

Abstract
Throughout the centuries, scientific observers have endeavoured to extend their knowledge of the interrelationships between the brain and its regulatory control of human emotions and behaviour. Since the time of physicians such as Aristotle and Galen and the more recent observations of clinicians and neuropathologists such as Broca, Papez, and McLean, the field of affective neuroscience has matured to become the province of neuroscientists, neuropsychologists, neurologists, and psychiatrists. It is accepted that the prefrontal cortex, amygdala, anterior cingulate cortex, hippocampus, and insula participate in the majority of emotional processes. New imaging technologies and molecular biology discoveries are expanding further the frontiers of knowledge in this arena. The advancements of knowledge on the interplay between the human brain and emotions came about as the legacy of the pioneers mentioned in this field. The aim of this paper is to describe the historical evolution of the scientific understanding of interconnections between the human brain, behaviour, and emotions.
Keywords: Limbic system, behaviour, emotions, neurosciences, historical article
1. INTRODUCTION
Emotions have been defined as a group of interrelated superior cerebral functions, resulting from states of reward and punishment [1, 2]. Behavioural rewarding conditions reinforce certain reactions, which are expressed by animals, including human primates, in a quest to experience a favourable result, which brings satisfaction, comfort, or wellbeing. As a principle, animals escape from and avoid punishment or harmful consequences [1]. A series of findings in the affective neurosciences have outlined the neural circuits encompassing cortical and subcortical structures, which are responsible for the generation of human emotions. It is currently accepted that the following areas participate in the majority of the emotional processes: prefrontal cortex, amygdala, anterior cingulate cortex, hippocampus, and insula [3].
The prosencephalon (forebrain), the mesencephalon (midbrain), and rhombencephalon (hindbrain) are the three primary brain structures formed during the early embryonic development of the human system [4]. The forebrain is formed by the telencephalon and the diencephalon. The telencephalon is the most cranial region of the human central nervous system, which matures to form the cerebrum. The dorsal part of the telencephalon, or pallium, develops into the cerebral cortex, while the ventral telencephalon, or subpallium, forms the basal ganglia. The diencephalon encompasses the thalamus, metathalamus, hypothalamus, epithalamus, prethalamus or subthalamus and pretectum. The midbrain is centrally located below the cerebral cortex and above the pons. It includes the tectum, the tegmentum, the ventricular mesocoelia, and the cerebral peduncles. The pons, or metencephalon, in association with the myelencephalon and additional subunits called rhombomeres, form the rhombencephalon (or hindbrain), which represents a transition to the spinal cord.
Some regions of the mesencephalon, diencephalon, and telencephalon are structurally and functionally interrelated so that they can be considered as a unique functional complex, the so-called limbic system [4]. On the whole, this system is characterized by direct involvement in processes put in place to guarantee the survival of the individual and species (Figures 1 and 2) [4–11].
Figure 1.
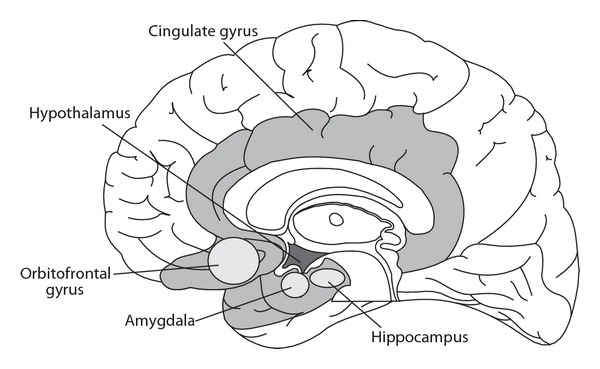
Anatomical illustration of important areas of the limbic system.
Figure 2.
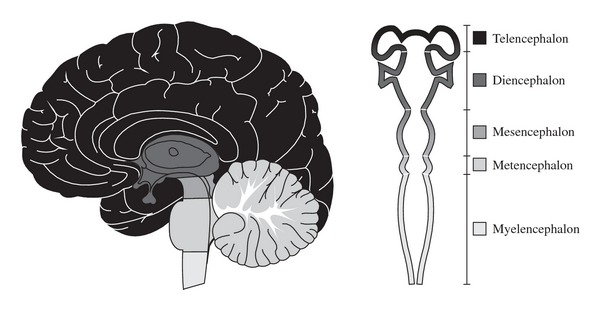
Anatomical representation of the developmental subdivisions of the brain.
