

Abstract
Summary: The review summarizes the abundant information on the 35 identified peptidoglycan (PG) hydrolases of Escherichia coli classified into 12 distinct families, including mainly glycosidases, peptidases, and amidases. An attempt is also made to critically assess their functions in PG maturation, turnover, elongation, septation, and recycling as well as in cell autolysis. There is at least one hydrolytic activity for each bond linking PG components, and most hydrolase genes were identified. Few hydrolases appear to be individually essential. The crystal structures and reaction mechanisms of certain hydrolases having defined functions were investigated. However, our knowledge of the biochemical properties of most hydrolases still remains fragmentary, and that of their cellular functions remains elusive. Owing to redundancy, PG hydrolases far outnumber the enzymes of PG biosynthesis. The presence of the two sets of enzymes acting on the PG bonds raises the question of their functional correlations. It is difficult to understand why E. coli keeps such a large set of PG hydrolases. The subtle differences in substrate specificities between the isoenzymes of each family certainly reflect a variety of as-yet-unidentified physiological functions. Their study will be a far more difficult challenge than that of the steps of the PG biosynthesis pathway.
INTRODUCTION
Peptidoglycan (PG) (or murein) is a continuous covalent heteropolymer extending on the outside the cytoplasmic membrane of most eubacteria (194, 220, 232, 294). Its main functions are to preserve cell integrity by withstanding internal osmotic pressure and to maintain the shape of the bacterial cell. PG has been extensively studied owing to its importance as an essential structural cell wall component, its involvement in cellular morphogenesis, and the fact that steps of its biosynthesis are specific targets for well-known antibiotics and potential ones for novel antibacterials (71).
PG consists of linear glycan chains, which contain alternating β-1,4-linked units of GlcNAc and MurNAc-peptide and which are interconnected by the cross-linking of their peptide subunits (Fig. 1). Its biosynthesis is a two-stage process. First, the PG monomer unit is assembled in the form of a lipid intermediate by enzymes located in the cytoplasm or at the inner side of the cytoplasmic membrane (7, 26, 286, 288). In the second stage, extracytoplasmic glycosyltransferases catalyze the formation of the glycan chains using the lipid intermediate as a substrate (76, 229, 287). Finally, cross-linking between peptide subunits by transpeptidases leads to the PG network (77, 229).
Fig. 1.

Structure of E. coli peptidoglycan. (A) Glycan chain with monomer peptide subunits. (B) PG dimer with the d-Ala→(d)-meso-A2pm cross-linkage. (C) PG dimer with the (l)-meso-A2pm→(d)-meso-A2pm cross-linkage. (D) Tripeptide subunit with linked Lpp lipoprotein. Abbreviations: G, N-acetyl-β-d-glucosamine (GlcNAc); M, N-acetyl-β-d-muramic acid (MurNAc); red C, carbon of the d-configuration; →, CO-NH linkage.
Besides the enzymes of the PG biosynthesis pathway, eubacteria generally harbor a more or less important set of endogenous hydrolases that specifically cleave PG linkages (238, 295). A hydrolytic activity has been identified for each bond linking the PG amino acid and sugar components. It has long been speculated that PG hydrolases are involved in many different aspects of PG metabolism and bacterial cell physiology (238, 240, 295). In particular, the uncontrolled action of PG hydrolases operating as autolysins plays a major role in the bacterial autolysis observable under various conditions (215).
The discovery of PG hydrolases in Escherichia coli paralleled the structural characterization of its cell wall PG (304). Initially, they were elicited by incubating hydrolase-containing cell extracts with PG muropeptides and analyzing the reaction products (198, 199). Later, isolated radiolabeled PG (sacculi) was used as substrate (89). Another approach entailed the use of the zymogram method initially developed for Gram-positive organisms (137) and later adapted to Gram-negative organisms (16). Nine PG hydrolases from E. coli were visualized in this way. Finally, sequence comparisons by database analysis became a powerful way to identify new hydrolases. Data on the E. coli PG hydrolases have concerned mainly but not exclusively strain K-12.
In E. coli, at least 35 PG hydrolases have been identified and classified into 12 distinct families (Table 1). As in many other eubacteria, they include mainly glycosidases, peptidases, and N-acetylmuramoyl-l-alanine amidases. So far, only the PG hydrolases of Bacillus subtilis have been studied to a similar extent (240). The multiplicity and redundancy of the E. coli hydrolases raise the question of their respective functions in various aspects of PG metabolism. Work up to 1995 has been reviewed (99, 100, 102). More recent reviews have concerned specific hydrolase families (72, 73, 92, 103, 185, 196, 230, 295).
Table 1.
E. coli peptidoglycan hydrolasesa
| Hydrolase | Gene | Localization | 3D structure reference | Substrate(s) |
|---|---|---|---|---|
| β-N-Acetylglucosaminidase NagZ | nagZ | C | SC + PG | |
| Lytic transglycosylases | ||||
| Slt70 | slt | P | 257 | PG |
| MltA | mltA | OM | 289 | SC + PG |
| MltB | mltB | OM | 277 | SC + PG |
| MltC | mltC | OM | PG | |
| MltD | mltD | OM | 9 | PG |
| MltE | mltE | OM | 3 | SC + PG |
| MltF | yfhD | OM | PG | |
| dd-Carboxypeptidases | ||||
| PBP4b | yfeW | CM | SC | |
| PBP5 | dacA | CM | 46 | SC + PG |
| PBP6 | dacC | CM | 36 | SC |
| PBP6b | dacD | CM | PG | |
| dd-Endopeptidases | ||||
| PBP7 | pbpG | P | PG | |
| MepA | mepA | P | 161 | SC + PG |
| EnvC | envC | CM/P | ||
| dd-Peptidases | ||||
| PBP4 | dacB | P | 125 | SC + PG |
| AmpH | ampH | CM | SC + PG | |
| ld-Carboxypeptidases | ||||
| LdcA | ldcA | C | SC | |
| 12 kDa | SC | |||
| 86 kDa | P | SC | ||
| ld-Transpeptidases | ||||
| ErfK | erfK | P | PG | |
| YbiS | ybiS | P | PG | |
| YcbB | ycbB | P | PG | |
| YcfS | ycfS | P | PG | |
| YnhG | ynhG | P | PG | |
| ld-Endopeptidase, 34 kDa | SC | |||
| Endoamidase MpaA | mpaA | C | SC | |
| MurNAc-l-Ala amidases | ||||
| AmiA | amiA | P | PG | |
| AmiB | amiB | P | PG | |
| AmiC | amiC | P | PG | |
| AmpD | ampD | C | 147 | SC |
| AmiD | amiD | OM | 124 | SC + PG |
| 39 kDa | P | SC | ||
| l-Ala-d/l-Glu epimerase YcjG | ycjG | C | 84 | SC |
| d-Ala-d-Ala dipeptidase DdpX | ddpX | C | 144 | SC |
dd-Peptidases show both dd-carboxypeptidase and dd-endopeptidase activities. C, cytoplasm; CM, cytoplasmic membrane; P, periplasm; OM, outer membrane; SC, soluble component (precursor, muropeptide, peptide, or glycan chain); PG, isolated PG.
The aim of the present work is to bring together the past and present abundant information on the PG hydrolases of E. coli and to attempt a critical assessment of their functions in PG maturation, turnover, elongation, septation, and recycling as well as in cell autolysis. Except for a few examples, comparisons with the abundant literature concerning PG hydrolases of other organisms were not attempted. The present review should be helpful for developing future comparative studies.
E. COLI PEPTIDOGLYCAN GLYCOSIDASES
In E. coli, several glycosidases cleave the GlcNAc-(1→4)-MurNAc (β-N-acetylglucosaminidase) or MurNAc-(1→4)- GlcNAc (N-acetylmuramidase) bonds of PG glycan chains (Fig. 2). In contrast with other bacterial organisms, no N-deacetylases acting on the GlcNAc or MurNAc units have been reported for E. coli (296). However, an N-acetylglucosamine-6-phosphate deacetylase acting on GlcNAc-6-phosphate (306) and a d-lactyletherase acting on MurNAc-6-phosphate (119) were characterized.
Fig. 2.

Sites of action of hydrolases in PG, precursor, and muropeptide.
N-Acetylglucosaminidase
A β-N-acetylglucosaminidase removing GlcNAc from the disaccharide-tripeptide muropeptide (Fig. 2) was initially identified in a cell extract from E. coli (198). Using monomer and dimer muropeptides or p-nitrophenyl-β-N-acetylglucosaminide as a substrate, it was purified as a cytoplasmic enzyme (155, 310). An activity-deficient mutant showed normal growth and morphology (311). Later, the encoding nagZ gene was identified (37), and an overproduced recombinant form was purified (298). A null mutant completely lacked NagZ activity, thereby establishing that NagZ was the only β-N-acetylglucosaminidase present in E. coli (37). The findings that this mutant grew normally in rich medium and had normal morphology proved that NagZ is not essential for cell growth. However, it is required for the PG recycling process (196). NagZ is specific for the β linkage and is active on monomer and dimer muropeptides (155, 310) but not on the tetrasaccharide muropeptide (155). It cleaves the GlcNAc-(1→4)-1,6-anhydro-MurNAc disaccharide (271). Its cytoplasmic location excludes any in vivo effect on PG, but NagZ releases GlcNAc from isolated radiolabeled PG, presumably by acting as an exoenzyme at the nonreducing end of the glycan chains (310).
Lytic Transglycosylases
Lytic transglycosylases (LTs) are N-acetylmuramidases that not only cleave the β-1,4 glycosidic bond between the MurNAc and GlcNAc units but also concomitantly catalyze an intramolecular glycosyl transfer reaction whereby a 1,6-anhydro bond is formed between C1 and C6 of N-acetylmuramic acid (Fig. 2 and see Fig. 4). The LT activity producing 1,6-anhydro- MurNAc muropeptides was initially characterized with phage λ and phage ViII endolysins (253, 254). The first E. coli LT was designated Slt70, for soluble lytic transglycosylase (98). Subsequently, six other identified E. coli LTs were designated MltA, MltB, MltC, MltD, MltE, and MltF, for membrane-bound lytic transglycosylases (Table 1). LTs were found in many other organisms (230, 295) and were classified into four families on the basis of consensus sequence motifs (23). Slt70, MltC, MltD, and MltE belong to family 1, which are homologous to goose-type lysozyme, whereas MltA and MltB are classified as belonging to families 2 and 3, respectively. Phage λ endolysin belongs to the fourth LT family.
Fig. 4.
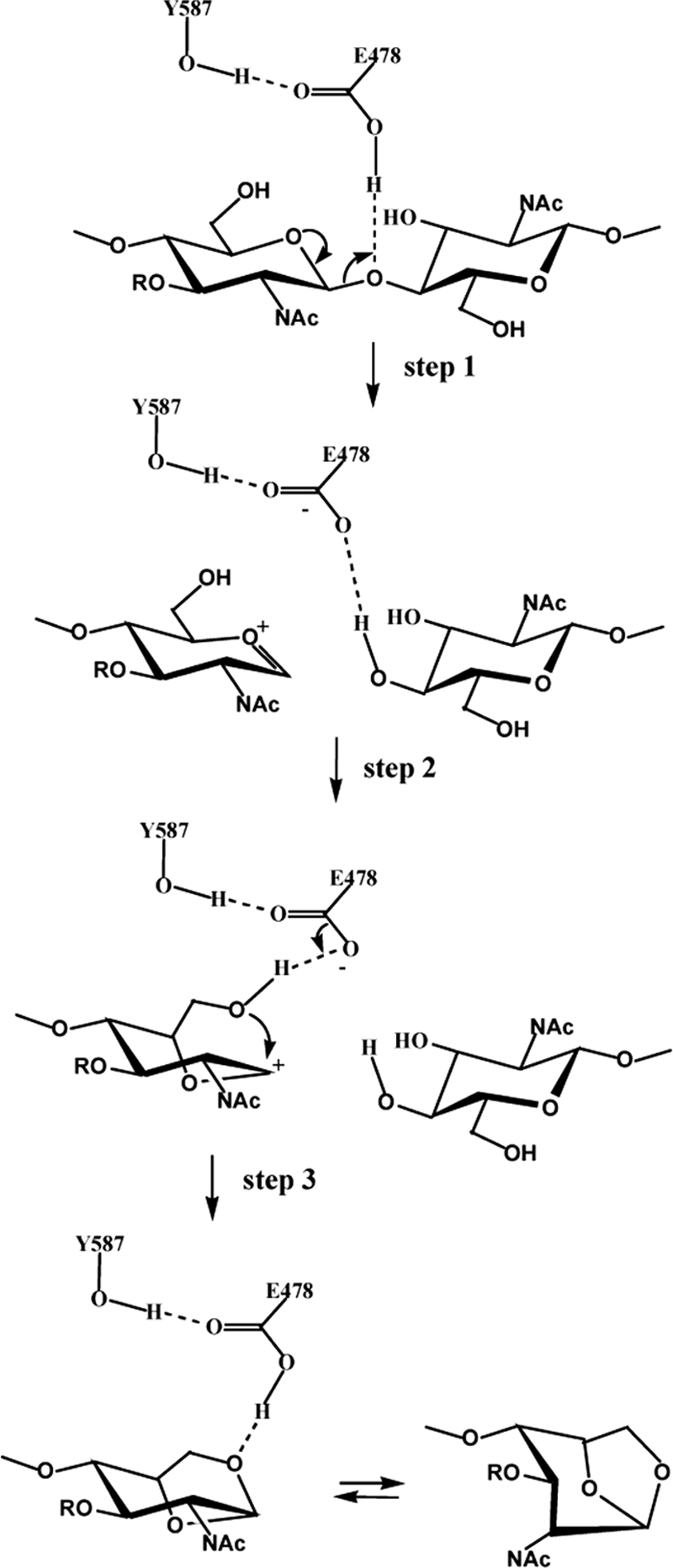
Reaction mechanism of the Slt70 LT proposed by Thunnissen et al. (259). R, d-lactoyl peptide. Step 1 is the cleavage of the MurNAc-(1→4)-GlcNAc bond with the formation of an oxocarbonium ion intermediate. Step 2 is an intramolecular nucleophilic attack on the C1 carbon of the oxocarbonium ion. Step 3 is the extraction of a proton by catalytic Glu478 and the formation of the 1,6-anhydro-MurNAc-peptide.
Slt70 lytic transglycosylase.
Slt70 was identified in the soluble fraction of disrupted cells by its action on isolated diaminopimelic acid (A2pm)-radiolabeled PG yielding 1,6-anhydro-MurNAc muropeptides and was purified (98, 135). By immunoelectron microscopy, it was shown to be localized mainly in the cell wall, with only 20 to 30% remaining in the cytoplasm. Tight binding to the outer surface of isolated sacculi was demonstrated as well (299). The sequence of the structural slt gene (21) codes for a polypeptide with a typical signal sequence cleaved off during maturation, confirming a periplasmic localization (66). The deletion of slt led to no changes in growth, cell morphology, or PG muropeptide composition but had an effect on the response toward various β-lactams (255). Similarly, bulgecin A, which reversibly and noncompetitively inhibits isolated Slt70, also modified the response to β-lactams. Growth inhibition without extensive lysis was observed with overproduced Slt70 (66). However, the inhibition of protein synthesis induced the rapid autolysis of the overproducing strains (21). Affinity chromatography studies (223, 297) revealed protein-protein interactions between Slt70 and different penicillin-binding proteins (PBPs) (PBP2, PBP3, and PBP7/8), suggesting a possible participation of Slt70 in multienzyme complexes.
The structures of native Slt70 and its complex with a 1,6-anhydro-MurNAc muropeptide were solved at 1.65-Å and 1.90-Å resolutions, respectively (257, 258, 276). Slt70 has α-helices assembled into three distinct domains, generating a doughnut-like shape (Fig. 3a). The N-terminal and central domains form a closed ring with a large central hole. The catalytic site is located in the C-terminal domain and shows close structural similarity to that of goose-type lysozyme (258). It has a bilobal structure with a long substrate-binding groove and a catalytic acid/base glutamic acid. The crystal structure of Slt70 complexed with a 1,6-anhydro-MurNAc muropeptide showed that the recognition site for the peptide moiety is located at the interface of the three domains (276). The structure of Slt70 complexed with bulgecin A was determined at a 2.8-Å resolution and revealed the binding of one inhibitor molecule to the active site of the enzyme (259).
Fig. 3.

Three-dimensional structures of 10 E. coli PG hydrolases. (a) Slt70. (Reprinted from reference 276 with permission of Elsevier.) (b) MltA. (Reprinted from reference 289 with permission of Elsevier.) (c) MltB. (Reprinted from reference 277 with permission of Elsevier.) (d) MltE. (Reprinted from reference 3 with permission from the publisher. Copyright 2011 American Chemical Society.) (e) PBP4. (Reprinted from reference 125 with permission from the publisher. Copyright 2006 American Chemical Society.) (f) PBP5. (Reprinted from reference 46 with permission of the publisher.) (g) PBP6. (Reprinted from reference 36 with permission from the publisher. Copyright 2009 American Chemical Society.) (h) MepA. (Reprinted from reference 161 with permission of the publisher.) (i) YcjG epimerase. (Reprinted from reference 84 with permission of the publisher. Copyright 2001 American Chemical Society.) (j) AmiD dimer. (Reprinted from reference 124 with permission of Elsevier.)
MltA lytic transglycosylase.
In an slt deletion mutant, an LT was detected in the membrane fraction, solubilized with detergents, and purified (221, 275). The membrane enzyme differed from Slt70 by accepting soluble peptide-free poly-[GlcNAc-(1→4)-MurNAc] glycan chains as substrate besides cross-linked PG (221, 275). Initially referred to as Mlt38 (275), it was redesignated MltA (64), and its encoding gene was identified (149). Surprisingly, MltA shares no region of sequence similarity with Slt70, MltB, or MltC. It contains a typical lipoprotein consensus sequence. Its lipoprotein nature was demonstrated by [3H]palmitate labeling, and its localization in the outer membrane was established by membrane fractionation. In an MltA deletion mutant, growth was unaffected, and no change of morphology was observed (149). In contrast, the overproduction of MltA resulted in the formation of spheroplasts and lysis at 30°C but not at 37°C, which is consistent with the purified enzyme having a temperature optimum at 30°C (149). Affinity chromatography experiments using purified proteins revealed protein-protein interactions with PBPs and periplasmic proteins (293). In particular, the periplasmic protein MipA mediates the MltA-PBP1b interaction, suggesting the participation of MltA in a PG-synthesizing complex.
The crystal structure of a functional soluble form of MltA lacking its membrane anchor was determined at a 2.0-Å resolution (289). It revealed two domains separated by a large groove (Fig. 3b), which can accommodate a six-residue-long glycan chain and which has a centrally located aspartic acid residue conserved in all known MltAs (206, 289). This first structure of a member of LT family 2 showed a fold different from those of the other known LTs. While the latter proteins share a catalytic core with a lysozyme-like fold, the catalytic domain of MltA contains a double-psi β-barrel fold similar to that of members of the barwin-like endoglucanase superfamily.
MltB lytic transglycosylase.
Soluble LT activity was also detected in a strain carrying a chromosomal slt deletion. The LT was purified as a fully soluble 35-kDa enzyme distinct from Slt70 and was named Slt35 (67). The identified encoding gene was designated mltB (56, 64). The coding region displayed a consensus lipoprotein signal at the N-terminal end, and the presumed prolipoprotein form accumulated in the presence of globomycin (64). The soluble LT activity associated with MltB-overproducing cells increased with time as the result of proteolytic degradation. The lipoprotein character of MltB was proven by [3H]palmitate labeling, and the localization of the enzyme at the outer membrane was demonstrated (64). Two overproduced soluble forms of MltB were fully active (64, 249). In particular, one form was able to cleave the middle MurNAc-(1→4)-GlcNAc bond of a synthetic soluble tetrasaccharide muropeptide, yielding disaccharide muropeptides (249). An MltB deletion mutant showed no significant change in growth or in PG muropeptide composition, whereas the overproduction of MltB led to spherical cells and subsequent lysis (64).
The structure of the Slt35 soluble form of MltB was determined at a 1.7-Å resolution (277, 278) and revealed an ellipsoid molecule with three domains, called α, β, and core (Fig. 3c). The fold of the core domain resembles that of lysozyme and that of the catalytic domain of Slt70. The active site is located in a deep groove formed by residues from the α and core domains and has a catalytic acid/base glutamic acid. A study of Slt35 complexed with chitobiose, the muropeptide GlcNAc-(1→4)-MurNAc-l-Ala-d-Glu, or bulgecin A revealed the location of four sugar-binding sites and two peptide-binding sites, all close to the catalytic glutamic acid residue (278).
MltC lytic transglycosylase.
The search for LTs in the Haemophilus influenzae genome revealed an open reading frame having slt-derived fingerprints and a strong homology with an E. coli open reading frame at centisome 66.8 containing a putative consensus lipoprotein signal sequence (57). The newly discovered E. coli LT was designated MltC, and three forms were overproduced without lysis and were analyzed by zymography. The full-length sequence with the putative lipoprotein signal sequence as well as the one without it showed significant PG hydrolase activity. The form with only the C-terminal domain also had a high level of activity. All forms produced 1,6-anhydro-MurNAc muropeptides from isolated PG (57).
MltD lytic transglycosylase.
The sequence of the E. coli yafG open reading frame at centisome 5.1 contains the three slt fingerprints indicative of muramidase activity (57), and the gene was designated mltD (58). The N-terminal end of MltD displayed a lipoprotein signal and an LT domain, whereas the C-terminal end had two repeats of the LysM PG-binding motif (9, 58). The LysM domain is a widespread protein module, and there is evidence suggesting that it is a general PG-binding module. Often encountered in cell wall-degrading enzymes, it probably acts to anchor the catalytic domain to PG (9). The PG hydrolase activities of full-length MltD and of a recombinant form lacking the N-terminal signal peptide and the membrane-anchoring residues were revealed by zymography (58, 249). The overproduction of MltD led to spheroplast formation and lysis (58).
Structural studies of MltD have been limited to its C-terminal domain containing the two LysM repeats (9). The peptide encompassing the two motifs was overproduced, and assignments were obtained by using a range of two-dimensional (2D) and three-dimensional (3D) heteronuclear nuclear magnetic resonance (NMR) experiments. The LysM domain has a βααβ secondary structure, with the two helices packing onto the same side of an antiparallel β sheet. The structure showed no similarity to other bacterial cell surface domains. A shallow groove on the protein surface was identified as a potential binding site.
MltE lytic transglycosylase.
An LT accepting unsubstituted PG glycan chains as a substrate was detected in membrane extracts from an mltA mutant and was named MltE. Partial purification led to a 22-kDa protein (132). In a second step, an open reading frame at centisome 26.7 was identified by using a motif common to Slt70 and MltC. Initially named emtA, for endo-type membrane-bound lytic transglycosylase (132), it was later redesignated mltE (92, 249). Its coding region started with a signal peptide followed by a lipoprotein-processing site. It was cloned, and the lipoprotein character of the mltE-encoded protein was demonstrated by [3H]palmitate labeling (132). The PG hydrolase activity of MltE on isolated PG was revealed by zymography (132, 249). However, no products were released when isolated radiolabeled PG was used as a substrate (132). Membranes from cells overproducing MltE degraded isolated PG glycan chains having seven disaccharide units (132), and the specificity of an endotransglycosylase activity was revealed by the cleavage of internal glycosidic bonds, leading to shorter chains with 1,6-anhydro-MurNAc ends. The overproduction of MltE led to a shortening of the PG glycan chains and to an increased (l)-meso-A2pm→(d)-meso-A2pm cross-linkage content. These changes occurred without rapid cell lysis (132).
