

Abstract
As markers of DNA synthesis, [3H]thymidine ([3H]dT) and the later developed analog bromodeoxyuridine (BrdU) have revolutionized our ability to identify dividing cells and follow their fate in various tissues, including the nervous system. However, the effect of the incorporation of these molecules into DNA on cell proliferation, migration, differentiation, and function is not fully understood. Here, we compare the number and distribution of labeled cells in the cerebral cortex of postnatal macaque monkeys exposed to either [3H]dT or BrdU as embryos. The large size and prolonged brain development in this species allows higher resolution of cellular events and more accurate discrimination between the two methods. Our analysis revealed substantial differences in the number and distribution of labeled cells. The data indicate that random incorporation of the thymidine analog BrdU into the genes of dividing cells makes the fate of postmitotic neurons more prone to unpredictable errors than the incorporation of the more natural DNA constituent nucleotide [3H]dT. These findings have implications for the interpretation of results obtained by BrdU as an index of the number of neurons produced, and their migration, placement, subsequent connectivity, function, and survival.
Introduction
For decades, [3H]thymidine ([3H]dT) autoradiography dominated the studies of cell proliferation, birth dating, migration, and fate in the developing brain (Sidman et al., 1959; Altman, 1963, 1969; Angevine, 1965; Rakic and Sidman, 1968; Rakic, 1974, 2002b; Schlessinger et al., 1975; Nowakowski and Rakic, 1979). However, because of cost, logistic problems in handling radiolabeled substances, and the lengthy process of developing autoradiographs (3–12 weeks), contemporary studies are commonly performed with the thymidine analog bromodeoxyuridine (5-bromo-2′-deoxyuridine) (BrdU), a halopyrimidine with a different molecular structure than [3H]dT.
[3H]dT and BrdU incorporate into the nuclear DNA during the S-phase of the cell cycle (Nowakowski et al., 1989), and both are known to be toxic (Ehmann et al., 1975; Kolb et al., 1999; Nowakowski and Hayes, 2000; Sekerková et al., 2004; Breunig et al., 2007; Kuwagata et al., 2007) (for review, see Taupin, 2007). However, since DNA synthesis can be initiated independently of mitosis (e.g., during gene duplication, repair, or apoptosis), both markers are indicators of only DNA synthesis and not of cell division (Yang et al., 2001; Rakic, 2002c; Kuan et al., 2004) (for review, see Breunig et al., 2007) as commonly assumed. Nevertheless, [3H]dT and BrdU are generally used as specific markers of neuronal birth during embryonic, postnatal, and adult neurogenesis, mostly without taking into consideration their possible effects on cell division and subsequent cell function and fate.
Use of each DNA marker has advantages and disadvantages. For example, because the half-distance of the beta particle emitted by tritium atom decay is only ∼1 μm, detection of [3H]dT-labeled cells is restricted to a few micrometers deep from the surface of the section (Sidman, 1970; Rogers, 1973, Bisconte, 1979). However, the intensity of [3H]dT labeling is stoichiometric (Nowakowski and Rakic, 1974; Rakic, 2002a,b) and can be combined with anatomical tracing methods (Nowakowski et al., 1975). In contrast, BrdU has the advantage that its presence can rapidly be revealed by immunohistochemical methods (Gratzner, 1982), which also allow colabeling with various phenotypic markers. Furthermore, although BrdU labeling is not stoichiometric (Nowakowski and Hayes, 2000), immunohistochemistry in 50-μm-thick tissue efficiently labels all cells throughout the section.
Previous studies in rodent CNS indicated that results obtained with [3H]dT and BrdU are similar (Miller and Nowakowski, 1988; del Rio and Soriano, 1989). This study was done in primate because its large brain and slow development provide higher spatial and temporal resolution to study the effects of these markers on cellular events. Although data of the time at which labeling of CNS cells takes place in this species, obtained by [3H]dT and BrdU, are in basic qualitative agreement (Rakic, 2002c), it is important to examine whether the results obtained by these two markers are also quantitatively the same. To make sure that our results are a function of the different effect of these molecules on DNA translation rather than a reflection of differences in injection times, we examined monkeys injected with either [3H]dT or BrdU at two postconception times. The necessary corrections of inherent methodological disparities in handling the tissue and other technical biases were also performed.
Materials and Methods
Animals.
Animal care and experimentation were conducted in accordance with institutional guidelines. Animal breeding and dating of pregnancies have previously been described (Rakic, 1973; Kornack and Rakic, 1998). Fourteen rhesus monkeys (Macaca mulatta) were used in this study. Seven monkeys were injected with [3H]dT and seven with BrdU at various embryonic days. All animals delivered at term (∼E165) and were killed ∼6 months postinjection, at postnatal days P48 to P98 (Table 1). The distribution of labeled cells in the cerebral cortex was examined in every case. In addition, in the animals in which the days of injection matched precisely (Table 1), quantitative comparisons between the two markers were performed in five cytoarchitectonically and functionally different cortical areas (Tables 2, 3).
Table 1.
Animals
| Animals | ||||
|---|---|---|---|---|
| BrdU | Sex | Comparison | [3H]dT | Sex |
| E55–P75 | F |  |
E40–P62 | U |
| E55–P75 | F | E50–P61 | U | |
| E56–P66 | U | |||
| E70–P70 | F |  |
||
| E70–P75 | M | E62–P50 | U | |
| E70–P75 | M | E70–P98 | U | |
| E70–P75 | F | E80–P48 | U | |
| E70–P91 | F | E85–P94 | U | |
Qualitative assessment (n = 14) of the position of labeled cells was performed in all cases. Quantitative comparisons (n = 9) were made only for cases in which the injection time closely matched for both markers. F, Female; M, male; U, unknown; E, embryonic age at time of injection (days); P, postnatal day at time of killing (days).
Table 2.
Details on labeled cell numbers and percentage comparisons
| Labeled cell numbers and percentages |
|||||
|---|---|---|---|---|---|
| PFC | LIM | MOT | SOM | VIS | |
| Injections at E70 | |||||
| Number of labeled cells | |||||
| BrdU | |||||
| Total | 1136 | 928 | 995 | 1206 | 2307 |
| % Dead | 59.9 | 73.0 | 75.1 | 68.3 | 16.3 |
| [3H]dTa | |||||
| Total-1 | 11,320 | 13,740 | 16,000 | 15,220 | 11,020 |
| Total-2 | 5660 | 6870 | 8000 | 7610 | 5510 |
| Total-3 | 2830 | 3435 | 4000 | 3805 | 2755 |
| % Dead | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Percentage largest accumulation of labeled cells within a single 100 μm bin | |||||
| BrdU | |||||
| % | 12.4 | 17.4 | 12.7 | 16.2 | 13.4 |
| Depth (μm) | 250 | 250 | 250 | 250 | 1050 |
| [3H]dT | |||||
| % | 21.0 | 20.2 | 14.5 | 16.2 | 36.8 |
| Depth (μm) | 650 | 350 | 450 | 550 | 850 |
| Percentage largest accumulation of labeled cells within a single 100 μm bin, corrected for cortical thicknessb | |||||
| BrdU | |||||
| % | 12.2 | 13.2 | 12.5 | 15.7 | 15.7 |
| Depth (μm) | 250 | 250 | 250 | 250 | 1150 |
| [3H]dT | |||||
| % | 16.6 | 12.7 | 10.0 | 12.8 | 28.9 |
| Depth (μm) | 950 | 450 | 650 | 350 | 1250 |
| Percentage accumulation of labeled cells within 5 and 10 (100 μm) bins | |||||
| BrdU | |||||
| 5 bins | 48.2 | 44.1 | 45.8 | 49.6 | 89.3 |
| 10 bins | 79.3 | 61.7 | 70.3 | 72.8 | 99.6 |
| [3H]dT | |||||
| 5 bins | 77.4 | 81.4 | 64.8 | 70.4 | 54.6 |
| 10 bins | 99.8 | 96.9 | 96.1 | 95.5 | 85.0 |
| Percentage accumulation of labeled cells within 5 and 10 (100 μm) bins, corrected for cortical thicknessb | |||||
| BrdU | |||||
| 5 bins | 47.3 | 49.0 | 45.9 | 51.2 | 57.1 |
| 10 bins | 79.4 | 64.7 | 69.6 | 73.1 | 85.6 |
| [3H]dT | |||||
| 5 bins | 67.1 | 56.8 | 43.4 | 55.8 | 86.2 |
| 10 bins | 97.9 | 87.2 | 73.1 | 90.4 | 95.3 |
| Percentage accumulation of labeled cells deeper than 1900 μm from pia (deep bins) | |||||
| BrdU | |||||
| % | 4.8 | 21.8 | 14.7 | 15.8 | 0.2 |
| %b | 4.7 | 22.0 | 14.1 | 15.6 | 0.2 |
| [3H]dT | |||||
| % | 0.0 | 0.0 | 0.4 | 1.1 | 0.0 |
| %b | 0.0 | 0.0 | 8.1 | 0.9 | 0.0 |
| Injections at E55 | |||||
| Number of labeled cells | |||||
| BrdU | |||||
| Total | 779 | 1105 | 1219 | 994 | 1593 |
| Mean | 194.8 | 276.3 | 304.8 | 248.5 | 398.3 |
| % Dead | 75.9 | 49.5 | 57.3 | 21.1 | 45.9 |
| [3H]dTa | |||||
| Total | 2420 | 1640 | 2140 | 945 | 2210 |
| Mean | 806.7 | 546.7 | 713.3 | 315.0 | 736.7 |
| Percentage largest accumulation of labeled cells within a single 100 μm bin, corrected for cortical thicknessb | |||||
| BrdU | |||||
| % | 13.2 | 9.1 | 5.3 | 11.3 | 12.8 |
| Depth (μm) | 1050 | 1050 | 1850 | 1150 | 1350 |
| [3H]dT | |||||
| % | 20.7 | 22.2 | 9.8 | 22.3 | 39.4 |
| Depth (μm) | 750 | 950 | 1450 | 750 | 1150 |
| Percentage accumulation of labeled cells deeper than 1900 μm from pia (deep bins) | |||||
| BrdU | |||||
| %b | 4.2 | 2.9 | 35.5 | 10.1 | 8.5 |
| [3H]dT | |||||
| %b | 0.0 | 0.0 | 24.1 | 0.0 | 0.0 |
Percentage accumulation within 5 and 10 bins is not shown for injections at E55; they follow the same trends of those shown for injections at E70.
aCorrected by increasing [3H]dT cell numbers as follows: total-1 detected cell numbers multiplied 20× represents the case in which detection occurred on the top 2.5 μm of the tissue; total-2 (10×) and total-3 (5×) represent cases in which detection occurred within 5 and 10 μm from the top, correspondingly. For [3H]dT, we assumed survival of 100% until detection; for details, see Results, and Materials and Methods, Technical considerations and corrections of methodological differences.
bAfter normalizing cortical thickness per area against that in BrdU material.
