

Abstract
Brain tumors have been suggested to possess a small population of stem cells that are the root cause of tumorigenesis. Neurosphere assays have been generally adopted to study the nature of neural stem cells, including those derived from normal and tumorous tissues. However, appreciable amounts of differentiation and cell death are common in cultured neurospheres likely due to sub-optimal condition such as accessibility of all cells within sphere aggregates to culture medium.
Medulloblastoma, the most common pediatric CNS tumor, is characterized by its rapid progression and tendency to spread along the entire brain-spinal axis with dismal clinical outcome. Medulloblastoma is a neuroepithelial tumor of the cerebellum, accounting for 20% and 40% of intracranial and posterior fossa tumor in childhood, respectively1. It is now well established that Shh signaling stimulates proliferation of cerebellar granule neuron precursors (CGNPs) during cerebellar development 2-4. Numerous studies using mouse models, in which the Shh pathway is constitutively activated, have linked Shh signaling with medulloblastoma 5-9.
A recent report has shown that a subset of medulloblastoma cells derived from Patched1LacZ/+ mice are cancer stem cells, which are capable of initiating and propogating tumors 10. Here we describe an efficient method to isolate, enrich and maintain tumor stem cells derived from several mouse models of medulloblastoma, with constitutively activated Shh pathway due to a mutation in Smoothened (11, hereon referred as SmoM2), a GPCR that is critical for Shh pathway activation. In every isolated medulloblastoma tissue, we were able to establish numerous highly proliferative colonies. These cells robustly expressed several neural stem cell markers such as Nestin and Sox2, can undergo serial passages (greater than 20) and were clonogenic. While these cultured tumor stem cells were relatively small, often bipoar with high nuclear to cytoplasmic ratio when cultured under conditions favoring stem cell growth, they dramatically altered their morphology, extended multiple cellular processes, flattened and withdrew from the cell cycle upon switching to a cell culture medium supplemented with 10% fetal bovine serum. More importantly, these tumor stem cells differentiated into Tuj1+ or NeuN+ neurons, GFAP+ astrocytes and CNPase+ oligodendrocytes, thus highlighting their multi-potency. Furthermore, these cells were capable of propagating secondary medulloblastomas when orthotopically transplanted into host mice.
Protocol
1. Micro-dissection of Tumor-bearing Cerebellum, Dissociation of Tumor Tissue and Plating
- Retrieval of tumor tissue
- Sick mice bearing medulloblastoma were often runted, displayed hydrocephaly and typical neurological symptoms, including posterior paralysis and failure to regain posture when overturned. To retrieve tumor tissue, euthanize mice by carbon dioxide inhalation. It is important not to perform cervical dislocation, a procedure that generates pressure to the posterior skull and can compromise tumor tissue integrity.
- Decapitation is performed immediately after death using a pair of scissors, removing hair and muscle tissue as much as possible for good visualization of the skull. Clean the surface of the skull with Kimwipe soaked with 95% ethanol.
- Use fine scissors to cut an opening along the midline of the skull, and remove skull tissue using fine tweezers, at which point the whole brain including tumor-bearing cerebellum is exposed.
- While the cerebella of healthy adults display well-defined hemispheres and vermis, the cerebella of tumor-bearing mice are often enlarged, amorphous with a smooth surface and conspicuous blood vessels. Using sterile techniques, retrieve the cerebellar tumor using tweezers and place in ice-cold PBS without Mg2+ and Ca2+.
- Dissociation of tumor tissue
- Transfer the tumor tissue from PBS to 50% Accutase (diluted in PBS) that is about 4 times the volume of the tumor tissue, mince the tissue with fine scissors for 3 minutes at room temperature, followed by incubation at 37°C for 4 minutes, after which the tissue undergoes repetitive pipeting with a 1-mL Pipetman for additional 3 minutes. This method should yield a mixture of single cells and small cellular aggregates.