The crystal structure of MltE was determined at a 2.0-Å resolution (3) and revealed a two-lobe ellipsoid shape (Fig. 3d). Its overall fold contains 10 α-helices and resembles that of goose-type lysozyme. The active site containing eight saccharide-binding subsites is sequestered in a deep grove spanning the two lobes and is similar to that of Slt70. The absence of a peptide-binding subsite, which allows processivity in the functioning of exolytic Slt70, explains the endolytic specificity of MltE.
MltF lytic transglycosylase.
A potential LT was recognized in the sequence of the yfhD open reading frame of E. coli containing the three slt fingerprints indicative of muramidase activity (57). Based on sequence alignments and the presence of consensus motifs, the hypothetical YfhD protein was the archetype of LT family 1B (23). It had a predicted cleavable signal sequence and a transmembrane helix. Experimental proof confirmed its LT activity, and it was named MltF (231). Full-length overproduced MltF was associated with the outer membrane (231). An overproduced soluble truncated form lacking the N-terminal signal sequence and the membrane anchor catalyzed the release of 1,6-anhydro-MurNAc muropeptides from isolated PG. The deletion of mltF led to no change in cell growth or morphology, whereas the overproduction of MltF had a lytic effect.
MltF appeared to be a bimodular structure composed of an N-terminal non-LT domain and a C-terminal LT domain (231). To confirm this modular structure, the mltF gene was engineered to encode each domain separately. The overproduced C-terminal domain retained full LT activity, whereas the N-terminal domain was devoid of PG-binding capacity and LT activity. A prediction of the 3D structures of the N-terminal and C-terminal domains was made by using a protein fold recognition server.
Enzymatic properties of E. coli LTs.
The seven identified E. coli LTs are all associated with the cell envelope and are presumably located close to their PG substrates. However, Slt70 is tightly bound to PG, whereas the six other LTs are bound to the outer membrane as lipoproteins (MltA to MltE) or by a membrane anchor (MltF). The seven LTs are active on isolated PG, but the release of 1,6-anhydro-disaccharide-peptide units was established only with Slt70, MltA, MltB, and MltF (67, 98, 231, 275). These four LTs are thus exoenzymes processively degrading PG glycan chains at one end. Unfortunately, it is not clear at which end degradation proceeds. The crystal structure of Slt70 suggests that binding occurs at the 1,6-anhydro-MurNAc end (276), whereas the kinetics of the release of muropeptides from PG with specifically radiolabeled GlcNAc ends show that degradation proceeds at the nonreducing end of the glycan chain (12). Among the other specificity particularities, MltA accepts the peptide-free poly- (GlcNAc-1,4-MurNAc) glycan chains as substrate (275), and MltB is active on a short synthetic glycan fragment (249). MltE acts only with endo activity, cleaving internal MurNAc-(1→4)-GlcNAc bonds with the formation of shorter chains (132). The functioning of MltC as an LT is based on the release of 1,6-anhydro-MurNAc-containing muropeptides (57), whereas that of MltD is based on sequence comparisons with other LTs (58). It should be stressed that the use of isolated PG as an insoluble substrate has not favored extensive studies of LT enzymatic properties.
Analysis of the crystal structures of Slt70, MltA, MltB, and MltE as well as inhibition studies led to the same two-step reaction mechanism (Fig. 4) valid for different LTs (3, 206, 213, 221, 230, 255, 258, 259, 276, 277, 278, 289, 290). The first step entails the cleavage of the glycosidic bond between MurNAc and GlcNAc by the action of a glutamic or aspartic acid residue functioning as a single-acid catalyst donating a proton. This results in the formation of an oxocarbenium ion-like transition state stabilized by the formation of an oxazolinium intermediate. In the second step of the reaction, the deprotonated catalytic residue acts as a general base to abstract a proton from the OH on C6 of MurNAc, thereby allowing for an intramolecular nucleophilic attack at C1, collapsing the oxazolinium intermediate with the concomitant formation of the 1,6-anhydro-MurNAc-peptide (230). Experimental evidence for this reaction mechanism was obtained by studies of the inhibitory effect of bulgecin A and N-acetylglucosamine thiazoline functioning on LTs as structural analogs of the putative oxazolinium intermediate (213, 221, 255, 258, 259). Site-directed mutagenesis confirmed the crucial role played by the glutamic or aspartic acid residue in the mechanism (213, 276, 289).
E. COLI PEPTIDOGLYCAN PEPTIDASES
In E. coli, there are at least 20 peptidases (Table 1) cleaving peptide bonds in isolated PG, muropeptides, or precursors (Fig. 1 and 2). Their high enzymatic specificities are due to the presence of nonprotein amino acids (d-Ala, d-Glu, and meso-A2pm) in the peptide subunits and to the unusual stereochemistry [dd, dl, ld, and γ-d-Glu-(l)-meso-A2pm] of the peptide bonds (Fig. 1). The E. coli PG peptidases are either penicillin-binding or penicillin-insensitive enzymes.
Penicillin-Binding Peptidases
Penicillin-binding proteins (PBPs) are a family of bacterial acyl-serine transferases of common evolutionary origin specifically inhibited by the covalent binding of β-lactam antibiotics to their active sites (77, 156, 229, 312). E. coli PBPs are visualized by polyacrylamide gel electrophoresis after the treatment of cells with radiolabeled or fluorescence-labeled β-lactams and are designated according to their electrophoretic migrations (6, 49, 72, 95, 103, 186, 243, 291, 315). Most PBPs are anchored to the cytoplasmic membrane with their active site in the periplasm (73). The five high-molecular-mass (HMM) PBPs (PBP1a, PBP1b, PBP1c, PBP2, and PBP3) catalyze the extracytoplasmic reactions of PG polymerization and are devoid of hydrolytic activity (76, 229). Among the eight low-molecular-mass (LMM) PBPs (PBP4, PBP4b, PBP5, PBP6, PBP6b, PBP7/8, AmpC, and AmpH), only the β-lactamase AmpC has no recognized dd-peptidase activity (95). The E. coli LMM PBPs belong to four subcategories of the class C subdivision of the PBP classification (229): type 4 (PBP4), type 5 (PBP5, PBP6, and PBP6b), type 7 (PBP7/8), and type AmpH (PBP4b and AmpH). The description of the LMM PBPs will follow the order of increasing electrophoretic mobility.
dd-Peptidase PBP4.
The PBP4 amino acid sequence encoded by the dacB gene is similar to those of class C β-lactamases and other LMM PBPs (130, 181). PBP4 was purified from wild-type and overproducing strains (39, 130, 146, 182). The dacB gene encodes a signal peptide, which directs the protein to the periplasm and which is lacking in purified PBP4. Isolated without detergents, PBP4 behaves as a soluble protein. Its primary structure reveals no evidence of a transmembrane helix, an amphiphilic membrane anchor, or a lipid anchor (181). Nevertheless, PBP4 is associated with the membrane, as established by its presence in inner membrane vesicles (117, 143) and its purification from the membrane fraction (130). This behavior supports the view that PBP4 is bound to some membrane protein. Affinity chromatography experiments suggested a possible interaction with MltB (297).
PBP4 shows dd-endopeptidase activity (Fig. 2) by hydrolyzing the d-Ala→(d)-meso-A2pm bond in the soluble dimer muropeptide (130) and in the dimer units of isolated PG (39, 130, 146). PBP4 is identical to the soluble and periplasmic forms of the d-alanine carboxypeptidase IB activity, cleaving the d-Ala→d-Ala bond of the UDP-MurNAc-pentapeptide precursor and the cross-linkage of the dimer muropeptide (111, 165, 252). PBP4 thus exhibits both dd-carboxypeptidase and dd-endopeptidase activities (Fig. 2).
The high-resolution (1.6-Å) structures of PBP4 and covalent complexes with different antibiotics are related to those of other PBPs and β-lactamases (125). The overall structure shows three distinct domains, with the C terminus ending close to the N terminus in domain 1 (Fig. 3e). More recently, a mutational analysis of the substrate specificity of PBP4 revealed the importance of a subsite in accommodating the meso-A2pm residue of the UDP-MurNAc-pentapeptide substrate (39).
The loss of PBP4 fails to induce growth defects or detectable morphological alterations (49, 165, 170). The changes in PG structure accompanying PBP4 overproduction indicated that PBP4 acts in vivo as a dd-endopeptidase and a dd-carboxypeptidase but not as a dd-transpeptidase (130). Moreover, the decrease in d-Ala→(d)-meso-A2pm cross-linkage contents was partially compensated for by an increase in the (l)-meso-A2pm→(d)-meso-A2pm cross-linkage content, possibly explaining the tolerance of E. coli for high-level PBP4 overproduction.
dd-Carboxypeptidase PBP4b.
PBP4b was recognized as an LMM PBP dd-carboxypeptidase by a study of E. coli open reading frame yfeW, encoding a 51-kDa polypeptide which had the three PBP motifs and strong homology with AmpH (291). Its N-terminal signal peptide serving for the anchoring to the cytoplasmic membrane had no putative signal peptidase sequence. An overproducing strain showed binding to radiolabeled β-lactam, as visualized by polyacrylamide gel electrophoresis. Overproduced PBP4b displayed dd-carboxypeptidase activity with Nα,Nε-diacetyl-l-lysyl-d-alanyl-d-alanine. Gene inactivation demonstrated the nonessential character of PBP4b and led to viable mutants with unchanged morphologies. Interestingly, the yfeW gene encoding PBP4b is in an operon together with the murP and murQ genes, which are required for growth on MurNAc (118).
dd-Carboxypeptidase PBP5.
As the major PBP of E. coli (60, 243), PBP5 has been extensively studied (72, 205, 280, 301). It was identified with d-alanine carboxypeptidase IA activity encoded by the dacA gene (30, 166, 191, 242). Its primary structure shows sequence similarity with PBP4, PBP6, and β-lactamases (30, 163). Synthesized by the removal of a hydrophobic N-terminal signal sequence, PBP5 localizes in the cytoplasmic membrane, with the bulk of the protein extending into the periplasm (208, 209). The 18-amino-acid-long amphiphilic helix at the C-terminal end is essential for tight anchoring to the membrane (114, 115), and the mechanism of this attachment has been discussed (187).
Initially, PBP5 was purified to homogeneity from wild-type E. coli by affinity chromatography (1). Various N- and C-terminally truncated forms were produced (68, 205, 280). The soluble and membrane-bound forms of PBP5 recognize the same range of substrates. They display a broad substrate specificity by catalyzing the cleavage of the C-terminal d-Ala→d-Ala bond (Fig. 2) in many soluble substrates (UDP-MurNAc-pentapeptide, lipid II, muropentapeptides from different sources, pentapeptides, and synthetic compounds) and in un-cross-linked and cross-linked PG (1, 188, 205, 246, 281). In cross-linked PG, PBP5 recognizes d-Ala→d-Ala bonds in both the monomer and dimer subunits (205).
The crystal structure of a soluble form of PBP5 (Fig. 3f) revealed two distinct, roughly perpendicular domains (46, 189). The active site located in domain 1 showed high structural similarity with those of other PBPs and β-lactamases. Domain 2, which has a hydrophobic surface, is of unknown function but is essential for stabilizing the protein (280) and for the proper in vivo functioning of PBP5 (186).
PBP5-defective mutants (166, 191, 244) show normal cell growth and an increased pentapeptide subunit content (50, 134, 146, 226). This finding indicates that PBP5 is acting in vivo on PG. Morphological defects accompanying the loss of PBP5 are visible by microscopic inspection (185), and shape changes with slight abnormalities at the poles are detectable by fluorescence-activated cell sorting (170). The overproduction of PBP5 is lethal, causing E. coli to grow as spherical cells before lysing (162, 247). The proposed function of PBP5 is to regulate the availability of pentapeptide subunits for the formation of the cross-linkages by transpeptidation (48, 205).
dd-Carboxypeptidase PBP6.
PBP6, encoded by dacC, has a high level of sequence homology with PBP5 (30). The cleavage of the signal sequence yields a mature protein anchored to the cytoplasmic membrane by its C-terminal amphiphilic helix, with the bulk of the protein extending into the periplasm (73, 208). PBP6 was purified to homogeneity at the same time and in the same way as PBP5 (1, 2). With UDP-MurNAc-pentapeptide as a substrate, it shows a much lower level of dd-carboxypeptidase activity than PBP5. A later report asserted that both native and overproduced soluble forms of PBP6 have no detectable dd-carboxypeptidase activity with the nucleotide substrate (279). However, a soluble form of PBP6 cleaves the d-Ala→d-Ala bond of MurNAc-pentapeptide (36).
The crystal structure of PBP6 lacking its signal peptide and membrane anchor was determined at a 2.1-Å resolution, while the structures of complexes with ampicillin and MurNAc-pentapeptide were determined at a 1.8 Å-resolution (36). Like PBP5, PBP6 has two perpendicular domains: a large N-terminal domain harboring the active site and a smaller β-sheet-rich C-terminal domain (Fig. 3g).
The deletion of PBP6 has no effect on cell growth (28, 49) and, unlike PBP5, leads to no increase of the PG pentapeptide subunit content (226). The overproduction of the native or soluble form is not bacteriolytic (279). The expression of dacC mRNA is far more dependent on the BolA transcriptional regulator than that of dacA mRNA (226). Despite homology, the marked differences between PBP5 and PBP6 point to distinct biological functions. Owing to its unusually high level in stationary-phase cells (31), PBP6 could play a role in the stabilization of PG at the end of the growth phase (279).
dd-Carboxypeptidase PBP6b.
PBP6b was identified by a study of an E. coli open reading frame showing the capacity to encode a polypeptide with a predicted amino acid sequence strikingly similar to those of PBP5 and PBP6 (6). The gene was designated dacD. Its product bound penicillin and showed dd-carboxypeptidase activity in vivo on PG pentapeptide subunits. As with PBP5 and PBP6, PBP6b is associated with the cytoplasmic membrane by its C-terminal end, which has the capacity to form the amphiphilic α-helical secondary structure required for anchoring to the membrane (73). The dacD gene is expressed at a very low level, and PBP6b is not essential for cell growth (6).
dd-Peptidase AmpH.
A PBP different from the previously known ones was observed in E. coli cells infected with λ phages from the Kohara genomic library (95). After cloning, the established sequence was identical to that of the open reading frame yaiH at centisome 8.5. Owing to its relationship to other amp genes of the class C family, the gene was named ampH. In wild-type E. coli, ampH is expressed at a low level, and its deletion leads to no morphological alterations. It is nonessential under normal laboratory conditions. However, morphological alterations are observable when an ampH deletion is associated with PBP1a and PBP5 deficiencies, indicating a role in PG metabolism (95).
Initially, AmpH had no recognized enzymatic activity (95). The recently reported cloning and purification of overproduced forms have enabled a better characterization of its enzymatic specificities (79). Like the other PBPs, AmpH shows a very low level of β-lactamase activity. On the other hand, it displays a wide spectrum of high-level dd-peptidase activities on monomer, dimer, and oligomer muropeptides as well as on isolated PG (Fig. 2). AmpH dd-carboxypeptidase activity hydrolyzes disaccharide-pentapeptide rapidly but the synthetic Nα,Nε-diacetyl-l-lysyl-d-alanyl-d-alanine tripeptide slowly. Like PBP4, AmpH shows both dd-carboxypeptidase and dd-endopeptidase activities.
An inspection of the amino acid sequence revealed an N-terminal sequence peptide but no hydrophobic membrane-anchoring sequence (95). The mature overproduced 42-kDa form remained membrane associated and was solubilized by detergent (79). The removal of the signal peptide and the analysis of spheroplasts suggested a localization at the outer surface of the inner membrane. Like PBP4, the attachment of AmpH implies interactions with other membrane constituents.
dd-Endopeptidase PBP7/PBP8.
The identification and cloning of the pbpG gene encoding PBP7 were key to an understanding of the initially confused status of PBP7 and PBP8 (94), characterized by the inconsistency of their appearance in PBP assays and by differences in their assignments (222). PBP8 is an artifact of sample preparation due to the proteolytic truncation of the C-terminal end of PBP7 by the protease OmpT (93). Together, PBP7 and PBP8 make up a significant fraction of the total PBP pool and are as abundant as PBP5 (60). PBP7 displays some sequence identity with PBP4, PBP5, and PBP6. PBP7 and PBP8 have a common N-terminal end and are processed in the same way as the other LMM PBPs (222). The amino acid sequence of PBP7 revealed no transmembrane helix, amphiphilic helix, or lipid anchor (94). PBP7 and PBP8 are loosely membrane-associated periplasmic proteins, since substantial amounts are released upon spheroplast formation or by osmotic shock and by salt treatment of a membrane fraction (222).
Purified PBP7 and PBP8 are dd-endopeptidases (Fig. 2) with a strict requirement for intact PG as a substrate. They both cleave d-alanyl→(d)-meso-A2pm cross-linkages in isolated PG but cleave (l)-meso-A2pm→(d)-meso-A2pm cross-linkages poorly (222). Moreover, they have no activity on isolated dimer muropeptides, and they are devoid of any dd-carboxypeptidase activity (222). PBP7 and PBP8 thus differ from the dd-endopeptidases PBP4, AmpH, and MepA, which cleave the d-Ala→(d)-meso-A2pm cross-linkage both in soluble dimer muropeptides and in isolated PG. These results emphasize the unusually narrow substrate specificity of PBP7/PBP8.
E. coli grows normally when pbpG is inactivated or overexpressed (94). A protein-protein interaction between PBP7/8 and Slt70 was demonstrated by affinity chromatography (223). Furthermore, PBP8 stabilizes and stimulates the activity of Slt70 in vivo. It was speculated that Slt70, PBP7/8, and PBP3 could form a multienzyme complex in vivo (223). PBP7 also plays a role in the penem-induced lysis of nongrowing cells (266).
Penicillin-Insensitive Peptidases
dd-Endopeptidase MepA.
Early work on E. coli PBP dd-peptidases revealed the presence of a penicillin-insensitive peptidase degrading isolated PG and cleaving the d-Ala→(d)-meso-A2pm cross-linkage of the dimer muropeptide (89, 252). Purification to homogeneity yielded a 30-kDa soluble enzyme, which bound no known β-lactam antibiotic and which remained fully active in their presence (122, 123, 260). It operated exclusively as an endopeptidase (Fig. 2) and displayed no detectable dd-carboxypeptidase activity with UDP-MurNAc-pentapeptide as a substrate (122). Single-stranded DNA and polynucleotides had an inhibitory effect (122, 260). Two deficient mutants, named mepA and mepB, for murein endopeptidase, were isolated (107). Comparison with the N-terminal sequence of purified endopeptidase showed that mepA is the structural encoding gene and that MepA is synthesized initially with an N-terminal signal peptide, indicating a periplasmic location (123).
The crystal structure of MepA (Fig. 3h) was determined at a 1.4-Å resolution (161). The C-terminal catalytic domain contains a zinc-binding site, in agreement with the sensitivity of MepA to metal chelators. The active site resembles that of the LAS metallopeptidases, which include lysostaphin, d-alanyl-d-alanine carboxypeptidase, and sonic hedgehog protein, their initials forming the term LAS. The MepA sequence has two conserved LAS-characteristic motifs but no PBP Ser-Xaa-Xaa-Lys motif. The reaction mechanism of MepA thus differs from that of the LMM PBPs (123). Furthermore, there is no significant sequence homology with the endopeptidases PBP4 and PBP7, and overproduction leads to no change in the normal degree of cross-linking (123). Mutational analysis revealed that the metal ligands conserved in MepA and its homologs are required for folding and catalysis (69).
EnvC endopeptidase.
A mutant forming chains of cells of irregular lengths was isolated from E. coli, and the mutated gene was designated envC (219) and mapped (120). As septation was partially blocked, some cells became longer than normal, and daughter cell poles remained fused to each other for several generations. The mutant showed hypersensitivity to dyes, a leakage of periplasmic proteins, and other defects indicating the perturbation of the outer membrane (121). Later, envC was recognized as the yibP open reading frame, and the single-residue substitution causing the mutation was identified (88).
The cleavage of the signal sequence yields a mature protein anchored to the cytoplasmic membrane, and a 43-kDa form is released upon spheroplast formation (88). The N-terminal half of the mature form is composed mostly of sequences with a repetitive pattern characteristic of a coiled-coil structure possibly involved in the oligomerization of EnvC or in its interaction with another protein (88). The activation of AmiA and AmiB by EnvC (273) suggests a possible protein-protein interaction between EnvC and these amidases. The C-terminal domain of EnvC resembles a zinc metallopeptidase and has homology with the staphylococcal LytM lysostaphin, which is known to degrade PG cross-linkages (88). The PG hydrolase activity of EnvC was assessed by zymography (20). Furthermore, EnvC hydrolyzes β-casein (106) and binds to PG in cosedimentation assays (273). These facts are consistent with PG endopeptidase activity. However, this finding is disputed by the absence of the zinc-chelating residues of the LytM active site and by the fact that different forms of EnvC have no effect on isolated dye-labeled PG (273). Therefore, it is difficult to decide whether EnvC truly has peptidase activity.
The disruption of chromosomal envC causes the same defects as the envC point mutation, indicating that the gene is dispensable for growth but important for normal septation and cell envelope integrity. The overexpression of envC leads to growth inhibition, morphological aberrations, and cell lysis (88, 273). EnvC, together with three other LytM factors (NlpD, YgeR, and YebA) discovered in E. coli, is required for septal PG splitting and daughter cell separation (272).
ld-Endopeptidase releasing d-alanyl-d-alanine from UDP-MurNAc-pentapeptide.
A peptidase cleaving the (l)-meso-A2pm→d-Ala bond of UDP-MurNAc-pentapeptide (Fig. 2) with the release of UDP-MurNAc-tripeptide and d-alanyl-d-alanine was identified in E. coli (78). Partially purified as a 34-kDa soluble enzyme, this ld-endopeptidase was devoid of any MurF synthetase activity catalyzing the addition of d-alanyl-d-alanine onto UDP-MurNAc-tripeptide. Conversely, purified MurF had no hydrolytic effect on UDP-MurNAc-pentapeptide. However, in B. subtilis, purified MurF catalyzes the removal of d-alanyl-d-alanine from UDP-MurNAc-pentapeptide but only in the presence of Co2+ ions (63). This is not strictly a reverse reaction, since no ATP is formed in the presence of ADP and phosphate.