The medial cortex was named by Broca (1824–80), as “the great limbic lobe,” due to its oval shape (in French, limbique means hoop). Subsequently; however, the limbic lobe started to be called rhinencephalon, which means olfactory brain, due to its apparent involvement with the olfactory process and behaviours generated by olfaction [6]. In order to understand the concept of limbic system, it is important to understand the term rhinencephalon, whose origins are difficult to trace [12]. The term was firstly used by Saint-Hillarie to name a one-eyed monster. Soon after, Owen (1804–1902) used the term which means cerebral nose in a neuroanatomical context, referring to the olfactory bulb and the peduncle [12]. Later, Turner (1832–1916) extended its meaning to include the pyriform lobe. In fact, some neuroscientists consider many of the limbic structures as integrant parts of the rhinencephalon, which is entirely confined to the telencephalon [4].
2. ANCIENT TIMES: THE FIRST EMPIRIC THEORIES
Since ancient times, searches for the evidence for the existence of controlling centres of the emotions commenced with Aristotle (384 BC–322 BC), in Ancient Greece, who stated that the centre of intelligence and emotions was the heart and that memory would generate learning based on emotions and feelings [13]. However, after a long-lasting hegemony of the Aristotelian theories, some changes took place, which allowed for a better understanding of human psychic foundations. These changes occurred as a result of increasing interest in the dissection of corpses and the resulting progress in the understanding of human anatomy [14, 15]. Galen (130 AD–200 AD) conveyed the most complete and influential comments on neuroanatomy. He also developed the most sophisticated investigations into cerebral functions before the Renaissance period. Among others contributions, Galen developed theories on the somatic senses, having described the anatomy of the cranial nerves and the autonomic nervous system [14]. Contrary to Aristotle, he believed that the brain, and not the heart, was the centre of intelligence and that the “animal spirits,” although originating in the heart, were sent to the cerebral ventricles via the circulatory system [14].
Later, with the anatomical contributions put forward by Galen, the cerebral ventricles were considered the centres of reason and emotions [14]. According to the “cerebral ventricles theory of emotions,” the information resulting from the five senses—touch, taste, smell, hearing and sight—would be processed in the cerebral ventricle, by the so-called “common sense,” and grouped as a unique perception. This perceptual input would then travel through the “internal senses”: fantasy, imagination, cognition, estimate, and memory. This was how the emotional process was thought to be generated. A ventricular theory of emotions was also proposed by Saint Augustine in 500 AD [6]. Although the exact date when this theory was conceived is unknown, it certainly evolved from Galen's fundamental theories of brain anatomy [14, 15].
3. MIDDLE AGES—MODERN AGE: THE SCIENTIFIC FOUNDATION
Da Vinci (1452–1519) made significant contributions to the development of neuroscience, particularly in neuroanatomy and neurophysiology. Regarding the development of human emotional processes, he directed his research to the quest for a biological explanation of the brain processes responsible for visual perception, as well as other sensorial modalities, trying to integrate these senses with an understanding of the mind [16, 17]. Da Vinci correlated cerebral structures to superior cerebral functions, and, for this reason, he can be considered the forerunner of the theory developed centuries later by a Viennese doctor named Franz J. Gall, called phrenology [5].
The printing press, invented by the Johannes Gutenberg around 1440, made possible for the first time the rapid creation of metal movable type in large quantities, which subsequently led to a boom in printing activities in Europe between 1450 and 1500. The ability to publish manuscripts was an important technological advance that contributed to the understanding of neuroanatomy and neurology, as known today [15, 18]. Thus, in 1499 Peyligk graphically described part of the cerebral anatomy, including the dura mater and the pia mater, as well as the ventricles, in his work entitled Compendium philosophiae naturalis. In 1543, the publication of the groundbreaking work De humani corporis fabrica libri septem by Vesalius (1514–1564) truly revolutionized neuroanatomy. It was the most complete and detailed work in this field and corrected several inaccuracies from the works of Galen. It revealed details about the cerebral ventricles, cranial and peripheral nerves, pituitary gland, meninges, ocular structures, cerebral vascular supply, and spinal cord [15].
