

Abstract
Individuals face evolutionary trade-offs between the acquisition of costly but accurate information gained firsthand and the use of inexpensive but possibly less reliable social information. American crows (Corvus brachyrhynchos) use both sources of information to learn the facial features of a dangerous person. We exposed wild crows to a novel ‘dangerous face’ by wearing a unique mask as we trapped, banded and released 7–15 birds at five study sites near Seattle, WA, USA. An immediate scolding response to the dangerous mask after trapping by previously captured crows demonstrates individual learning, while an immediate response by crows that were not captured probably represents conditioning to the trapping scene by the mob of birds that assembled during the capture. Later recognition of dangerous masks by lone crows that were never captured is consistent with horizontal social learning. Independent scolding by young crows, whose parents had conditioned them to scold the dangerous mask, demonstrates vertical social learning. Crows that directly experienced trapping later discriminated among dangerous and neutral masks more precisely than did crows that learned through social means. Learning enabled scolding to double in frequency and spread at least 1.2 km from the place of origin over a 5 year period at one site.
Keywords: American crow, Corvus brachyrhynchos, fear learning, mobbing, public information, social learning
1. Introduction
An animal's world includes environmental challenges that select for adaptive traits across generations [1], as well as those that change within individual lifetimes and necessitate adaptive learning [2]. Learning from direct trial and error experiences allows for a wide range of species to rapidly adjust their behaviour to changing conditions [3–5], but errors can be costly [6,7]. Species able to observe and imitate others [8], even crudely [9], can forgo this cost by adjusting to changing environments through social learning and cultural evolution [10,11].
In our increasingly human-dominated world, animals frequently face new dangers and opportunities that would profitably be solved through social learning. Nowhere may this be more apparent than in our cities, where changing abiotic conditions; novel anthropogenic foods and other subsidies; and exotic predators, diseases and competitors are potent, novel and erratic selective forces [12,13]. The behaviour of individual people towards animals is often changing, and recent studies have demonstrated that an ability to discern differences between humans [14,15] may enable successful species to adapt to, and even coevolve with, human behaviour [16]. Because human actions often threaten animals, learning socially about individual people's habits would be advantageous.
Learning dangers by observing others is well understood in the laboratory [17], but difficult to demonstrate in nature because public information can influence an animal's behaviour without the need for imitative social learning [18,19]. Society can influence an individual's behaviour by stimulating (contagion), exposing or supporting an action that already exists in the animal's repertoire. Such socially influenced traits are subject to cultural transmission, but at best provide weak evidence for social learning as new behaviours need not be acquired [20,21]. By contrast, social animals often model adaptive traits that enable individuals to learn the object, location or circumstances that enhance fitness. Such social learning (i.e. stimulus enhancement or observational conditioning; [19]) is more rudimentary than imitating complex behaviours or learning about goals from others, but it is common among socially tolerant animals [22,23].
The American crow (Corvus brachyrhynchos) is a successful resident of North American cities [24] with a typical fission–fusion social structure within which many traditions and vocalizations appear to be socially learned [16,25]. One such tradition is the tendency of some crow groups to habitually follow or scorn particular humans. In experiments that we expand upon here, crows were capable of discriminating among individual people and remembering those who have aided or threatened them in the past [26]. Threatening people are scolded with harsh vocalizations and may be mobbed by groups of scolding crows. Individual crows learn from a single experience when they are captured and scold their captors, but in our experiments many more crows appeared knowledgeable than the few with direct, individual experience. Crows recruit and tolerate others of their own and different species in mobs that form around dangerous people. This social tolerance during a situation when a typically neutral stimulus (a person) is paired with a signal of danger (scolding) could allow naive crows to learn about dangerous situations, locations and individual humans. Alternatively, many naive crows may simply be stimulated to mob dangerous people because of the contagious nature of a mob [27].
Mobbing is a compound social stimulus that includes both visual and auditory signals in response to predators that is found in a variety of taxa including fishes, birds and mammals [28]. Mobbing has multiple functions including predation, deterrence and learning to identify predators [29–32]. Here, we are concerned with the latter and seek to distinguish contagious from socially acquired behaviour to understand the relative roles of individual learning and social learning in the acquisition of knowledge about people by crows. We test the hypothesis that observing others mob an object conditions an individual to react to that object and thus subsequently scold it [29]. Our experiments in a field setting advance the relatively poor understanding of social learning pathways in nature [22].
2. Methods
(a). Field procedures
We experimentally investigated the roles of individual and social learning by pairing an aversive experience (capture, banding and release) with a novel human face (a full face mask worn by people trapping and handling the birds). After trapping, we investigated the strength of the ecologically relevant, unconditioned response (fear, scolding or mobbing) to the conditioned stimulus (people wearing the mask used during trapping, ‘dangerous face’) and to similar stimuli (people wearing other masks not used for trapping or no mask, ‘neutral faces’; see the electronic supplementary material, also see Marzluff et al. [26] for details on masks, previous research at sites and discriminatory abilities of subjects).
We trapped and then observed crows respond to masks at five sites. We began research at the University of Washington (UW) main campus, where we contrasted crow responses over a 5 year period (and ongoing) to an extraordinary dangerous mask. To test the generality of our findings at UW, and test discrimination among ordinary faces in a fully balanced design, where each mask was dangerous at one site and neutral at all other sites, we conducted five-month long observations at four additional sites (see the electronic supplementary material).
While our goals varied among sites, our field procedures were standardized. Two trappers each wore the same mask during capture with a net launcher and banding. In total, birds were held for 10–30 min by the masked trappers, during which time groups of crows who were not captured circled and gave alarm vocalizations (the electronic supplementary material). After releasing the banded birds, we removed our masks while inside a nearby building. Each bird was aged based on plumage and colour of the mouth lining ([33]; electronic supplementary material). Adults were only marked using leg bands (including colour bands that did not reinforce coloration known to be associated with status) to minimize long-term stress on the crows. All trapping was carried out at the start of experiments; once trapping was completed, no additional individual learning by experiencing or observing trapping was possible.