Table 3.
Statistical details
| PFC | LIM | MT | SS | VIS | ||
|---|---|---|---|---|---|---|
| Statistical results for injections at E70 | ||||||
| Number of plotted cells | ||||||
| BrdU | ||||||
| Total | 1136 | 928 | 995 | 1206 | 2307 | |
| Mean | 227.2 | 185.6 | 199.0 | 241.2 | 461.4 | |
| SEM | 13.0 | 16.1 | 12.0 | 12.8 | 15.0 | |
| PFC | — | 0.0318 | 0.1574 | 0.3392 | 0.0015 | Multiple comparisons (p values–LSD contrast) |
| LIM | — | — | 0.4112 | 0.0037 | <0.0001 | |
| MT | — | — | — | 0.0237 | <0.0002 | |
| SS | — | — | — | — | 0.0140 | |
| [3H]dT | ||||||
| Total | 566 | 687 | 800 | 761 | 551 | |
| Mean | 113.2 | 137.4 | 160.0 | 152.2 | 110.2 | |
| SEM | 10.4 | 16.5 | 26.8 | 9.0 | 29.9 | |
| BrdU–[3H]dT difference in number of plotted cells | ||||||
| p | 0.0001 | 0.0719 | 0.2215 | 0.0005 | <0.0001 | ANOVA, single factor (t test) |
| BrdU–[3H]dT difference in cell numbers, corrected for section cut thickness and [3H]dT detection abilitya | ||||||
| p | 0.0002 | 0.0003 | 0.0021 | <0.0001 | 0.5679 | |
| Comparison of the percentage distributions of BrdU versus [3H]dT | ||||||
| χ2 | 7.74 | 10.52 | 6.15 | 8.16 | 3.61 | Kruskal–Wallis test |
| p | 0.0054 | 0.0012 | 0.0132 | 0.0043 | 0.0575 | |
| F (p) | 0.35 (0.0066) | 0.26 (0.0006) | 0.38 (0.0111) | 0.43 (0.0265) | 0.27 (0.0007) | F test of dispersion |
| Correlation coefficient of percentage of BrdU versus [3H]dT | ||||||
| rs, p | 0.83, p < 0.0001 | 0.77, p < 0.0001 | 0.82, p < 0.0001 | 0.79, p < 0.0001 | 0.69, p < 0.0001 | Spearman's rank |
| Comparison of the percentage distributions of BrdU versus [3H]dT, corrected for cortical thicknessb | ||||||
| χ2 | 4.08 | 6.11 | 1.4 | 3.58 | 1.22 | Kruskal–Wallis test |
| p | 0.0434 | 0.0135 | 0.2365 | 0.0583 | 0.2691 | |
| F (p) | 2.04 (0.0594) | 2.28 (0.0298) | 1.50 (0.2839) | 1.33 (0.4528) | 3.02 (0.0040) | F test of dispersion |
| Correlation coefficient for mean percentage distributions of BrdU versus [3H]dTb | ||||||
| rs, p | 0.89, p < 0.0001 | 0.68, p < 0.0001 | 0.78, p < 0.0001 | 0.65, p < 0.0001 | 0.88, p < 0.0001 | Spearman's rank |
| Statistical results for injections at E55 | ||||||
| Number of plotted cells | ||||||
| BrdU | ||||||
| Total | 779 | 1105 | 1219 | 994 | 1593 | |
| Mean | 194.8 | 276.3 | 304.8 | 248.5 | 398.3 | |
| SEM | 34.2 | 45.9 | 23.3 | 34.9 | 46.3 | |
| [3H]dT | ||||||
| Total | 484 | 328 | 428 | 189 | 442 | |
| Mean | 161.3 | 109.3 | 142.7 | 63.0 | 147.3 | |
| SEM | 14.2 | 9.9 | 22.3 | 12.2 | 11.8 | |
| Comparison of the percentage distributions of BrdU versus [3H]dT | ||||||
| χ2 | 2.37 | 2.75 | 0.35 | 5.17 | 8.19 | Kruskal–Wallis test |
| p | 0.1238 | 0.0974 | 0.5541 | 0.0230 | 0.0042 | |
| F (p) | 0.42 (0.0234) | 0.28 (0.0009) | 0.20 (<0.0001) | 0.34 (0.0047) | 0.13 (0.0001) | F test of dispersion |
| Correlation coefficient of the percentage of BrdU versus [3H]dT | ||||||
| rs, p | 0.67, p < 0.0001 | 0.90, p < 0.0001 | 0.86, p < 0.0001 | 0.60, p = 0.0005 | 0.61, p = 0.0004 | Spearman's rank |
| Correlation coefficient of the percentage of BrdU versus [3H]dTb | ||||||
| rs, p | 0.89, p < 0.0001 | 0.59, p = 0.0005 | 0.79, p < 0.0001 | 0.81, p < 0.0001 | 0.88, p < 0.0001 | Spearman's rank |
Only essential details are shown for each group, and not all details are shown for E55, which follows the same trends observed for E70.
aCorrected by increasing [3H]dT cell numbers by 500%. This corresponds to the case for which we had the worse possible comparisons (Table 2, Total-3, for the number of [3H]dT cells; i.e., the case in which we assume [3H]dT cells are detected within 10 μm). For the other two cases, the estimated numbers of [3H]dT cells are so high compared with the number of BrdU cells that all per corresponding area comparisons were always statistically significant.
bAfter normalizing cortical thickness per area against that in BrdU material.
Values in bold are statistically significant.
Injections.
All animals received a single intravenous injection of one of two DNA replication markers. [3H]dT (PerkinElmer Life and Analytical Sciences; 10 mCi per kilogram of body weight) and BrdU (Sigma-Aldrich; 50 mg per kilogram of body weight; dissolved in 0.9% NaCl with 0.007 m NaOH) injections in nonsedated pregnant monkeys took place while the monkey was sitting in a special chair. Because results of BrdU labeling may change with different antibodies and staining procedures (Leuner et al., 2009), our BrdU protocols were identical in all cases, and BrdU detection was done using the same antibody, mouse monoclonal anti-BrdU (clone BU 33; catalog #B2531; Sigma-Aldrich).
Perfusions, immunohistochemistry, and tissue processing.
Methods for perfusions and processing of monkey brains for autoradiography and BrdU have been previously published (Rakic and Sidman, 1970; Rakic, 1973, 1976; Kornack and Rakic, 1998). In all BrdU cases, sections were cut at 50 μm. In [3H]dT cases, sections were cut at 8–10 or 30 μm. All brains were cut in a freezing microtome in the coronal plane. In every case, every 10th section was stained for Nissl and an adjacent section was treated to reveal [3H]dT or BrdU. BrdU sections were counterstained with thionine and [3H]dT sections with aqueous toluidine blue. Having been processed >30 years ago, the mounting media on [3H]dT sections had deteriorated. After immersion in xylenes, old coverslips were removed, and new mounting media and coverslips were used to recover the slides. The quality of the [3H]dT autoradiography was not compromised (see Fig. 1).
Figure 1.

[3H]dT versus BrdU staining. Examples of [3H]dT (A) and BrdU (B) staining in the visual cortex are shown.
Microscopy and sampling.
Section outlines and cell plotting were done using a Zeiss Axioskop microscope fully motorized and interfacing to a Dell computer running StereoInvestigator software (MicroBrightField). Outlines and fiduciary marks were drawn at 5×. Boxes were placed using bright field (10×), and were sequentially scanned at 100× oil-immersion using a Zeiss Plan-Neofluar objective lens with a 1.3 optical aperture. Once a labeled cell was encountered, its position was plotted using one of the symbols provided by the program. Data were harvested using Neurolucida Explorer. To plot cells, a “counting” rectangular box (500 × 3000 μm) was placed in the cortical area to be sampled, the short side aligned to the pia border while the long side extended toward the white matter (WM). A 200-μm-long extension of the rectangle beyond pia served as a guide for positioning. Boxes were located in five different cortical areas: prefrontal cortex (PFC), in the ventral bank of the principal sulcus in area 46V; limbic cortex (LIM), in the cingulate gyrus, area 24a/b; motor cortex (MOT) in area 4 dorsal to the superior precentral dimple; somatosensory cortex (SOM), in area 1–2 in the laterodorsal surface of the post-central gyrus; and in the visual cortex (VIS) in the ventral back of the calcarine sulcus (area 17) (Fig. 2).