Furthermore, it was suggested that the E. coli ld-endopeptidase could be the ld-transpeptidase catalyzing the formation of the (l)-meso-A2pm→(d)-meso-A2pm cross-linkages (145). This proposal is consistent with the finding that the ld-transpeptidase from Enterococcus faecalis, which catalyzes the formation of dimers by the removal of d-alanyl-d-alanine from the branched disaccharide-heptapeptide monomer muropeptide, also displays ld-endopeptidase activity catalyzing the removal of d-alanyl-d-alanine from the same substrate (158).
ld-Carboxypeptidases cleaving the tetrapeptide (l)-meso-A2pm→d-Ala bond.
ld-Carboxypeptidase activity cleaving the (l)-meso-A2pm→d-Ala bond of the disaccharide-tetrapeptide muropeptide or of the UDP-MurNAc-tetrapeptide (Fig. 2) with the release of d-alanine was initially detected in E. coli (112, 198) and was later detected in many other organisms (179, 256). The use of UDP-MurNAc-tetrapeptide as a substrate allowed the characterization of three E. coli ld-carboxypeptidases (14, 180, 274). Two of the enzymes were released by Tris-EDTA treatment and were purified as 86-kDa and 12-kDa proteins (14, 180). The osmotic shock treatment confirmed the periplasmic localization of the 86-kDa enzyme (13, 14), but that of the 12-kDa enzyme remained uncertain (180).
The third enzyme was the best-studied ld-carboxypeptidase. Its inhibition by nocardicin A allowed its purification by affinity chromatography as a 32-kDa enzyme found exclusively in the cytoplasm (274). The identified encoding gene was designated ldcA (256). The substrate specificity of LdcA was examined with various derivatives carrying the l-Ala-γ-d-Glu-(l)-meso-A2pm-(l)-d-Ala tetrapeptide (142, 256, 274). Only monomeric muropeptides (with either a MurNAc or a 1,6-anhydro-MurNAc residue), free tetrapeptides, and UDP-MurNAc-tetrapeptide are substrates. LdcA interacts with neither isolated PG nor cross-linked muropeptides (274). No ld-carboxypeptidase activity was detectable in soluble extracts from an ldcA deletion mutant (256). Moreover, the growth and morphology of the mutant are severely affected at the onset of the stationary phase, and it is prone to lysis. The high-level overproduction of LdcA has no effect on the growth rate or cell viability (256). LdcA plays a major role in the recycling process by converting the potentially harmful l-Ala-γ-d-Glu-(l)-meso-A2pm-(l)-d-Ala tetrapeptide into the l-Ala-γ-d-Glu-(l)-meso-A2pm tripeptide (196).
Among the five ld-transpeptidases (ErfK, YcfS, YbiS, YcbB, and YnhG) recently discovered in E. coli, three (ErfK, YcfS, and YbiS) anchor the Lpp lipoprotein to PG, whereas two (YcbB and YnhG) are responsible for the formation of the (l)-meso-A2pm→(d)-meso-A2pm cross-linkages (157, 159). They also catalyze the exchange of d-alanine with glycine in PG tetrapeptide subunits. All five ld-transpeptidases show ld-carboxypeptidase activity catalyzing the formation of tripeptide subunits from tetrapeptide subunits. They appear as periplasmic proteins possessing a sole cysteine residue essential for activity (54), like other ld-transpeptidases (160). The identification of either one of these E. coli ld-transpeptidases with the purified 12-kDa and 86-kDa ld-carboxypeptidases or with the d-alanyl-d-alanine-releasing ld-endopeptidase remains an open question.
MpaA amidase cleaving the γ-d-Glu→(l)-meso-A2pm bond.
In the PG peptide subunits, the γ-d-Glu→(l)-meso-A2pm bond is not sensu stricto a peptide bond, and the hydrolases cleaving it are designated γ-d-Glu→(l)-meso-A2pm amidases. Among the various reported examples, sporulation-related endopeptidase 1 (or ENP1 amidase) from Bacillus sphaericus has been well studied, and its C-terminal catalytic domain was shown to contain a zinc-binding site (105). The release of the dipeptide (l)-meso-A2pm-d-Ala into the culture medium of growing cells was the first indication of a similar activity in E. coli (82). Later, the presence of the dipeptide l-Ala-d-Glu was demonstrated in an mpl mutant lacking the UDP-N-acetylmuramate:tripeptide ligase and accumulating large amounts of the l-Ala-γ-d-Glu-(l)-meso-A2pm tripeptide (176).
Regions of the E. coli ycjI open reading frame displayed significant homology with the catalytic domain of ENP1 and its zinc-binding triad (269). The encoding gene was named mpaA, for murein peptide amidase A, and its in vivo activity was assessed by the accumulation of the l-Ala-γ-d-Glu-(l)-meso-A2pm tripeptide in an mpl mpaA double mutant (269). MpaA recognizes tri- and tetrapeptides (Fig. 2) but not disaccharide-peptides or UDP-MurNAc-peptide precursors (269). The finding of an absence of a signal sequence in mpaA was in favor of a cytoplasmic localization (269). However, the release of (l)-meso-A2pm-d-Ala in the growth medium as a PG turnover product does not exclude the possible presence of some MpaA activity in the periplasm (82).
l-Alanyl-d/l-glutamate dipeptide epimerase.
The search for members of the enolase superfamily in the E. coli and B. subtilis genomes led to the discovery of the l-alanyl-d/l-glutamate epimerases YcjG and YkfB, respectively, which convert l-Ala-d-Glu to l-Ala-l-Glu (233). Their crystal structures (Fig. 3i) were determined at 2.6- to 2.7-Å resolutions, and their reaction mechanism was studied (84, 128). The location of the gene encoding the E. coli YcjG epimerase downstream of mpaA suggested that it could play a role in PG metabolism. Its likely cytoplasmic localization excludes any action on PG. In E. coli, the l-Ala-l-Glu reaction product of the epimerase is a substrate for the broad-specificity PepD dipeptidase (234). Thus, the sequential actions of MpaA, YcjG, and PepD can release all of the amino acids from the PG tripeptide. However, this is a minor pathway, since in the recycling process, the Mpl ligase converts most of the cell tripeptide to UDP- MurNAc-tripeptide (196).
DdpX dipeptidase cleaving d-alanyl-d-alanine.
In PG-containing bacteria, d-alanyl-d-alanine is an essential cytoplasmic metabolite used in the formation of UDP-MurNAc-pentapeptide (286). The Ddl ligase catalyzes the ATP-driven condensation of two d-alanine molecules to form d-alanyl-d-alanine, and MurF synthetase catalyzes the addition of the dipeptide onto UDP-MurNAc-tripeptide. In the mechanism of resistance to vancomycin in enterococci, the dipeptidase VanX is responsible for the hydrolysis of the dipeptide (214). Analyses of the E. coli genome database revealed the presence of a VanX homolog with 27% similarity, originally referred to as EcoVanX and later renamed DdpX (144, 145). DdpX was overproduced and purified, and its activity was validated. DdpX conserves the active-site residues and auxiliary residues maintaining the active-site topology of VanX, and a kinetic analysis showed the same substrate specificity (144). Homology modeling of DdpX with the crystal structure of VanX from Enterococcus faecium (EntVanX) revealed a striking similarity in the overall structures, with the highest identity seen within the key catalytic residues (144).
DdpX has a potential lethal activity owing to the hydrolysis of the d-alanyl-d-alanine necessary for PG biosynthesis. However, DdpX is present preferentially at the onset of the stationary phase (144, 145). Therefore, in growing cells, DdpX plays no role in the regulation of the dipeptide pool controlled by the feedback inhibition of the Ddl ligase (286). The tight regulation of the d-alanyl-d-alanine pool is required to avoid the depletion of the alanine pool. In the stationary phase, DdpX could hydrolyze d-alanyl-d-alanine and supply d-alanine for use as metabolic fuel (145).
Enzymatic properties of E. coli PG peptidases.
The identified PG peptidases (Table 1) are characterized by their narrow specificity for a given peptide bond, except for PBP4 and AmpH, which recognize both d-Ala→d-Ala and d-Ala→(d)-meso-A2pm bonds. Some are unique for a given bond (MepA and YcjG). Redundancies concern the d-Ala→d-Ala bond (PBPs 4, 5, and 6; AmpH; and DdpX), the cross-bridges (PBP4, PBP7, AmpH, MepA, and EnvC), and the (l)-meso-A2pm→d-Ala bond (ld-carboxypeptidases and ld-transpeptidases), but there are great differences in substrate specificity between isoenzymes of a same family. Structural data (Fig. 3) are now available for six peptidases (PBPs 4, 5, and 6; MepA; DdpX; and YcjG), and reaction mechanisms have been proposed for some of them.
LMM PBPs have a common two-step reaction mechanism (77). The initial acylation step (Fig. 5a) involves the nucleophilic attack of the d-Ala→d-Ala or the d-Ala→(d)-meso-A2pm bond by a serine at the carbonyl carbon of the penultimate d-Ala residue, which results in a tetrahedral high-energy intermediate. The collapse of this species gives rise to an acyl enzyme intermediate and the release of the C-terminal d-Ala or of the acceptor peptide unit. In the subsequent deacylation step, a water molecule attacks the acyl enzyme to generate another tetrahedral high-energy species, which in turn collapses to yield the shortened peptide subunit and the regenerated PBP.
Fig. 5.

Reaction mechanisms of the LMM PBPs and d-Ala-d-Ala dipeptidases. (a) Two-step catalytic mechanism with the formation of an acyl enzyme intermediate and its attack by a water molecule. (b) Attack of the dipeptide carbonyl by a water molecule and formation of a bidentate complex with the catalytic Zn2+.
Several penicillin-insensitive E. coli PG peptidases (MepA, EnvC, MpaA, and DdpX) bind Zn2+ ions and have no PBP active-site sequence motifs, indicating that their reaction mechanisms differ from that of the PBPs (70). The strong structural similarity of DdpX with VanX and the conservation of the active-site topology (144) suggest that DdpX could follow the same reaction mechanism as that of VanX (32, 167). This implies (Fig. 5b) that the incoming d-alanyl-d-alanine dipeptide displaces a zinc-bound water molecule toward Glu181. The water molecule, activated by the influence of the catalytic zinc and Glu181, attacks the polarized carbonyl to form a tetrahedral intermediate, yielding a bidentate complex with the catalytic zinc. Subsequently, Glu181 donates the proton to the scissile nitrogen, completing the reaction with the release of d-alanine.
E. COLI N-ACETYLMURAMOYL-l-ALANINE AMIDASES
In bacterial PG, the peptide subunits are bound to the glycan chains by an amide bond between their N-terminal l-alanine residue and the d-lactoyl moiety of the MurNAc units (Fig. 1). Amidases cleaving the N-acetylmuramoyl→l-alanine bond are present in many eubacteria (238, 240, 295). In E. coli, an N-acetylmuramoyl-l-alanine amidase acting on monomer and dimer muropeptides was first detected in a crude cell extract (198). So far, six E. coli N-acetylmuramoyl-l-alanine amidase activities (Table 1) have been reported: five (AmiA, AmiB, AmiC, AmpD, and AmiD) were genetically well defined, while another one (39-kDa amidase) was only biochemically characterized. Based on the similarity of their amino acid sequences, AmiA, AmiB, and AmiC are members of the amidase_3 superfamily of the Pfam database, whereas AmiD and AmpD belong to the amidase_2 superfamily of the same database (200).
AmiA, AmiB, and AmiC N-Acetylmuramoyl-l-Alanine Amidases
Homology studies led to the identification of AmiA, AmiB, and AmiC as N-acetylmuramoyl-l-alanine amidases. Upstream of the hemF gene at centisome 55, an open reading frame codes for a 32-kDa protein homologous to the CwlB amidase from B. subtilis (262). The designation amiA is now used for the encoding gene (91) and must not be confused with the same previously proposed designation (261) for the 39-kDa amidase (197, 283). A second open reading frame at centisome 94.7 encodes a 48-kDa protein with homology to AmiA and other bacterial amidases, and the gene was designated amiB (263). Finally, an open reading frame at centisome 63.4 showed significant sequence similarity to other amidases, and the gene encoding a 45-kDa protein was named amiC (91). The overexpression of amiB or amiC led to cell lysis (91).
The presence of a signal sequence in each amidase indicated their periplasmic localization (19, 91). AmiA and AmiC are exported to the periplasm by the Tat system (19, 113), whereas AmiB is supposedly exported by the more general Sec pathway (19). The subcellular periplasmic localization of AmiA and AmiC was studied by use of a functional fusion to green fluorescent protein (GFP) (19). AmiA-GFP remains dispersed in the periplasm throughout the cell cycle, and AmiC-GFP remains dispersed only in nondividing cells. In contrast, AmiC-GFP was concentrated mostly at the septal ring in dividing cells, as confirmed by an electron microscopy analysis of the metallothionein-tagged protein (55).
The release of the l-Ala-γ-d-Glu-(l)-meso-A2pm tripeptide and the l-Ala-γ-d-Glu-(l)-meso-A2pm-(l)-d-Ala tetrapeptide was observed after the incubation of isolated PG with overproduced AmiA and AmiC (91). An analysis of changes in PG structure revealed the amidase activity of AmiB. The release of soluble material upon incubation with isolated dye-labeled PG confirmed the activity of all three amidases (273). AmiA, which is a zinc enzyme, cleaves un-cross-linked PG fragments having at least a tetrasaccharide glycan chain (154). Neither amidase cleaves 1,6-anhydro-MurNAc→l-Ala bonds (270). Mutants devoid of any one of the three amidases were constructed and showed no major changes in growth (91). Morphological changes appeared mostly in the late stationary phase, with the AmiA and AmiC mutants growing in chains of 3 to 6 cells. None of the single mutants showed any major change in PG structure.
AmpD Anhydro-N-Acetylmuramoyl-l-Alanine Amidase
The ampD gene of enterobacteria was first shown to encode a cytoplasmic protein acting as a negative regulator of β-lactamase induction (148). It was later established that AmpD is an N-acetylmuramoyl-l-alanine amidase exhibiting an exclusive specificity for substrates containing 1,6-anhydro-MurNAc residues (101, 116). In the cytoplasm, AmpD rapidly cleaves 1,6-anhydro-MurNAc-peptides but has no effect on UDP- MurNAc-pentapeptide (196). This high specificity is confirmed by the cleavage of the 1,6-anhydro-MurNAc→l-Ala bond of the 1,6-anhydro-MurNAc-peptide located at the end of isolated PG chains (100). GlcNAc-(1→4)-1,6-anhydro-MurNAc-tetrapeptide is also recognized as a substrate (97). The accumulation of 1,6-anhydro-MurNAc-tripeptide in an AmpD-deficient mutant suggests that it is the in vivo substrate of AmpD in the PG-recycling process (196). AmpD from Citrobacter freundii is 90% identical in sequence to the E. coli enzyme, and its 3D structure determined by NMR revealed a deep substrate-binding pocket (147). To explain the specificity of AmpD for low-molecular-mass substrates, a model of the enzyme-substrate complex was established.
AmiD Anhydro-N-Acetylmuramoyl-l-Alanine Amidase
A second enzyme with 1,6-anhydro-MurNAc-l-alanine amidase activity was identified when a nagZ ampD mutant was found to accumulate not only GlcNAc-(1→4)-1,6-anhydro-MurNAc-tripeptide, as expected, but also a significant amount of the GlcNAc-(1→4)-1,6-anhydro-MurNAc disaccharide in the cytoplasm (270). A search of the genome revealed a paralog, YbjR, having 40% identity with a 123-amino-acid sequence of AmpD, and it was named AmiD. The presence of a short signal sequence and a lipobox motif in ybjR suggested that AmiD is a lipoprotein, and it was demonstrated to be in the outer membrane (270). Membrane and soluble forms of AmiD were overproduced, purified, inactivated by chelators, and shown to bind a zinc ion required for activity (200, 270).
A null mutation had no effect on cell growth or morphology (270). The deletion of amiD from the nagZ ampD double mutant produced a strain totally lacking 1,6-anhydro-MurNAc-l-Ala amidase activity, demonstrating that AmpD and AmiD were the only enzymes in E. coli able to cleave the 1,6-anhydro-MurNAc→l-Ala bond (270). AmiD has a broader substrate specificity than AmpD. It can cleave both MurNAc→l-Ala and 1,6-anhydro-MurNAc→l-Ala bonds in muropeptides and in MurNAc-tri- or tetrapeptides, with the disaccharide-tetrapeptide muropeptide being the best substrate (200, 270). AmiD presumably corresponds to the MurNAc-l-Ala amidase activity detected previously by using radiolabeled disaccharide-tetrapeptide as a substrate (80). Moreover, the digestion of isolated PG leads to the release of free tri-, tetra-, and octapeptides, whereas AmiD has no effect on MurNAc-l-Ala-d-Glu, UDP-MurNAc-tripeptide, UDP-MurNAc-pentapeptide, lipid I, or lipid II (200, 270).
The crystal structure of AmiD devoid of its lipid anchorage was determined at a 1.8-Å resolution (124). The overall shape is a 45-Å-long and 30-Å-thick cylinder, with the zinc ion critical for activity located at the intersection of the two branches of the L-shaped groove of the active site (Fig. 3j). The structures of AmiD in complexes with 1,6-anhydro-MurNAc-tripeptide or the tripeptide reaction product were also determined. AmiD shares a common fold with AmpD from Citrobacter freundii, bacteriophage T7 lysozyme zinc amidase, and PG recognition proteins (124).
39-Kilodalton Amidase Cleaving MurNAc-Tripeptide
An N-acetylmuramoyl-l-alanine amidase activity from E. coli was biochemically characterized by use of radiolabeled MurNAc-tripeptide as a substrate (197, 261, 282, 283, 308). After cell disruption by sonication, it appeared mainly (261, 283) but not exclusively (308) in the soluble fraction. Most of the activity was released upon spheroplast formation, but none was released by osmotic shock, suggesting binding to a periplasmic structure or an outer membrane location (197).
The enzyme was purified to near homogeneity from the spheroplast supernatant as a 39-kDa protein (283) and was shown to have a narrow substrate specificity limited to MurNAc-peptides (197). The enzyme had practically no effect on the disaccharide-tetrapeptide muropeptide and 1,6-anhydro-MurNAc-containing muropeptides or on isolated PG. A mutant showing 95% reduced amidase activity with MurNAc-tripeptide was isolated and showed no distinct physiological changes (261). However, it was not established whether the mapped gene, named amiA, was truly the structural gene of the 39-kDa amidase. Owing to clear differences in specificities, the identification of the 39-kDa amidase with any of the five amidases genetically characterized so far is unlikely. It could play a role in the processing of MurNAc-peptides originating from the turnover reaction products or from exogenous PG fragments used as metabolic fuel.
Enzymatic Properties of the E. coli Amidases
The substrate specificities of the E. coli PG amidases differ considerably from one another. Some act on isolated PG (AmiA, AmiB, and AmiC), while others recognize only low-molecular-mass PG fragments (AmpD and 39-kDa amidase). The amidase AmiD has the broadest specificity by acting on both types of substrates. Several amidases (AmiA, AmpD, and AmiD) are zinc metalloenzymes. So far, the biochemical properties of only AmiD (200) and the 39-kD amidase (197) have been studied to some extent. In particular, AmiD exhibits a biphasic time course, which is explained by the presence of a branched pathway involving a substrate inactivation process (200). It is unknown whether this unusual kinetic behavior has an in vivo regulatory significance.
The determination of the crystal structure of AmiD led to the proposal of a reaction mechanism (Fig. 6) similar to that of the thermolysin-related zinc peptidases (124). Upon the approach of the substrate, the water molecule bound to active-site Zn2+ is shifted toward residue Glu104, which favors the nucleophilic attack of the amide bond, producing a tetrahedral conformation of the amide carbon and a transient pentameric coordination of the zinc ion. Hydrogen interactions involving the carbonyl oxygen of the scissile bond stabilize the tetrahedral intermediate. The next step is proton transfer from the protonated glutamic acid to the leaving nitrogen. Mutation analysis demonstrated the critical role played by Glu104 in the catalytic mechanism.
Fig. 6.

Reaction mechanism of the AmiD amidase proposed by Kerff et al. (124). The scheme describes the catalytic mechanism for the AmiD hydrolysis of the MurNAc-l-Ala amide bond. (A) Activation of the catalytic water molecule by Zn2+ and Glu104 upon binding of the substrate; (B) formation of the tetrahedral intermediate stabilized by Zn2+ and Lys159; (C) rotation of the amide bond leading to an ideal tetrahedral conformation of the amide bond nitrogen; (D) transfer of the hydrogen from the protonated Glu104 to the amide nitrogen and cleavage of the amide bond. R, remaining peptide moiety.
HYDROLASE-DEFECTIVE MUTANTS
To determine possible functional roles, single defective mutants were constructed for most E. coli PG hydrolases. Except for the deletions of the dd-carboxypeptidase PBP5 (170, 185), the ld-carboxypeptidase LdcA (256), the endopeptidase EnvC (219), and the amidases AmiA and AmiC (92), most mutants grew well and displayed no morphological changes. Therefore, many hydrolases are not individually essential, at least under conditions of laboratory culture. This is in sharp contrast with the functionally well-defined enzymes of the PG biosynthesis pathway leading to lipid II, which are unique and essential (5, 224, 286). For this reason, the construction of mutants with multiple deletions of PG hydrolases was undertaken. Since access to mutants with all possible combinations of deletions is not readily feasible, only a limited number of multiple mutants of the same or different enzymatic specificities have been considered so far.
Multiple Deletions of the LTs
Several multiple-deletion mutants of the LTs were constructed (92, 149). The double mutants lacking Slt70, MltA, or MltB as well as the triple mutants lacking all three LTs displayed no obvious phenotype and no effect on morphology (149). The triple mutant showed a reduced rate of PG turnover and a low level of β-lactamase induction, but neither the rate of PG synthesis nor the cell PG content varied (133). A triple mutant lacking MltC, MltD, and MltE and a hextuple mutant with deletions of six LTs, not including MltF, grew with 30% to 50% of the cells in short chains containing three to eight cells (92). Only minor changes in the PG structure were detected, and no significant variation in the 1,6-anhydro-MurNAc content was observed (92). An attempt to isolate a multiple mutant deficient for all seven identified LTs failed (231).
Multiple Deletions of the LMM PBPs
Mutants with multiple deletions of the LMM PBPs were constructed (6, 29, 49, 62, 92, 226, 250, 291). In particular, mutants lacking all possible combinations of PBP4, PBP5, PBP6, PBP6b, and PBP7 (49) as well as a mutant lacking six LMM PBPs (291) were generated. All multiple deletions had little or no effect on growth under laboratory conditions. However, some multiple-PBP deletions produced cells with random shapes, lengths, and diameters (49, 62, 170, 185, 291). The minor alterations observable for some simple mutants were enhanced in multiple mutants. In particular, the modifications in the PBP5 single mutant were also present in the different multiple mutants lacking PBP5. They were exacerbated by the additional deletion of PBP4 or/and PBP7, with the greatest alterations being observed with the triple mutant (170). PBP4 and PBP7 are thus endopeptidases influencing cell shape, but the absence of shape abnormalities in mutants lacking only PBP4 and PBP7 stresses the importance of PBP5. A PBP4 PBP5 double mutant showed a higher increase of the PG pentapeptide subunit content than did the PBP5 single mutant (134). However, the triple deletion of the endopeptidases PBP4, PBP7, and MepA had no effect on morphology and only minor effects on the PG structure (92). Cells with a double deletion of PBP5 and PBP6 grew as filaments failing to construct and divide efficiently (226).