- Dilute the cellular suspension 3-fold with PBS and centrifuge for 5 minutes at 1000 g to pellet the cells. Resuspend the cell pellet in fresh neural stem cell culture medium and plate onto a gelatinized 60 mm Primaria tissue culture dish. We use Primaria dishes for enhanced attachment at first plating; subsequent passages may be plated onto regular tissue culture dishes.
Note: Coat culture dishes with 0.1% gelatin for at least 30 minutes. Prepare fresh neural stem cell culture medium consisting of Neurobasal medium with glutamine, Pen-Strep, N2, B27, human EGF (25 ng/ mL) and basic FGF (25ng/ mL).
2. Enrichment, Maintenance and Expansion of Medulloblastoma Tumor Cells by Serial Passages
We usually get many colonies after 1 week of initial plating (Figure 1). These colonies can be dissociated and replated onto new gelatinized dishes for enrichment of the tumor cell population. First change to new culture medium, then simply use a 1-mL Pipetman to mechanically detach colonies followed by repetitive pipeting for 4 minutes to yield a cellular suspension (no Accutase needed). Check under a microscope to ensure that large cell clumps have been dissociated. Then dispense cell suspension onto new gelatinized dishes for further culturing and go through the same procedure for additional passages. Unattached tumor cells, blood cells and other cell types are removed by serial re-platings, leaving the attached tumor cells to expand rapidly. Change culture medium on the second day following initial seeding, and every other day thereafter.
3. Immunofluorescent Staining and Examination of Cultured Cells
1. Grow tumor cells on glass coverslips
On day 1, place one glass coverslip in each of a 6-well plate, then add 0.1% gelatin for 30 minutes at 37°C. Seed approximately 2-4X 105 tumor cells per mL. The cells should attach overnight and the neural stem cell medium changed on day 3. Staining is performed on day 4.
2 Immunofluorescent staining
Remove culture medium and wash cells twice with ice-cold PBS. Fix the cells with freshly-made 4%PFA for 15 minutes. Briefly wash cells twice with PBS and permeablize with 0.3% Triton (in PBS) for 5 minutes. Briefly wash cells twice with PBS and block with 10% goat serum for 40 minutes. Incubate the cells with primary antibody for 90 minutes, then wash cells three times with PBS, for 5 minutes each. Incubate the cells with secondary antibody for 60 minutes, then wash three times with PBS, 5 minutes each. Mount coverslips onto slides with 5 mL of an aqueous mounting medium and perform confocal microscopy.
4. Differentiation of Tumor Cells
Remove neural stem cell medium and briefly wash cells with PBS twice. Add differentiation medium on day 1, change medium on day 4 and determine level of differentiation by immunofluorescence staining on day 7. The differentiation medium consists of DMEM, Pen-Strep and 10% fetal bovine serum.
5. Representative Results
Morphology results
After Accutase treatment and gentle repetitive pipeting, the tumor tissue should be mostly dissociated into small cellular aggregates and single cells. Many sizable colonies can be observed as early as 5 days after initial plating, as shown by Figure 1. Highly proliferative tumor cells are often bi-polar with high nuclear/cytoplasmic ratio. These cells are seen typically radiating from small cellular aggregates attached to the gelatinized surface. Blood cells, small and round, are usually removed by subsequent media changes and serial platings. After several passages, one can observe uniformly distributed, proliferative Ki67+ tumor cells and little contamination with other cell types (Figure 2).
Expression of neural stem cell markers, clonal analysis and differentiation into multiple lineages
To determine whether the proliferative cells derived from dissociated cerebellar tumor tissue show characteristics of tumor stem cells, we performed further characterization by stem cell marker expression, clonal analysis and differentiation potency. We found that these isolated tumor cells highly express neural stem cell markers such as Sox2 and Nestin (Figure 3). As we retrieved tissues from SmoM2-YFP tumor, all tumor cells express YFP. Consistent with a recent study reporting that primary cilia are important for the development of Shh pathway-dependent medulloblastoma 12, SmoM2-YFP expression was clearly localized to the cilia of Sox2+ tumor cells (Figure 3). When plated at clonal density of 300 cells per mL of culture medium, we observed clonal expansion from a single tumor cell to a sizable colony (Figure 4). Furthermore, these cells were able to differentiate into various neuronal and glial cell types when cultured under pro-differentiation conditions (Figure 5).