Figure 2.
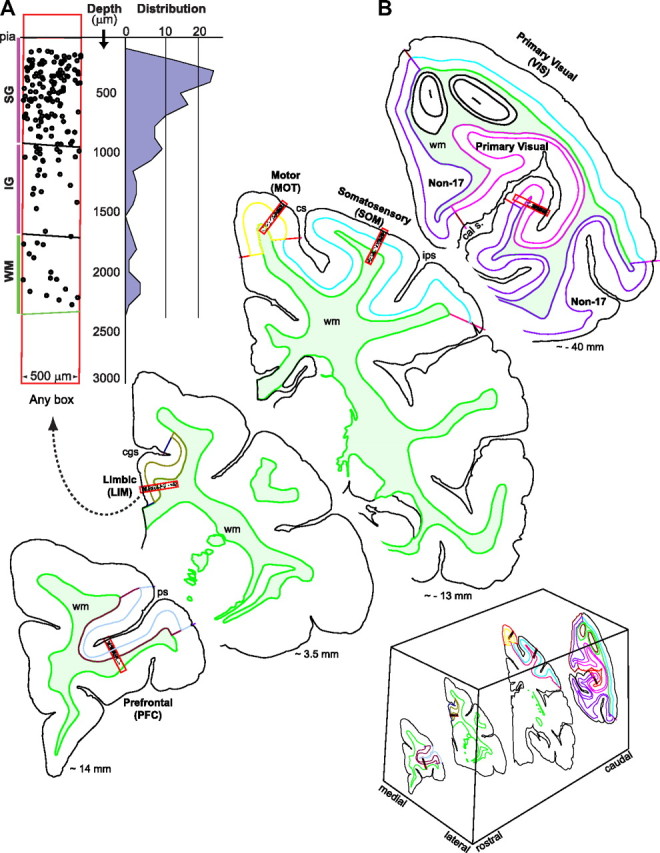
Sampling strategy. A, B, Hemicoronal sections illustrating approximate rostrocaudal levels and cortical sites where sampling boxes were placed. A random box is illustrated to indicate how cell distributions from pia to the white matter (WM, wm; filled light green color) are generated. The black dots inside boxes represent labeled cells. cal s., Calcarine sulcus; cgs, cingulate sulcus; cs, central sulcus; ips, intraparietal sulcus; ps, principal sulcus; SG, supragranular, IG, infragranular. Values in millimeters indicate approximate distance to bregma according to the atlas of Paxinos et al. (2000). The abbreviations for the five cortical areas studied apply to all other figures and tables.
In the BrdU cases at E70, we obtained 1 box of data per cortical area per brain, 25 boxes in total. From the single [3H]dT case at E70 (Table 1), we also obtained a total of 25 boxes of data, 5 boxes from every cortical area. In the BrdU cases at E55, we obtained 4 boxes of data per cortical area, for a total of 20 boxes, 3 boxes per cortical area in one brain and 1 box per cortical area in the second brain. In the corresponding case for [3H]dT, we obtained 3 boxes per cortical area for a total of 15 boxes. The rostrocaudal position of the selected sections was closely matched between brains and the corresponding boxes were placed using the same orientation and at approximately the same location with respect to anatomical landmarks [i.e., boxes were always placed at matching positions between the different sections to sample the corresponding cortical area (Fig. 2)].
We investigated cell numbers and positions of cells with respect to the pia and we compared the results for corresponding cortical areas between BrdU and [3H]dT before and after applying correction factors to account for cortical thickness differences and tissue shrinkage. We collapsed the plotted cells to the pia–WM plane and calculated the distribution of labeled cells from pia to the WM (inclusive) that resulted from both BrdU and [3H]dT labeling. To plot the distribution of cells, we measured the straight distance from each plotted cell to pia and then calculated the mean number of cells encountered at each depth in bins of 100 μm size. Because of differences in the total number of labeled cells both within the same region using the same label and between regions using different labels, we normalized the data by calculating the binned mean percentage of cells with respect to depth from pia and plotted the histogram distribution of mean percentages (per bin) of both populations. The relationships between the corresponding distributions, per cortical area, were investigated using nonparametric tests (see below, Statistical analyses).
Criteria for positive labeling.
In both [3H]dT and BrdU material, the staining consists of a gradient in amount and intensity of the corresponding precipitate (silver grains in [3H]dT and DAB in BrdU cases). The nucleus of a [3H]dT cell was considered positive when it contained a minimum of three discrete grains that would clearly outnumber any grains over similar surface areas in the background (Rakic, 1973). Regions of high background where many silver grains could be observed outside of cellular profiles were not used for counting; and a new cleaner area, usually in a different section, was selected. Similarly, nuclei of cells were considered to be BrdU positive if the amount and intensity of their labeling far exceeded that of their background. In this manner, for both [3H]dT and BrdU, the whole gradient of positive cells, from lightly to heavily labeled, was plotted. We differentiated only between positively stained and unstained cells and not between heavily and lightly stained cells. This strategy assures that there is no bias in the sampling and therefore allows comparison of labeled cell numbers and their distribution.
Statistical analyses.
For quantitative comparisons and detailed statistical analyses between BrdU and [3H]dT, we examined animals injected at E70 and E55 (Table 1). Statistical analyses were performed with Analyze-it, version 2.12 for Microsoft Excel. Univariate statistics [means ± SEM, frequencies (percentage)] were performed for all study variables. The distributions of different data were examined to check for normalcy and homoscedasticity to confirm the appropriateness of the statistical procedures. Normality was assessed using the Shapiro–Wilk test. Homogeneity of variance was tested with an F test. ANOVA was used to compare means among normally distributed populations with equal variance. Nonparametric ANOVA alternatives were performed with the Mann–Whitney and/or Kruskal–Wallis tests. Agreement between distributions was evaluated using Bland–Altman analysis (Altman and Bland, 1983; Bland and Altman, 1986). Further analyses included the computation of χ2 values and Spearman correlation coefficients. When appropriate, least square difference (LSD), Tukey's, or Bonferroni's corrections were applied.
Technical considerations and corrections of methodological differences.
To compare the actual number of cells that the plotted cells represent, it is necessary to make several corrections relevant to the methodological differences between the two approaches. First, BrdU sections were cut 50 μm thick, while [3H]dT sections were 8–10 μm (E70) and 30 μm (E55) thick, respectively. Because immunocytochemistry exposes BrdU-positive cells throughout the thickness of the section, we consider the number of BrdU plotted cells to represents 100% of the BrdU-labeled cells that are present in the examined tissue. Although previous studies have shown that the number of labeled cells observed following exposure to BrdU is sensitive to multiple parameters (Hayes and Nowakowski, 2002), BrdU immunohistochemistry in 50-μm-thick sections is not limited by penetration issues. The same, as mentioned before, does not apply to [3H]dT-labeled cells because detection of [3H]dT is restricted to a few micrometers deep from the surface of the section. This is due to the limited impregnation of the tissue and the fact that the half-distance of the beta particle emitted by tritium atom decay is ∼1 μm. Previous experiments, in methacrylate-embedded tissue, showed that only the top 2–3 μm contain silver grains. But this also depended on the thickness of the sections and emulsion (Caviness and Barkley, 1971). In our case, the tissue was not embedded, making the surface more wavy and more accessible to the emulsion, which would result in slightly deeper detection. Also, if nuclei are of the order of 10 μm in diameter, in 50-μm-thick tissue, ∼20% of them are exposed to the surface and a higher percentage would be exposed with detectability to deeper locations (e.g., ∼26–30% to the top 3–5 μm). Considering the variables at play and our microscopic observations, since we cannot know with absolute certainty the ratio of the number of [3H]dT-positive nuclei detected to [3H]dT-positive nuclei present in the sample, we calculated the number of [3H]dT-positive cells detected to represent detection within 2.5, 5, and 10 μm (Table 2). In any case, the number of plotted [3H]dT-labeled cells in a section represents only a fraction of those present in the section at the time of detection. Therefore, to be able to compare [3H]dT and BrdU cell numbers, we multiplied 5×, 10×, or 20× the number of [3H]dT cells detected, corresponding to scenarios in which [3H]dT cells were detected within 10, 5, or 2.5 μm from the surface of the tissue, correspondingly. The multiplication factor equalizes all [3H]dT tissue to a corresponding thickness of 50 μm, the same thickness of the BrdU material. Since detectability within 10 μm is the extreme case best scenario and the one that would make the number of detected [3H]dT and BrdU cells the closest, we used this case for comparison reasoning that, if differences in cell numbers are still detected, they must indeed exist.
Shrinkage corrections were done in 3D using the appropriate tool in StereoInvestigator. To get an accurate measure of cortical thickness, we measured the distance from pia to the WM along the sides of every counting box and added a third measurement in the middle. Then we obtained an average cortical thickness at the exact place where cells were being plotted and averaged those averages to get a measure of thickness for each cortical area, for both the BrdU and [3H]dT material. The thickness of each cortical area, section by section, was then standardized to the average thickness obtained in the BrdU material, which was the most homogeneous. Another possible correction should take into consideration the number of generations that are detectable by the two methods. In principle, the amount of label per nucleus should be halved with each cell cycle. BrdU allows detection of only approximately three cell cycles (Hayes and Nowakowski, 2002). For [3H]dT, since we are using ∼3 grains per nucleus (provided a clean background), and assuming an average maximum of 50 grains/nucleus, we might be detecting between three and four cell cycles. However, because the great majority of nuclei counted had more than just three grains, we expect the number of positive labeled cells we detect on both cases to correspond roughly to a maximum of three cell cycles for both markers.