Multiple Deletions of the N-Acetylmuramoyl-l-Alanine Amidases
Mutants lacking two or all three AmiA, AmiB, and AmiC amidases showed normal growth (91). In the three double mutants, cell separation took place, but 5 to 20% of the cells had a tendency to grow in chains. The triple mutant grew in long chains, and the change in morphology was accompanied by an increase of trimer and tetramer contents as well as a higher 1,6-anhydro-MurNAc content. The double deletion of amiA and amiC rendered the integrity of the cell envelope highly defective, resulting in an increased permeability of the outer membrane to hydrophobic antibiotics and detergents (113).
Multiple Deletions of Hydrolases of Different Enzymatic Specificities
An early study concerned mutants with multiple deletions in the amidases AmiA, AmiB, and AmiC combined with deletions in the endopeptidases PBP4, PBP7, and MepA or in LTs (92). The average number of cells in the chains increased significantly compared to the average number of cells in chains of amidase mutants. A multiple-amidase mutant also lacking the endopeptidases MepA, PBP4, and PBP7 formed chains containing 6 to 80 cells. A mutant with deletions in the three amidases, the three endopeptidases, and the LT Slt70 formed the longest chains, consisting of 20 to 100 cells. The conclusion was that all three types of activities are involved in the splitting of the PG septum (92). Some mutants showed lower outer membrane permeabilities and greater susceptibilities to antibiotics, whereas LT activity was essential for β-lactamase induction (131).
Recently, mutants combining multiple deletions of the LMM PBPs and the AmiA, AmiB, AmiC amidases were constructed to determine which hydrolases facilitate cell separation (210, 211). Particular combinations of PBPs and amidases affected the overall septum geometry in that certain mutants formed highly abnormal cell chains. Amidase mutants lacking PBP4 grew as longer chains, identifying a specific biological role for this endopeptidase in cell separation. PBP7 appeared to play a minor role. The deletion of PBP5 from a mutant lacking both AmiA and AmiC led to twisted chains (210).
Several mutants with multiple deletions of the LdcA, AmiD, AmpD, and NagZ hydrolases involved in the recycling process were constructed (196, 256, 270). In particular, a mutant lacking AmpD, AmiD, NagB, and NagZ was used to establish the important role of AmiD in the recycling process (270).
HYDROLASES AND PEPTIDOGLYCAN MATURATION
If PG metabolism was restricted to the formation of the glycan chains by transglycosylation and their cross-linking by transpeptidation using pentapeptide subunits, mature PG would contain only tetrapeptide subunits in the donor position of the dd-cross-linkages and unused pentapeptide subunits. However, the analysis of the muropeptide composition of isolated E. coli PG (74) revealed a far greater structural complexity (Fig. 1) characterized by the presence, in particular, of various truncated peptide subunits, ld-cross-linkages, glycan chains with nonreducing 1,6-anhydro-MurNAc ends, and substitutions by the Lpp lipoprotein (61) or glycine (74). These modifications are due to maturation reactions taking place in the periplasm during the formation of new PG. Pulse and pulse-chase labeling experiments carried out with radiolabeled A2pm revealed that the various reactions follow different kinetics from incipient to mature PG (33, 47, 51, 75). Several envelope-associated PG hydrolases are involved in the process.
Tetrapeptide Subunits
The most obvious feature of the muropeptide composition of PG from growing E. coli cells is the predominance of tetrapeptide subunits, which amount to ca. 60% of the total peptide subunits, and the presence of less than 1% pentapeptide subunits (74). Short pulse experiments showed the rapid formation of tetrapeptide subunits from pentapeptide subunits very early in the synthesis of nascent PG (47, 75). The pentapeptide subunits are used as donors in the formation of the d-Ala→(d)-meso-A2pm cross-linkages by dd-transpeptidation, and the unused ones are converted to tetrapeptide subunits by the removal of their C-terminal d-alanine residue. Pulse-chase experiments revealed a complex time course for the formation and use of the tetrapeptide subunits, which are at the crossroads of several reactions (33, 75). Tetrapeptide subunits are used as acceptors in the dd-transpeptidation reactions but also as energy donors in the formation of (l)-meso-A2pm→(d)-meso-A2pm cross-linkages by ld-transpeptidation, the linking of the Lpp lipoprotein, and the replacement of d-alanine residues by glycine. At the same time, tetrapeptide units are formed as a PG turnover product by the dd-endopeptidase cleavage of d-Ala→(d)-meso-A2pm cross-linkages.
PBP5 appears to be the major dd-carboxypeptidase responsible for the conversion of the pentapeptide subunits not used in transpeptidation to tetrapeptide subunits. The mutational loss of PBP5 leads to an increased pentapeptide subunit content (50, 134, 146). PBP5 cleaves the d-Ala→d-Ala bonds of un-cross-linked and cross-linked PG (1, 205). The additional deletion of PBP4 enhances the pentapeptide content, as observed for a double-defective mutant (134). The deletion of PBP4 alone leads to no great increase in numbers of pentapeptide subunits. This finding suggests that both PBPs cooperate in some way in controlling the amount of pentapeptide subunits available for transpeptidation and regulating the degree of PG cross-linkage (48, 170, 185, 205). The known 3D structures of PBP5 and bifunctional PBP1 have given rise to a model (Fig. 7) for the coordinated removal of d-alanine from PG by hydrolytic cleavage and transpeptidation (205). The close attachment of PBP5 to the outer face of the cytoplasmic membrane restricts its in vivo action to pentapeptide subunits of incipient PG.
Fig. 7.

Model showing the concomitant actions of PBP5 and PBP1b on the pentapeptide subunits of nascent PG. Arrows represent the pentapeptide subunits linked to the glycan chains (in black). (Reprinted from reference 205 with permission of John Wiley & Sons.)
Tripeptide Subunits
In the tripeptide subunits of E. coli PG, the l-carboxyl of the C-terminal meso-A2pm residue is either free or engaged in an (l)-meso-A2pm→(d)-meso-A2pm cross-linkage or linked to the Lpp lipoprotein or glycine (74). The unsubstituted and substituted forms account for ca. 13% and 10%, respectively, of the total PG peptide subunits. In growing cells, their formation follows a slow time course. Cells harvested without rapid cooling lead to higher free tripeptide subunit contents (303). In the stationary phase (74) or during amino acid starvation (268), a considerable increase of the tripeptide subunit content is observed in parallel with those of (l)-meso-A2pm→(d)-meso-A2pm cross-linkages and bound Lpp lipoprotein.
The formation of tripeptide subunits by the cleavage of the (l)-meso-A2pm→d-Ala bond of tetrapeptide subunits by the LdcA ld-carboxypeptidase is unlikely, considering its cytoplasmic localization and the absence of an effect on isolated PG (256, 274). Some tripeptide subunits could originate from the use of the UDP-MurNAc-tripeptide precursor in parallel with UDP-MurNAc-pentapeptide in the biosynthesis pathway (15, 285). However, recent results point to the major role played by ld-transpeptidases. A sharp decrease of the tripeptide subunit content was observed with a quadruple mutant lacking the ErfK, YcfS, YbiS, and YcbB ld-transpeptidases (157), and a total loss of tripeptide subunits accompanied their deletion together with that of the YnhG ld-transpeptidase (159). Thus, these enzymes are not only acting as ld-transpeptidases in the formation of (l)-meso-A2pm→(d)-meso-A2pm cross-linkages and the binding of the Lpp lipoprotein but are also responsible for the formation of the PG tripeptide subunits by the ld-carboxypeptidase cleavage of tetrapeptide subunits. Furthermore, the overexpression of each ld-transpeptidase gene led to the accumulation of tripeptide subunits. Biochemical data confirming their ld-carboxypeptidase activity are still lacking. The large increases in amounts of bound Lpp lipoprotein and (l)-meso-A2pm→(d)-meso-A2pm cross-linkages in the stationary phase parallel the increase in amounts of tripeptide subunits (74), indicating a close connection between ld-transpeptidase and ld-carboxypeptidase activities. The balance between the two activities is presumably dependent on the availability of the acceptors (d-amino-meso-A2pm groups and free Lpp lipoprotein). The location of free Lpp lipoprotein at the outer side of the outer membrane (42) could be a factor limiting its use. An enzyme showing both ld-transpeptidase and ld-carboxypeptidase activities was reported previously for E. faecalis (158).
Dipeptide Subunits
The presence of 3% of dipeptide subunits in isolated PG (74) could result from the cleavage of the γ-d-Glu→(l)-meso-A2pm bond in tri- and tetrapeptide subunits by the MpaA endoamidase. However, having no effect on disaccharide muropeptides and being located in the cytoplasm, its in vivo action on PG is unlikely (196, 269). Another possibility is that dipeptide subunits originate to a small extent from the use of the UDP-MurNAc-dipeptide precursor by the biosynthesis pathway in parallel with UDP-MurNAc-pentapeptide. The ratio of dipeptide/pentapeptide precursor pool levels is in agreement with the observed 3% of dipeptide subunits (172). A high PG dipeptide subunit content was observed previously for a Staphylococcus aureus mutant accumulating large amounts of UDP-MurNAc-dipeptide (192).
1,6-Anhydro-N-Acetylmuramic Acid
An important structural feature of E. coli PG is the presence of a 1,6-anhydro-MurNAc residue at the reducing end of the glycan chains, as substantiated by the extremely low reducing power of isolated PG (304) and the presence of 1,6-anhydro-MurNAc muropeptides accounting for 3 to 4% of the total muropeptide composition (74). This percentage is a measure of the average length of the glycan chains. By pulse and pulse-chase experiments, it was established that the glycan chains of nascent PG undergo a shortening from 50 to 60 disaccharide units to 31 disaccharide units in less than 5 min after formation, implying the action of LTs (75).
The formation of the glycan chains in the periplasm proceeds by the repetitive addition of the disaccharide units, with the reducing end of the growing chain remaining bound to undecaprenyl pyrophosphate (287). It was proposed that the termination of the polymerization reaction entails the release of the newly synthesized glycan chains from undecaprenyl pyrophosphate and the formation of 1,6-anhydro-MurNAc ends (132). Owing to their enzymatic activity, LTs should in some way be responsible for this process. However, a multiply defective mutant lacking six (Stl70, MltA, MltB, MltC, MltD, and MltE) out of the seven identified LTs exhibited no significant decrease of the 1,6-anhydro-MurNAc muropeptide content (92). Moreover, the PG material synthesized in vitro by purified glycosyltransferases remained bound to undecaprenyl pyrophosphate (8, 300). Thus, neither glycosyltransferases nor most LTs appear to be directly involved in the termination step. The enzymes catalyzing the release of undecaprenyl pyrophosphate, the formation of the 1,6-anhydro ends, and the shortening of the glycan chains during maturation remain to be clearly identified.
N-Acetylmuramoyl-l-Alanine Linkage
The muropeptide composition of isolated E. coli PG showed that MurNAc residues and peptide subunits were present in roughly equivalent amounts, and no muropeptide lacking the GlcNAc-(1→4)-MurNAc disaccharide was reported (74, 304). Therefore, if not nil, the number of cleaved MurNAc→l-Ala bonds in PG is low. The free N-terminal l-alanine amino groups in PG from cells grown under different conditions accounted for 1 to 2.5% of the total muramic acid content and were presumably those of truncated oligomer units (197). Cells harvested without rapid cooling have more cleaved MurNAc→l-Ala bonds, and their muropeptide composition shows the presence of free disaccharide and a truncated dimer (197, 283). Supposedly, only the amidase AmiA, AmiB, AmiC, or AmiD, having a known action on isolated PG (91, 200, 270, 273), is responsible for the in vivo cleavage of MurNAc→l-Ala bonds, which has also been observed for many other organisms.
HYDROLASES AND PEPTIDOGLYCAN TURNOVER
As PG is an essential continuous structural component of the cell envelope, its synthesis is correlated with cell growth and division. The incorporation of new PG material has been studied by various labeling and microscopy approaches (48, 313). The results are consistent with the presence of different PG-synthesizing machineries in a given organism. In E. coli, one is associated with cell elongation, while another one is involved in cell septation (48). The muropeptide composition is the same in lateral and septal PG, suggesting that the maturation reactions are also the same throughout the cell cycle (47, 183).
In growing E. coli cells, PG synthesis is associated not only with maturation reactions but also with a 40 to 50% turnover per generation of the cell wall PG (83, 196). It is possible to study lateral wall and septal PG turnovers independently from one another by specifically blocking septation or elongation (48, 271) or by using synchronously growing cells (47). Lateral wall and septal PGs undergo 60 and 30% turnovers, respectively, but cell pole PG does not and remains stable (33, 52). In the process, PG is continuously degraded in the periplasm to GlcNAc-(1→4)-1,6-anhydro-MurNAc-peptide monomers, GlcNAc-(1→4)-1,6-anhydro-MurNAc disaccharides, and free peptides (83, 196). Owing to size exclusion by the pores of the outer membrane, only some free peptides are released into the medium (82), whereas the greater part of the turnover products is reused in a recycling process (83, 196). The formation of mostly truncated monomer PG fragments is evidence for the combined action of LT, endopeptidase, and amidase activities. It is presently difficult to say which specific hydrolases play a role in lateral wall or septal PG turnover and whether they function randomly or in a defined order.
HYDROLASES AND LATERAL WALL PEPTIDOGLYCAN
The formation of nascent lateral wall PG proceeds by the incorporation of precursors at a limited number of discrete sites, which are highly mobile and distributed over the cell surface (48). Since nascent PG does not exist as free material in E. coli (33, 81), the extension of the growing glycan chains must proceed simultaneously with cross-linking to preexisting PG. Furthermore, the enlargement of lateral wall PG implies the cleavage of old bonds to enable the intercalation of the new glycan chains while preserving PG integrity. Different models describing the insertion of new glycan chains into lateral wall PG have been proposed (34, 41, 47, 103, 193).
In the cut-and-insert model (193), the insertion of a new single glycan chain is associated with the simultaneous action of an endopeptidase opening cross-linkages in preexisting PG (34). To avoid any rupture in the stress-bearing PG layer, the process implies a tight coordination between the endopeptidase activity and the cross-linkage-forming transpeptidases. In this model, the insertion of incipient PG and PG turnover do not appear to be necessarily linked processes (196, 271).
The three-for-one model is an elaborate two-step process (103). First, a triplet of new glycan chains is cross-linked to preexisting glycan chains, and this addition occurs adjacently to the cytoplasmic membrane. In the second step, the removal of a nearby preexisting chain by the endopeptidase cleavage of its cross-linkages leaves space for the incoming new material. As a result, three new glycan chains replace the preexisting one with a preservation of PG integrity, hence the name of the model. The released chain is degraded at the outer side of the multilayered PG (75). According to this inside-to-outside growth mechanism, insertion and turnover are two closely coupled processes (75).
It has not yet been determined which specific endopeptidases or LTs are involved in the enlargement process. Although all possible mutational combinations of hydrolase have not yet been considered, many of the single or multiple mutants of the endopeptidases and LTs constructed so far show normal growth, and it has been questioned whether these hydrolases are truly essential for the synthesis of lateral wall PG (92, 211). In vitro protein-protein interactions between Slt70 or MltA and PBP1b or PBP2 have been described (293, 297). An association of PG hydrolases with the multienzyme complex (elongase) responsible for lateral wall PG synthesis has been considered (48), but evidence is still lacking.
HYDROLASES AND SEPTAL PEPTIDOGLYCAN
During cell division, it is obvious that septal PG must be split to promote the separation of the daughter cells and that hydrolases are involved in the process. The formation and turnover of septal PG were investigated by the specific PG labeling of growing cells (47, 48, 52, 271). The septum is made exclusively of 40% of the newly synthesized cell PG, over 30% of which is rapidly degraded during septation, while the remaining hydrolysis-resistant part ends up in the stable poles formed after cell separation. Although quantitatively less important than that of lateral wall PG, septal PG turnover proceeds at a much higher rate, since it is functioning over a shorter time of the cell cycle. It was proposed that the rapid removal of excess septal PG is responsible for the separation of the new poles of the daughter cells (271). The involvement of LTs, endopeptidases, and amidases in septal turnover was substantiated by an analysis of the degradation products originating from radiolabeled septal PG (271). How the degradation of septal PG correlates with the splitting of the septum remains a matter of debate (92, 271). Supposedly, cell division follows an ordered sequence of events (initiation, invagination, septation, and cell separation), with different hydrolases functioning at different stages of the process.
Studies of certain defective mutants and subcellular localizations have given a few hints as to how hydrolases are involved in cell division. The role of the AmiA, AmiB, and AmiC amidases in septal PG turnover is illustrated by a triple-defective amidase mutant characterized by long chains of cells connected by both complete and incomplete septa, as revealed by electron microscopy analysis of whole cells and isolated sacculi (91, 211). Fluorescence microscopy confirmed the existence of rings on sacculi (211). The septal rings are material accumulating by continued PG synthesis after the arrest of turnover (211). They are observable in any of the three possible double amidase mutants but not in any of the single amidase mutants, indicating that all three amidases have a function in septal PG turnover (211). During the cell cycle, AmiC physically localizes mostly to the septal ring in the periplasm. The N-terminal nonamidase domain of AmiC is both necessary and sufficient for the targeting of the enzyme to the septal ring (19). Moreover, AmiC requires FtsN for recruitment to the septal ring and appears localized at the time of constriction initiation (20).
Presently, there are limited data on the role played by the PBP4, PBP7, and MepA endopeptidase activities in septal PG turnover. Their deletion has no morphological effect except when made in the triple-amidase-defective mutant (91, 210). On the other hand, the deletion of the putative EnvC endopeptidase alone or in combination with the NlpD LytM factor leads to morphological changes similar to those observed for the amidase mutants (272). Experiments with fluorescent fusion proteins revealed that both EnvC and NlpD are recruited to the septal ring during cell division, indicating that they play a role in septation (272). EnvC functions only at a very early stage of cell septation, since it localizes to the nascent division site prior to the start of active invagination and disappears from the septal ring in deeply constricted cells (20, 91, 92). A striking finding is the activation of the AmiA and AmiB activities by EnvC and that of AmiC by NlpD observed in in vitro assays with purified proteins and dye-labeled PG (273). EnvC and NlpD thus appear as regulators of the amidases involved in septal PG turnover. The mechanism of this activation remains to be determined.
The dd-carboxypeptidase PBP5 is involved in both elongation and septation, as established by immunolocalization and the use of fusions with green fluorescent protein (48, 205). When septation is inhibited, PBP5 shows a random and dispersed membrane localization, whereas it localizes almost exclusively at the septum in spherical cells obtained after the inhibition of elongation and even prior to the appearance of visible constriction. This indicates that PBP5 is where the peptidoglycan-synthesizing activity is highest, meaning that its interaction with pentapeptide subunits of incipient PG determines its localization. Interactions of PBP5 with proteins of the elongation or septation complexes are not required (48, 205). The importance of PBP5 in determining and maintaining cell morphology by controlling the availability of pentapeptide subunits for transpeptidation has been stressed (170, 185), but many aspects of the process remain unknown (205).
HYDROLASES AND PEPTIDOGLYCAN RECYCLING
In E. coli, over 90% of the products generated by PG turnover return to the cell for reuse (196). The pathways of this recycling have been worked out and reviewed in detail (196, 204). Most enzymes of the process were characterized, and the effect of their deletion was studied. Briefly, recycling proceeds according to three successive steps: (i) transport of the turnover degradation products generated in the periplasm into the cytoplasm, (ii) further intracellular degradation, and (iii) reuse of the final products.
After PG degradation in the periplasm, the resulting GlcNAc- (1,4)-1,6-anhydro-MurNAc-peptides are transferred into the cytoplasm by the highly specific AmpG permease (38), while the free PG peptides not released into the medium are taken up by peptide permeases (83, 195). The incorporated fragments are further degraded by the cytoplasmic AmpD, NagZ, and LdcA hydrolases into the l-Ala-γ-d-Glu-(l)-meso-A2pm tripeptide, d-alanine, GlcNAc, and 1,6-anhydro-MurNAc (196). The tripeptide is ligated into UDP-MurNAc by the Mpl ligase, and the resulting tripeptide nucleotide precursor returns to the biosynthesis pathway (96, 176). GlcNAc and 1,6-anhydro-MurNAc are both converted into GlcNAc-6-phosphate, which is available for reuse (119, 196, 204). The kinase NagK produces GlcNAc-6-P directly from GlcNAc, whereas the conversion of 1,6-anhydro-MurNAc is a two-step process. The kinase AnmK produces the MurNAc-6-phosphate converted to GlcNAc-6-phosphate by the MurQ etherase. A structural model of the E. coli MurQ etherase was generated, and a reaction mechanism for the cleavage of the d-lactylether bond by MurQ yielding MurNAc-6-phosphate was proposed (119).
The deletion of AmpD or NagZ has no effect on cell growth (196). However, the ld-carboxypeptidase LdcA is essential at the onset of the stationary phase, since an LdcA-deficient mutant forms thick oval cells, many of which lyse (256). A similar phenotype was observed with dithiazoline-inhibited LdcA (10). In the absence of LdcA activity due to mutation or inhibition, the l-Ala-γ-d-Glu-(l)-meso-A2pm-(l)-d-Ala tetrapeptide accumulates and is used by Mpl to form UDP- MurNAc-tetrapeptide (96, 256). Tetrapeptide subunits are incorporated into PG instead of pentapeptide subunits, leading to a cross-linkage deficiency and lysis. The effect due to the absence of LdcA activity suggests that at the onset of the stationary phase, recycling may be of critical importance for sustaining the prolonged synthesis of structurally modified PG resistant to autolysis (24).
Besides recycling its own PG, E. coli is able to grow on PG fragments, GlcNAc, and MurNAc as the sole carbon and nitrogen sources (118, 197, 306). Presumably, hydrolases of the recycling process, together with other cell enzymes, have an important role in the uptake and use of these exogenous products as metabolic fuel. In particular, this is exemplified by the action of the MurQ etherase in connection with growth on MurNAc (119).
HYDROLASES AND AUTOLYSIS
Being responsible for the mechanical resistance of the bacterial cell wall, PG is essential for preserving cell integrity. Its degradation by the uncontrolled action of endogenous PG hydrolases is potentially dangerous for cell survival, leading to the rupture of the cell envelope and lysis (238, 295). Although not as readily prone to autolysis as other bacterial organisms (248), E. coli cells lyse under a wide variety of conditions not always explicitly correlated with PG degradation. Here, attention is limited to autolysis induced by the inhibition of PG synthesis in growing cells, to the resistance of nongrowing cells to autolysis, and to the autolysis of harvested cells. Various aspects of the role of E. coli PG hydrolases functioning as autolysins have been reviewed (53, 99, 100, 102, 103, 264, 265).