As a corollary to the evolution of the medical sciences, it became evident that the advancement of scientific knowledge, specifically neuroanatomy, depended on the capacity to create accurate reproduction of images. Some scholars believe that one reason that accounted for a lack of a major advance in Medicine during the Renaissance was the inability to reproduce with graphical perfection the scientific and anatomical findings [15].
The study of anatomy flourished in the 17th and 18th centuries, given that famous artists, including Michelangelo and Rembrandt, studied anatomy, attended dissections, and published their drawings. Certified anatomists were allowed to perform dissections in many European cities, depending on the availability of fresh bodies [19]. These developments enabled a more accurate description of complex structures of the nervous system. Moreover, the rise of neurochemistry took place in the 1780s in France. Antoine-François de Fourcroy, a leading investigator trained in medicine and chemistry, was puzzled by the preserved status of the brains of exhumed bodies during the removal of a cemetery in Paris. He led studies to examine the nature of brain substances that could retard putrefaction [19]. As a result, through the convergence of previously distinct biological disciplines including anatomy and chemistry, it became possible to speculate about the molecular biology of the cerebral systems responsible for the production of emotions [20].
At the end of the eighteenth century, neurology had developed from a science with poor anatomical groundwork to a more concise, practical, and less philosophical combination of anatomy, pathology, and neurochemistry [15, 21].
4. CONTEMPORARY AGE: THE TERM LIMBIC SYSTEM
In the nineteenth century, medical knowledge gained new impetus as important discoveries occurred. Henle combined anatomy with human biology to create the field of physiology, whereas Virchow and Pasteur established the fields of cellular pathology and microbiology, respectively. In addition, major advancement allowed for safer and painless surgical procedures in that period. Lister advocated for the disinfection of surgical equipment, whereas Morton developed anaesthetic techniques. In addition, new concepts in neuroscience emerged [22]. The invention of the microscope and the development of histological staining made it possible for Ramón y Cajal to identify the neuron as the anatomical and functional unit of the nervous system. Neurology flourished as a discipline and became similar to what it is today: an independent field of research of the complex functions and dysfunctions of the nervous system [15].
In the nineteenth century Gall believed, similar to several of his precursors, that the brain was organized according to different abilities and physically defined specific functions. Gall suggested that each of these functions would be generated in an appropriate “organ” of the brain, as, for example, intelligence, speech, and memory [6]. This theory was called “craniology” (science of the head) by Gall. Later, the nomenclature was modified to “organology” (science of the organs of the brain), and currently it is called “phrenology” (science of the mind) [6, 23]. Nowadays it is known that the theories of Gall were not correct. However, phrenology was the first theory to consider the cerebral location of specific functions, being the precursor of modern theories due to its strong emphasis on localization of cerebral function [6].
In an effort to advance knowledge of the limbic system, Broca established the Société d'Anthropologie (Society of Anthropology), where debates about the origin of the family, human race, human intelligence, and the organization of the human brain took place [24]. The year 1860 was marked by discussions about whether the cerebral hemispheres acted as independent units and whether there were specialized regions in the human brain [24]. In 1861, Broca examined the brain of one of his patients and concluded that the centre of speech was located in the inferior frontal gyrus in the dominant hemisphere. Broca had related injuries in this area to the loss of speech and referred for the first time to the French word aphémie. Later, the eponyms Broca's aphasia and Broca's area became widely known [24]. Broca also studied eight cases of left frontal injuries and subsequently developed the concept of cerebral dominance [25, 26]. Furthermore, he also coined the term “Great Limbic Lobe,” which was later named “Broca's Great Limbic Lobe” [11, 12, 27–31].
Substantial progress in the understanding of the association between cortical damage and behavioural changes came from an observation by Harlow in Vermont, USA, in 1848. A healthy twenty-five-year-old man suffered an accident, in which an iron bar passed through his skull, affecting the pre-frontal cortex region [32, 33]. The patient was purportedly in perfect physical condition less than two months later, except that a bizarre behavioural change had developed. Testing of his executive functioning indicated that he had lost the ability to use anticipatory planning as well as becoming socially awkward [32–36].