To assess the responses of the crow populations at each site to people with masks after capture, we surveyed crows and their responses to a person slowly walking a 2–3.8 km-long route for 1–2 h (figures 1 and 3; electronic supplementary material). Surveys were carried out by the authors and others who were blind to the experimental objectives and expectations. Observations made by blind observers are conservatively biased (typically ascribing less discriminatory ability to crows because some scold vocalizations were not recognized as such; [26]). Each route included the trapping sites. At each site, the order of trials in which observers wore the dangerous trapping mask and the neutral masks was randomized. During surveys, which were limited to no more than one per day, observers paused briefly to look at each crow encountered and categorized its behaviour. We defined scolding as a harsh, alarm ‘kaw’ directed repeatedly at the observer accompanied by agitated wing and tail flicking [34]. Mobbing (or a mob) is defined as contagious activity where more than one crow jointly scolds. Mobbing may be accompanied by diving and following the masked observer. Observers did not carry binoculars (so as not to cue birds), but noted the unique band combination of each crow when possible, enabling us to determine if a particular individual was previously captured. Although attraction of birds along the route to mobs may increase the chance of repeatedly counting the same birds within an experiment, we have no suggestion that it occurred. Our routes were long, many of the crows were territorial and wandered little beyond their centres of activity, and we never observed a banded bird in more than a single mob within a trial.
Figure 1.

Responses of lone, unbanded crows (solid symbols, scolding; open symbols, silent) at site B (urban and dense suburban Bellevue, WA; for other sites see the electronic supplementary material). Each of the 11 repeated trials in which the dangerous mask was worn is represented as a straight line with segments proportional to the actual route (lines 1–11 under the study route, letters orient lines to route; dashed lines delineate the major segments). The locations of all mobs given in response to the dangerous person's presence and the responses of all lone unbanded (never captured) crows are plotted in a spatially explicit manner for each trial. Scolding crows exposed to a mob (observed within 100 m of the location where mobbing occurred on a previous trial; these crows have a plus symbol) are evidence of peer-to-peer social learning. Scolding crows that were not exposed to a mob are evidence of social learning by observing the trapping event (trial 1), evidence of inherent scolding of a masked person (trials 2–11) or the conservative nature of our assessment (trials 2–11).
Figure 3.

Spatial increase in intensity and extent of scolding at the UW site. Locations of consistent (cross hatched circles) and inconsistent (filled circles) scolding to the dangerous mask from (a) two weeks, (b) 1.25 years and (c) 2.8 years post-trapping. Circle diameter is proportional to the number of scolding crows: smallest circles represent single crows; largest represent groups of seven crows. Solid line marks survey route. (d) Responses with scolding (filled circles) and no scolding (open circles) during the expanded search (large dashed circle superimposed on street network) carried out 2.7 years after trapping.
(b). Temporal and spatial effect of social learning at the University of Washington
Ongoing observations at our single long-term research site (UW) enabled us to describe the potential effect of social learning by detailing the nature of involvement in scolding by birds that were not trapped. We did this on a temporal basis by comparing the change in the number of birds scolding after trapping with two typical models of population-level cultural learning: linear and exponential rise to maximum [35,36]. Network-based approaches [37] were not possible because many birds were not individually marked. Rejection of the linear model would be consistent with lack of social learning. Support for the linear model would be consistent with ongoing social learning. Support for the exponential rise to maximum model would be consistent with learning by observation of the trapping event, but little or no further learning by observing scolding.
We quantified the geographical spread in scolding by mapping the locations and average number of scolding birds during the first two weeks immediately after trapping, 1.25 years after trapping, and 2.7 years post-trapping (9, 31 and 38 cumulative number of times crows were exposed to the dangerous mask prior to each time point, respectively). Since the first 2 years of data suggested that the response area was increasing, we expanded our search of knowledgeable birds off of the typical route at the 2.7 year mark. We surveyed in a circular search area with a radius of 1160 m, positioned around our typical route. We started at the perimeter of the circle and worked our way in towards the centre. We drove until we saw a crow perched within 10 m of the ground, put on the mask, approached the crow on foot to observe its response, then removed the mask once back inside the vehicle. We surveyed this area on a weekly basis from 9 October 2008 until 30 November 2008.
(c). Distinguishing contagion from horizontal social learning at new sites with a short-term experiment
To differentiate horizontal social learning from contagious behaviour, we investigated the temporal change in scolding by lone crows that had never been trapped. The responses of lone unbanded crows during the first trial after trapping at each of the four non-UW sites indicate the degree to which crows may have learned about the dangerous mask by observing us during the previous week's capture events. The responses of lone unbanded crows in 10 subsequent trials could indicate the extent to which they learned about the dangerous mask via observation of trapping or horizontal transmission of social information within mobs. Because these birds were not marked, we cannot tell whether observing trapping or horizontal learning is responsible for changes in the behaviour of lone crows. But, we believe the latter is most probable, especially in the second half of the experiment, as we have no reason to suspect some crows who observed trapping then left the study area for a time or remained silent when initially, but not subsequently, encountering the dangerous face. Limited observations at UW, where some crows were banded prior to the start of our mask experiments, support this belief. Lone crows that were present but not trapped at UW remained consistent in their behaviour during subsequent experimental trials (six consistently scolded, three consistently did not and one scolded after exposure to a mob).
Because we could not individually identify lone, unbanded crows, we used the less precise approach of analysing the behaviour of crows at discrete locations where they were encountered along the route. We did this only at sites A, B, C and D because the short duration of experiments (five months), the defence of small exclusive territories by adult crows (territories typically include no more than 200 m of the survey route [24], while adjacent lone crows were 100–1000 m apart; figure 1 and the electronic supplementary material), and the fact that scolding crows never followed us for more than 100 m before they returned to the place we originally encountered them makes it likely that some of the unbanded crows we repeatedly encountered within 100 m of a location were the same individuals. We assumed this to be the case and mapped the locations of lone unbanded crows and mobs encountered during trials with the dangerous masks at each of the four sites. We classified each location as either being exposed (within 100 m) to a mob or not, and as having a lone unbanded crow that scolded or did not during each of the 11 trials described above (see figure 1 and the electronic supplementary material for layout of routes, trapping locations, occurrence of mobs and responses of lone crows).