Autolysis of Growing Cells by Inhibition of PG Synthesis
In growing E. coli cells, the steady formation and insertion of new material in the lateral wall and septum compensate for the loss of PG material by turnover. It can be speculated that any abrupt slowdown or arrest of the polymerization reactions will result in an unbalanced state where hydrolases presumably continue to cleave PG bonds unabated, leading to excessive PG degradation and cell lysis. In this case, autolysis is not due to any specific induction of autolysins but rather to an uncoupling between synthesis and degradation owing to an absence of a control of the turnover hydrolases. Autolysis by the inhibition of PG synthesis is thus a two-step process (203), with the initial one being a deficiency in PG polymerization due to the depletion of the lipid II pool or to the inhibition of the HMM PBPs. In the second step, the hydrolases involved in turnover are presumably responsible for PG degradation. This simple model is supported by the similar release of monomer PG fragments during autolysis (127) and turnover (83, 196), indicating the same combined action of LT, endopeptidase, and amidase activities. However, this does not exclude the action of hydrolases other than those of the turnover process. Another way of promoting lysis is not to inhibit PG synthesis but to favor an increase of the rate of PG degradation. This could be one cause of the lytic effect observed with the overproduction of certain hydrolases (Slt70, MltA, MltB, MltD, MltF, EnvC, AmiB, and AmiC).
Autolysis of Growing Cells by Depletion of the Lipid II Pool
In E. coli, most enzymes of the linear biosynthesis pathway leading to lipid II are unique and essential (5, 224, 286). Interference with any step has an effect on the downstream precursors and, consequently, on the lipid II pool level. This is illustrated by the autolytic effect of specific inhibitors such as DON (6-diazo-5-oxo-l-norleucine), fosfomycin, or d-cycloserine (40, 129, 153, 173). Vancomycin, which complexes lipid II, can also bring about the lysis of E. coli at high concentrations (136). Many temperature-sensitive mutations specifying enzymes of the biosynthesis pathway and auxotrophic conditional-lethal mutations involving essential metabolites (Table 2) have a phenotype of osmotic fragility. The shift of the mutants to nonpermissive conditions brings about cell lysis. More generally, the depletion of virtually any metabolite essential for a step of the biosynthesis pathway has an autolytic effect. This explains why autolysis occurs under many different circumstances. The 40 to 50% PG breakdown observed prior to cell lysis (174, 177) is consistent with the lowest PG content found to be compatible with cell viability (207). Table 2 summarizes examples of autolysis due to interference with the early steps of PG biosynthesis.
Table 2.
Examples of autolysis of E. coli by inhibition of lipid II synthesis
| Metabolic target | Inhibitor | Mutanta | Reference(s) |
|---|---|---|---|
| GlmS synthetase | glmS | 227, 309 | |
| DON | 40 | ||
| GlmM mutase | glmM | 177 | |
| GlmU synthetase | glmU | 175 | |
| MurA transferase | Fosfomycin | 173, 292 | |
| Protein A2 | 18 | ||
| d-Ala racemase | alr | 307 | |
| d-Cycloserine | 45 | ||
| Alaphosphin | 4 | ||
| d-Ala-d-Ala ligase | ddl | 153 | |
| d-Cycloserine | 45 | ||
| d-Glu racemase | murI | 59 | |
| A2pm synthesis | a2pm | 169 | |
| MurC synthetase | murC | 151 | |
| Excess l-serine | 314 | ||
| MurE synthetase | murE | 152, 178 | |
| MurF synthetase | murF | 151 | |
| Diarylquinoline | 11 | ||
| Promoter mra | mra | 87 | |
| MraY transferase | mraY | 27 | |
| Protein E | 17 | ||
| Colicin M | 65 | ||
| MurG transferase | murG | 225 | |
| murG | 174 | ||
| Vancomycin | 136 | ||
| Colicin M | 65 |
Mutant refers to a point mutation or a gene deletion.
Autolysis of Growing Cells by Inhibition of PG Polymerization
E. coli HMM PBPs are responsible for the formation of the glycan chains by transglycosylation and of the d-Ala→(d)-meso-A2pm cross-linkages by dd-transpeptidation (109, 110, 202, 251). PBPs 1a, 1b, and 1c catalyze both types of reactions, whereas PBP2 and PBP3 are limited to transpeptidation (48, 229). The inhibition of transglycosylation by moenomycin (287, 305) has a lytic effect on E. coli (129, 140, 141, 284). In particular, the PG degradation of growing cells treated with moenomycin does not exceed 30% at the onset of lysis, accompanied by an irregular attack of the PG layer (141).
The inhibition of dd-transpeptidation by β-lactams like penicillin, which has similar affinities for all five HMM PBPs, leads to lysis (44, 241). The highest rate of lysis is observable only when all PBPs are β-lactam saturated (228). The specific targeting of PBPs 1a and 1b by cefsulodin also has a lytic effect, whereas the preferential binding of amdinocillin or aztreonam to PBP2 or PBP3, respectively, has morphological effects (spherical or filamentous cells), with later lysis, at least in fast-growing cells (44, 202, 228, 241, 245). Similar results were found when singly and multiply defective mutations specifying the HMM PBPs were considered (49, 245, 250). Furthermore, the simultaneous use of two β-lactams targeting different HMM PBPs has a synergistic effect that also causes rapid lysis, even though each drug is at a sublytic concentration (85). Yet another kind of interference with PG polymerization leading to lysis was reported when inactive variants of the polymerase PBP1b were overproduced in cells with a normal level of wild-type PBP1b (171).
Morphological Changes Accompanying Autolysis of Growing Cells
Phase-contrast or electron microscopy of whole cells or isolated sacculi revealed the important morphological changes observed for E. coli prior to the onset of autolysis (53, 86, 126, 138, 141, 150, 212, 237). These changes include an increase in size, a loss of regular morphology, the appearance of protrusions (usually in the middle of the cell), filamentation, and spheroplasting.
The penicillin-induced formation of spheroplasts initially revealed the correlation between the loss of PG integrity and autolysis in E. coli (86, 138). When growing cells are treated with penicillin, the rate of PG synthesis is reduced by 55 to 75% prior to the onset of lysis (129, 184), and the rate of the loss of PG with different β-lactams does not exceed 20 to 35% (126, 141). The correlation between the β-lactam inhibition of PG transpeptidation, PG degradation by endogenous hydrolases, and the induction of autolysis was established (127, 235, 236, 237).
In penicillin-treated cells, bulges appear at the septum or at the presumed septum location, and sacculi are split at the midcell precisely into two halves (237). The lytic responses to β-lactam treatment correlate with the inhibition of lateral wall or septal PG synthesis (53, 237). An electron microscopy study of sacculi from cefsulodin-treated cells showed that lysis started earlier and was clearly faster in a strain with septal PG synthesis than in one without septal PG synthesis, and cells were cut at the septum (53). In contrast, strains with only lateral wall PG synthesis showed a general weakening of the PG network with random breakpoints. These results are consistent with the far-higher rate of septal turnover (271).
Resistance of Nongrowing Cells to Autolysis
The rate of autolysis of E. coli is highest for fast-growing cells but low or nil for nongrowing cells (24, 25, 86, 139, 164, 264). During entry into stationary phase, PG progressively undergoes considerable structural modifications. It becomes more cross-linked and has more covalently bound Lpp lipoprotein, more (l)-meso-A2pm→(d)-meso-A2pm cross-linkages, and shorter glycan chains (201, 267). It was suggested that these changes are responsible for the greater resistance to autolysis observed in the stationary phase (268). In particular, nongrowing cells are generally not prone to lysis when treated with most β-lactams and are characterized by the phenomenon of phenotypic tolerance (265).
The inhibition of the protein synthesis of growing cells by amino acid deprivation gives rise to resistance to antibiotic-induced autolysis after the arrest of growth (265). The rapid modifications of the PG structure taking place after amino acid starvation could be responsible for the resistance to autolysin attack (268). The stringent response (35) also has a role in resistance to autolysis via the inhibition of phospholipid synthesis, which is essential for both PG synthesis and autolysin activity (216, 217). However, when protein synthesis is inhibited in the absence of stringent control (the use of chloramphenicol or a relA mutant), the observed β-lactam-induced lysis is due to a too-low level of PG polymerase activity (218).
Certain β-lactams (mostly penems) can induce the lysis of amino-acid-starved cells although retaining an intact stringent response (264, 265). They all bind to PBP7, and conversely, β-lactams without a lytic effect do not bind well to PBP7 (43, 266). It can be speculated that by its enzymatic specificity for d-Ala→(d)-meso-A2pm cross-linkages (222), PBP7 endopeptidase activity has a role in the PG structural changes generating autolysis resistance at the onset of the stationary phase. Inhibition by penems could favor the maintenance of growing-phase d-Ala→(d)-meso-A2pm cross-linkages and a PG structure sensitive to lytic attack. However, the importance of the binding of penems to PBP7 is disputed by the fact that PBP7 mutants are fully viable (94). Thus, it is unclear what the binding of penems to PBP7 means as far as cell lysis is concerned. Another possibility is that penems inhibit the ld-transpeptidases responsible for forming (l)-meso-A2pm→(d)- meso-A2pm cross-linkages and thereby contribute to the maintenance of an autolysin-sensitive PG structure. A penem-sensitive ld-transpeptidase from Enterococcus faecium has been reported (160).
Autolysis of Harvested Cells
Wild-type E. coli cells generally do not lyse spontaneously at a high rate after harvesting and resuspension in an NaCl solution or buffer (90, 140, 248). Obviously, after the arrest of cell growth, PG hydrolases undergo tight control with the setup of a barrier between them and PG by an unknown mechanism. Supposedly, the harvesting of cells leads to a change in the dynamic status of the cell envelope and to the segregation of the autolysins from PG. Some kind of triggering is necessary to promote the breakdown of the barrier and the attack of PG by the autolysins (90). This is brought about by submitting harvested cells to osmotic shocks or by treating them with high-salt concentrations, sucrose, trichloroacetic acid (TCA), EGTA, EDTA, or chaotropic agents (90, 108, 139, 239). These treatments disorganize the cell envelope and lead to high rates of extensive cell lysis, with over 60 to 80% PG breakdown, as visualized by the disappearance of the PG material over the whole cell surface (141). During TCA-induced autolysis, monomer muropeptides and free peptides are released, indicating that, like with growing cells, LTs, endopeptidases, and amidases are operating (127).
Inhibitory Effect of Mg2+ Ions on Autolysis
The dramatic effect of preventing lysis by the addition of 20 mM Mg2+ was initially reported for penicillin-treated cells (168) and can be observed for many types of autolyses of both growing and harvested E. coli cells (90, 140). The mechanism of the Mg2+ effect is unknown. The importance of divalent cations in the organization of the outer membrane and their role in the stabilization of the lipopolysaccharide (LPS) monolayer portion of the outer membrane has been stressed (190). It can be speculated that the binding of Mg2+ ions to LPS induces a change of the outer membrane structure favoring the segregation of the autolysins from the normally closely associated PG layer. On the other hand, the removal of the Mg2+ ions by chelators will lead to the destabilization of the LPS monolayer, the disorganization of the outer membrane, and the uncontrolled action of autolysins.
CONCLUDING REMARKS
In E. coli, there is a hydrolytic activity for each bond linking the PG amino acids and sugars. Owing to redundancy, PG hydrolases far outnumber the enzymes of the biosynthesis pathway. The presence of two sets of highly specific enzymes acting on the same bonds raises the question of possible functional correlations or interferences. When cytoplasmic and cell envelope localizations are considered, few hydrolases, if any, are found simultaneously in both compartments, at least in nonoverproducing strains (Table 1).
The cytoplasmic hydrolases appear to act independently from the enzymes responsible for the assembly of the PG monomer unit, which are cytoplasmic or operating at the inner side of the cytoplasmic membrane (286). No interaction between them has been reported, and interferences are avoided in different ways. The hydrolases NagZ, AmpD, and MpaA, involved in recycling, have little or no action on the nucleotide precursors (196). The ld-carboxypeptidase LdcA, which prevents the deleterious accumulation of UDP-MurNAc-tetrapeptide, also has no effect on the precursors owing to a specificity limited to tetrapeptide-containing monomer derivatives (142, 256, 274). Furthermore, the dipeptidase DdpX is without an effect on the d-alanyl-d-alanine pool in growing cells produced in the stationary phase. Finally, a possible hydrolytic activity associated with an enzyme of the biosynthesis pathway has been seldom considered (63, 78) and has not been reported for E. coli. The reversibility of biosynthesis steps has been observed in a few cases but only in the presence of the reaction products (7). Thus, PG hydrolases and biosynthesis enzymes have both distinct reaction mechanisms and no overlapping functions.
The envelope-bound hydrolases localize to the outer membrane, in the periplasm, or at the outer side of the inner membrane. Few of them appear to function at the inner side of the cytoplasmic membrane, and it is thus unlikely that any envelope-bound hydrolases have an action on the strictly cytoplasm-located nucleotide precursors, which can nonetheless be used as substrates in in vitro assays. The formation of lipid precursors I and II at the inner side of the cytoplasmic membrane by the transferases MraY and MurG is characterized by their low pool levels and high turnover rates (26, 288). The importance of protecting these lipid intermediates from the potential deleterious effect of hydrolases has been stressed (37). Presumably, the direct channeling of lipid I from MraY to MurG and that of lipid II from MurG to the extracytoplasmic polymerases, perhaps via the hypothetical translocase MurJ (224), avoid their presence as free components in the membrane and their attack by hydrolases.
The way in which the hydrolase activities involved in PG maturation and turnover correlate with the PG-forming HMM PBPs remains largely unknown. Protein-protein interactions with HMM PBPs (293, 297) and subcellular localizations suggest possible associations of hydrolases with the PG-synthesizing machineries responsible for lateral wall and septal PG syntheses. However, clear evidence that PG hydrolases have an essential function in lateral wall PG synthesis is an uncertain matter (104). Although considered a possibility (48), no PG hydrolase has yet been reported to be associated with the elongation complex.
Fifty years of research work have led to the discovery of at least 35 PG hydrolases in E. coli, and perhaps more can exist, as judged by the set of PG hydrolases proposed for B. subtilis (240). Despite these efforts, our present knowledge of these activities is still fragmentary. Many questions concerning genetic, biochemical, and physiological aspects remain unanswered. It is unknown whether most PG hydrolases are produced constitutively or not. A few examples show that there are variations in hydrolase activity with the growth phase or the cell cycle. This is the case for PBP6 (31, 226), DdpX (144), and ld-carboxypeptidase (13, 14). Although nearly all PG hydrolase genes have been identified, mutational studies have led to disappointingly modest results (49, 92), since many single deficiencies have no effect on growth, morphology, or PG structure.
After purification to homogeneity, the crystal structures and reaction mechanisms of certain E. coli hydrolases have been investigated, while the study of the biochemical properties of other ones has not yet been initiated. Moreover, the in vivo levels of activity of many hydrolases remain poorly defined. Cell copy numbers are available for some PBPs (60, 243), but we have no idea at what level hydrolases identified only by their gene sequence are present in wild-type cells.
PG hydrolases are involved in various important structural modifications of PG taking place during the growth and stationary phases. The proper control of the PG hydrolases and mostly of those involved in the turnover process is critical for cell integrity and survival. Our present knowledge of the mechanisms regulating the E. coli PG hydrolases is yet limited to a few examples: the stringent control modulating Slt70 activity (22); BolA controlling the levels of transcription of PBP5 and PBP6 (226); the activation of AmiA, AmiB, and AmiC amidase activities by EnvC and NlpD (272); and the upregulation of the transcription of amiA and amiC by the CpxR/CpxA system (302).
A few hydrolases have reasonably well-defined functions. This is true for PBP5 (205), the ld-transpeptidases involved in maturation (159), and the hydrolases of the recycling process (196). The ld-transpeptidases can seemingly function as both hydrolases and transferases, a possibility not often considered for PG hydrolases (104). LT, endopeptidase, and amidase activities are clearly responsible for turnover and autolysis, but it is difficult to assess whether they fulfill other specific functions.
The exact cellular functions of many PG hydrolases remain elusive, and it is difficult to understand why E. coli keeps such a large set of PG lytic enzymes, many of which are seemingly nonessential under laboratory culture conditions. The redundancy of the PG hydrolases and subtle differences in substrate specificities between the enzymes of each family certainly reflect a large variety of as-yet-unidentified specific physiological functions. Their study will be a far more difficult challenge and a longer task than the elucidation of the steps of the PG biosynthesis pathway.
ACKNOWLEDGMENTS
This work was supported by grant UMR8619 from the Centre National de la Recherche Scientifique.
I thank Michel Arthur (Université Pierre and Marie Curie, Paris, France), Juan Ayala (Universidad Autónoma, Madrid, Spain), Didier Blanot (Université Paris-Sud, Orsay, France), Miguel de Pedro (Universidad Autónoma, Madrid, Spain), Waldemar Vollmer (University of Newcastle, Newcastle, United Kingdom), and Kevin Young (University of Arkansas, Little Rock, AR) for critical comments, suggestions, and help in improving the manuscript.
Biography

Jean van Heijenoort is an emeritus CNRS Director of Research at the University of Orsay, France. He received his M.Sc. in chemistry and physics from the University of Paris. He then joined the department of biochemistry of Edgar Lederer at Orsay, where he prepared a thesis on the chemistry of diaminopimelic acid. After a postdoctoral stay with J.-M. Ghuysen at the University of Lie`ge and one with J. T. Park at Tufts University, he returned to Orsay to head a research group on bacterial cell walls. His scientific work has focused on the structure and metabolism of peptidoglycan and on antibiotics inhibiting its biosynthesis. He is the author of over 150 research articles, review articles, and book chapters. His group has collaborated with many other academic and private laboratories.