Subsequently, a significant revolution in the concept of the emotions took place under the influence of Darwin's seminal ideas. With his book “Expression of emotions in man and animals,” Darwin became the precursor, together with the pioneering American psychologist William James twelve years later, of a research field currently called “affective neuroscience” [3, 37]. Darwin proposed two major postulates in relation to mammalian emotional processes. The first was that emotions in animals would be similar to human emotions—a logical extension of his work on the evolution of the species [37, 38]. Darwin also proposed that humans expressed vestigial patterns of mammalian emotional behaviour, by exposing the front teeth when expressing sadness through anger or crying [37, 38].
The second postulate proposed by Darwin states that there is a set of basic or fundamental emotions that are present throughout distinct species and are independent of cultures or societal norms. These emotions include anger, fear, surprise, and sadness [37, 38]. Both tenets were of great relevance for the field of affective neuroscience, since they have spawned investigations involving animals as a resource to understand human emotions. Moreover, these ideas generated new research on distinct neural substrates for a series of emotional expressions [39].
The quest for a reasonable explanation for the emotional processes began in 1884, with James' article entitled “What is an emotion?” [40]. James proposed an innovative theory whereby various human emotions occurred in response to afferent feedback loops from sensory receptors in the skin, muscles, cartilage, and other organs which produced, although unknown at that time, physical changes that were subsequently encoded into the cerebral cortex memory storage to determine the subjective quality of the stimuli being experienced as temperature change, pain, vibration, and so forth [37, 40, 41].
According to James' theory, emotions are just one form of experience of a wider array of physical changes that occur in response to emotional stimuli. James advocated that a sensorial feedback occurred from the corporal periphery to the cerebral cortex in the context of an emotionally laden behaviour, thus, determining the subjective quality of such a behavioural experience [1]. James understood that different corporal memory processes encoded different emotions [37, 40, 41]. According to his theory, tremor is the cause of fear and not its consequence, as cry is the cause of sadness [6]. Similar ideas were proposed in the same period by the Danish physician and psychologist Lange [42]. In 1885, this hypothesis was called the “James-Lange theory of emotions” [37]. The James-Lange theory states that the autonomic nervous system generates physiological events as a response to humans' experiential interaction with the world. According to this theory, emotions are feelings that occur as a consequence—instead of being the cause—of physiological changes.
Experiments developed by authors such as Exner, Freud, and Waynbaum led to further advancements [43]. In spite of the then limited knowledge about the cerebral anatomical interconnections, the ideas advocated by these proponents were in line with current tendencies. Sigmund Exner (1846–1926), a physiologist at the University of Vienna and one of the charter members of the German Society of Psychology, described in 1894 a neural circuit model that explained the interactions between sensations of pleasure and aversion in the brain [41, 42]. This model, based on his knowledge of animal experimentation, detailed how sensorial events acquire emotional meaning and produce motor and autonomic responses, anticipating what subsequently would be elaborated in recent neurobiological theories [41]. Thus, the thalamus would function as the centre of sensory integration and as a filter that would direct only intense stimuli to the aversive centre. The aversion would be processed in a structure composed of neuronal bodies, which encompassed, under modern neurobiological perspectives, the amygdala.
This theory underpinned Sigmund Freud's ideas (1856–1939) who, at the beginning of his scientific activities (1895), had described psychological phenomena as forms of nervous energy in neuronal systems, which consisted of diverse cellular types, each with only one function [35, 43–45]. During a painful experience, for example, the representation of a dangerous object would stimulate specific “neuron-keys” that could trigger aversive emotions. Thus, aversive reactions would be manifested during the mnemonic representation of the aversive experience. This is the basis of Freud's neuronal theory [43, 44].
Freud's first postulate, called inertia, is similar to what is today known as homeostasis. It states that an organism, when stimulated, attempts to return to the unstimulated condition. The excitation of states of inertia is conceived in neurophysiologic terms or as “quantities of excitation in flow.” For Freud, a neuron may “fill” or become “cathected” with excitation without necessarily producing any demonstrable physiological response [46]. Neurologically, the affects result when cathexis increases (negative affects) or decreases (positive affects). These changes in cortical cathexis follow the activation of traces imprinted in the nuclear system on prior occasions during episodes of negative or positive affects. The affects, consequently, modulate current experiences with imprints from past impressions [46].