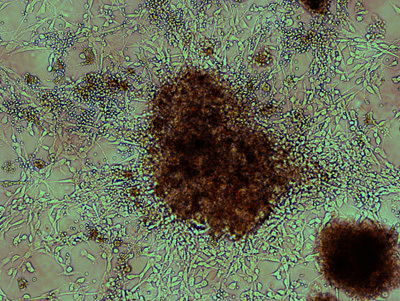 Figure 1. Morphology of a highly proliferative colony after first plating of dissociated medulloblastoma tissue. Generally, a sizable colony forms within 5 days after initial seeding of dissociated tumor tissue, and a representative one is illustrated here in bright field. Bi-polar, elongated cells radiate from a dense core cell aggregate. Attached to the tumor cells are small, round red blood cells, which are gradually depleted by serial platings.
Figure 1. Morphology of a highly proliferative colony after first plating of dissociated medulloblastoma tissue. Generally, a sizable colony forms within 5 days after initial seeding of dissociated tumor tissue, and a representative one is illustrated here in bright field. Bi-polar, elongated cells radiate from a dense core cell aggregate. Attached to the tumor cells are small, round red blood cells, which are gradually depleted by serial platings.
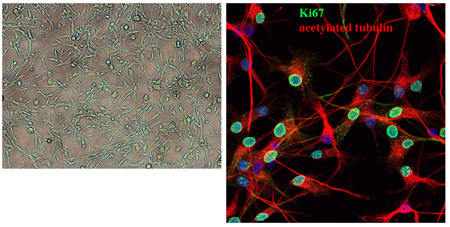 Figure 2. Morphology of established medulloblastoma cells beyond three passages. This bright field image shows the typical appearance of established medulloblastoma cells after several passages. The cells remain mostly bi-polar with high nuclear/cytoplasmic ratio. We show acetylated tubulin staining of tumor cells, most of which are cycling as shown by strong Ki67 expression.
Figure 2. Morphology of established medulloblastoma cells beyond three passages. This bright field image shows the typical appearance of established medulloblastoma cells after several passages. The cells remain mostly bi-polar with high nuclear/cytoplasmic ratio. We show acetylated tubulin staining of tumor cells, most of which are cycling as shown by strong Ki67 expression.
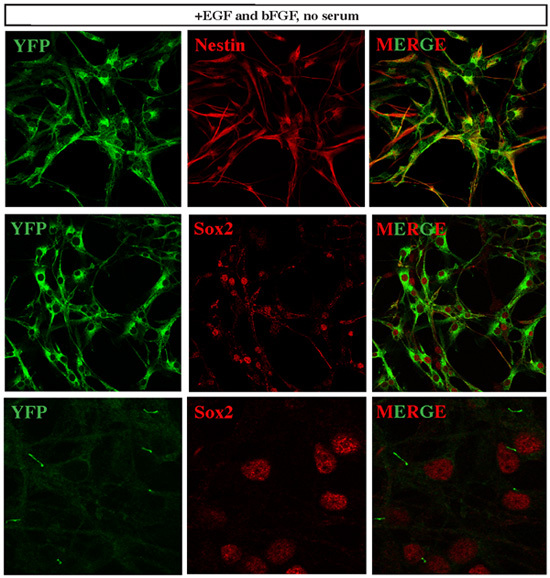 Figure 3. Established medulloblastoma cells express neural stem cell markers. SmoM2-YFP marks cells expressing constitutively active Smo, hence Shh pathway activity. All cells of the established medulloblastoma cell lines expressed YFP, and most of them co-expressed high levels of various neural stem cell markers, such as Nestin and Sox2. Interestingly, when we performed YFP signal detection with minimal laser power during confocal microscopy, we detected concentrated YFP signal in the cilia of Sox2+ cells. This observation is consistent with a recent report 12 that described the essential role of cilia in the development of Shh pathway-dependent medulloblastoma.