Results
In every cortical area analyzed and for both injection times, [3H]dT-outnumber BrdU-labeled cells. In addition, [3H]dT-labeled cells formed narrow (a few hundred micrometers wide) bands at discrete depths from pia, while BrdU-labeled cells were much more widespread.
Assessment of labeled cell numbers
Even assuming the extreme case in which [3H]dT is detected within 10 μm from the top surface of the tissue (see Materials and Methods, Technical considerations and corrections of methodological differences), at both time points and in each cortical area, the number of [3H]dT-labeled cells was substantially larger than that of BrdU (Table 2, Fig. 3A,B). Although the incorporation of markers could affect the cell cycle, in the absence of precise data to this effect, we assumed that the cell cycle itself does not suffer major alterations. Hence, since both markers label cells in the S-phase of cell division, we suggest that the difference in the number of labeled cells is likely due to differential cell death and that, for the case under consideration, on average, at least ∼60 and ∼50% of BrdU cells died after injections at E70 and E55, correspondingly (Table 2). These percentages are even higher if detectability of [3H]dT is reduced from 10 to 5 or 2.5 μm.
Figure 3.

Cell numbers. Labeled (A, B) and plotted (C, D) mean number of BrdU and [3H]dT cells in a 500-μm-wide slab of cortex from pia to WM. Although more BrdU than [3H]dT cells were plotted (C, D) and some of the differences reached statistical significance (ANOVA at the p < 0.05 level); after corrections (E, F) (see Materials and Methods, Technical considerations and corrections of methodological differences), it was determined that there are more [3H]dT than BrdU-labeled cells (A, B). E and F illustrate the factor by which each cortical area was corrected to normalize the cortical thickness to the mean cortical thickness measured in the BrdU cases. Cortical thickness was highly variable in the [3H]dT cases and rather constant in BrdU cases. *Denotes statistical significance. Error bars indicate SEM.
For postnatal animals injected at E70, the distributions of the mean number of [3H]dT- and BrdU-labeled cells per cortical area were all normal and had equal variances. [3H]dT-labeled cells outnumber BrdU-labeled cells in all cortical areas, and the difference is statistically significant in all areas except the VIS cortex (Fig. 3A). If detectability of [3H]dT is considered for 2.5 and 5 μm, then, even for the VIS cortex, the difference is statistically significant. The mean number of plotted BrdU cells across different cortical areas (Table 3) was broad, in the range (186–461) with highly significant differences (F(4,20) = 66.58; p < 0.001), a result in agreement with Kruskal–Wallis testing (χ2 = 16.45; df = 4; p = 0.003). In contrast, the mean number of plotted [3H]dT cells across different cortical areas (Table 3) was tight (range, 110.2–160), with no region having significantly more positive cells than any other region (F(4,20) = 1.21; p = 0.337). Still, if due to the large difference in the BrdU cell numbers between the VIS cortex and other cortical areas, we remove the VIS cortex from the analysis, there is a significant difference in the new groups (F(3,16) = 3.52; p = 0.039). Hence, to investigate how the mean number of BrdU plotted cells differed among cortical areas, we performed LSD-corrected multiple-comparisons tests and established statistically significant differences between different cortical areas (Table 3). With a more stringent Bonferroni correction, the differences between the VIS and other cortical areas are still detected.
For postnatal animals injected at E55, the mean number of plotted [3H]dT cells across different cortical areas was less tight than in the E70 cases (range, 63–161), due mostly to a very low number of cells in the SOM cortex. If the SOM is removed from the sample, the numbers are in a similar range to those at E70. There was also similar variability in BrdU plotted cell numbers (range, 195–398) across cortical regions; and here, as in the E70 injections, the VIS cortex contained the largest number of BrdU cells (Table 3, Fig. 3C,D). Here too, when using a [3H]dT maximum detectability within 10 μm (see Technical considerations and corrections of methodological differences, above), a comparison of plotted cell numbers indicates statistically significant differences in all cortical areas except the PFC. After corrections, a comparison of labeled cell numbers shows that [3H]dT cells outnumber BrdU-labeled cells in all cortical areas studied and that differences are statistically significant in all areas except the SOM cortex (Fig. 3B–D). If detectability of [3H]dT is considered for 2.5 and 5 μm, then all [3H]dT and BrdU differences in labeled cell numbers are statistically significant.
BrdU-labeled cells are more widely dispersed than [3H]dT-labeled cells
All animals were killed between 2.5 and 3 months of age, in this species a time when neurons have completed migration to their final areal and laminar positions (Rakic, 1974, 2002a). With and without shrinkage corrections (including normalization of cortical thickness), plotting of the histograms representing the continuous distribution of mean percentage BrdU and [3H]dT cells in bins of 100 μm from pia to (and including) the WM indicated that, for the same injection times and for corresponding cortical areas, the peak accumulation of BrdU and [3H]dT cells does not coincide and BrdU-labeled cells are more widely dispersed (Figs. 4A–D, 5A–D).
Figure 4.
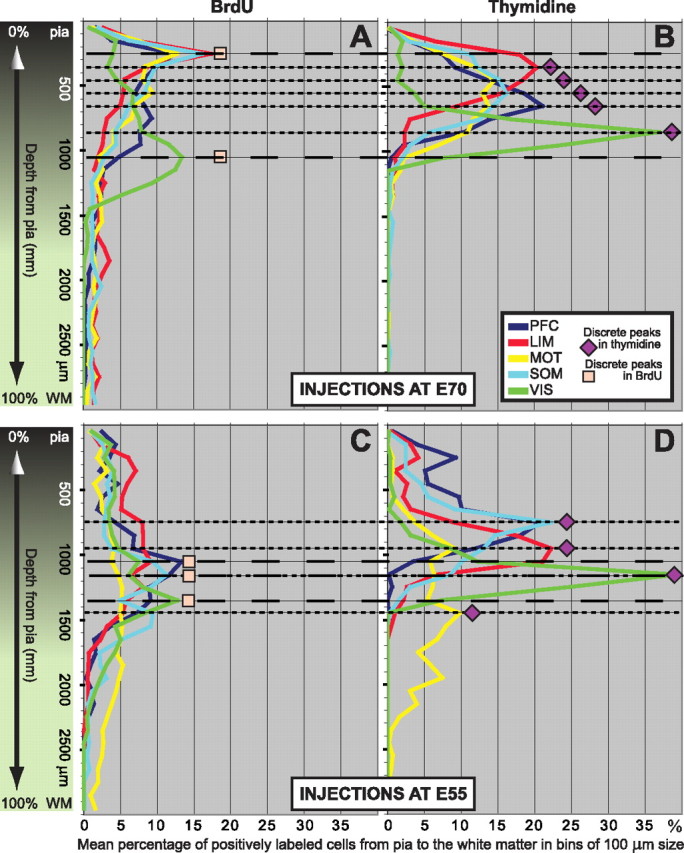
BrdU-positive cells are significantly more dispersed than [3H]dT-positive cells. A–D, Distribution plots of the mean percentage BrdU and [3H]dT cells in bins of 100 μm from pia to (and including) WM. As indicated by color gradient, the transition from gray matter to WM is not an abrupt one. WM border usually initiates at ∼1800 μm deep, but this depends on the region, angle of cut, etc. A, B, Injections at E70. A, BrdU peaks for the PFC, LIM, MOT, and SOM cortices coincide in depth between 200 and 300 μm from pia. The peak for VIS cortex is between 1000 and 1100 μm deep. In addition to these peaks, for all BrdU cases there are other quasi-salient peaks at different depths, and in most cases >10% of the cells accumulate deeper to where no [3H]dT cells accumulate. Between 2 and 3% of the cells are found at very deep locations including within the WM (>1900 μm from pia). Because of the spreading of the distributions, the largest accumulation of BrdU cells in a single bin is low, in the range from ∼12 to 17%. B, [3H]dT peaks are discrete (i.e., separate from each other) and compact. In order of depth are LIM (at 300–400 μm), MOT (at 400–500 μm), SOM (at 500–600 μm), PFC (at 600–700 μm), and VIS (at 800–900 μm) cortices. There is no accumulation of cells at deeper locations and no cells found in the WM. Consequently, in each cortical area the largest accumulation of [3H]dT cells in a single bin is high, in the range from ∼15 to 37%. C, D, Injections at E55. C, D, BrdU (C) and [3H]dT (D) distributions are wider when compared with the corresponding ones for injections at E70 but, when compared with each other, BrdU cells (C) are still more dispersed than the corresponding [3H]dT cells (D). As with the E70 cases, the total percentage accumulation per bin for BrdU is consequently lower that of [3H]dT. The spreading of the BrdU cells is therefore a function of the marker and not of the time at which it was injected. To facilitate comparisons, all scales are the same and the position of BrdU or [3H]dT distribution peaks are indicated by long and short dashed lines, respectively.
Figure 5.