(d). Distinguishing contagion from vertical social learning at short- and long-term study sites
To determine the occurrence of vertical transmission of social information, we exposed adult pairs to the dangerous mask and observed their responses and those of their dependent young to that mask and to a paired control trial when no mask was worn. Development of a response to the dangerous mask that was eventually given by young birds without the contagious stimulation of their parents is consistent with vertical transmission of information. Discrimination by young birds between a person wearing the dangerous mask and a person not wearing a mask indicates learning about a new danger from the actions of their parents. We did not test the ability of young birds to discriminate among dangerous and neutral masks because previously we demonstrated that crows respond similarly to neutral people whether they are masked or not and readily distinguish both types of experimental controls from people wearing dangerous masks [26]. All exposures were begun early in the fledgling period when young crows typically communicate with parents via begging vocalizations and do not yet utter scolding calls. The response of young crows was monitored later in the nestling period when they were beginning to disperse from the natal territory.
We monitored 24 young from 10 different pairs of crows at the UW site in 2007. The 10 adult pairs, who all reliably scolded the dangerous mask were identified by either having unique colour-bands from a previous banding event or by nest location. To reduce direct learning by the young, we did not capture and band them, thus we could only follow them until they regularly left the natal territory. Because we could not reliably distinguish among multiple unmarked young on each territory, we considered them as 10 families not 24 individuals in our analyses (see the electronic supplementary material). In 2008, we included banded young from the other four sites. At three to five weeks of age, within a week of fledging, we captured by hand, banded and radio tagged the young of four adult pairs of crows in which at least one bird had previously been captured by the dangerous mask and both members of the pair consistently scolded the mask. We reinforced the association between the mask and danger that these adult crows had previously learned by approaching the nest and climbing the nest tree while wearing the dangerous mask. We did not directly expose the young to the mask; while just underneath the nest we took off the mask and quickly put the young into a pouch preventing their view. These capture attempts took 20 min per bird in which we put on a unique colour-band combination and attached 11 g radio transmitters (total weight less than 3% of body weight) by a backpack harness [38]. During tagging, we took 300 ml blood samples from the ulnar vein of juvenile birds to determine sex.
To initiate learning, we presented the dangerous mask (and at UW the neutral unmasked face) to both banded and unbanded juvenile crows while they were still on the natal territory, letting them observe their parents scold us wearing the mask. We visited each of the 14 territories 10–24 times for 5–20 min while wearing the mask to allow the parents to scold while their offspring had clear view of us. After this period, we tested the reaction of fledglings to the dangerous mask and neutral face. In 2007 at UW, we tested families 25–35 days after fledging only on the natal territory when parents were and were not present. In 2008 at the other four sites, we tested radio-tagged young while on the natal territory with their parents, and after dispersal from the territory without their parents. We exposed families to either the dangerous or neutral face no more than once per day in a random order. Because mortality is frequent on fledglings, we were unable to observe some families respond to both stimuli when the parents were not present (11 families were exposed to the dangerous face, nine to the neutral and eight to both). During each presentation of the mask, we categorized behaviour as mobbing, scolding or neither for adults and either begging, scolding or neither for the young.
(e). Analyses
All general statistical tests were performed using SPSS for Windows v. 12.0 [39]. Curve-fitting was done with SigmaPlot v. 11 [40]. Exact tests were conducted using StatXact v. 6 [41]. We used ordinary least-squares regression to quantify the degree and pattern of temporal autocorrelation among repeated trials within a site to the same type of mask. We assumed the responses of individual crows to various masks were independent when we could not identify specific individuals, and where repeated encounters were spread over weeks to years and were directed at many different observers over a range of environmental conditions. Where possible (repeated tests on known crow families; balanced and repeated testing of dangerous and neutral masks at sites A, B, C and D) we employed repeated-measures analyses. In one such analysis, the responses of birds to dangerous versus neutral masks during the first and second half of trials were square root-arcsine-transformed and analysed as repeated measures scored by each of n = 10 blind observers at four sites.
3. Results
(a). Observations consistent with social learning over a 5 year period
We previously demonstrated an immediate increase in scolding the dangerous mask relative to neutral masks at our long-term study site (UW) after trapping [26]. The number of crows scolding the dangerous mask continued to increase for 5 years after trapping, as expected if social learning or social stimulation were present (adjusted r2 = 0.17; F1,37 = 8.75, p = 0.005; figure 2a). As we conducted trials at UW, walking with the dangerous mask along the route, our actions presented opportunities for crows to observe or be stimulated by scolding. The number of crows encountered was consistent across trials (n = 38, r = −0.11, p = 0.51), but the number that scolded the dangerous mask increased steadily (figure 2b). The temporal increase in the number of crows scolding the dangerous mask was consistent with the linear model (Y = 22.3 + 1.0X; adjusted r2 = 0.27; F1,37 = 14.49, p = 0.0005), and not with the exponential rise to maximum model (Y = 15.0 + 35 (X/0.75X); adjusted r2 = 0.00; F2,37 = 0.95, p = 0.40). No change in the rate of scolding over time to the neutral mask was evident (poor fit to the data of the linear model: Y = 12.8–0.03X; adjusted r2 = 0.00; p = 0.93).
Figure 2.

Scolding of the (a,b) dangerous and (c) neutral mask at our long-term research site at the University of Washington (UW) campus. The same responses to the dangerous face are plotted as a function of real time (a) and the number of trials during which birds could witness others scolding (exposures to dangerous mask; b). Responses were obtained during the breeding and non-breeding seasons by the authors and naive observers blind to the study design. A linear model was fit to all the data (solid lines with confidence intervals; a,c). An exponential rise to maximum model was also fit to the responses to the dangerous mask (dashed line; b).
