCNJ10 potassium channel gene with idiopathic generalized epilepsy. Epilepsy Res. 2005;63:113–118. doi: 10.1016/j.eplepsyres.2005.01.002. [DOI] [PubMed] [Google Scholar]
- Li L, Head V, Timpe LC. Identification of an inward rectifier potassium channel gene expressed in mouse cortical astrocytes. Glia. 2001;33:57–71. doi: 10.1002/1098-1136(20010101)33:1<57::aid-glia1006>3.0.co;2-0. [DOI] [PubMed] [Google Scholar]
- Neusch C, Papadopoulos N, Müller, Maletzki I, Winter SM, Hirrlinger J, Handschuh M, Bähr M, Richter DW, Kirchhoff F, Hülsmann S. Lack of the Kir4.1 channel subunit abolishes K+ buffering properties of astrocytes in the ventral respiratory group: impact on extracellular K+ regulation. J Neurophysiol. 2006;95:1843–1852. doi: 10.1152/jn.00996.2005. [DOI] [PubMed] [Google Scholar]
- Noël G, Belda M, Guadagno E, Micoud J, Klöcker N, Moukhles H. Dystroglycan and Kir4.1 coclustering in retinal Müller glia is regulated by laminin-1 and requires the PDZ-ligand domain of Kir4.1. J Neurochem. 2005;94:691–702. doi: 10.1111/j.1471-4159.2005.03191.x. [DOI] [PubMed] [Google Scholar]
- Olsen ML, Campbell SL, Sontheimer H. Differential distribution of Kir4.1 in spinal cord astrocytes suggest regional differences in K+ homeostasis. J Neurophysiol. 2007;98:786–793. doi: 10.1152/jn.00340.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Poopalasundaram S, Knott C, Shamotienko OG, Foran PG, Dolly JO, Ghiani CA, Gallo V, Wilkin GP. Glial heterogeneity in expression of the inwardly rectifying K(+) channel, Kir4.1, in adult rat CNS. Glia. 2000;30:362–372. doi: 10.1002/(sici)1098-1136(200006)30:4<362::aid-glia50>3.0.co;2-4. [DOI] [PubMed] [Google Scholar]
- Rothstein JD, Dykes-Hoberg M, Pardo CA, Bristol LA, Jin L, Kuncl RW, Kanai Y, Hediger MA, Wang YF, Schielke JP, Welty DF. Knockout of glutamate transporters reveals a major role for astroglial transport in excitotoxicity and clearance of glutamate. Neuron. 1996;16:675–686. doi: 10.1016/s0896-6273(00)80086-0. [DOI] [PubMed] [Google Scholar]
- Schröder W, Hinterkeuser S, Seifert G, Schramm J, Jabs R, Wilkin GP, Steinhäuser C. Functional and molecular properties of human astrocytes in acute hippocampal slices obtained from patients with temporal lobe epilepsy. Epilepsia. 2000;41(Suppl 6):S181–S184. doi: 10.1111/j.1528-1157.2000.tb01578.x. [DOI] [PubMed] [Google Scholar]
- Schröder W, Seifer G, Huttmann K, Hinterkeuser S, Steinhäuser C. AMPA receptor-mediated modulation of inward rectifier K+ channels in astrocytes of mouse hippocampus. Mol Cell Neurosci. 2002;19:447–458. doi: 10.1006/mcne.2001.1080. [DOI] [PubMed] [Google Scholar]
- Scholl UI, Choi M, Liu T, Ramaekers VT, Häusler MG, Grimmer J, Tobe SW, Farhi A, Nelson-Williams C, Lifton RP. Seizures, sensorineural deafness, ataxia, mental retardation, and electrolyte imbalance (SeSAME syndrome) caused by mutations in KCNJ10. PNAS. 2009;16:5842–5847. doi: 10.1073/pnas.0901749106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schopf S, Ruge H, Bringmann A, Reichenbach A, Skatchkov SN. Switch of K+ buffering conditions in Müller glial cells of rabbit retina during postnatal development. Neurosci Lett. 2004;365:167–170. doi: 10.1016/j.neulet.2004.04.074. [DOI] [PubMed] [Google Scholar]
- Seifert G, Küttman K, Binder DK, Harmann C, Wyczynski A, Neusch C, Steinhä0user C. Analysis of astroglial K+ channel expression in the developing hippocampus reveals a predominant role of the Kir4.1 subunit. J Neurosci. 2009;29:7474–7488. doi: 10.1523/JNEUROSCI.3790-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shang L, Lucchese CJ, Haider S, Tucker SJ. Functional characterisation of missense variations in the Kir4.1 potassium channel (KCNJ10) associated with seizure susceptibility. Brain Res Mol Brain Res. 2005;139:178–183. doi: 10.1016/j.molbrainres.2005.05.003. [DOI] [PubMed] [Google Scholar]
- Skatchkov SN, Krusek J, Reichenbach A, Orkand RK. Potassium buffering by Müller cells isolated from the center and periphery of the frog retina. Glia. 1999;27:171–180. [PubMed] [Google Scholar]
- Stockklausner C, Klöcker N. Surface expression of inward rectifier potassium channels is controlled by selective Golgi export. J Biol Chem. 2003;278:17000–17005. doi: 10.1074/jbc.M212243200. [DOI] [PubMed] [Google Scholar]
- Takumi T, Ishii T, Horio Y, Morishige K, Takahashi N, Yamada M, Yamashita T, Kiyama H, Sohmiya K, Nakanishi S, Kurachi Y. A novel ATP-dependent inward rectifier potassium channel expressed predominantly in glial cells. J, Biol, Chem. 1995;270:16339–16346. doi: 10.1074/jbc.270.27.16339. [DOI] [PubMed] [Google Scholar]
- Tanaka K, Watase K, Manabe T, Yamada K, Watanabe M, Takahashi K, Iwama H, Nishikawa T, Ichihara N, Hori S, Takimoto M, Wada K. Epilepsy and exacerbation of brain injury in mice lacking the glutamate transporter GLT1. Science. 1997;276:1699–1702. doi: 10.1126/science.276.5319.1699. [DOI] [PubMed] [Google Scholar]
- Tian G-F, Azmi H, Takano T, Xu Q, Peng W, Lin J, Oberheim N, Lou N, Want X, Zielke HR, Kang J, Nedergaard M. An astrocytic basis of epilepsy. Nature Med. 2005;11:973–981. doi: 10.1038/nm1277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Traynelis SF, Dingledine R. Potassium-induced spontaneous electrographic seizures in the rat hippocampal slice. J Neurophysiol. 1988;59:259–276. doi: 10.1152/jn.1988.59.1.259. [DOI] [PubMed] [Google Scholar]
- Yaari Y, Konnerth A, Heinemann U. Nonsynaptic epileptogenesis in the mammalian hippocampus in vitro. II. Role of extracellular potassium. J Neurophysiol. 1986;56:424–438. doi: 10.1152/jn.1986.56.2.424. [DOI] [PubMed] [Google Scholar]
- Zhou M, Kimelberg HK. Freshly isolated astrocytes from rat hippocampus show two distinct current patterns and different [K+]o uptake capabilities. J Neurophysiol. 2000;84:2746–2757. doi: 10.1152/jn.2000.84.6.2746. [DOI] [PubMed] [Google Scholar]
- Zuckermann EC, Glaser GH. Hippocampal epileptic activity induced by localized ventricular perfusion with high-potassium cerebrospinal fluid. Exp Neurol. 1968;20:87–110. doi: 10.1016/0014-4886(68)90126-x. [DOI] [PubMed] [Google Scholar]