REFERENCES
- 1. Amanuma H., Strominger J. L. 1980. Purification and properties of penicillin-binding proteins 5 and 6 from Escherichia coli membranes. J. Biol. Chem. 255: 11173–11180 [PubMed] [Google Scholar]
- 2. Amanuma H., Strominger J. L. 1984. Purification and properties of penicillin-binding proteins 5 and 6 from the dacA mutant strain of Escherichia coli (JE 11191). J. Biol. Chem. 259: 1294–1298 [PubMed] [Google Scholar]
- 3. Artola-Recolons C., et al. 2011. High-resolution crystal structure of MltE, an outer membrane-anchored endolytic peptidoglycan lytic transglycosylase from Escherichia coli. Biochemistry 50: 2384–2386 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Atherton F. R., et al. 1979. Phosphonopeptides as antibacterial agents: mechanism of action of alaphosphin. Antimicrob. Agents Chemother. 15: 696–705 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Baba T., et al. 2006. Construction of Escherichia coli K-12 in-frame, single-gene knockout mutants: the Keio collection. Mol. Syst. Biol. 2: 1–11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Baquero M.-R., Bouzon M., Quintela J. C., Ayala J. A., Moreno F. 1996. dacD, an Escherichia coli gene encoding a novel penicillin-binding protein (PBP6b) with dd-carboxypeptidase activity. J. Bacteriol. 178: 7106–7111 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Barreteau H., et al. 2008. Cytoplasmic steps of peptidoglycan biosynthesis. FEMS Microbiol. Rev. 32: 168–207 [DOI] [PubMed] [Google Scholar]
- 8. Barrett D., et al. 2007. Analysis of glycan polymers by peptidoglycan glycosyltransferases. J. Biol. Chem. 282: 31964–31971 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Bateman A., Bycroft M. 2000. The structure of a LysM domain from E. coli membrane-bound lytic murein transglycosylase D (MltD). J. Mol. Biol. 299: 1113–1119 [DOI] [PubMed] [Google Scholar]
- 10. Baum E. Z., et al. 2005. Identification of a dithiazoline inhibitor of Escherichia coli l,d-carboxypeptidase A. Antimicrob. Agents Chemother. 49: 4500–4507 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Baum E. Z., et al. 2009. MurF inhibitors with antibacterial activity: effect on muropeptide levels. Antimicrob. Agents Chemother. 53: 3240–3247 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Beachey E. H., Keck W., de Pedro M. A., Schwarz U. 1981. Exoenzymatic activity of transglycosylase isolated from Escherichia coli. Eur. J. Biochem. 116: 355–358 [DOI] [PubMed] [Google Scholar]
- 13. Beck B. D., Park J. T. 1976. Activity of three murein hydrolases during the cell division cycle of Escherichia coli K-12 as measured in toluene-treated cells. J. Bacteriol. 126: 1250–1260 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Beck B. D., Park J. T. 1977. Basis for the observed fluctuation of carboxypeptidase II activity during the cell cycle in BUG 6, a temperature-sensitive division mutant of Escherichia coli. J. Bacteriol. 130: 1292–1302 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Begg K. J., et al. 1990. The balance between different peptidoglycan precursors determines whether Escherichia coli cells will elongate or divide. J. Bacteriol. 172: 6697–6703 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Bernadsky G., Beveridge T. J., Clarke A. J. 1994. Analysis of the sodium dodecyl sulfate-stable peptidoglycan autolysins of select gram-negative pathogens by using renaturing polyacrylamide gel electrophoresis. J. Bacteriol. 176: 5225–5232 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Bernhardt T. G., Struck D. K., Young R. 2001. The lysis protein E of φX174 is a specific inhibitor of the MraY-catalyzed step in peptidoglycan synthesis. J. Biol. Chem. 276: 6093–6097 [DOI] [PubMed] [Google Scholar]
- 18. Bernhardt T. G., Wang I.-N., Struck D. K., Young R. 2001. A protein antibiotic in the phage Qβ virion: diversity in lysis targets. Science 292: 2326–2329 [DOI] [PubMed] [Google Scholar]
- 19. Bernhardt T. G., de Boer P. A. J. 2003. The Escherichia coli amidase AmiC is a periplasmic septal ring component exported via the twin-arginine transport pathway. Mol. Microbiol. 48: 1171–1182 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Bernhardt T. G., de Boer P. A. J. 2004. Screening for synthetic lethal mutants in Escherichia coli and identification of EnvC (YibP) as a periplasmic septal ring factor with murein hydrolase activity. Mol. Microbiol. 52: 1255–1269 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Betzner A. S., Keck W. 1989. Molecular cloning, overexpression and mapping of the slt gene encoding the soluble lytic transglycosylase of Escherichia coli. Mol. Gen. Genet. 219: 489–491 [DOI] [PubMed] [Google Scholar]
- 22. Betzner A. S., Ferreira L. C. S., Höltje J.-V., Keck W. 1990. Control of the activity of the soluble lytic transglycosylase by the stringent response in Escherichia coli. FEMS Microbiol. Lett. 67: 161–164 [DOI] [PubMed] [Google Scholar]
- 23. Blackburn N. T., Clarke A. J. 2001. Identification of four families of peptidoglycan lytic transglycosylases. J. Mol. Evol. 52: 78–84 [DOI] [PubMed] [Google Scholar]
- 24. Blasco B., Pisabarro A. G., de Pedro M. A. 1988. Peptidoglycan biosynthesis in stationary-phase cells of Escherichia coli. J. Bacteriol. 170: 5224–5228 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Boman H. G., Eriksson K. G. 1963. Penicillin induced lysis in Escherichia coli. J. Gen. Microbiol. 31: 339–352 [DOI] [PubMed] [Google Scholar]
- 26. Bouhss A., Trunkfield A. E., Bugg T. D. H., Mengin-Lecreulx D. 2008. The biosynthesis of peptidoglycan lipid-linked intermediates. FEMS Microbiol. Rev. 32: 208–233 [DOI] [PubMed] [Google Scholar]
- 27. Boyle D. S., Donachie W. D. 1998. mraY is an essential gene for cell growth in Escherichia coli. J. Bacteriol. 180: 6429–6432 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Broome-Smith J. K., Spratt B. G. 1982. Deletion of the penicillin-binding protein 6 gene of Escherichia coli. J. Bacteriol. 152: 904–906 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Broome-Smith J. K. 1985. Construction of a mutant of Escherichia coli that has deletions of both the penicillin-binding protein 5 and 6 genes. J. Gen. Microbiol. 131: 2115–2118 [DOI] [PubMed] [Google Scholar]
- 30. Broome-Smith J. K., Ioannidis I., Edelman A., Spratt B. G. 1988. Nucleotide sequences of the penicillin-binding protein 5 and 6 genes of Escherichia coli. Nucleic Acids Res. 16: 1617. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Buchanan C. E., Sowell M. O. 1982. Synthesis of penicillin-binding protein 6 by stationary-phase Escherichia coli. J. Bacteriol. 151: 491–494 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Buissiere D. E., et al. 1998. The structure of VanX reveals a novel amino-dipeptidase involved in mediating transposon-based vancomycin resistance. Mol. Cell 2: 75–84 [DOI] [PubMed] [Google Scholar]
- 33. Burman L. G., Park J. T. 1983. Changes in the composition of Escherichia coli murein as it ages during exponential growth. J. Bacteriol. 155: 447–453 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Burman L. G., Park J. T. 1984. Molecular model for elongation of the murein sacculus of Escherichia coli. Proc. Natl. Acad. Sci. U. S. A. 81: 1844–1848 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Cashel M., Gentry D. R., Hernandez V. J., Vinella D. 1996. The stringent response, p. 1459–1496 In Neidhardt F. C., et al. (ed.), Escherichia coli and Salmonella typhimurium: cellular and molecular biology. ASM Press, Washington, DC [Google Scholar]
- 36. Chen Y., et al. 2009. Crystal structures of penicillin-binding protein 6 from Escherichia coli. J. Am. Chem. Soc. 131: 14345–14354 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Cheng Q., Li H., Merdek K., Park J. T. 2000. Molecular characterization of the β-N-acetylglucosaminidase of Escherichia coli and its role in cell wall recycling. J. Bacteriol. 182: 4836–4840 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Cheng Q., Park J. T. 2002. Substrate specificity of the AmpG permease required for recycling of cell wall anhydro-muropeptides. J. Bacteriol. 184: 6434–6436 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Clarke T. B., et al. 2009. Mutational analysis of the substrate specificity of Escherichia coli penicillin binding protein 4. Biochemistry 48: 2675–2683 [DOI] [PubMed] [Google Scholar]
- 40. Coggin J. H., Jr., Martin W. R. 1965. 6-Diazo-5-oxo-l-norleucine inhibition of Escherichia coli. J. Bacteriol. 89: 1348–1353 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Cooper S., Hsieh M. L. 1988. The rate and topography of cell wall synthesis during the division cycle of Escherichia coli using N-acetylglucosamine as a peptidoglycan label. J. Gen. Microbiol. 134: 1717–1721 [DOI] [PubMed] [Google Scholar]
- 42. Cowles C. E., Li Y., Semmelhack M. F., Cristea I. M., Silhavy T. J. 2011. The free and bound forms of Lpp occupy distinct cellular locations in Escherichia coli. Mol. Microbiol. 79: 1168–1181 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Cozens R. M., Markiewicz Z., Tuomanen E. 1989. Role of autolysins in the activities of imipenem and CGP 31608, a novel penem, against slowly growing bacteria. Antimicrob. Agents Chemother. 33: 1819–1821 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Curtis N. A. C., Orr D., Ross G. W., Boulton M. G. 1979. Affinities of penicillins and cephalosporins for the penicillin-binding proteins of Escherichia coli K-12 and their antibacterial activity. Antimicrob. Agents Chemother. 16: 533–539 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Curtiss R., III, Charamella L. J., Berg C. M., Harris P. E. 1965. Kinetic and genetic analyses of d-cycloserine inhibition and resistance in Escherichia coli. J. Bacteriol. 90: 1238–1250 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Davies C., White S. W., Nicholas R. A. 2001. Crystal structure of a deacylation-defective mutant of penicillin-binding protein 5 at 2.3-Å resolution. J. Biol. Chem. 276: 616–623 [DOI] [PubMed] [Google Scholar]
- 47. de Jonge B. L. M., et al. 1989. Peptidoglycan synthesis during the cell cycle of Escherichia coli: composition and mode of insertion. J. Bacteriol. 171: 5783–5794 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. den Blaauwen T., de Pedro M. A., Nguyen-Distèche M., Ayala J. A. 2008. Morphogenesis of rod-shaped sacculi. FEMS Microbiol. Rev. 32: 321–344 [DOI] [PubMed] [Google Scholar]
- 49. Denome S. A., Elf P. K., Henderson T. A., Nelson D. E., Young K. D. 1999. Escherichia coli mutants lacking all possible combinations of eight penicillin-binding proteins: viability, characteristics, and implications for peptidoglycan synthesis. J. Bacteriol. 181: 3981–3993 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. de Pedro M. A., Schwarz U., Nishimura Y., Hirota Y. 1980. On the biological role of penicillin-binding proteins 4 and 5. FEMS Microbiol. Lett. 9: 219–221 [Google Scholar]
- 51. de Pedro M. A., Schwarz U. 1981. Heterogeneity of newly inserted and preexisting murein in the sacculus of Escherichia coli. Proc. Natl. Acad. Sci. U. S. A. 78: 5856–5860 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. de Pedro M. A., Quintela J. C., Höltje J.-V., Schwarz H. 1997. Murein segregation in Escherichia coli. J. Bacteriol. 179: 2823–2834 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. de Pedro M. A., Höltje J.-V., Schwarz H. 2002. Fast lysis of Escherichia coli filament cells requires differentiation of potential division sites. Microbiology 148: 79–86 [DOI] [PubMed] [Google Scholar]
- 54. Depuydt M., et al. 2009. A periplasmic reducing system protects single cysteine residues from oxidation. Science 326: 1109–1111 [DOI] [PubMed] [Google Scholar]
- 55. Diestra E., Fontana J., Guichard P., Marco S., Risco C. 2009. Visualization of proteins in intact cells with a clonable tag for electron microscopy. J. Struct. Biol. 165: 157–168 [DOI] [PubMed] [Google Scholar]
- 56. Dijkstra A. J., Hermann F., Keck W. 1995. Cloning and controlled overexpression of the gene encoding the 35 kDa soluble lytic transglycosylase from Escherichia coli. FEBS Lett. 366: 115–118 [DOI] [PubMed] [Google Scholar]
- 57. Dijkstra A. J., Keck W. 1996. Identification of new members of the lytic transglycosylase family in Haemophilus influenzae and Escherichia coli. Microb. Drug Resist. 2: 141–145 [DOI] [PubMed] [Google Scholar]
- 58. Dijkstra A. J. 1997. The lytic transglycosylase family of Escherichia coli. Ph.D. thesis Rijksuniversiteit Groningen, Groningen, Netherlands [Google Scholar]
- 59. Doublet P., van Heijenoort J., Bohin J.-P., Mengin-Lecreulx D. 1993. The murI gene of Escherichia coli is an essential gene that encodes a glutamate racemase activity. J. Bacteriol. 175: 2970–2979 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60. Dougherty T. J., Kennedy K., Kessler R. E., Pucci M. J. 1996. Direct quantitation of the number of individual penicillin-binding proteins per cell in Escherichia coli. J. Bacteriol. 178: 6110–6115 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61. Dramsi S., Magnet S., Davison S., Arthur M. 2008. Covalent attachment of proteins to peptidoglycan. FEMS Microbiol. Rev. 32: 307–320 [DOI] [PubMed] [Google Scholar]
- 62. Edwards D. H., Donachie W. D. 1993. Construction of a triple deletion of penicillin-binding proteins 4, 5, and 6 in Escherichia coli, p. 369–374 In Pedro M. A. de., Höltje J.-V., Löffelhardt W. (ed.), Bacterial growth and lysis. Plenum Press, New York, NY [Google Scholar]
- 63. Egan A., Lawrence P., Strominger J. L. 1973. Enzymatic synthesis of the peptide in bacterial uridine nucleotides. V. Co ++ dependent reversal of peptide bond formation. J. Biol. Chem. 248: 3122–3130 [PubMed] [Google Scholar]
- 64. Ehlert K., Höltje J.-V., Templin M. F. 1995. Cloning and expression of a murein hydrolase lipoprotein from Escherichia coli. Mol. Microbiol. 16: 761–768 [DOI] [PubMed] [Google Scholar]
- 65. El Ghachi M., et al. 2006. Colicin M exerts its bacteriolytic effect via enzymatic degradation of undecaprenyl phosphate-linked peptidoglycan precursors. J. Biol. Chem. 281: 22761–22772 [DOI] [PubMed] [Google Scholar]
- 66. Engel H., Kazemier B., Keck W. 1991. Murein-metabolizing enzymes from Escherichia coli: sequence analysis and controlled overexpression of the slt gene, which encodes the soluble lytic transglycosylase. J. Bacteriol. 173: 6773–6782 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67. Engel H., Smink A. J., van Wijngaarden L., Keck W. 1992. Murein-metabolizing enzymes from Escherichia coli: existence of a second lytic transglycosylase. J. Bacteriol. 174: 6394–6403 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68. Ferreira L. C., et al. 1988. Properties and crystallization of a genetically engineered, water-soluble derivative of penicillin-binding protein 5 of Escherichia coli K12. Eur. J. Biochem. 171: 11–16 [DOI] [PubMed] [Google Scholar]
- 69. Firczuk M., Bochtler M. 2007. Mutational analysis of peptidoglycan amidase MepA. Biochemistry 46: 120–128 [DOI] [PubMed] [Google Scholar]
- 70. Firczuk M., Bochtler M. 2007. Folds and activitues of peptidoglycan amidases. FEMS Microbiol. Rev. 31: 676–691 [DOI] [PubMed] [Google Scholar]
- 71. Gale E. F., Cundiffe E., Reynolds P. E., Richmond M. H., Warning M. J. 1981. The molecular basis of antibiotic action. John Wiley & Sons, London, United Kingdom [Google Scholar]
- 72. Ghosh A. S., Chowdhury C., Nelson D. E. 2008. Physiological functions of D-alanine carboxypeptidases in Escherichia coli. Trends Microbiol. 16: 309–317 [DOI] [PubMed] [Google Scholar]
- 73. Gittins J. R., Phoenix D. A., Pratt J. M. 1994. Multiple mechanisms of membrane anchoring of Escherichia coli penicillin-binding proteins. FEMS Microbiol. Rev. 13: 1–12 [DOI] [PubMed] [Google Scholar]
- 74. Glauner B., Höltje J.-V., Schwarz U. 1988. The composition of the murein of Escherichia coli. J. Biol. Chem. 263: 10088–10095 [PubMed] [Google Scholar]
- 75. Glauner B., Höltje J.-V. 1990. Growth pattern of the murein sacculus of Escherichia coli. J. Biol. Chem. 265: 18988–18996 [PubMed] [Google Scholar]
- 76. Goffin C., Ghuysen J.-M. 1998. Multimodular penicillin-binding proteins: an enigmatic family of orthologs and paralogs. Microbiol. Mol. Biol. Rev. 62: 1079–1093 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77. Goffin C., Ghuysen J.-M. 2002. Biochemistry and comparative genomics of SxxK superfamily acyltransferases offer a clue to the mycobacterial paradox: presence of penicillin-susceptible target proteins versus lack of efficiency of penicillin as therapeutic agent. Microbiol. Mol. Biol. Rev. 66: 702–738 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78. Gondré B., Flouret B., van Heijenoort J. 1973. Release of D-alanyl-D-alanine from the precursor of the cell wall peptidoglycan by a peptidase of Escherichia coli K12. Biochimie 55: 685–691 [DOI] [PubMed] [Google Scholar]
- 79. Gonzalez-Leiza S., de Pedro M. A., Ayala J. A. 14 October 2011. AmpH, a bifunctional DD-endopeptidase and DD-carboxypeptidase of Escherichia coli. J. Bacteriol. doi: 10.1128/JB.05764-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80. Goodell E. W., Schwarz U. 1977. Enzymes synthesizing and hydrolyzing murein in Escherichia coli. Topographical distribution over the cell envelope. Eur. J. Biochem. 81: 205–210 [DOI] [PubMed] [Google Scholar]
- 81. Goodell E. W., Markiewicz Z., Schwarz U. 1983. Absence of oligomeric murein intermediates in Escherichia coli. J. Bacteriol. 156: 130–135 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82. Goodell E. W., Schwarz U. 1985. Release of cell wall peptides into culture medium by exponentially growing Escherichia coli. J. Bacteriol. 162: 391–397 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83. Goodell E. W. 1985. Recycling of murein by Escherichia coli. J. Bacteriol. 163: 305–310 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84. Gulick A. M., Schmidt D. M. Z., Gerlt J. A., Rayment I. 2001. Evolution of enzymatic activities in the enolase superfamily: crystal structures of the L-Ala-D/L-Glu epimerases from Escherichia coli and Bacillus subtilis. Biochemistry 40: 15716–15724 [DOI] [PubMed] [Google Scholar]
- 85. Gutmann L., et al. 1986. Involvement of penicillin-binding protein 2 with other penicillin-binding proteins in lysis of Escherichia coli by some β-lactam antibiotics alone and in synergistic lytic effect of amdinocillin (mecillinam). Antimicrob. Agents Chemother. 30: 906–912 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86. Hahn F. E., Ciak J. 1957. Penicillin-induced lysis of Escherichia coli. Science 125: 119–120 [DOI] [PubMed] [Google Scholar]
- 87. Hara H., Yasuda S., Horiuchi K., Park J. T. 1997. A promoter for the first nine genes of the Escherichia coli mra cluster of cell division and cell envelope biosynthesis genes, including ftsI and ftsW. J. Bacteriol. 179: 5802–5811 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88. Hara H., et al. 2002. Identification and characterization of the Escherichia coli envC gene encoding a periplasmic coiled-coil protein with putative peptidase activity. FEMS Microbiol. Lett. 212: 229–236 [DOI] [PubMed] [Google Scholar]
- 89. Hartmann R., Höltje J.-V., Schwarz U. 1972. Targets of penicillin action in Escherichia coli. Nature 235: 426–429 [DOI] [PubMed] [Google Scholar]
- 90. Hartmann R., Bock-Hennig S. B., Schwarz U. 1974. Murein hydrolases in the envelope of Escherichia coli. Properties in situ and solubilization from the envelope. Eur. J. Biochem. 41: 203–208 [DOI] [PubMed] [Google Scholar]
- 91. Heidrich C., et al. 2001. Involvement of N-acetylmuramyl-L-alanine amidases in cell separation and antibiotic-induced autolysis of Escherichia coli. Mol. Microbiol. 41: 167–178 [DOI] [PubMed] [Google Scholar]
- 92. Heidrich C., Ursinus A., Berger J., Schwarz H., Höltje J.-V. 2002. Effects of mutiple deletions of murein hydrolases on viability, septum cleavage, and sensitivity to large toxic molecules in Escherichia coli. J. Bacteriol. 184: 6093–6099 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93. Henderson T. A., Dombrosky P. M., Young K. D. 1994. Artifactual processing of penicillin-binding proteins 7 and 1b by the OmpT protease of Escherichia coli. J. Bacteriol. 176: 256–259 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94. Henderson T. A., Templin M., Young K. D. 1995. Identification and cloning of the gene encoding penicillin-binding protein 7 of Escherichia coli. J. Bacteriol. 177: 2074–2079 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95. Henderson T. A., Young K. D., Denome S. A., Elf P. K. 1997. AmpC and AmpH, proteins related to the class C β-lactamases, bind penicillin and contribute to the normal morphology of Escherichia coli. J. Bacteriol. 179: 6112–6121 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96. Hervé M., Boniface A., Gobec S., Blanot D., Mengin-Lecreulx D. 2007. Biochemical characterization and physiological properties of Escherichia coli UDP-N-acetylmuramate:l-alanyl-γ-d-glutamyl-meso-diaminopimelate ligase. J. Bacteriol. 189: 3987–3995 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97. Hesek D., Lee M., Zhang W., Noll B. C., Mobashery S. 2009. Total synthesis of N-acetylglucosamine-1,6-anhydro-N-acetylmuramylpentapeptide and evaluation of its turnover by AmpD from Escherichia coli. J. Am. Chem. Soc. 131: 5187–5193 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98. Höltje J.-V., Mirelman D., Sharon N., Schwarz U. 1975. Novel type of murein transglycosylase in Escherichia coli. J. Bacteriol. 124: 1067–1076 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99. Höltje J.-V., Schwarz U. 1985. Biosynthesis and growth of the murein sacculus, p. 77–119 In Nanninga N. (ed.), Molecular cytology of Escherichia coli. Academic Press, London, United Kingdom [Google Scholar]
- 100. Höltje J.-V., Tuomanen E. 1991. The murein hydrolases of Escherichia coli: properties, functions and impact on the course of infections in vivo. J. Gen. Microbiol. 137: 441–454 [DOI] [PubMed] [Google Scholar]
- 101. Höltje J.-V., Kopp U., Ursinus A., Wiedemann B. 1994. The negative regulator of β-lactamase induction AmpD is a N-acetyl-anhydromuramyl-L-alanine amidase. FEMS Microbiol. Lett. 122: 159–164 [DOI] [PubMed] [Google Scholar]
- 102. Höltje J.-V. 1995. From growth to autolysis: the murein hydrolases in Escherichia coli. Arch. Microbiol. 164: 243–254 [DOI] [PubMed] [Google Scholar]
- 103. Höltje J.-V. 1998. Growth of the stress-bearing and shape-maintaining murein sacculus of Escherichia coli. Microbiol. Mol. Biol. Rev. 62: 181–203 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104. Höltje J.-V., Heidrich C. 2001. Enzymology of elongation and constriction of the murein sacculus of Escherichia coli. Biochimie 83: 103–108 [DOI] [PubMed] [Google Scholar]
- 105. Hourdou M.-L., et al. 1993. Characterization of the sporulation-related γ-D-glutamyl-(L)meso-diaminopimelic-acid-hydrolysing peptidase I of Bacillus sphaericus NCTC 9602 as a member of the metallo(zinc) carboxypeptidase A family. Modular design of the protein. Biochem. J. 292: 563–570 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106. Ichimura T., Yamazoe M., Maeda M., Wada C., Hiraga S. 2002. Proteolytic activity of YibP protein in Escherichia coli. J. Bacteriol. 184: 2595–2602 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107. Iida K., Hirota Y., Schwarz U. 1983. Mutants of Escherichia coli defective in penicillin-insensitive murein DD-endopeptidase. Mol. Gen. Genet. 189: 215–221 [DOI] [PubMed] [Google Scholar]