Figure 3. Established medulloblastoma cells express neural stem cell markers. SmoM2-YFP marks cells expressing constitutively active Smo, hence Shh pathway activity. All cells of the established medulloblastoma cell lines expressed YFP, and most of them co-expressed high levels of various neural stem cell markers, such as Nestin and Sox2. Interestingly, when we performed YFP signal detection with minimal laser power during confocal microscopy, we detected concentrated YFP signal in the cilia of Sox2+ cells. This observation is consistent with a recent report 12 that described the essential role of cilia in the development of Shh pathway-dependent medulloblastoma.
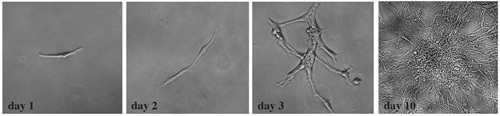 Figure 4. Established tumor cells undergo clonal expansion. After dissociation into single cell suspension, tumor cells were plated onto a 24 well-plate at a clonal density of 300 cells per mL of culture medium. In each well we observed clonally expanded colonies. This series of bright field images demonstrate typical changes over a 10-day period of culture.
Figure 4. Established tumor cells undergo clonal expansion. After dissociation into single cell suspension, tumor cells were plated onto a 24 well-plate at a clonal density of 300 cells per mL of culture medium. In each well we observed clonally expanded colonies. This series of bright field images demonstrate typical changes over a 10-day period of culture.
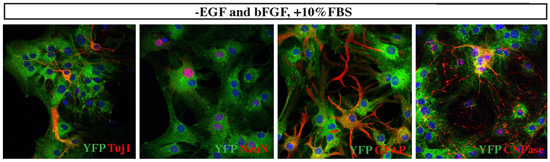 Figure 5. Differentiation analyses of established medulloblastoma cells. When tumor cells were switched from EGF/bFGF containing serum-free medium to DMEM/10% FBS, YFP+ tumor cells significantly altered their morphology and differentiated into various cell types, including Tuj1+ or NeuN+ neurons, GFAP+ astroglial cells or CNPase+ oligodendrocytes.
Figure 5. Differentiation analyses of established medulloblastoma cells. When tumor cells were switched from EGF/bFGF containing serum-free medium to DMEM/10% FBS, YFP+ tumor cells significantly altered their morphology and differentiated into various cell types, including Tuj1+ or NeuN+ neurons, GFAP+ astroglial cells or CNPase+ oligodendrocytes.
Discussion
We describe an efficient way to isolate, enrich and maintain medulloblastoma stem cells from primary tumor tissue retrieved from mutant mice with constitutively active Hedgehog signaling. We found that one critical step in successfully establishing a healthy tumor stem cell line is the Accutase treatment used during dissociation of the primary tumor tissue. In our experience, when first dissociating tumor tissue from the cerebellum, 4 minutes of 50% Accutase treatment at 37°C, followed by 3 minutes of repetitive pipeting using a 1-mL Pipetman will generally result in successful first seeding. It is also important to wait until sizable colonies are formed before re-plating (Figure 1). After the first seeding, Accutase treatment is no longer necessary; repetitive pipeting for 3-4 minutes using a 1-mL Pipetman is sufficient to dislodge and dissociate cell clumps for re-plating. Do not use smaller volume Pipetman as the finer tips will damage the cells extensively. An established cell line can be generated from each tumor tissue isolated and these cells are highly proliferative, robustly express several neural stem cell markers and can differentiate into both neuronal and glial cell types. We have performed orthotopic transplantation of these cells into immuno-compromised recipient mice which subsequently developed secondary medulloblastomas. These tumor cells in culture rarely exhibited spontaneous differentiation or apoptosis, features that are common to neurosphere culture methods. This protocol is designed for the successful propagation of tumor stem cells derived from primary medulloblastomas and thus, will enable us to test the effects of candidate genes and chemical inhibitors on Hedgehog-driven tumor cell growth and behavior.