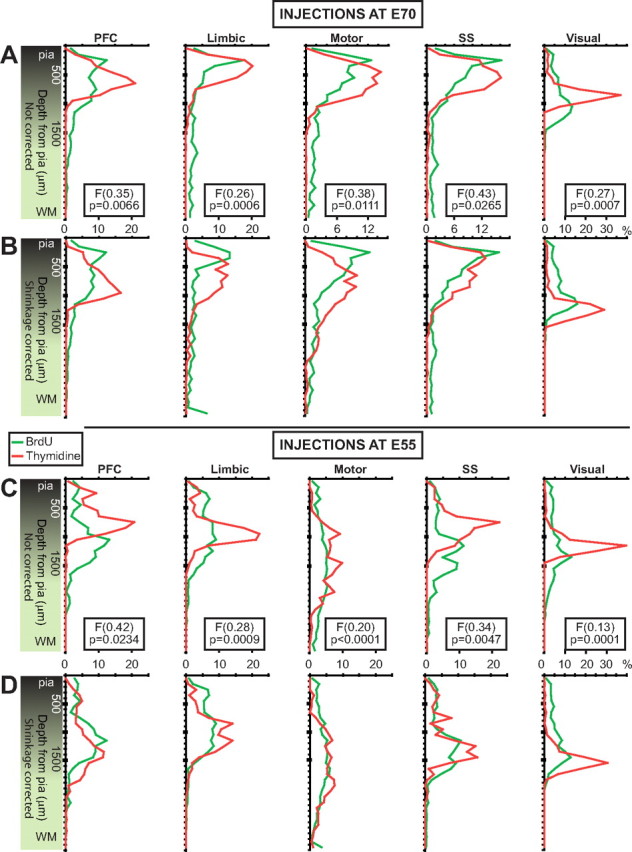
BrdU versus [3H]dT cell dispersion per cortical area. To facilitate comparison, the distributions are now shown per cortical area. Corresponding uncorrected (top) and corrected (bottom) distributions for injections at E70 (A, B) and at E55 (C, D). In all cases, independently of whether or not the cortical thickness is normalized, BrdU and [3H]dT peaks do not coincide in depth from pia. Normalization has little effect on BrdU distributions but decreases the amplitude and deepens (with respect to pia) the peaks of the [3H]dT distributions. [3H]dT distributions are more compact than BrdU distributions indicating that BrdU cells are more widespread. All the F scores for dispersion have significant p values. F scores are not provided for the normalized cases because normalization renders the F test less powerful.
In the monkey exposed to [3H]dT at E70, the apices of the distributions of labeled cells for different cortical areas (Fig. 4B) are each confined within distinct bins situated at different depths from the pia. In all cortical areas, the distributions are relatively narrow, and the peak amplitudes are high. Most labeled cells are close together and form a distinct band that is visible with both light and dark-field microscopy. In contrast, the apices of the distributions of BrdU-labeled cells for all cortical areas, except VIS, are located within the same bin (Fig. 4A), and the distributions are wide and their amplitude low. Importantly, there are more BrdU than [3H]dT cells at deeper locations, including in the WM. Without correcting for cortical thickness, the accumulation of BrdU cells does not surpass 18% at any given bin, while the distribution of [3H]dT cells is much more compact with PFC, LIM, and VIS, all accumulating >20% of the cells in a single bin (Table 2). Normalization of cortical thickness per cortical area displaces the [3H]dT distribution curves more than the corresponding BrdU distribution curves (Fig. 5). Consequently, normalization has most of the effect on the [3H]dT percentage accumulation (lower per 100 μm bin) and positional depth of the highest peaks (deeper, closer to WM) and almost no effect on the BrdU distributions. After cortical thickness normalization, the accumulation of BrdU cells does not surpass 16% at any given bin, while the corresponding peaks in the [3H]dT distributions drop (Table 2). However, BrdU cells are still much more broadly distributed, while [3H]dT cells are still more compactly distributed (Fig. 5, compare A, B).
Similarly, in the monkeys exposed to [3H]dT at E55, the corresponding distributions are much more compact than the BrdU distributions. PFC, LIM, SOM, and VIS cortices all accumulate >20% of the cells within a single bin (Table 2). For BrdU, the apices of the distributions of labeled cells in the PFC and LIM coincide in depth, SOM and VIS cortices have discrete peak apices, and MOT cortex does not really have a salient peak (Fig. 4C). Here, much more pronounced than in the E70 cases, there are quasi-salient peaks at different depths particularly for the LIM, MOT, and SOM cortices. Also, BrdU-labeled cells distribute broadly, and their accumulation, even after cortical thickness normalization, does not surpass 14% at any given bin.
The effect of normalizing cortical thickness in monkeys exposed at E55 is identical with that described for the E70 cases. The [3H]dT peaks drop in amplitude and are displaced to deeper locations, but the differences between BrdU and [3H]dT distributions persist (Fig. 5, compare C, D). A sizable percentage of BrdU cells labeled by injection at E70 accumulate in deep bins, while in the animals exposed to [3H]dT at the same embryonic age, most labeled cells reach more superficial bins (Table 2). Deeper than 1900 μm, which in the PFC, LIM, and VIS correspond to the WM, and in MOT and SOM cortices correspond to either WM or lower layer VI, the difference between the mean percentage numbers of BrdU and [3H]dT cells, in all cortical areas examined, is substantial (Table 2).
In animals injected at E55, many labeled cells are normally destined for the deeper layers; despite this, as was the case at E70, there are more BrdU than [3H]dT cells found at very deep locations, including in the WM (Table 2).
Visual examination of the distributions for corresponding injection times, independently of cortical thickness normalization, indicates that the central locations (median) of the BrdU and [3H]dT percentage distributions are different between corresponding cortical areas. Furthermore, nonparametric Kruskal–Wallis analysis demonstrates that these differences are statistically significant in several areas and F tests indicate the differences are due to dispersion (Table 3). Together, these analyses validate the observation that although smaller in number, the BrdU-labeled cells are more dispersed than [3H]dT-labeled cells.
The distributions of BrdU and [3H]dT-labeled cells display strong correlation but poor agreement
To investigate the strength of the association between the mean percentage distributions of BrdU- and [3H]dT-labeled cells for corresponding cortical areas, we calculated the corresponding Spearman rank correlation coefficients (rs). This analysis indicates the degree of association between the distribution obtained with [3H]dT and the corresponding one obtained with BrdU. A significant correlation was found in every case, with all the rs having a significant p value with and without normalization of cortical thickness (Table 3). This is not surprising since we expected the position of both [3H]dT and BrdU cells to be correlated. Their presence and position is a function of the precise time at which cells have been labeled (i.e., of injection times) and of the migratory processes that follow. However, comparison of final cell position is not a matter of correlation but a matter of agreement. Because a significant correlation does not automatically imply that there is good agreement between the two methods and the distributions do look different, the degree of agreement between the corresponding distributions was evaluated using Bland–Altman analysis. Here, we address the question of whether or not, despite the difference in dispersion between BrdU and [3H]dT cells, as methods, BrdU and [3H]dT labeling can be used interchangeably. The results, in accord with the previous analysis, indicate clearly that there are significant differences between the distributions of BrdU and [3H]dT cells in all cortical areas (Fig. 6).
Figure 6.
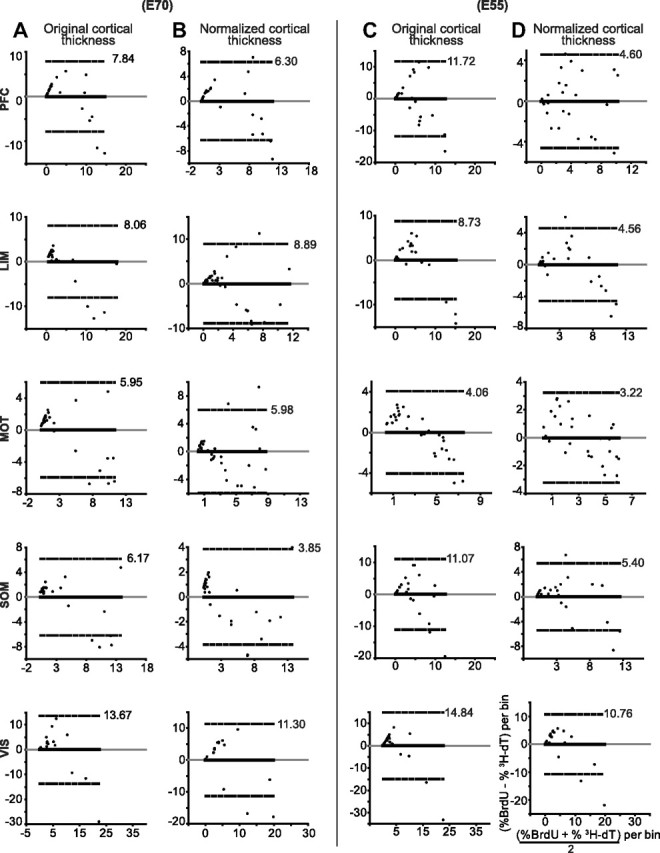
Agreement between BrdU and [3H]dT cell distributions is poor. A–D, Bland–Altman difference plots (also called Tukey's mean difference plots) indicate poor agreement between BrdU and [3H]dT distributions for all injection times and in all cortical areas. The smallest limits of agreement with semibalance differences occur in the MOT cortex at E55 probably due to the poor development of the MOT cortex at the time of injection. The widest limits of agreement are all in the VIS cortex (a very active yet undeveloped area for both injection times). Normalization of cortical thickness improves the limits of agreement for most distributions, but limits are still wide. Bias (solid black) and identity (gray) lines are all at zero because the distributions are for normalized (percentage) data. The dashed lines represent the 95% agreement limits, of which only the positive value is shown. Since the 3000-μm-long counting boxes were divided in bins of 100 μm height, each of the 30 dots in each plot represents an ordered pair in which the ordinate is the difference in percentage and the abscissa is the mean percentage for the corresponding bins (i.e., bins at the same depth from pia) of the BrdU and [3H]dT distributions. The coordinate labels in the last plot apply to all plots.