- 108. Ingram L. O. 1981. Mechanism of lysis of Escherichia coli by ethanol and other chaotropic agents. J. Bacteriol. 146: 331–336 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109. Ishino F., Mitsui K., Tamaki S., Matsuhashi M. 1980. Dual enzyme activities of cell wall peptidoglycan synthesis, peptidoglycan transglycosylase and penicillin-sensitive peptidoglycan transpeptidase, in purified preparations of Escherichia coli penicillin-binding protein 1A. Biochem. Biophys. Res. Commun. 97: 287–293 [DOI] [PubMed] [Google Scholar]
- 110. Ishino F., Tamaki S., Spratt B. G., Matsuhashi M. 1982. A mecillinam-sensitive crosslinking reaction in Escherichia coli. Biochem. Biophys. Res. Commun. 109: 689–696 [DOI] [PubMed] [Google Scholar]
- 111. Iwaya M., Strominger J. L. 1977. Simultaneous deletion of D-alanine carboxypeptidase IB-C and penicillin-binding component IV in a mutant of Escherichia coli K12. Proc. Natl. Acad. Sci. U. S. A. 74: 2980–2984 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112. Izaki K., Strominger J. L. 1968. Biosynthesis of the peptidoglycan of bacterial cell walls. XIV. Purification and properties of two D-alanine carboxypeptidases from Escherichia coli. J. Biol. Chem. 243: 3193–3201 [PubMed] [Google Scholar]
- 113. Ize B., Stanley N. R., Buchanan G., Palmer T. 2003. Role of the Escherichia coli Tat pathway in outer membrane integrity. Mol. Microbiol. 48: 1183–1193 [DOI] [PubMed] [Google Scholar]
- 114. Jackson M. E., Pratt J. M. 1987. An 18 amino acid amphiphilic helix forms the membrane-anchoring domain of the Escherichia coli penicillin-binding protein 5. Mol. Microbiol. 1: 23–28 [DOI] [PubMed] [Google Scholar]
- 115. Jackson M. E., Pratt J. M. 1988. Analysis of the membrane-binding domain of penicillin-binding protein 5 of Escherichia coli. Mol. Microbiol. 2: 563–568 [DOI] [PubMed] [Google Scholar]
- 116. Jacobs C., et al. 1995. AmpD, essential for both β-lactamase regulation and cell wall recycling, is a novel cytosolic N-acetylmuramyl-L-alanine amidase. Mol. Microbiol. 15: 553–559 [DOI] [PubMed] [Google Scholar]
- 117. Jacoby G. H., Young K. D. 1988. Unequal distribution of penicillin-binding proteins among inner membrane vesicles of Escherichia coli. J. Bacteriol. 170: 3660–3667 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118. Jaeger T., Arsic M., Mayer C. 2005. Scission of the lactyl ether bond of N-acetylmuramic acid by Escherichia coli “etherase.” J. Biol. Chem. 280: 30100–30106 [DOI] [PubMed] [Google Scholar]
- 119. Jaeger T., Mayer C. 2008. N-Acetylmuramic acid 6-phosphate lyases (MurNAc etherases): role in cell wall metabolism, distribution, structure, and mechanism. Cell. Mol. Life Sci. 65:928–939 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120. Karibian D., Pellon G., Starka J. 1981. Autolysis of a division mutant of Escherichia coli. J. Gen. Microbiol. 126: 55–61 [DOI] [PubMed] [Google Scholar]
- 121. Karibian D., Michel G. P. F., Starka J. 1985. The EnvC phenotype and its expression in various Escherichia coli K12 strains. FEMS Microbiol. Lett. 27: 319–324 [Google Scholar]
- 122. Keck W., Schwarz U. 1979. Escherichia coli murein-dd-endopeptidase insensitive to β-lactam antibiotics. J. Bacteriol. 139: 770–774 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123. Keck W., van Leeuwen A. M., Huber M., Goodell E. W. 1990. Cloning and characterization of mepA, the structural gene of the penicillin-insensitive murein endopeptidase from Escherichia coli. Mol. Microbiol. 4: 209–219 [DOI] [PubMed] [Google Scholar]
- 124. Kerff F., et al. 2010. Specific structural features of the N-acetylmuramoyl-L-alanine amidase AmiD from Escherichia coli and mechanistic implications for enzymes of this family. J. Mol. Biol. 397: 249–259 [DOI] [PubMed] [Google Scholar]
- 125. Kishida H., et al. 2006. Crystal structure of penicillin binding protein 4 (dacB) from Escherichia coli, both in the native form and covalently linked to various antibiotics. Biochemistry 45: 783–792 [DOI] [PubMed] [Google Scholar]
- 126. Kitano K., Tomasz A. 1979. Triggering of autolytic cell wall degradation in Escherichia coli by beta-lactam antibiotics. Antimicrob. Agents Chemother. 16: 838–848 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127. Kitano K., Tuomanen E., Tomasz A. 1986. Transglycosylase and endopeptidase participate in the degradation of murein during autolysis of Escherichia coli. J. Bacteriol. 167: 759–765 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128. Klenchin V. A., Schmidt D. M., Gerlt J. A., Rayment I. 2004. Evolution of enzymatic activities in the enolase superfamily: structure of a substrate-liganded complex of the L-Ala-D/L-Glu epimerase from Bacillus subtilis. Biochemistry 43: 10370–10378 [DOI] [PubMed] [Google Scholar]
- 129. Kohlrausch U., Höltje J.-V. 1991. Analysis of murein and murein precursors during antibiotic-induced lysis of Escherichia coli. J. Bacteriol. 173: 3425–3431 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130. Korat B., Mottl H., Keck W. 1991. Penicillin-binding protein 4 of Escherichia coli: molecular cloning of the dacB gene, controlled overexpression, and alterations in murein composition. Mol. Microbiol. 5: 675–684 [DOI] [PubMed] [Google Scholar]
- 131. Korsak D., Liebscher S., Vollmer W. 2005. Susceptibility to antibiotics and β-lactamase induction in murein hydrolase mutants of Escherichia coli. Antimicrob. Agents Chemother. 49: 1404–1409 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132. Kraft A. R., Templin M. F., Höltje J.-V. 1998. Membrane-bound lytic transglycosylase in Escherichia coli. J. Bacteriol. 180: 3441–3447 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133. Kraft A. R., Prabhu J., Ursinus A., Höltje J.-V. 1999. Interference with murein turnover has no effect on growth but reduces β-lactamse induction in Escherichia coli. J. Bacteriol. 181: 7192–7198 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134. Kraus W., Höltje J.-V. 1987. Two distinct transpeptidation reactions during murein synthesis in Escherichia coli. J. Bacteriol. 169: 3099–3103 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135. Kusser W., Schwarz U. 1980. Escherichia coli murein transglycosylase. Purification by affinity chromatography and interaction with polynucleotides. Eur. J. Biochem. 103: 277–281 [DOI] [PubMed] [Google Scholar]
- 136. Lara B., Mengin-Lecreulx D., Ayala J. A., van Heijenoort J. 2005. Peptidoglycan precursor pools associated with MraY and FtsW deficiencies or antibiotic treatments. FEMS Microbiol. Lett. 250: 195–200 [DOI] [PubMed] [Google Scholar]
- 137. Leclerc D., Asselin A. 1989. Detection of bacterial cell wall hydrolases after denaturing polyacrylamide gel electrophoresis. Can. J. Microbiol. 35: 749–753 [DOI] [PubMed] [Google Scholar]
- 138. Lederberg J. 1956. Bacterial protoplasts induced by penicillin. Proc. Natl. Acad. Sci. U. S. A. 42: 574–577 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139. Leduc M., van Heijenoort J. 1980. Autolysis of Escherichia coli. J. Bacteriol. 142: 52–59 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140. Leduc M., Kasra R., van Heijenoort J. 1982. Induction and control of the autolytic system of Escherichia coli. J. Bacteriol. 152: 26–34 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141. Leduc M., Frehel C., van Heijenoort J. 1985. Correlation between degradation and ultrastructure of peptidoglycan during autolysis of Escherichia coli. J. Bacteriol. 161: 627–635 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142. Leguina J. I., Quintela J. C., de Pedro M. A. 1994. Substrate specificity of Escherichia coli ld-carboxypeptidase on biosynthetically modified muropeptides. FEBS Lett. 339: 249–252 [DOI] [PubMed] [Google Scholar]
- 143. Leidenix M. J., Jacoby G. H., Henderson T. A., Young K. D. 1989. Separation of Escherichia coli penicillin-binding proteins into different membrane vesicles by agarose electrophoresis and sizing chromatography. J. Bacteriol. 171: 5680–5686 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144. Lessard I. A. D., et al. 1998. Homologs of the vancomycin resistance d-Ala-d-Ala dipeptidase VanX in Streptomyces toyocaensis, Escherichia coli and Synechocystis: attributes of catalytic efficiency, stereoselectivity and regulation with implications for function. Chem. Biol. 5: 489–504 [DOI] [PubMed] [Google Scholar]
- 145. Lessard I. A. D., Walsh C. T. 1999. VanX, a bacterial D-alanyl-D-alanine dipeptidase: resistance, immunity, or survival function? Proc. Natl. Acad. Sci. U. S. A. 96: 11028–11032 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146. Li S.-Y., Höltje J.-V., Young K. D. 2004. Comparison of high-performance liquid chromatography and fluorophore-assisted carbohydrate electrophoresis methods for analyzing peptidoglycan composition of Escherichia coli. Anal. Biochem. 326: 1–12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147. Liepinsh E., Généreux C., Dehareng D., Joris B., Otting G. 2003. NMR structure of Citrobacter freundii AmpD, comparison with bacteriophage T7 lysozyme and homology with PGRP domains. J. Mol. Biol. 327: 833–842 [DOI] [PubMed] [Google Scholar]
- 148. Lindquist S., Galleni M., Lindberg F., Normark S. 1989. Signalling proteins in enterobacterial AmpC β-lactamase regulation. Mol. Microbiol. 3: 1091–1102 [DOI] [PubMed] [Google Scholar]
- 149. Lommatzsch J., Templin M. F., Kraft A. R., Vollmer W., Höltje J.-V. 1997. Outer membrane localization of murein hydrolases: MltA, a third lipoprotein lytic transglycosylase in Escherichia coli. J. Bacteriol. 179: 5465–5470 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150. Lounatmaa K., Sarvas M., Wartiovaara J. 1974. Ultrastructure of the envelope during glucosamine starvation and penicillin treatment of a mutant of Escherichia coli. J. Gen. Microbiol. 83: 199–203 [DOI] [PubMed] [Google Scholar]
- 151. Lugtenberg E. J. J., van Schijndel-van Dam A. 1972. Temperature-sensitive mutants of Escherichia coli K12 with low activities of the L-alanine adding enzyme and the D-alanyl-D-alanine adding enzyme. J. Bacteriol. 110: 35–40 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152. Lugtenberg E. J. J., van Schijndel-van Dam A. 1972. Temperature-sensitive mutants of Escherichia coli K12 with low activity of the diaminopimelic acid adding enzyme. J. Bacteriol. 110: 41–46 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153. Lugtenberg E. J. J., van Schijndel-van Dam A. 1973. Temperature-sensitive mutant of Escherichia coli K12 with an impaired d-alanine:d-alanine ligase. J. Bacteriol. 113: 96–104 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154. Lupoli T. J., et al. 2009. Studying a cell division amidase using defined peptidoglycan substrates. J. Am. Chem. Soc. 131: 18230–18231 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155. Maass D., Pelzer H., Weidel W. 1964. Reinigung, Eigenschaften und Substratspezifität einer N-Acetylglucosaminidase aus E. coli B. Z. Naturforsch. B 19: 413–418 [DOI] [PubMed] [Google Scholar]
- 156. Macheboeuf P., Contreras-Martel C., Job V., Dideberg O., Dessen A. 2006. Penicillin binding proteins: key players in bacterial cell cycle and drug resistance processes. FEMS Microbiol. Rev. 30: 673–691 [DOI] [PubMed] [Google Scholar]
- 157. Magnet S., et al. 2007. Identification of the l,d-transpeptidases responsible for attachment of the Braun lipoprotein to Escherichia coli peptidoglycan. J. Bacteriol. 189: 3927–3931 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158. Magnet S., et al. 2007. Specificity of l,d-transpeptidases from gram-positive bacteria producing different peptidoglycan chemotypes. J. Biol. Chem. 282: 13151–13159 [DOI] [PubMed] [Google Scholar]
- 159. Magnet S., Dubost L., Marie A., Arthur M., Gutmann L. 2008. Identification of the l,d-transpeptidases for peptidoglycan cross-linking in Escherichia coli. J. Bacteriol. 190: 4782–4785 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160. Mainardi J.-L., et al. 2007. Unexpected inhibition of peptidoglycan l,d-transpeptidase from Enterococcus faecium by the β-lactam imipenem. J. Biol. Chem. 282: 30414–30422 [DOI] [PubMed] [Google Scholar]
- 161. Marcyjaniak M., Odintsov S. G., Sabala I., Bochtler M. 2004. Peptidoglycan amidase MepA is a LAS metallopeptidase. J. Biol. Chem. 279: 43982–43989 [DOI] [PubMed] [Google Scholar]
- 162. Markiewicz Z., Broome-Smith J. K., Schwarz U., Spratt B. G. 1982. Spherical E. coli due to elevated levels of d-alanine carboxypeptidase. Nature 297: 702–704 [DOI] [PubMed] [Google Scholar]
- 163. Massova L., Mobashery S. 1998. Kinship and diversification of bacterial penicillin-binding proteins and β-lactamases. Antimicrob. Agents Chemother. 42: 1–17 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164. Mathys E., van Gool A. 1979. Sensitivity of Escherichia coli to cephaloridine at different growth rates. J. Bacteriol. 138: 642–646 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165. Matsuhashi M., et al. 1977. Mutants of Escherichia coli lacking in highly penicillin-sensitive d-alanine carboxypeptidase activity. Proc. Natl. Acad. Sci. U. S. A. 74: 2976–2979 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166. Matsuhashi M., Tamaki S., Curtis S. J., Strominger J. L. 1979. Mutational evidence for identity of penicillin-binding protein 5 in Escherichia coli with the major d-alanine carboxypeptidase IA activity. J. Bacteriol. 137: 644–647 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167. McCafferty D. G., Lessard I. A. D., Walsh C. T. 1997. Mutational analysis of potential zinc-binding residues in the active site of the enterococcal d-Ala-d-Ala dipeptidase VanX. Biochemistry 36: 10498–10505 [DOI] [PubMed] [Google Scholar]
- 168. McQuillen K. 1958. Lysis resulting from metabolic disturbances. J. Gen. Microbiol. 18: 498–512 [DOI] [PubMed] [Google Scholar]
- 169. Meadow P., Hoare D. S., Work E. 1957. Interelationships between lysine and αε-diaminopimelic acid and their derivatives and analogues in mutants of Escherichia coli. Biochem. J. 66: 270–282 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170. Meberg B. M., Paulson A. L., Priyadarshini R., Young K. D. 2004. Endopeptidase penicillin-binding proteins 4 and 7 play auxiliary roles in determining uniform morphology of Escherichia coli. J. Bacteriol. 186: 8326–8336 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171. Meisel U., Höltje J.-V., Vollmer W. 2003. Overproduction of inactive variants of the murine synthase PBP1B causes lysis in Escherichia coli. J. Bacteriol. 185: 5342–5348 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172. Mengin-Lecreulx D., van Heijenoort J. 1985. Effect of growth conditions on peptidoglycan content and cytoplasmic steps of its biosynthesis in Escherichia coli. J. Bacteriol. 163: 208–212 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173. Mengin-Lecreulx D., van Heijenoort J. 1990. Correlation between the effects of fosfomycin and chloramphenicol on Escherichia coli. FEMS Microbiol. Lett. 66: 129–134 [DOI] [PubMed] [Google Scholar]
- 174. Mengin-Lecreulx D., Texier L., Rousseau M., van Heijenoort J. 1991. The murG gene of Escherichia coli codes for the UDP-N-acetylglucosamine:N-acetylmuramyl-(pentapeptide) pyrophosphoryl-undecaprenol N-acetylglucosamine transferase involved in the membrane steps of peptidoglycan synthesis. J. Bacteriol. 173: 4625–4636 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175. Mengin-Lecreulx D., van Heijenoort J. 1993. Identification of the glmU gene encoding N-acetylglucosamine-1-phosphate uridyltransferase in Escherichia coli. J. Bacteriol. 175: 6150–6157 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176. Mengin-Lecreulx D., van Heijenoort J., Park J. T. 1996. Identification of the mpl gene encoding UDP-N-acetylmuramate:l-alanyl-γ-d-glutamyl-meso-diaminopimelate ligase in Escherichia coli and its role in recycling of cell wall peptidoglycan. J. Bacteriol. 178: 5347–5352 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177. Mengin-Lecreulx D., van Heijenoort J. 1996. Characterization of the essential gene glmM encoding phosphoglucosamine mutase in Escherichia coli. J. Biol. Chem. 271: 32–39 [DOI] [PubMed] [Google Scholar]
- 178. Mengin-Lecreulx D., et al. 1999. Expression of the Staphylococcus aureus UDP-N-acetylmuramoyl-l-alanyl-d-glutamate:l-lysine ligase in Escherichia coli and effects on peptidoglycan biosynthesis and cell growth. J. Bacteriol. 181: 5909–5914 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 179. Metz R., Henning S., Hammes W. P. 1986. ld-Carboxypeptidase activity in Escherichia coli. I. The ld-carboxypeptidase activity in ether treated cells. Arch. Microbiol. 144: 175–180 [DOI] [PubMed] [Google Scholar]
- 180. Metz R., Henning S., Hammes W. P. 1986. ld-Carboxypeptidase activity in Escherichia coli. II. Isolation, purification and characterization of the enzyme from E. coli K 12. Arch. Microbiol. 144: 181–186 [DOI] [PubMed] [Google Scholar]
- 181. Mottl H., Terpstra P., Keck W. 1991. Penicillin-binding protein 4 of Escherichia coli shows a novel type of primary structure among penicillin-interacting proteins. FEMS Microbiol. Lett. 62: 213–220 [DOI] [PubMed] [Google Scholar]
- 182. Mottl H., Keck W. 1991. Purification of penicillin-binding protein 4 of Escherichia coli as a soluble protein by dye-affinity chromatography. Eur. J. Biochem. 200: 767–773 [DOI] [PubMed] [Google Scholar]
- 183. Nanninga N. 1991. Cell division and peptidoglycan assembly in Escherichia coli. Mol. Microbiol. 5: 791–795 [DOI] [PubMed] [Google Scholar]
- 184. Nathenson S. G., Strominger J. L. 1961. Effects of penicillin on the biosynthesis of the cell walls of Escherichia coli and Staphylococcus aureus. J. Pharmacol. Exp. Ther. 131: 1–6 [PubMed] [Google Scholar]
- 185. Nelson D. E., Young K. D. 2000. Penicillin binding protein 5 affects cell diameter, contour, and morphology of Escherichia coli. J. Bacteriol. 182: 1714–1721 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 186. Nelson D. E., Young K. D. 2001. Contributions of PBP 5 and DD-carboxypeptidase penicillin binding proteins to maintenance of cell shape in Escherichia coli. J. Bacteriol. 183: 3055–3064 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 187. Nelson D. E., Ghosh A. S., Paulson A. L., Young K. D. 2002. Contribution of membrane-binding and enzymatic domains of penicillin binding protein 5 to maintenance of uniform cellular morphology of Escherichia coli. J. Bacteriol. 184: 3630–3639 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 188. Nicholas R. A., Strominger J. L. 1988. Site-directed mutants of a soluble form of penicillin-binding protein 5 from Escherichia coli and their catalytic properties. J. Biol. Chem. 263: 2034–2040 [PubMed] [Google Scholar]
- 189. Nicholas R. A., Krings S., Tomberg J., Nicola G., Davies C. 2003. Crystal structure of wild-type penicillin-binding protein 5 from Escherichia coli. Implications for deacylation of the acyl-enzyme complex. J. Biol. Chem. 278: 52826–52833 [DOI] [PubMed] [Google Scholar]
- 190. Nikaido H., Vaara M. 1985. Molecular basis of bacterial outer membrane permeability. Microbiol. Rev. 49: 1–32 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 191. Nishimura Y., Suzuki H., Hirota Y., Park J. T. 1980. A mutant of Escherichia coli defective in penicillin-binding protein 5 and lacking d-alanine carboxypeptidase IA. J. Bacteriol. 143: 531–534 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192. Ornelas-Soares A., de Lencastre H., de Jonge B. L., Tomasz A. 1994. Reduced methicillin resistance in a new Staphylococcus aureus transposon mutant that incorporates muramyl dipeptides into the cell wall peptidoglycan. J. Biol. Chem. 269: 27246–27250 [PubMed] [Google Scholar]
- 193. Park J. T., Burman L. G. 1985. Elongation of the murein sacculus of Escherichia coli. Ann. Inst. Pasteur Microbiol. 136A: 51–58 [DOI] [PubMed] [Google Scholar]
- 194. Park J. T. 1996. The murein sacculus, p. 48–57 In Neidhardt F. C., et al. (ed.), Escherichia coli and Salmonella typhimurium: cellular and molecular biology. ASM Press, Washington, DC [Google Scholar]
- 195. Park J. T., Raychaudhuri D., Li H., Normark S., Mengin-Lecreulx D. 1998. MppA, a periplasmic binding protein essential for import of the bacterial cell wall peptide l-alanyl-γ-d-glutamyl-meso-diaminopimelate. J. Bacteriol. 180: 1215–1223 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 196. Park J. T., Uehara T. 2008. How bacteria consume their own exoskeletons (turnover and recycling of cell wall peptidoglycan). Microbiol. Mol. Biol. Rev. 72: 211–227 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 197. Parquet C., Flouret B., Leduc M., Hirota Y., van Heijenoort J. 1983. N-Acetylmuramoyl-L-alanine amidase of Escherichia coli K12. Possible physiological functions. Eur. J. Biochem. 133: 371–377 [DOI] [PubMed] [Google Scholar]
- 198. Pelzer H. 1963. Mucopeptidhydrolasen in Escherichia coli. I. Nachweis und Wirkungsspefizität. Z. Naturforsch. B 18: 950–956 [PubMed] [Google Scholar]
- 199. Pelzer H. 1963. Mucopeptidhydrolasen in Escherichia coli B. II. Quantitative Bestimungsverfahren und säulenchromatographische Trennung einiger Mucopeptid-hydrolasen. Z. Naturforsch. B 18: 956–964 [PubMed] [Google Scholar]
- 200. Pennartz A., Généreux C., Parquet C., Mengin-Lecreulx D., Joris B. 2009. Substrate-induced inactivation of the Escherichia coli AmiD N-acetylmuramoyl-L-alanine amidase highlights a new strategy to inhibit this class of enzyme. Antimicrob. Agents Chemother. 53: 2991–2997 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 201. Pisabarro A. G., de Pedro M. A., Vázquez D. 1985. Structural modifications in the peptidoglycan of Escherichia coli associated with changes in the state of growth of the culture. J. Bacteriol. 161: 238–242 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 202. Pisabarro A. G., Prats R., Vázquez D., Rodríguez-Tébar A. 1986. Activity of penicillin-binding protein 3 from Escherichia coli. J. Bacteriol. 168: 199–206 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 203. Pisabarro A. G., de Pedro M. A., Ishiguro E. E. 1990. Dissociation of the ampicillin-induced lysis of amino acid-deprived Escherichia coli into two stages. J. Bacteriol. 172: 2187–2190 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 204. Plumbridge J. 2009. An alternative route for recycling of N-acetylglucosamine from peptidoglycan involves the N-acetylglucosamine phosphotransferase system in Escherichia coli. J. Bacteriol. 191: 5641–5647 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 205. Potluri L., et al. 2010. Septal and lateral wall localization of PBP5, the major D,D-carboxypeptidase of Escherichia coli, requires substrate recognition and membrane attachment. Mol. Microbiol. 77: 300–323 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 206. Powell A. J., Liu Z.-J., Nicholas R. A., Davies C. 2006. Crystal structures of the lytic transglycosylase MltA from N. gonorrhoeae and E. coli: insights into interdomain movements and substrate binding. J. Mol. Biol. 359: 122–136 [DOI] [PubMed] [Google Scholar]
- 207. Prats R., de Pedro M. A. 1989. Normal growth and division of Escherichia coli with a reduced amount of murein. J. Bacteriol. 171: 3740–3745 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 208. Pratt J. M., Holland I. B., Spratt B. G. 1981. Precursor forms of pencillin-binding proteins 5 and 6 of E. coli cytoplasmic membrane. Nature 293: 307–309 [DOI] [PubMed] [Google Scholar]