Scolding the dangerous mask at UW has gradually involved more crows, spread over a larger area (figure 3). In the first two weeks after trapping, an average of 26 per cent of crows encountered (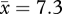 individuals) scolded the dangerous mask consistently at one location covering 3060 m2 and intermittently at seven locations (9000 m2). By 1.25 years post-trapping, 30.4 per cent of crows encountered (
individuals) scolded the dangerous mask consistently at one location covering 3060 m2 and intermittently at seven locations (9000 m2). By 1.25 years post-trapping, 30.4 per cent of crows encountered (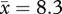 individuals) scolded the dangerous mask consistently at two locations (6000 m2) and intermittently at 15 (16 900 m2). In autumn, 2008, 2.7 years after trapping ceased, 66 per cent of crows encountered (
individuals) scolded the dangerous mask consistently at two locations (6000 m2) and intermittently at 15 (16 900 m2). In autumn, 2008, 2.7 years after trapping ceased, 66 per cent of crows encountered (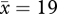 individuals) scolded the dangerous mask consistently at seven locations (10 922 m2) and intermittently at six locations (8050 m2). At this time, we expanded surveys beyond the original route and four out of 65 individuals encountered scolded the dangerous mask at three new locations 541, 667 and 1248 m away from the trapping locations. Considering that only 3.0 per cent of 167 crows scolded the dangerous mask on campus prior to trapping in 2005 [26], we expected only 3 per cent of naive crows to respond to the dangerous mask in 2008 (two of 65 encountered). The frequency of scolding at UW has continued to increase beyond this time (figure 2a), but we have not reassessed the spatial extent of scolding.
individuals) scolded the dangerous mask consistently at seven locations (10 922 m2) and intermittently at six locations (8050 m2). At this time, we expanded surveys beyond the original route and four out of 65 individuals encountered scolded the dangerous mask at three new locations 541, 667 and 1248 m away from the trapping locations. Considering that only 3.0 per cent of 167 crows scolded the dangerous mask on campus prior to trapping in 2005 [26], we expected only 3 per cent of naive crows to respond to the dangerous mask in 2008 (two of 65 encountered). The frequency of scolding at UW has continued to increase beyond this time (figure 2a), but we have not reassessed the spatial extent of scolding.
(b). Public information available to crows at all sites
We documented three distinct sources of public information revealing the identity of dangerous people. Each of these sources could have produced the increased frequency and extent of scolding that we documented over a 5 year period at the UW site.
— At the time of capture, a number of crows, whose identities were not known, witnessed people trapping and wearing the dangerous mask. After birds were trapped, masked people were exposed for 15–45 min marking, measuring and releasing birds. At each site, except D, a mob of 15–40 crows circled above each trap site at a height of 10–30 m in response to capture, and scolded for up to 10 min before dispersing (see the electronic supplementary material). At site D, where crows were occasionally killed by homeowners, such a group (five crows) only assembled in response to one of four trapping events.
— After trapping, during our experiments, crows that did not have a direct, negative experience with the mask associated with other crows that were captured by a person wearing the dangerous mask. Scolding vocalizations directed at the dangerous mask typically (82% of 126 scolding events) attracted a mob of crows who scolded the masked person. Mob size averaged 6.4 crows (range 2–21, n =103) and included both crows previously captured and those not previously captured by masked people. Most of these birds were not trapped by us while we wore the dangerous mask; 86.9 per cent of the 160 scolding birds we identified as either not banded or trapped and banded at least 2 years earlier by unmasked people. While scolding, crows maintained a consistent minimum distance of 17.4 m (n = 126, s.e.m. = 1.20) between themselves and the masked person while the mob perched around and moved with, but rarely dove at, the person. Mobs escorted the masked person for distances of 10–100 m before returning to their previous activities.
— Young birds hatched after our trapping was concluded, observed and joined their parents scolding the dangerous mask. We observed fledglings scold the dangerous mask in the company of their parents at all territories where parents had previously been conditioned (n = 10 at UW in 2007 and n = 2 at other sites in 2008). By contrast, fledglings and parents rarely (n = 2 of 9 territories) scolded neutral observers (n = 8 territories at UW in 2007, and n = 1 territory at other sites in 2008).
(c). Use of public information for stimulus enhancement during the short-term study
Scolding the dangerous mask by lone, unbanded individuals in the first trial after capture is consistent with learning about the trapping site and the trapper from observation or participation in the mob that formed in response to the capture event (i.e. stimulus enhancement). We only have evidence consistent with this mode of learning for one lone crow (figure 1). This bird was near the trap site on our first trial and a mob had also formed 0.5 km earlier on the route. In each study area, we also recorded mobs forming on our first trials, typically near the trap sites (figure 1 and the electronic supplementary material), suggesting that other crows learned by observing the capture, or were influenced by the contagion of those who had so learned.
(d). Use of public information for observational conditioning during the short-term study
The contagious nature of scolding behaviour was indicated by the formation of mobs (see above). Many or all individuals in a mob may have learned the identity of the dangerous person by observation, but our tagging design only enabled us to answer this question outside of the mob. The actions of lone, unbanded crows are consistent with both horizontal and vertical social learning.
Consistent with horizontal transmission of information, as the number of opportunities to observe mobs scold the mask increased, birds that had never been captured increasingly scolded the mask used for trapping, and more accurately discriminated it from the neutral mask. An opportunity to observe other crows scold the dangerous mask occurred each time we used the dangerous mask in an experimental trial. Scolding by lone, unbanded crows to the dangerous mask was more likely in the second half of our trials at sites A, B, C and D (9.9% of 158 birds encountered scolded) relative to the first half (4.5% of 124 scolded). During these same trials, lone unbanded birds reduced their scolding of neutral masks (5.6% of 562 scolded during the first half of trials; 3.4% of 732 scolded during the second half). The increasing ability of lone, unbanded birds to discriminate dangerous from neutral faces was significant (interaction of mask type with trial half: F1,32 = 4.44, p = 0.043).