BrdU cells both bypass and lag behind their targets compared with [3H]dT cells
Since the apices of the tallest peaks in the distributions of BrdU and [3H]dT cells do not coincide with respect to depth from pia, we examined the proportion of BrdU-labeled cells above and below their laminar target. Assuming that the distributions of [3H]dT cells were closer to the proper and correct distributions, we normalized the cortical thickness per cortical area and calculated the percentage of [3H]dT and of BrdU cells correspondingly above and below the apex of the largest [3H]dT peak (or center location between two largest peaks). The results indicate that, for animals injected at E55, a certain percentage of the BrdU cells bypass their proper depth and locate themselves more superficially than they should in the PFC, LIM, and MOT cortices, while in the SOM and VIS cortices a certain percentage lags behind (i.e., remains deeper in the cortex) (Fig. 7). However, later on, as the results of animals injected at E70 indicate, the tendency reverses and in the PFC, LIM, and MOT cortices BrdU cells lag behind while they bypass their proper depth in the SOM and VIS cortices. Overall, our data indicate that, while some BrdU-labeled cells bypass their target position, other may lag behind and take deeper positions.
Figure 7.
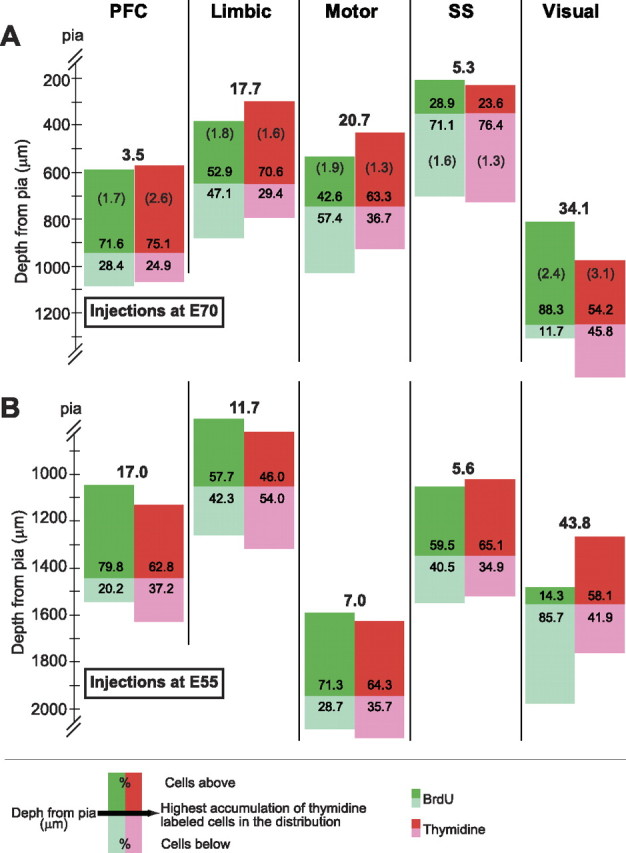
Some BrdU cells are lost: they can bypass or lag behind their target positions. We calculated the percentage accumulation of BrdU and [3H]dT cells with respect to the highest peak(s) in the [3H]dT distributions after normalization of cortical thickness per cortical area. The difference (percentage) between the distributions is indicated in bold numbers above them. A, For injections at E70, some BrdU cells lag behind their proper depth in the PFC, LIM, and MOT cortices, while in SOM and VIS cortices they overshoot their target and position themselves more superficially than they should. B, Early on (at E55) the pattern was exactly the opposite, some BrdU cells in the PFC, LIM, and MOT cortices bypass their target depth with reference to pia and position themselves more superficially that they should, while in SOM and VIS cortices they lag behind. The numbers in parentheses (E70 cases; a single pair of animals in each case) indicate that the within-group differences are smaller than the between-group differences for each marker and within each cortical area.
Importantly, the differences between groups ([3H]dT vs BrdU) are greater than the differences within a group of animals labeled with the same marker. To illustrate this, two BrdU (two males; both E70–P75) and two [3H]dT cases (unknown gender; E62–P50 vs E70–P98) were chosen for closer comparison. Since the difference between groups is smallest (3.5%) in the PFC for injections at E70, it is important to establish that this difference is still larger than that between two animals labeled with the same marker. This was indeed the case. The average difference in the PFC between the two BrdU cases was 1.7%, and the average difference between the two [3H]dT cases was 2.6%. These differences were similarly small in all areas compared with the between group differences. The largest difference within the same area was 3.1% in the visual cortex of the [3H]dT–E70 case. Within-group differences were not calculated for injections at E55 (Fig. 7).
Discussion
Our analysis shows that incorporation of BrdU into the cell has a sizable effect on labeled cell numbers and final positions. Compared with [3H]dT, lower BrdU cell numbers are likely due to decreased cell survival, rather than decreased cell proliferation. Final cell position is affected by faulty migration that gives rise to higher BrdU-labeled cell dispersion. We suggest that both effects may be mediated by BrdU toxicity, which compromises DNA translational function and gene expression.
BrdU is more toxic than [3H]dT
We hypothesize that higher toxicity of BrdU than [3H]dT mediates random effects on gene expression making the resulting changes difficult to predict and assess. In fact, it should not be surprising that incorporation of BrdU into DNA is more toxic than incorporation of [3H]dT. The only difference between [3H]dT and the normal endogenous nucleotide is an extra neutron in a H atom. Consequently, DNA with [3H]dT possibly reflects closer DNA in the naive noninjected animal. BrdU has a different structure and integrates a totally foreign atom (Br) into replicating DNA when it introduces bromouracil in lieu of thymidine (Brockman and Anderson, 1963; Hitchings and Elion, 1967). This causes the subsequent faulty base pairing of bromouracil with guanine instead of adenine (Roy-Burman, 1970). Since BrdU is chemically and structurally significantly different from [3H]dT, genes that use it as its substitute are unlikely to transcribe correctly into RNA and eventually the proper protein. It is almost impossible to expect that such drastic changes in the double helix would not have deleterious effects on cell fate and function.
BrdU toxicity has been documented in a host of studies both in vitro and in vivo in a dose-dependent manner, and toxic effects have been shown at even lower doses than what we and most other laboratories use (Biggers et al., 1987; Kolb et al., 1999; Sekerková et al., 2004) (for review, see Breunig et al., 2007). BrdU has detrimental effects on cell cycle, differentiation, and survival of neural progenitor cells (Lehner et al., 2011), and its adverse effects on chromosomes have been known for almost 50 years (Hsu and Somers, 1961). BrdU produces morphological and behavioral abnormalities; increases sister chromatid exchanges, mutations, and breaks; alters DNA stability; and may render cells unable to divide or carry out normal division (Bannigan and Langman, 1979; Saffhill and Ockey, 1985; Morris, 1991; Kolb et al., 1999). At higher doses, (400–600 mg/kg; mouse), BrdU causes an increase in the duration of the S-phase and mitosis, making the cell cycle 5 h longer than normal with the additive effects of cell death and retardation of the cell cycle causing a 15% deficit of Purkinje cells in the postnatal cerebellum without interfering with cell differentiation (Bannigan and Langman, 1979). Similar effects have been reported in neocortex (Webster et al., 1973) where BrdU prolonged the duration of mitosis, did not block cell division, and also caused cell death.
BrdU affects cell numbers and/or cell survival
Our results indicate that BrdU toxicity compromises the number of labeled cells and/or the number of cells that survive. In rodents, the laminar distribution of [3H]dT and BrdU cells closely corresponds and the number of labeled cells is similar after simultaneous injections of either marker at E19 with killing 3 d later (Miller and Nowakowski, 1988). In contrast, the longer waiting period (∼6 months) between injection and evaluation of cell numbers and positions and the larger primate brain in our study provide higher resolution for detecting intermarker differences.
The lower number of BrdU- than [3H]dT-labeled cells we observed is unlikely due to a lower comparative dosage. The amount of time that the marker is available for incorporation in relation to the length of the S-phase may cause differences in labeling. For instance, low-intensity labeling with both markers may result from their availability only during a portion of the S-phase although also from marker dilution due to multiple cell cycles. To our knowledge, there are no studies determining exactly what comparable doses of [3H]dT and BrdU are in vivo for the primate brain. Although larger doses of BrdU could result in longer availability and increase the pool of precursors that could be labeled (Böswald et al., 1990), they would also be more toxic and likely increase cell death. Thus, lower BrdU cell numbers may be caused by inhibition of cell proliferation as well as effects on cell cycle exit behavior or increased cell death.
The length of the S-phase progressively lengthens over successive cell cycles in mice (3–5 h) and rats (5–9 h), while in macaque monkey it first lengthens (from 5.2 h at E40 to 14.9 h at E60) and subsequently shortens (4.6 h at E80) (von Waechter and Jaensch, 1972; Miller and Nowakowski, 1991; Reznikov and van der Kooy, 1995; Takahashi et al., 1995) (for review, see Kornack and Rakic, 1998). For ages E55 and E70, we estimate the length of the S-phase to be ∼10 and ∼12 h, correspondingly [Kornack and Rakic (1998), their Fig. 3].