- 209. Pratt J. M., Jackson M. E., Holland I. B. 1986. The C terminus of penicillin-binding protein 5 is essential for localisation to the E. coli inner membrane. EMBO J. 5: 2399–2405 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 210. Priyadarshini R., Popham D. L., Young K. D. 2006. Daughter cell separation by penicillin-binding proteins and peptidoglycan amidases in Escherichia coli. J. Bacteriol. 188: 5345–5355 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 211. Priyadarshini R., de Pedro M. A., Young K. D. 2007. Role of peptidoglycan amidases in the development and morphology of the division septum in Escherichia coli. J. Bacteriol. 189: 5334–5347 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 212. Pulvertaft R. J. V. 1951. The effect of antibiotics on growing cultures of bacterium coli. J. Pathol. Bacteriol. 64: 75–89 [DOI] [PubMed] [Google Scholar]
- 213. Reid C. W., Blackburn N. T., Legaree B. A., Auzanneau F.-I., Clarke A. J. 2004. Inhibition of membrane-bound lytic transglycosylase B by NAG-thiazoline. FEBS Lett. 574: 73–79 [DOI] [PubMed] [Google Scholar]
- 214. Reynolds P. E., Depardieu F., Dutka-Malen S., Arthur M., Courvalin P. 1994. Glycopeptide resistance mediated by enterococcal transposon Tn 1546 requires production of VanX for hydrolysis of d-alanyl-d-alanine. Mol. Microbiol. 13: 1065–1070 [DOI] [PubMed] [Google Scholar]
- 215. Rice K. C., Bayles K. W. 2008. Molecular control of bacterial death and lysis. Microbiol. Mol. Biol. Rev. 72: 85–109 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 216. Rodionov D. G., Pisabarro A. G., de Pedro M. A., Kusser W., Ishiguro E. E. 1995. β-Lactam-induced bacteriolysis of amino acid-deprived Escherichia coli is dependent on phospholipid synthesis. J. Bacteriol. 177: 992–997 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 217. Rodionov D. G., Ishiguro E. E. 1995. Direct correlation between overproduction of guanosine 3′,5′-bispyrophosphate (ppGpp) and penicillin tolerance in Escherichia coli. J. Bacteriol. 177: 4224–4229 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 218. Rodionov D. G., Ishiguro E. E. 1996. Effects of inhibitors of protein synthesis on lysis of Escherichia coli induced by β-lactam antibiotics. Antimicrob. Agents Chemother. 40: 899–903 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 219. Rodolakis A., Thomas P., Starka J. 1973. Morphological mutants of Escherichia coli. Isolation and ultrastructure of a chain-forming envC mutant. J. Gen. Microbiol. 75: 409–416 [DOI] [PubMed] [Google Scholar]
- 220. Rogers H. J., Perkins H. R., Ward J. B. 1980. Microbial cell walls and membranes. Chapman and Hall, London, United Kingdom [Google Scholar]
- 221. Romeis T., Vollmer W., Höltje J.-V. 1993. Characterization of three different lytic transglycosylases in Escherichia coli. FEMS Microbiol. Lett. 111: 141–146 [DOI] [PubMed] [Google Scholar]
- 222. Romeis T., Höltje J.-V. 1994. Penicillin-binding protein 7/8 of Escherichia coli is a DD-endopeptidase. Eur. J. Biochem. 224: 597–604 [DOI] [PubMed] [Google Scholar]
- 223. Romeis T., Höltje J.-V. 1994. Specific interaction of penicillin-binding proteins 3 and 7/8 with soluble lytic transglycosylase in Escherichia coli. J. Biol. Chem. 269: 21603–21607 [PubMed] [Google Scholar]
- 224. Ruiz N. 2008. Bioinformatics identification of MurJ (MviN) as the peptidoglycan lipid II flippase in Escherichia coli. Proc. Natl. Acad. Sci. U. S. A. 105: 15553–15557 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 225. Salmond G. P. C., Lutkenhaus J. F., Donachie W. D. 1980. Identification of new genes in a cell envelope-cell division gene cluster of Escherichia coli: cell envelope gene murG. J. Bacteriol. 144: 438–440 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 226. Santos J. M., Lobo M., Matos A. P. A., de Pedro M. A., Arraiano C. M. 2002. The gene bolA regulates dacA (PBP5), dacC (PBP6) and ampC (AmpC), promoting normal morphology in Escherichia coli. Mol. Microbiol. 45: 1729–1740 [DOI] [PubMed] [Google Scholar]
- 227. Sarvas M. 1971. Mutant of Escherichia coli K-12 defective in D-glucosamine biosynthesis. J. Bacteriol. 105: 467–471 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 228. Satta G., et al. 1995. Target for bacteriostatic and bactericidal activities of β-lactam antibiotics against Escherichia coli resides in different penicillin-binding proteins. Antimicrob. Agents Chemother. 39: 812–818 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 229. Sauvage E., Kerff F., Terrak M., Ayala J. A., Charlier P. 2008. The penicillin-binding proteins: structure and role in peptidoglycan biosynthesis. FEMS Microbiol. Rev. 32: 234–258 [DOI] [PubMed] [Google Scholar]
- 230. Scheurwater E., Reid C. W., Clarke A. J. 2008. Lytic transglycosylases: bacterial space-making autolysins. Int. J. Biochem. Cell Biol. 40: 586–591 [DOI] [PubMed] [Google Scholar]
- 231. Scheurwater E. M., Clarke A. J. 2008. The C-terminal domain of Escherichia coli YfhD functions as a lytic transglycosylase. J. Biol. Chem. 283: 8363–8373 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 232. Schleifer K. H., Kandler O. 1972. Peptidoglycan types of bacterial cell walls and their taxonomic implication. Bacteriol. Rev. 36: 407–477 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 233. Schmidt D. M. Z., Hubbard B. K., Gerlt J. A. 2001. Evolution of enzymatic activities in the enolase superfamily: functional assignment of unknown proteins in Bacillus subtilis and Escherichia coli as l-Ala-d/l-Glu epimerases. Biochemistry 40: 15707–15715 [DOI] [PubMed] [Google Scholar]
- 234. Schroeder U., Henrich B., Fink J., Plapp R. 1994. Peptidase D of Escherichia coli K-12, a metallopeptidase of low substrate specificity. FEMS Microbiol. Lett. 123: 153–159 [DOI] [PubMed] [Google Scholar]
- 235. Schwarz U., Weidel W. 1965. Zum Wirkungsmechanismus von Penicillin. I. Isolierung und Charakterisierung 2.6-Diaminopimelinsäure enthaltender, niedermolecularer Peptide aus Penicillinsphäroplasten von Escherichia coli. Z. Naturforsch. B 20: 147–153 [PubMed] [Google Scholar]
- 236. Schwarz U., Weidel W. 1965. Zum Wirkungsmechanismus von Penicillin. II. Nachweis eines Penicillin-induzierten hydrolytischen Abbaus von Murein und Mureinvorstufen in E. coli B. Z. Naturforsch. B 20: 153–157 [PubMed] [Google Scholar]
- 237. Schwarz U., Asmus A., Frank H. 1969. Autolytic enzymes and cell division of Escherichia coli. J. Mol. Biol. 41: 419–429 [DOI] [PubMed] [Google Scholar]
- 238. Shockman G. D., Höltje J.-V. 1994. Microbial peptidoglycan (murein) hydrolases, p. 131–166 In Ghuysen J.-M., Hakenbeck R. (ed.), New comprehensive biochemistry, vol. 27 Bacterial cell wall. Elsevier Science B.V., Amsterdam, Netherlands [Google Scholar]
- 239. Signoretto C., del Mar Lleò M., Canepari P. 2002. Modification of the peptidoglycan of Escherichia coli in the viable but nonculturable state. Curr. Microbiol. 44: 125–131 [DOI] [PubMed] [Google Scholar]
- 240. Smith T. J., Blackman S. A., Foster S. J. 2000. Autolysins of Bacillus subtilis: multiple enzymes with multiple functions. Microbiology 146: 249–262 [DOI] [PubMed] [Google Scholar]
- 241. Spratt B. G. 1975. Distinct penicillin-binding proteins involved in the division, elongation, and shape of Escherichia coli K12. Proc. Natl. Acad. Sci. U. S. A. 72: 2999–3003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 242. Spratt B. G., Strominger J. L. 1976. Identification of the major penicillin-binding proteins of Escherichia coli as d-alanine carboxypeptidase IA. J. Bacteriol. 127: 660–663 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 243. Spratt B. G. 1977. Properties of the penicillin-binding proteins of Escherichia coli K12. Eur. J. Biochem. 72: 341–352 [DOI] [PubMed] [Google Scholar]
- 244. Spratt B. G. 1980. Deletion of penicillin-binding protein 5 gene of Escherichia coli. J. Bacteriol. 144: 1190–1192 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 245. Spratt B. G. 1983. Penicillin-binding proteins and the future of β-lactam antibiotics. J. Gen. Microbiol. 129: 1247–1260 [DOI] [PubMed] [Google Scholar]
- 246. Stefanova M. E., Davies C., Nicholas R. A., Gutheil W. G. 2002. pH, inhibitor, and substrate specificity studies on Escherichia coli penicillin-binding protein 5. Biochim. Biophys. Acta 1597: 292–300 [DOI] [PubMed] [Google Scholar]
- 247. Stoker N. G., Broome-Smith J. K., Edelman A., Spratt B. G. 1983. Organization and subcloning of the dacA-rodA-pbpA cluster of cell shape genes in Escherichia coli. J. Bacteriol. 155: 847–853 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 248. Sturges W. S., Rettger L. F. 1922. Bacterial autolysis. J. Bacteriol. 7: 551–577 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 249. Suvorov M., Lee M., Hesek D., Boggess B., Mobashery S. 2008. Lytic transglycosylase MltB of Escherichia coli and its role in recycling of peptidoglycan strands of bacterial cell wall. J. Am. Chem. Soc. 130: 11878–11879 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 250. Suzuki H., Nishimura Y., Hirota Y. 1978. On the process of cellular division in Escherichia coli: a series of mutants of E. coli altered in the penicillin-binding proteins. Proc. Natl. Acad. Sci. U. S. A. 75: 664–668 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 251. Suzuki H., et al. 1980. In vitro peptidoglycan polymerization catalysed by penicillin binding protein 1b of Escherichia coli K-12. FEBS Lett. 110: 245–249 [DOI] [PubMed] [Google Scholar]
- 252. Tamura T., Imae Y., Strominger J. L. 1976. Purification to homogeneity and properties of two d-alanine carboxypeptidases I from Escherichia coli. J. Biol. Chem. 251: 414–423 [PubMed] [Google Scholar]
- 253. Taylor A., Das B. C., van Heijenoort J. 1974. Isolation and characterization of N-acetylglucosaminyl-β-1-4-1,6-anhydro-N-acetylmuramic acid. C. R. Acad. Sci. Hebd. Seances Acad. Sci. D 278: 1127–1129 [PubMed] [Google Scholar]
- 254. Taylor A., Das B. C., van Heijenoort J. 1975. Bacterial cell wall peptidoglycan fragments produced by phage λ or ViII endolysin and containing 1,6-anhydro-N-acetylmuramic acid. Eur. J. Biochem. 53: 47–54 [Google Scholar]
- 255. Templin M. F., Edwards D. H., Holtje J.-V. 1992. A murein hydrolase is the specific target of bulgecin in Escherichia coli. J. Biol. Chem. 267: 20039–20043 [PubMed] [Google Scholar]
- 256. Templin M. F., Ursinus A., Höltje J.-V. 1999. A defect in cell wall recycling triggers autolysis during the stationary growth phase of Escherichia coli. EMBO J. 18: 4108–4117 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 257. Thunnissen A.-M. W. H., et al. 1994. Doughnut-shaped structure of a bacterial muramidase revealed by X-ray crystallography. Nature 367: 750–753 [DOI] [PubMed] [Google Scholar]
- 258. Thunnissen A.-M. W. H., Isaacs N. W., Dijkstra B. W. 1995. The catalytic domain of a lytic bacterial transglycosylase defines a novel class of lysozymes. Proteins 22: 245–258 [DOI] [PubMed] [Google Scholar]
- 259. Thunnissen A.-M. W. H., Rozeboom H. J., Kalk K. H., Dijkstra B. W. 1995. Structure of the 70-kDa lytic transglycosylase complexed with bulgecin A. Implications for the enzymatic mechanism. Biochemistry 34: 12729–12737 [DOI] [PubMed] [Google Scholar]
- 260. Tomioka S., Matsuhashi M. 1978. Purification of penicillin-insensitive DD-endopeptidase, a new cell wall peptidoglycan-hydrolyzing enzyme in Escherichia coli, and its inhibition by deoxyribonucleic acids. Biochem. Biophys. Res. Commun. 84: 978–984 [DOI] [PubMed] [Google Scholar]
- 261. Tomioka S., Nikaido T., Miyakawa T., Matsuhashi M. 1983. Mutation of the N-acetylmuramyl-L-alanine amidase gene of Escherichia coli K-12. J. Bacteriol. 156: 463–465 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 262. Troup B., Jahn M., Hungerer C., Jahn D. 1994. Isolation of the hemF operon containing the gene for the Escherichia coli aerobic coproporphyrinogen III oxidase by in vivo complementation of a yeast HEM13 mutant. J. Bacteriol. 176: 673–680 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 263. Tsui H.-C. T., Zhao G., Feng G., Leung H.-C. E., Winkler M. E. 1994. The mutL repair gene of Escherichia coli K-12 forms a superoperon with a gene encoding a new cell-wall amidase. Mol. Microbiol. 11: 189–202 [DOI] [PubMed] [Google Scholar]
- 264. Tuomanen E., Tomasz A. 1986. Induction of autolysis in nongrowing Escherichia coli. J. Bacteriol. 167: 1077–1080 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 265. Tuomanen E. 1986. Phenotypic tolerance: the search for β-lactam antibiotics that kill nongrowing bacteria. Rev. Infect. Dis. 8: S279–S291 [DOI] [PubMed] [Google Scholar]
- 266. Tuomanen E., Schwartz J. 1987. Penicillin-binding protein 7 and its relationship to lysis of nongrowing Escherichia coli. J. Bacteriol. 169: 4912–4915 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 267. Tuomanen E., Cozens R. 1987. Changes in peptidoglycan composition and penicillin-binding proteins in slowly growing Escherichia coli. J. Bacteriol. 169: 5308–5310 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 268. Tuomanen E., Markiewicz Z., Tomasz A. 1988. Autolysis-resistant peptidoglycan of anomalous composition in amino-acid-starved Escherichia coli. J. Bacteriol. 170: 1373–1376 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 269. Uehara T., Park J. T. 2003. Identification of MpaA, an amidase in Escherichia coli that hydrolyzes the γ-d-glutamyl-meso-diaminopimelate bond in murein peptides. J. Bacteriol. 185: 679–682 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 270. Uehara T., Park J. T. 2007. An anhydro-N-acetylmuramyl-L-alanine amidase with broad specificity tethered to the outer membrane of Escherichia coli. J. Bacteriol. 189: 5634–5641 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 271. Uehara T., Park J. T. 2008. Growth of Escherichia coli: significance of peptidoglycan degradation during elongation and septation. J. Bacteriol. 190: 3914–3922 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 272. Uehara T., Dinh T., Bernhardt T. G. 2009. LytM-domain factors are required for daughter cell separation and rapid ampicillin-induced lysis in Escherichia coli. J. Bacteriol. 191: 5094–5107 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 273. Uehara T., Parzych K. R., Dinh T., Bernhardt T. G. 2010. Daughter cell separation is controlled by cytokinetic ring-activated cell wall hydrolysis. EMBO J. 29: 1412–1422 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 274. Ursinus A., Steinhaus H., Höltje J.-V. 1992. Purification of a nocardicin A-sensitive LD-carboxypeptidase from Escherichia coli by affinity chromatography. J. Bacteriol. 174: 441–446 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 275. Ursinus A., Höltje J.-V. 1994. Purification and properties of a membrane-bound lytic transglycosylase from Escherichia coli. J. Bacteriol. 176: 338–343 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 276. van Asselt E. J., Thunnissen A.-M. W. H., Dijkstra B. W. 1999. High resolution crystal structures of the Escherichia coli lytic transglycosylase Slt70 and its complex with a peptidoglycan fragment. J. Mol. Biol. 291: 877–898 [DOI] [PubMed] [Google Scholar]
- 277. van Asselt E. J., et al. 1999. Crystal structure of Escherichia coli lytic transglycosylase Slt35 reveals a lysozyme-like catalytic domain with an E-F hand. Structure 7: 1167–1180 [DOI] [PubMed] [Google Scholar]
- 278. van Asselt E. J., Kalk K. H., Dijkstra B. W. 2000. Crystallographic studies of the interactions of Escherichia coli lytic transglycosylase Slt35 with peptidoglycan. Biochemistry 39: 1924–1934 [DOI] [PubMed] [Google Scholar]
- 279. van der Linden M. P. G., de Haan L., Hoyer M. A., Keck W. 1992. Possible role of Escherichia coli penicillin-binding protein 6 in stabilization of stationary-phase peptidoglycan. J. Bacteriol. 174: 7572–7578 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 280. van der Linden M. P. G., de Haan L., Keck W. 1993. Domain organization of penicillin-binding protein 5 from Escherichia coli analyzed by C-terminal truncation. Biochem. J. 289: 593–598 [PMC free article] [PubMed] [Google Scholar]
- 281. van der Linden M. P. G., de Haan L., Dideberg O., Keck W. 1994. Site-directed mutagenesis of proposed active-site residues of penicillin-binding protein 5 from Escherichia coli. Biochem. J. 303: 357–362 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 282. van Heijenoort Y., van Heijenoort J. 1971. Study of the N-acetylmuramyl-L-alanine amidase activity in Escherichia coli. FEBS Lett. 15: 137–141 [DOI] [PubMed] [Google Scholar]
- 283. van Heijenoort J., Parquet C., Flouret B., van Heijenoort Y. 1975. Envelope-bound N-acetylmuramyl-L-alanine amidase of Escherichia coli K 12. Purification and properties of the enzyme. Eur. J. Biochem. 58: 611–619 [DOI] [PubMed] [Google Scholar]
- 284. van Heijenoort Y., Leduc M., Singer H., van Heijenoort J. 1987. Effect of moenomycin on Escherichia coli. J. Gen. Microbiol. 133: 667–674 [DOI] [PubMed] [Google Scholar]
- 285. van Heijenoort Y., Gomez M., Derrien M., Ayala J., van Heijenoort J. 1992. Membrane intermediates in the peptidoglycan metabolism of Escherichia coli: possible roles of PBP 1b and PBP 3. J. Bacteriol. 174: 3549–3557 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 286. van Heijenoort J. 2001. Recent advances in the formation of the bacterial peptidoglycan monomer unit. Nat. Prod. Rep. 18: 503–519 [DOI] [PubMed] [Google Scholar]
- 287. van Heijenoort J. 2001. Formation of the glycan chains in the synthesis of bacterial peptidoglycan. Glycobiology 11: 25R–36R [DOI] [PubMed] [Google Scholar]
- 288. van Heijenoort J. 2007. Lipid intermediates in the biosynthesis of bacterial peptidoglycan. Microbiol. Mol. Biol. Rev. 71: 620–635 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 289. van Straaten K. E., Dijkstra B. W., Vollmer W., Thunnissen A.-M. W. H. 2005. Crystal structure of MltA from Escherichia coli reveals a unique lytic transglycosylase fold. J. Mol. Biol. 352: 1068–1080 [DOI] [PubMed] [Google Scholar]
- 290. van Straaten K. E., Barends T. R. M., Dijkstra B. W., Thunnissen A.-M. W. H. 2007. Structure of Escherichia coli lytic transglycosylase MltA with bound chitohexaose. Implications for peptidoglycan binding and cleavage. J. Biol. Chem. 282: 21197–21205 [DOI] [PubMed] [Google Scholar]
- 291. Vega D., Ayala J. A. 2006. The DD-carboxypeptidase activity encoded by pbp4B is not essential for the cell growth of Escherichia coli. Arch. Microbiol. 185: 23–27 [DOI] [PubMed] [Google Scholar]
- 292. Venkateswaran P. S., Wu H. C. 1972. Isolation and characterization of a phosphonomycin-resistant mutant of Escherichia coli K-12. J. Bacteriol. 110: 935–944 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 293. Vollmer W., von Rechenberg M., Höltje J.-V. 1999. Demonstration of molecular interactions between the murein polymerase PBP1B, the lytic transglycosylase MltA, and the scaffolding protein MipA of Escherichia coli. J. Biol. Chem. 274: 6726–6734 [DOI] [PubMed] [Google Scholar]
- 294. Vollmer W., Blanot D., de Pedro M. A. 2008. Peptidoglycan structure and architecture. FEMS Microbiol. Rev. 32: 1–19 [DOI] [PubMed] [Google Scholar]
- 295. Vollmer W., Joris B., Charlier P., Foster S. 2008. Bacterial peptidoglycan (murein) hydrolases. FEMS Microbiol. Rev. 32: 259–286 [DOI] [PubMed] [Google Scholar]
- 296. Vollmer W. 2008. Structural variation in the glycan strands of bacterial peptidoglycan. FEMS Microbiol. Rev. 32: 287–306 [DOI] [PubMed] [Google Scholar]
- 297. von Rechenberg M., Ursinus A., Höltje J.-V. 1996. Affinity chromatography as a means to study multienzyme complexes involved in murein synthesis. Microb. Drug Resist. 2: 155–157 [DOI] [PubMed] [Google Scholar]
- 298. Vötsch W., Templin M. F. 2000. Characterization of the β-N-acetylglucosaminidase of Escherichia coli and elucidation of its role in muropeptide recycling and β-lactamase induction. J. Biol. Chem. 275: 39032–39038 [DOI] [PubMed] [Google Scholar]
- 299. Walderich B., Höltje J.-V. 1991. Subcellular distribution of the soluble lytic transglycosylase in Escherichia coli. J. Bacteriol. 173: 5668–5676 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 300. Wang T.-S. A., Manning S. A., Walker S., Kahne D. 2008. Isolated peptidoglycan glycosyltransferases from different organisms produce different glycan chain lengths. J. Am. Chem. Soc. 130: 14068–14069 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 301. Waxman D. J., Strominger J. L. 1983. Penicillin-binding proteins and the mechanism of action of β-lactam antibiotics. Annu. Rev. Biochem. 52: 825–869 [DOI] [PubMed] [Google Scholar]
- 302. Weatherspoon-Griffin N., et al. 2011. The CpxR/CpxA two-component system up-regulates two Tat-dependent peptidoglycan amidases to confer bacterial resistance to antimicrobial peptide. J. Biol. Chem. 286: 5529–5539 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 303. Weidel W., Frank H., Leutgeb W. 1963. Autolytic enzymes as a source of error in the preparation and study of gram-negative cell walls. J. Gen. Microbiol. 30: 127–130 [DOI] [PubMed] [Google Scholar]
- 304. Weidel W., Pelzer H. 1964. Bagshaped macromolecules: a new outlook on bacterial cell walls. Adv. Enzymol. 26: 193–232 [DOI] [PubMed] [Google Scholar]
- 305. Welzel P. 2005. Syntheses around the transglycosylation step in peptidoglycan biosynthesis. Chem. Rev. 105: 4610–4660 [DOI] [PubMed] [Google Scholar]
- 306. White R. J. 1968. Control of amino sugar metabolism in Escherichia coli and isolation of mutants unable to degrade amino sugars. Biochem. J. 106: 847–858 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 307. Wijsman H. J. W. 1972. The characterization of an alanine racemase mutant of Escherichia coli. Genet. Res. (Camb.) 20: 269–277 [DOI] [PubMed] [Google Scholar]
- 308. Wolf-Watz H., Normark S. 1976. Evidence for a role of N-acetylmuramyl-l-alanine amidase in septum separation in Escherichia coli. J. Bacteriol. 128: 580–586 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 309. Wu H. C., Wu T. C. 1971. Isolation and characterization of a glucosamine-requiring mutant of Escherichia coli K-12 defective in glucosamine-6-phosphate synthetase. J. Bacteriol. 105: 455–466 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 310. Yem D. W., Wu H. C. 1976. Purification and propeties of a β-N-acetylglucosaminidase from Escherichia coli. J. Bacteriol. 125: 324–331 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 311. Yem D. W., Wu H. C. 1976. Isolation of Escherichia coli K-12 mutants with altered levels of β-N-acetylglucosaminidase. J. Bacteriol. 125: 372–373 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 312. Zapun A., Contreras-Martel C., Vernet T. 2008. Penicillin-binding proteins and β-lactam resistance. FEMS Microbiol. Rev. 32: 361–385 [DOI] [PubMed] [Google Scholar]
- 313. Zapun A., Vernet T., Pinho M. G. 2008. The different shapes of cocci. FEMS Microbiol. Rev. 32: 345–360 [DOI] [PubMed] [Google Scholar]
- 314. Zhang X., El-Hajj Z. W., Newman E. 2010. Deficiency in L-serine deaminase interferes with one-carbon metabolism and cell wall synthesis in Escherichia coli K-12. J. Bacteriol. 192: 5515–5525 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 315. Zhao G., Meier T. I., Kahl S. D., Gee K. R., Blaszczak L. C. 1999. Bocillin FL, a sensitive and commercially available reagent for detection of penicillin-binding proteins. Antimicrob. Agents Chemother. 43: 1124–1128 [DOI] [PMC free article] [PubMed] [Google Scholar]