The population-level increase in mobbing by lone, unbanded birds throughout our experiments was mirrored at the individual level. At locations where we repeatedly observed the actions of lone, unbanded crows and where we never observed a mob scold the dangerous mask, only 3.3 per cent of 181 lone, unbanded crows scolded the dangerous mask (see figure 1 and the electronic supplementary material). However, at locations where we recorded mobs scolding the dangerous mask, subsequently observed lone, unbanded crows were significantly more likely to scold the dangerous mask (27.8% of 54 crows; 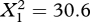 , p < 0.0001).
, p < 0.0001).
Consistent with vertical transmission of information, young crows that never witnessed the capture of their parents by people wearing the dangerous mask learned to scold the dangerous mask by observing and participating in mobs with their parents. Fledglings rarely scolded a masked or unmasked person in their natal territory that was not previously scolded by their parents (this is typical in our study area as in previous experiments only 3% of crows spontaneously scold a neutral mask or unmasked person; [26]). When parents were not present, families of fledglings who had observed their parents scold the dangerous mask were significantly more likely to scold the dangerous mask (7 of 11 territories) than they were to scold the same person without a mask (one of nine territories; Fisher's exact test: 1 d.f., p = 0.03). Increased likelihood of scolding the dangerous mask remained evident in a subset of eight families that were exposed to both the dangerous mask and the unmasked (neutral) observer when parents were not present (seven scolded only the dangerous, one scolded neither; McNemar's Test using the binomial distribution: p = 0.035).
(e). Accuracy of social learning
Whether crows directly or indirectly experienced the dangerous mask affected their discrimination abilities. At all sites, crows directly experiencing trapping by the dangerous mask later discriminated among dangerous and neutral masks more precisely than did crows only indirectly exposed to the danger of the trapper's face. We recorded the responses of 10 crows previously captured by masked trappers (of the 40 trapped) on a total of 41 instances as they later encountered masked people. These crows immediately and consistently scolded anyone wearing the dangerous mask (21 out of 23 encounters, 91.3%), but rarely scolded people wearing one of the neutral masks (2 out of 18 encounters, 11.1%: Fisher's exact test: 1 d.f., p < 0.0001). By contrast, crows that were not captured by masked trappers scolded people wearing the dangerous mask less frequently (137 out of 421 encounters, 33.0%). While this response was significantly below the response of crows captured by people wearing the dangerous mask (Fisher's exact test: 1 d.f., p < 0.0001), it was significantly greater than their response to neutral masks (birds not captured by the dangerous mask scolded neutral masks on 107 of 904 encounters, 11.8%; Fisher's exact test: 1 d.f., p < 0.0001).
4. Discussion
Individuals face evolutionary trade-offs between the acquisition of costly but accurate information gained firsthand and the use of inexpensive but possibly less reliable social information [7]. Theoretical models investigating the adaptive advantages of different forms of learning conclude that individuals should adopt flexible strategies that dictate the circumstances under which they copy others [42–45]. Our study exemplified such flexibility as American crows garnered knowledge of a dangerous event through direct experience, direct observation and subsequent horizontal and vertical transmissions of information about the event. Direct individual experience with a dangerous human enabled more precise learning by crows, but through observation of trapping and observing or joining others in mobs scolding the dangerous person [46], individuals also learned to accurately identify dangerous people. Few studies have demonstrated the combined influence of individual and social learning on alarm calling [47].
Mobbing of potential predators by corvids and many other birds is risky [48], but extremely beneficial as it enables self advertisement [49], reduces future predation risk [50], and may facilitate social learning of predator [29]. Our observations confirm the latter benefit, which is not mutually exclusive of other benefits. Scolding birds provide public information [8] that is used by members of the mob to learn about novel threats. Social learning from public information enables rapid information sharing among peers (horizontal transmission) and from parent to young (vertical transmission; [8,29,51]). Horizontal and vertical transmissions of information about danger for months to years after the dangerous act was committed appeared to account for much of the scolding we observed. In our experiments, direct experience with danger or direct observation of the dangerous activity accounted for less, albeit immediate, learning in the population. Mobs where the social learning occurred included groups of knowledgeable parents and naive offspring as well as groups of territory neighbours and non-breeding individuals. Learning by young birds provides a mechanism for threat recognition to spread several kilometres per generation with normal dispersal [38]; a distance consistent with our observations around the UW site.
The type of danger we simulated may in part account for the responses and routes of learning that we observed. While we demonstrated to crows a sure risk (being captured), we released all captured birds unharmed. Learning by observing a killing, rather than capture and release, may involve more direct use of public information at the scene. If true, our experiment may have underestimated the role of this learning pathway in more typical natural encounters with deadly predators. Continued learning from peers and parents however should remain important, perhaps even increasing in accuracy in response to increasing risk.
Our estimation of social learning by unbanded, lone crows is surely biased, but we do not think this bias is substantial. Our assumption that repeated observations of lone birds at a particular location were of one individual might overestimate social learning if these crows were actually different individuals in each trial. Such birds may have learned about the dangerous mask by watching the trapping event, then left the area immediately afterwards, only to gradually return and scold us. Our results suggest this is unlikely (there were only three locations where scolding by a lone crow after exposure to a mob had been preceded by an unexposed lone crow that also scolded (sites A and B; figure 1 and the electronic supplementary material). It is also likely that we underestimated horizontal social learning because some lone crows who scolded the dangerous mask at locations where mobs did not previously form may have experienced a mob elsewhere. We cannot estimate the frequency of this bias. However, the number of unbanded birds throughout each study area that scolded increased as the occurrence of mobs increased, suggesting that biased overestimation of peer-to-peer learning is less than biased underestimation.