Disclosures
No conflicts of interest declared.
Acknowledgments
This study was supported by grants from the Vanderbilt-Ingram Cancer Center Support Grant (P30 CA068485), the Childhood Brain Tumor Foundation and the National Institutes of Health (NS042205).
References
- Rossi A, Caracciolo V, Russo G. Medulloblastoma: From Molecular Pathology to Therapy. Clin Cancer Res. 2008;14(4):971–971. doi: 10.1158/1078-0432.CCR-07-2072. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wechsler-Reya RJ, Scott M. Control of Neuronal Precursor Proliferation in the Cerebellum by Sonic Hedgehog. P. Neuron. 1999;22(1):103–103. doi: 10.1016/s0896-6273(00)80682-0. [DOI] [PubMed] [Google Scholar]
- Dahmane N, Ruiz A, Altaba I. Sonic hedgehog regulates the growth and patterning of the cerebellum. Development. 1999;126(14):3089–3089. doi: 10.1242/dev.126.14.3089. [DOI] [PubMed] [Google Scholar]
- Wallace VA. Purkinje-cell-derived Sonic hedgehog regulates granule neuron precursor cell proliferation in the developing mouse cerebellum. Curr Biol. 1999;9(8):445–445. doi: 10.1016/s0960-9822(99)80195-x. [DOI] [PubMed] [Google Scholar]
- Fan X, Eberhart CG. Medullablastoma Stem Cells. J Clin Oncol. 2008;26(17):2821–2821. doi: 10.1200/JCO.2007.15.2264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoon JW, Gilbertson R, Iannaccone S. Defining a Role for Hedgehog Pathway Activation in Desmoplastic Medulloblastoma by Identifying GLI1 Target Genes. International journal of cancer. 2009;124(1):109–109. doi: 10.1002/ijc.23929. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O'Dorisio MS, Khanna G, Bushnell D. Combining anatomic and molecularly targeted imaging in the diagnosis and surveillance of embryonal tumors of the nervous and endocrine systems in children. Review. Cancer metastasis reviews. 2008;27(4):665–665. doi: 10.1007/s10555-008-9153-8. [DOI] [PubMed] [Google Scholar]
- Corcoran RB, Bachar Raveh T, Barakat MT. Insulin-like Growth Factor 2 Is Required for Progression to Advanced Medulloblastoma in patched1 Heterozygous Mice. Cancer research. 2008;68(21):8788–8788. doi: 10.1158/0008-5472.CAN-08-2135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gilbertson RJ, Ellison DW. The Origins of Medulloblastoma Subtypes. Annual review of pathology. 2008;3:341–341. doi: 10.1146/annurev.pathmechdis.3.121806.151518. [DOI] [PubMed] [Google Scholar]
- Ward RJ, Lee L, Graham K. Multipotent CD15+ Cancer Stem Cells in Patched-1 Deficient Mouse Medulloblastoma. Cancer research. 2009;69(11):4682–4682. doi: 10.1158/0008-5472.CAN-09-0342. [DOI] [PubMed] [Google Scholar]
- Mao J, Ligon KL, Rakhlin EY. A Novel Somatic Mouse Model to Survey Tumorigenic Potential Applied to the Hedgehog Pathway. Cancer research. 2006;66:10171–10171. doi: 10.1158/0008-5472.CAN-06-0657. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Han YG, Kim HJ, Dlugosz AA. Dual and opposing roles of primary cilia in medulloblastoma development. Nature medicine. 2009;15(9):1062–1062. doi: 10.1038/nm.2020. [DOI] [PMC free article] [PubMed] [Google Scholar]