In mice and rats, the time of tracer clearance from blood serum ranges from 0.5 to 1 h, which is short compared with the length of the S-phase and the reason why single injections are considered a single pulse of tracer (Lamerton and Fry, 1963; Cleaver, 1967; Packard et al., 1973; Nowakowski and Rakic, 1974) (for review, see Taupin, 2007). Also, the kinetics of [3H]dT and BrdU in mice is similar in that the loading time to label an S-phase cell to detectability is <0.2 h and cells that enter the S-phase continue to be detectable for 5–6 h (Hayes and Nowakowski, 2000). [3H]dT, after a single intravenous injection in a monkey at the dose we used, is available for incorporation for ∼10 min (Nowakowski and Rakic, 1974). Hence, if we assume similar loading times and kinetics for [3H]dT and BrdU transport across the placental barrier, then, even if availability ranges are somewhat different, given the length of the S-phase compared with marker availability and the prolonged tempo of corticogenesis in this species, injections of either marker at the same embryonic age should result in approximately the same number of labeled cells with similar distribution. Since this is not the case, it seems that, over time, BrdU toxicity interferes with cell survival. In addition, larger variability in the number of BrdU- than [3H]dT-labeled cells among cortical areas may indicate that the cell cycle kinetics of the corresponding progenitors may be different in each cortical area (Dehay et al., 1993) or that cells are affected differently by incorporation of BrdU. Furthermore, a very important but often neglected caveat is that BrdU can induce mitotic division that would not normally occur and also can be incorporated into DNA of nondividing postmitotic cells and into dying cells (Goldsworthy et al., 1992; Yang et al., 2001; Kuan et al., 2004; Qu et al., 2004; Caldwell et al., 2005; Breunig et al., 2007). Thus, cell death may be even larger if BrdU initially increased cell division. These issues can be resolved only by short time experiments not practical in primates.
BrdU disturbs cell migration and ultimately their final position
Our study indicates that BrdU compromises cell migration and influences final cell position. Normal migration and proper positioning of neurons in the cortex are processes genetically determined (Rakic, 1974; Shen et al., 2006). BrdU incorporation into the cell nucleus may have deleterious effects on gene expression necessary for providing postmitotic migrating cells with information regarding polarity, selection of pathway substrate, migration rate, guidance to proper lamina, and instructions for differentiation and connectivity.
After normalizing for cortical thickness, in all cortical areas for E70 cases and in all except MOT cortex for E55 cases, the highest peaks of the BrdU distributions were consistently more superficial than the corresponding [3H]dT peaks. One possible explanation is that many BrdU cells were labeled slightly later than [3H]dT cells and that this would be an indication of lengthening of BrdU availability, delays in the ability of BrdU cells to divide, or that some underwent a waiting period before continuing cell division. However, our findings also show more BrdU than [3H]dT cells at deeper locations, suggesting that BrdU cells are “lost” in their migration and cannot find their target position.
Our data indicate that cells found at a particular cortical area were destined to be there but made errors in finding their proper radial position. We consider differences in lamina positions even more striking than differences in labeled cell numbers because they are independent of technical corrections. We interpret both findings to be indicative of unequivocal biological differences between the two labels.
In conclusion, it is essential to take into account BrdU toxicity and its adverse effects when evaluating experimental results using this compound. This is especially important when investigators make multiple injections or use continuous exposure and assume that cells that have incorporated BrdU divide, migrate, or differentiate normally and then assume normal function. While BrdU and [3H]dT may be equally efficient to study areal neurogenesis during specific developmental periods, the details they provide on lamina formation, as well as magnitude of cell proliferation and survival rate, are quite different. We conclude that extreme caution should be exercised when interpreting the results of BrdU cell labeling as a possible indicator of the magnitude of cell division, pattern of migration, final position, and fate, and particularly, the accuracy of quantifications of these cell events based on this method.
Footnotes
This work was supported by a grant from NIH–NINDS. We thank Anita Begovic, Adi Begovic, Jacob A. Kravitz, Mariamma Pappy, and Marianne Horn for technical assistance.
The authors declare no competing financial interests.
References
- Altman D, Bland J. Measurement in medicine: the analysis of method comparison studies. Statistician. 1983;32:307–317. [Google Scholar]
- Altman J. Autoradiographic investigation of cell proliferation in the brains of rats and cats. Anat Rec. 1963;145:573–591. doi: 10.1002/ar.1091450409. [DOI] [PubMed] [Google Scholar]
- Altman J. DNA metabolism and cell proliferation. In: Lajtha A, editor. Handbook of neurochemistry. New York: Plenum; 1969. pp. 137–182. [Google Scholar]
- Angevine JB., Jr Time of neuron origin in the hippocampal region. An autoradiographic study in the mouse. Exp Neurol Suppl. 1965;2:1–70. [PubMed] [Google Scholar]
- Bannigan J, Langman J. The cellular effect of 5-bromodeoxyuridine on the mammalian embryo. J Embryol Exp Morphol. 1979;50:123–135. [PubMed] [Google Scholar]
- Biggers WJ, Barnea ER, Sanyal MK. Anomalous neural differentiation induced by 5-bromo-2′-deoxyuridine during organogenesis in the rat. Teratology. 1987;35:63–75. doi: 10.1002/tera.1420350110. [DOI] [PubMed] [Google Scholar]
- Bisconte JC. Kinetics analysis of cellular populations by means of the quantitative radioautography. In: Bourne GH, Danielli JF, editors. International review of cytology. New York: Academic; 1979. pp. 75–126. [DOI] [PubMed] [Google Scholar]
- Bland JM, Altman DG. Statistical methods for assessing agreement between two methods of clinical measurement. Lancet. 1986;1:307–310. [PubMed] [Google Scholar]
- Böswald M, Harasim S, Maurer-Schultze B. Tracer dose and availability time of thymidine and bromodeoxyuridine: application of bromodeoxyuridine in cell kinetic studies. Cell Tissue Kinet. 1990;23:169–181. doi: 10.1111/j.1365-2184.1990.tb01113.x. [DOI] [PubMed] [Google Scholar]
- Breunig JJ, Arellano JI, Macklis JD, Rakic P. Everything that glitters isn't gold: a critical review of postnatal neural precursor analyses. Cell Stem Cell. 2007;1:612–627. doi: 10.1016/j.stem.2007.11.008. [DOI] [PubMed] [Google Scholar]
- Brockman RW, Anderson EP. Biochemistry of cancer (metabolic aspects) Annu Rev Biochem. 1963;32:463–512. doi: 10.1146/annurev.bi.32.070163.002335. [DOI] [PubMed] [Google Scholar]
- Caldwell MA, He X, Svendsen CN. 5-Bromo-2′-deoxyuridine is selectively toxic to neuronal precursors in vitro. Eur J Neurosci. 2005;22:2965–2970. doi: 10.1111/j.1460-9568.2005.04504.x. [DOI] [PubMed] [Google Scholar]
- Caviness VS, Jr, Barkley DS. Section thickness and grain count variation in tritium autoradiography. Stain Technol. 1971;46:131–135. doi: 10.3109/10520297109067837. [DOI] [PubMed] [Google Scholar]
- Cleaver J. Tymidine metabolism and cell kinetics. Amsterdam: North-Holland; 1967. [Google Scholar]
- Dehay C, Giroud P, Berland M, Smart I, Kennedy H. Modulation of the cell cycle contributes to the parcellation of the primate visual cortex. Nature. 1993;366:464–466. doi: 10.1038/366464a0. [DOI] [PubMed] [Google Scholar]
- del Rio JA, Soriano E. Immunocytochemical detection of 5′-bromodeoxyuridine incorporation in the central nervous system of the mouse. Brain Res Dev Brain Res. 1989;49:311–317. doi: 10.1016/0165-3806(89)90033-3. [DOI] [PubMed] [Google Scholar]
- Ehmann UK, Williams JR, Nagle WA, Brown JA, Belli JA, Lett JT. Perturbations in cell cycle progression from radioactive DNA precursors. Nature. 1975;258:633–636. doi: 10.1038/258633a0. [DOI] [PubMed] [Google Scholar]
- Goldsworthy TL, Dunn CS, Popp JA. Dose effects of bromodeoxyuridine (BRUD) on rodent hepatocyte proliferation measurements. Toxicologist. 1992;12:265. [Google Scholar]
- Gratzner HG. Monoclonal antibody to 5-bromo- and 5-iododeoxyuridine: a new reagent for detection of DNA replication. Science. 1982;218:474–475. doi: 10.1126/science.7123245. [DOI] [PubMed] [Google Scholar]
- Hayes NL, Nowakowski RS. Exploiting the dynamics of S-phase tracers in developing brain: interkinetic nuclear migration for cells entering versus leaving the S-phase. Dev Neurosci. 2000;22:44–55. doi: 10.1159/000017426. [DOI] [PubMed] [Google Scholar]
- Hayes NL, Nowakowski RS. Dynamics of cell proliferation in the adult dentate gyrus of two inbred strains of mice. Brain Res Dev Brain Res. 2002;134:77–85. doi: 10.1016/s0165-3806(01)00324-8. [DOI] [PubMed] [Google Scholar]