In our long-term study area, a linear increase in scolding over a 5 year period as the number of opportunities to observe scolding, but not trapping, increased was most consistent with our observations. Steady increase in scolding for years after trapping is consistent with models of ongoing social learning [35,36], and suggests that mobs contain contagious naive observers who are learning and socially informed or directly experienced knowledgeable scolders who are demonstrating [25,29,52]. It is unlikely to result from a changing population (during the course of our research, the crow population in Seattle has remained high [38]; and more recently fluctuated between 93.4 crows per party-hour in 2005 and 80.7 crows per party-hour in 2009). Increased scolding, rather than extinction of it, years after trapping, may also demonstrate an ongoing, reinforcing reward of learning. In response to scolding, masked observers eventually left the study area, so preventing another aversive event (more trapping) by scolding, mobbing and becoming increasingly alert may reinforce mobbing by crows.
Our juvenile crows that learned of the dangerous mask by observing their parents, both joined in scolding with mobs of knowledgeable crows and also scolded without any other knowledgeable birds present. However, when the juveniles were scolding on their own, they never attracted other birds to join them and mob. Strength of response depends on age in a variety of animals [47,53–55]. In corvids, mobbing Florida scrub-jay fledglings were often ignored by adults that could not see the predator [53]. Thus, over time as the knowledgeable juveniles mature and fully develop their calls, we expect them to attract others to mob the dangerous mask.
The ability to recognize individual conspecifics is an essential feature for forming affiliative relationships, understanding relationships between third parties and classifying individuals based on their social class [56]. Thus, it stands to reason that it would be beneficial to recognize individual members of a potential predator species, especially one in which the individuals vary in their behaviour. Perhaps, no predator is as individualistic as a human. Humans can be beneficial, for example, during our study, several people we encountered said they fed the crows at their homes, or deadly; three crows were shot at one of our study sites. Thus, flexibility in response is crucial in order to maximize fitness by taking advantage of friendly humans and avoiding potentially lethal ones.
Acknowledgements
All crows were trapped, handled and marked using devices and protocols supported by the United States Bird Banding Laboratory (permit 22489) and the University of Washington Animal Care and Use Committee (protocol 3077-01).
The authors thank L. Aguirre, A. Borte, M. Brown, M. Campbell, C. Chan, F. Clinch, S. Collins, J. Eager, S. Hardwick, K. Helem, L. Hudnell, R. Kahlo, S. Kim, C. Larkin, J. Marzluff, K. Oh, H. and S. Pecararo, C. Perez, J. Phipps, B. Pochmershi, G. Rimbey, L. Sandstrom, J. Shen, K. Shipe, E. Shulenberger, T. Stilson, J. Tomasevic, W. Trepel, J. Vanderhoof, B. Wang, J. Warner, M. Whittaker, G. Wibisono and A. Yahnke for conducting blind trials. D. Craig provided field and intellectual assistance. J. DeLap prepared the figures. B. Clucas, J. DeLap, K. Dial, P. Marra, B. Shryock, C. Templeton, J. Tomasevic, M. Colon, K. Swift and M. Thomas provided editorial comments. Funding was provided by the US Army, the Denman professorship in the College of Forest Resources at the University of Washington, and by a Research Opportunity Award for NSF BCS Award no. 0508002.
References
- 1.Grant P. R., Grant B. R. 2007. How and why species multiply: the radiation of Darwin's finches. Princeton, NJ: Princeton University [Google Scholar]
- 2.Johnston T. D., Pietrewicz A. T. (eds) 1985. Issues in the ecological study of learning. Hillsdale, NJ: L. Erlbaum Associates [Google Scholar]
- 3.Staddon J. E. R. 1983. Adaptive behavior and learning. Cambridge, UK: Cambridge University Press [Google Scholar]
- 4.Marzluff J. M. 1988. Do pinyon jays use prior experience in their choice of a nest site? Anim. Behav. 36, 1–10 10.1016/S0003-3472(88)80244-6 (doi:10.1016/S0003-3472(88)80244-6) [DOI] [Google Scholar]
- 5.Feldman M. W., Aoki K., Kumm J. 1996. Individual versus social learning: evolutionary analysis in a fluctuating environment. Anthropol. Sci. 104, 209–231 [Google Scholar]
- 6.Rogers A. R. 1988. Does biology constrain culture? Am. Anthropol. 90, 819–831 10.1525/aa.1988.90.4.02a00030 (doi:10.1525/aa.1988.90.4.02a00030) [DOI] [Google Scholar]
- 7.Boyd R., Richerson P. J. 1995. Why does culture increase human adaptability? Ethol. Sociobiol. 15, 125–143 10.1016/0162-3095(94)00073-G (doi:10.1016/0162-3095(94)00073-G) [DOI] [Google Scholar]
- 8.Danchin E., Giraldeau L.-A., Valone T. J., Wagner R. H. 2004. Public information: from noisy neighbors to cultural evolution. Science 305, 487–491 10.1126/science.1098254 (doi:10.1126/science.1098254) [DOI] [PubMed] [Google Scholar]