- Hitchings G, Elion G. Mechanisms of action of purine and pyrimidine analogs. In: Brodsky I, Kahn S, Moyer J, editors. Cancer chemotherapy, I. New York: Grune and Stratton; 1967. p. 26. [Google Scholar]
- Hsu TC, Somers CE. Effect of 5-bromodeoxyuridine on mamalian chromosomes. Proc Natl Acad Sci U S A. 1961;47:396–403. doi: 10.1073/pnas.47.3.396. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kolb B, Pedersen B, Ballermann M, Gibb R, Whishaw IQ. Embryonic and postnatal injections of bromodeoxyuridine produce age-dependent morphological and behavioral abnormalities. J Neurosci. 1999;19:2337–2346. doi: 10.1523/JNEUROSCI.19-06-02337.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kornack DR, Rakic P. Changes in cell-cycle kinetics during the development and evolution of primate neocortex. Proc Natl Acad Sci U S A. 1998;95:1242–1246. doi: 10.1073/pnas.95.3.1242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuan CY, Schloemer AJ, Lu A, Burns KA, Weng WL, Williams MT, Strauss KI, Vorhees CV, Flavell RA, Davis RJ, Sharp FR, Rakic P. Hypoxia-ischemia induces DNA synthesis without cell proliferation in dying neurons in adult rodent brain. J Neurosci. 2004;24:10763–10772. doi: 10.1523/JNEUROSCI.3883-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuwagata M, Ogawa T, Nagata T, Shioda S. The evaluation of early embryonic neurogenesis after exposure to the genotoxic agent 5-bromo-2′-deoxyuridine in mice. Neurotoxicology. 2007;28:780–789. doi: 10.1016/j.neuro.2006.07.017. [DOI] [PubMed] [Google Scholar]
- Lamerton L, Fry R. Cell proliferation. Oxford: Blackwell Scientific Publications; 1963. [Google Scholar]
- Lehner B, Sandner B, Marschallinger J, Lehner C, Furtner T, Couillard-Despres S, Rivera FJ, Brockhoff G, Bauer HC, Weidner N, Aigner L. The dark side of BrdU in neural stem cell biology: detrimental effects on cell cycle, differentiation and survival. Cell Tissue Res. 2011;345:313–328. doi: 10.1007/s00441-011-1213-7. [DOI] [PubMed] [Google Scholar]
- Leuner B, Glasper ER, Gould E. Thymidine analog methods for studies of adult neurogenesis are not equally sensitive. J Comp Neurol. 2009;517:123–133. doi: 10.1002/cne.22107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miller MW, Nowakowski RS. Use of bromodeoxyuridine-immunohistochemistry to examine the proliferation, migration and time of origin of cells in the central nervous system. Brain Res. 1988;457:44–52. doi: 10.1016/0006-8993(88)90055-8. [DOI] [PubMed] [Google Scholar]
- Miller MW, Nowakowski RS. Effect of prenatal exposure to ethanol on the cell cycle kinetics and growth fraction in the proliferative zones of fetal rat cerebral cortex. Alcohol Clin Exp Res. 1991;15:229–232. doi: 10.1111/j.1530-0277.1991.tb01861.x. [DOI] [PubMed] [Google Scholar]
- Morris SM. The genetic toxicology of 5-bromodeoxyuridine in mammalian cells. Mutat Res. 1991;258:161–188. doi: 10.1016/0165-1110(91)90007-i. [DOI] [PubMed] [Google Scholar]
- Nowakowski RS, Hayes NL. New neurons: extraordinary evidence or extraordinary conclusion? Science. 2000;288:771. doi: 10.1126/science.288.5467.771a. [DOI] [PubMed] [Google Scholar]
- Nowakowski RS, Rakic P. Clearance rate of exogenous 3H-thymidine from the plasma of pregnant rhesus monkeys. Cell Tissue Kinet. 1974;7:189–194. doi: 10.1111/j.1365-2184.1974.tb00411.x. [DOI] [PubMed] [Google Scholar]
- Nowakowski RS, Rakic P. The mode of migration of neurons to the hippocampus: a Golgi and electron microscopic analysis in foetal rhesus monkey. J Neurocytol. 1979;8:697–718. doi: 10.1007/BF01206671. [DOI] [PubMed] [Google Scholar]
- Nowakowski RS, LaVail J, Rakic P. The correlation of the time of origin of neurons with their axonal projection: the combined use of [3H]thymidine autoradiography and horseradish peroxidase histochemistry. Brain Res. 1975;99:343–348. doi: 10.1016/0006-8993(75)90034-7. [DOI] [PubMed] [Google Scholar]
- Nowakowski RS, Lewin SB, Miller MW. Bromodeoxyuridine immunohistochemical determination of the lengths of the cell cycle and the DNA-synthetic phase for an anatomically defined population. J Neurocytol. 1989;18:311–318. doi: 10.1007/BF01190834. [DOI] [PubMed] [Google Scholar]
- Packard DS, Jr, Menzies RA, Skalko RG. Incorporation of thymidine and its analogue, bromodeoxyuridine, into embryos and maternal tissues of the mouse. Differentiation. 1973;1:397–404. doi: 10.1111/j.1432-0436.1973.tb00137.x. [DOI] [PubMed] [Google Scholar]
- Paxinos G, Huang X, Toga A. The rhesus monkey brain in stereotaxic coordinates. San Diego: Academic; 2000. [Google Scholar]
- Qu TY, Dong XJ, Sugaya I, Vaghani A, Pulido J, Sugaya K. Bromodeoxyuridine increases multipotency of human bone marrow-derived stem cells. Restor Neurol Neurosci. 2004;22:459–468. [PubMed] [Google Scholar]
- Rakic P. Kinetics of proliferation and latency between final cell division and onset of differentiation of cerebellar stellate and basket neurons. J Comp Neurol. 1973;147:523–546. doi: 10.1002/cne.901470407. [DOI] [PubMed] [Google Scholar]
- Rakic P. Neurons in rhesus monkey visual cortex: systematic relation between time of origin and eventual disposition. Science. 1974;183:425–427. doi: 10.1126/science.183.4123.425. [DOI] [PubMed] [Google Scholar]
- Rakic P. Prenatal genesis of connections subserving ocular dominance in the rhesus monkey. Nature. 1976;261:467–471. doi: 10.1038/261467a0. [DOI] [PubMed] [Google Scholar]
- Rakic P. Neurogenesis in adult primate neocortex: an evaluation of the evidence. Nat Rev Neurosci. 2002a;3:65–71. doi: 10.1038/nrn700. [DOI] [PubMed] [Google Scholar]
- Rakic P. Adult neurogenesis in mammals: an identity crisis. J Neurosci. 2002b;22:614–618. doi: 10.1523/JNEUROSCI.22-03-00614.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rakic P. Pre- and post-developmental neurogenesis in primates. Clin Neurosci Res. 2002c;2:29–39. [Google Scholar]
- Rakic P, Sidman RL. Supravital DNA synthesis in the developing human and mouse brain. J Neuropathol Exp Neurol. 1968;27:246–276. [PubMed] [Google Scholar]
- Rakic P, Sidman RL. Histogenesis of cortical layers in human cerebellum, particularly the lamina dissecans. J Comp Neurol. 1970;139:473–500. doi: 10.1002/cne.901390407. [DOI] [PubMed] [Google Scholar]
- Reznikov K, van der Kooy D. Variability and partial synchrony of the cell cycle in the germinal zone of the early embryonic cerebral cortex. J Comp Neurol. 1995;360:536–554. doi: 10.1002/cne.903600313. [DOI] [PubMed] [Google Scholar]
- Rogers AW. Techniques of autoradiography. Amsterdam: Elsevier; 1973. [Google Scholar]
- Roy-Burman P. Analogues of nucleic acid components. New York: Springer; 1970. [Google Scholar]
- Saffhill R, Ockey CH. Strand breaks arising from the repair of the 5-bromodeoxyuridine-substituted template and methyl methanesulphonate-induced lesions can explain the formation of sister chromatid exchanges. Chromosoma. 1985;92:218–224. doi: 10.1007/BF00348697. [DOI] [PubMed] [Google Scholar]
- Schlessinger AR, Cowan WM, Gottlieb DI. An autoradiographic study of the time of origin and the pattern of granule cell migration in the dentate gyrus of the rat. J Comp Neurol. 1975;159:149–175. doi: 10.1002/cne.901590202. [DOI] [PubMed] [Google Scholar]
- Sekerková G, Ilijic E, Mugnaini E. Bromodeoxyuridine administered during neurogenesis of the projection neurons causes cerebellar defects in rat. J Comp Neurol. 2004;470:221–239. doi: 10.1002/cne.11016. [DOI] [PubMed] [Google Scholar]
- Shen Q, Wang Y, Dimos JT, Fasano CA, Phoenix TN, Lemischka IR, Ivanova NB, Stifani S, Morrisey EE, Temple S. The timing of cortical neurogenesis is encoded within lineages of individual progenitor cells. Nat Neurosci. 2006;9:743–751. doi: 10.1038/nn1694. [DOI] [PubMed] [Google Scholar]
- Sidman RL. Autoradiographic methods and principles for study of the nervous system with thymidine-3H. New York: Springer; 1970. [Google Scholar]
- Sidman RL, Miale IL, Feder N. Cell proliferation and migration in the primitive ependymal zone: an autoradiographic study of histogenesis in the nervous system. Exp Neurol. 1959;1:322–333. doi: 10.1016/0014-4886(59)90024-x. [DOI] [PubMed] [Google Scholar]
- Takahashi T, Nowakowski RS, Caviness VS., Jr Early ontogeny of the secondary proliferative population of the embryonic murine cerebral wall. J Neurosci. 1995;15:6058–6068. doi: 10.1523/JNEUROSCI.15-09-06058.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taupin P. BrdU immunohistochemistry for studying adult neurogenesis: paradigms, pitfalls, limitations, and validation. Brain Res Rev. 2007;53:198–214. doi: 10.1016/j.brainresrev.2006.08.002. [DOI] [PubMed] [Google Scholar]
- von Waechter R, Jaensch B. Generation times of the matrix cells during embryonic brain development: an autoradiographic study in rats. Brain Res. 1972;46:235–250. doi: 10.1016/0006-8993(72)90018-2. [DOI] [PubMed] [Google Scholar]
- Webster W, Shimada M, Langman J. Effect of fluorodeoxyuridine, colcemid, and bromodeoxyuridine on developing neocortex of the mouse. Am J Anat. 1973;137:67–85. doi: 10.1002/aja.1001370106. [DOI] [PubMed] [Google Scholar]
- Yang Y, Geldmacher DS, Herrup K. DNA replication precedes neuronal cell death in Alzheimer's disease. J Neurosci. 2001;21:2661–2668. doi: 10.1523/JNEUROSCI.21-08-02661.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]