- 9.Castro L., Toro M. A. 2004. The evolution of culture: from primate social learning to human culture. Proc. Natl Acad. Sci. USA 101, 10 235–10 240 10.1073/pnas.0400156101 (doi:10.1073/pnas.0400156101) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Avital E., Jablonka E. 2000. Animal traditions: behavioural inheritance in evolution. Cambridge, UK: Cambridge University Press; [DOI] [PubMed] [Google Scholar]
- 11.Boyd R., Richerson P. J. 1988. Culture and the evolutionary process. Chicago, IL: University of Chicago Press [Google Scholar]
- 12.Marzluff J. M., Shulenberger E., Endlicher W., Alberti M., Bradley G., Ryan C., Simon U., ZumBrunnen C. (eds) 2008. Urban ecology: an international perspective on the interaction between humans and nature. New York, NY: Springer [Google Scholar]
- 13.Clucas B. A., Marzluff J. M. 2011. Coupled relationships between humans and other organisms in urban areas. In Urban ecology: patterns, processes, and applications (ed. Niemelä J.), pp. 135–147 Oxford, UK: Oxford University Press [Google Scholar]
- 14.Griffin A. S., Boyce H. M. 2009. Indian mynahs, Acridotheres tristis, learn about dangerous places by observing the fate of others. Anim. Behav. 78, 79–84 10.1016/j.anbehav.2009.03.012 (doi:10.1016/j.anbehav.2009.03.012) [DOI] [Google Scholar]
- 15.Levey D. J., Londono G. A., Ungvari-Martin J., Hiersoux M. R., Jankowski J. E., Poulsen J. R., Stracey C. M., Robinson S. K. 2009. Urban mockingbirds quickly learn to identify individual humans. Proc. Natl Acad. Sci. USA 106, 8959–8962 10.1073/pnas.0811422106 (doi:10.1073/pnas.0811422106) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Marzluff J. M., Angell T. 2005. Cultural coevolution: how the human bond with crows and ravens extends theory and raises new questions. Ecol. Anthropol. 9, 69–75 [Google Scholar]
- 17.Olsson A., Phelps E. A. 2007. Social learning of fear. Nat. Neurosci. 10, 1095–1102 10.1038/nn1968 (doi:10.1038/nn1968) [DOI] [PubMed] [Google Scholar]
- 18.Heyes C. M. 1994. Social learning in animals: categories and mechanisms. Biol. Rev. 69, 207–231 10.1111/j.1469-185X.1994.tb01506.x (doi:10.1111/j.1469-185X.1994.tb01506.x) [DOI] [PubMed] [Google Scholar]
- 19.Whiten A. 2000. Primate culture and social learning. Cogn. Sci. 24, 477–508 10.1207/s15516709cog2403_6 (doi:10.1207/s15516709cog2403_6) [DOI] [Google Scholar]
- 20.Zentall T. R. 2006. Imitation: definitions, evidence, and mechanisms. Anim. Cogn. 9, 335–353 10.1007/s10071-006-0039-2 (doi:10.1007/s10071-006-0039-2) [DOI] [PubMed] [Google Scholar]
- 21.Byrne R. W. 2005. Social cognition: imitation, imitation, imitation. Curr. Biol. 15, R498–R500 10.1016/j.cub.2005.06.031 (doi:10.1016/j.cub.2005.06.031) [DOI] [PubMed] [Google Scholar]
- 22.Box H. O., Russon A. E. 2004. Socially mediated learning among monkeys and apes: some comparative perspectives. In Comparative vertebrate cognition (eds Rogers L. J., Kaplan G.), pp. 97–140 New York, NY: Kluwer Academic Publishers [Google Scholar]
- 23.Griffin A. S. 2008. Socially acquired predator avoidance: is it just classical conditioning? Brain Res. Bull. 76, 264–271 10.1016/j.brainresbull.2008.02.005 (doi:10.1016/j.brainresbull.2008.02.005) [DOI] [PubMed] [Google Scholar]
- 24.Marzluff J. M., McGowan K. J., Donnelly R., Knight R. L. 2001. Avian ecology and conservation in an urbanizing world (eds Marzluff J. M., Bowman R., Donnelly R.), pp. 331–364 Boston, MA: Kluwer [Google Scholar]
- 25.Brown E. D. 1985. The role of song and vocal imitation among common crows (Corvus brachyrhynchos). Z. Tierpsychol. 68, 115–136 10.1111/j.1439-0310.1985.tb00119.x (doi:10.1111/j.1439-0310.1985.tb00119.x) [DOI] [Google Scholar]
- 26.Marzluff J. M., Walls J., Cornell H. N., Withey J. C., Craig D. 2010. Lasting recognition of threatening people by wild American crows. Anim. Behav. 79, 699–707 10.1016/j.anbehav.2009.12.022 (doi:10.1016/j.anbehav.2009.12.022) [DOI] [Google Scholar]
- 27.Zentall T. R., Sutton J. E., Shelborne L. M. 1996. True imitative learning in pigeons. Psychol. Sci. 7, 343–346 10.1111/j.1467-9280.1996.tb00386.x (doi:10.1111/j.1467-9280.1996.tb00386.x) [DOI] [Google Scholar]
- 28.Griffin A. S. 2004. Social learning about predators: a review and prospectus. Learn. Behav. 32, 131–140 10.3758/BF03196014 (doi:10.3758/BF03196014) [DOI] [PubMed] [Google Scholar]
- 29.Curio E., Ernst U., Vieth W. 1978. Cultural transmission of enemy recognition: one function of mobbing. Science 202, 899–901 10.1126/science.202.4370.899 (doi:10.1126/science.202.4370.899) [DOI] [PubMed] [Google Scholar]
- 30.Gursky S. 2006. Function of snake mobbing in spectral tarsiers. Am. J. Phys. Anthropol. 129, 601–608 10.1002/ajpa.20364 (doi:10.1002/ajpa.20364) [DOI] [PubMed] [Google Scholar]
- 31.Graw B., Manser M. B. 2007. The function of mobbing in cooperative meerkats. Anim. Behav. 74, 507–517 10.1016/j.anbehav.2006.11.021 (doi:10.1016/j.anbehav.2006.11.021) [DOI] [Google Scholar]
- 32.Ellis J. M. 2008. Which call parameters signal threat to conspecifics in white-throated magpie-jay mobbing calls? Ethology 114, 154–163 10.1111/j.1439-0310.2007.01439.x (doi:10.1111/j.1439-0310.2007.01439.x) [DOI] [Google Scholar]
- 33.Emlen J. T. 1936. Age determination in the American crow. Condor 38, 99–102 10.2307/1363907 (doi:10.2307/1363907) [DOI] [Google Scholar]
- 34.Yorzinski J. L., Vehrencamp S. L. 2009. The effect of predator type and danger level on the mob calls of the American crow. Condor 111, 159–168 10.1525/cond.2009.080057 (doi:10.1525/cond.2009.080057) [DOI] [Google Scholar]
- 35.Galef B. G., Jr 1991. Interpretation and explanation in the study of animal behaviour (eds Bekoff M., Jamieson D.), pp. 74–95 Boulder, CO: Westview [Google Scholar]
- 36.Lefebvre L. 1995. The opening of milk bottles by birds: evidence for accelerating learning rates, but against the wave-of-advance model of cultural transmission. Behav. Proc. 34, 43–53 10.1016/0376-6357(94)00051-H (doi:10.1016/0376-6357(94)00051-H) [DOI] [PubMed] [Google Scholar]
- 37.Hoppitt W., Boogert N. J., Laland K. N. 2010. Detecting social transmission in networks. J. Theor. Biol. 263, 544–555 10.1016/j.jtbi.2010.01.004 (doi:10.1016/j.jtbi.2010.01.004) [DOI] [PubMed] [Google Scholar]
- 38.Withey J. C., Marzluff J. M. 2005. Dispersal by juvenile American crows (Corvus brachyrhynchos) influences population dynamics across a gradient of urbanization. Auk 122, 205–221 10.1642/0004-8038(2005)122[0205:DBJACC]2.0.CO;2 (doi:10.1642/0004-8038(2005)122[0205:DBJACC]2.0.CO;2) [DOI] [Google Scholar]
- 39.SPSS 2003. SPSS for windows, v. 12.0. Chicago, IL: SPSS Inc [Google Scholar]
- 40.Systat 2008. SigmaPlot, v. 11.0. San Jose, CA: Systat Software, Inc [Google Scholar]
- 41.Cytel 2003. StatXact6. Cambridge, MA: Cytel Software Corporation [Google Scholar]
- 42.Laland K. N. 1996. Is social learning always locally adaptive? Anim. Behav. 52, 637–640 10.1006/anbe.1996.0205 (doi:10.1006/anbe.1996.0205) [DOI] [Google Scholar]
- 43.Laland K. N., Richerson P. J., Boyd R. 1996. Developing a theory of animal social learning. In Social learning in animals: the roots of culture (eds Heyes C. M., Galef B. G., Jr), pp. 129–154 San Diego, CA: Academic Press [Google Scholar]
- 44.Laland K. N., Kendal J. R. 2003. What the models say about social learning. In The biology of tradition: models and evidence (eds Perry S., Fragaszy D.), pp. 33–55 Cambridge, UK: Cambridge University Press [Google Scholar]
- 45.Laland K. N. 2008. Animal cultures. Curr. Biol. 18, 366–370 10.1016/j.cub.2008.02.049 (doi:10.1016/j.cub.2008.02.049) [DOI] [PubMed] [Google Scholar]
- 46.Griffin A. S., Haythorpe K. 2011. Learning from watching alarmed demonstrators: does the cause of alarm matter? Anim. Behav. 81, 1163–1169 [Google Scholar]
- 47.Hollén L. I., Radford A. N. 2009. The development of alarm call behaviour in mammals and birds. Anim. Behav. 78, 791–800 10.1016/j.anbehav.2009.07.021 (doi:10.1016/j.anbehav.2009.07.021) [DOI] [Google Scholar]
- 48.Curio E., Regelmann K. 1986. Predator harassment implies a real deadly risk: a reply to Hennessy. Ethology 72, 75–85 10.1111/j.1439-0310.1986.tb00607.x (doi:10.1111/j.1439-0310.1986.tb00607.x) [DOI] [Google Scholar]
- 49.Slagsvold T. 1984. The mobbing behaviour of the hooded crow, Corvus corone cornix: antipredator defence or self-advertisement? Fauna Norv. C Cinc. 7, 127–131 [Google Scholar]
- 50.Pavey C. R., Smyth A. K. 1998. Effects of avian mobbing on roost use and diet of powerful owls, Ninox strenua. Anim. Behav. 55, 331–338 10.1006/anbe.1997.0633 (doi:10.1006/anbe.1997.0633) [DOI] [PubMed] [Google Scholar]
- 51.Suboski M. D., Bain S., Carty A. E., McQuoid L. M., Seelen M. I., Seifert M. 1990. Alarm reaction in acquisition and social transmission of simulated-predator recognition by zebra danio fish (Brachydanio rerio). J. Comp. Psychol. 104, 101–112 10.1037/0735-7036.104.1.101 (doi:10.1037/0735-7036.104.1.101) [DOI] [Google Scholar]
- 52.Nicol C. J. 1995. The social transmission of information and behaviour. Appl. Anim. Behav. Sci. 44, 79–98 10.1016/0168-1591(95)00607-T (doi:10.1016/0168-1591(95)00607-T) [DOI] [Google Scholar]
- 53.Francis A., Hailman J. P., Woolfenden G. E. 1989. Mobbing by Florida scrub jays: behaviour, sexual asymmetry, role of helpers and ontogeny. Anim. Behav. 38, 795–816 10.1016/S0003-3472(89)80112-5 (doi:10.1016/S0003-3472(89)80112-5) [DOI] [Google Scholar]
- 54.Galef B. G., Laland K. N. 2005. Social learning in animals: empirical studies and theoretical models. Bioscience 55, 489–500 10.1641/0006-3568(2005)055[0489:SLIAES]2.0.CO;2 (doi:10.1641/0006-3568(2005)055[0489:SLIAES]2.0.CO;2) [DOI] [Google Scholar]
- 55.Hawkins L. A., Magurran A. E., Armstrong J. D. 2008. Ontogenetic learning of predator recognition in hatchery-reared Atlantic salmon, Salmo salar. Anim. Behav. 75, 1663–1671 10.1016/j.anbehav.2007.10.019 (doi:10.1016/j.anbehav.2007.10.019) [DOI] [Google Scholar]
- 56.Clayton N. S., Emery N. J. 2007. The social life of corvids. Curr. Biol. 17, R652–R656 10.1016/j.cub.2007.05.070 (doi:10.1016/j.cub.2007.05.070) [DOI] [PubMed] [Google Scholar]