5. TWENTIETH-CENTURY: THE THEORY OF THE LIMBIC SYSTEM
5.1. Understanding the Neural Network
Another model for the understanding of the emotional process was proposed in 1906-1907 by the French physician Israel Waynbaum. According to this model, sudden and unexpected sensory events can bring about an “emotional shock.” This process was explained by a hypothetical neural network, with the emotional centre functioning as a dominant element. According to this theory, the medulla oblongata would take on the function of the “general emotional centre,” with the aim of connecting external emotional perception to internal corporeal sensations [43, 44].
In the 1920s, physiological laboratory studies began to differentiate the psychological and neurophysiological domains, since previously both specialties had investigated only emotional processes [47]. Walter Cannon, a Harvard physiologist and pioneer in neurophysiological studies of emotional substrates, highlighted the fact that laboratory investigations of human emotions were partially hindered by the difficulty of inducing emotional states in animals and of maintaining these states for subsequent studies [48, 49]. The obstacles to conducting research on human emotions in laboratory settings were emphasized in numerous psychological and physiological studies. There were difficulties both in inducing genuine and intense emotional reactions in laboratories as well as the fact that emotions generated in the laboratory environment were considered extremely artificial [50–54].
In 1884, James suggested that emotions could be mediated by sensory and motor areas of the cerebral cortex. He believed that the sensory areas were essential to the immediate detection of stimuli and that the motor regions were responsible for the production of feedback reactions [6, 40]. In the 1920s, Cannon contradicted the prevailing peripheral emotional theory of James. He also proposed a new emotional theory based on investigations from Phillip Bard's laboratory, in which animal brains were longitudinally sectioned in the diencephalon in consecutive inferior anatomical planes [1, 55]. The intention was to find a transection plane that would suppress or diminish the emotional expression in the animal model. This region was defined, according to the experiments, as the caudal half of the hypothalamus and the posteroventral thalamus, which revealed to both Cannon and Bard that those structures were essential for the emotional brain [1, 6]. This theory was later called the “Cannon-Bard theory of emotions.” Additionally, many observations from these experiments corroborated the new theory: surgical decerebration of animals did not effect emotional behaviour and did not prevent generation of the “fictitious rage” process. Fictitious rage is a term coined by Cannon and Bard to describe the rage produced surgically in decerebrated animals. Consequently, physical and autonomic activities were insufficient to distinguish between distinct emotional states, and physical changes are extremely slow to generate emotions as a result of hormonal activation induced by such physical activity. They postulated that the hypothalamus could receive afferent impulses from the thalamus at the same time as the thalamus sends information to the cerebral cortex.
The hypothalamus would have access to emotions at the same time as the cortex and would, therefore, stimulate behavioural and autonomic bodily reactions typical of affective states. This would explain, according to Cannon and Bard, why decortication could not prevent the genesis of emotional patterns, a finding that opposed the James-Lange theory [1, 6, 55–58]. Findings demonstrated by the Swiss physiologist Walter R. Hess in the same period were in agreement with the Cannon-Bard theory. In experimental works Hess conducted research in which electrodes were implanted in the hypothalamic area of cats. After electrical stimulation, the animals presented a “defence affective reaction” that was associated with an increase in heart rate and a noticeable propensity to alertness and attack reactions [37].
Further advancement in laboratory investigations of emotions was achieved via the development of surgical decerebration in animals, albeit strong opposition and political pressure from animal liberation and antivivisection movements [47]. The decerebration made possible manipulation of animals without imposing painful sensations and allowed scientists to explore stimuli, which would otherwise cause pain to the unanaesthetised animals [47, 55]. This strategy set a benchmark upon which new behavioural models and hypotheses were based. This fact was ultimately responsible for a historic shift of paradigms in the neurosciences and human emotion research. The truncated brain, produced through decerebration processes, spurred a separation between research on emotions as conducted by psychologists and by physiologists such as Cannon who, therefore, brought about a major revolution in emotion physiology research [47].
The first evidence that the limbic system was responsible for the cortical representation of emotions was obtained in 1939, when Kluver and Bucy, in Chicago, demonstrated that the bilateral removal of the temporal lobes in monkeys—including the amygdala and the hippocampal formation, as well as the non-limbic temporal cortex—produced an extreme behavioural syndrome [59]. After temporal lobectomy, the hitherto aggressive monkeys became docile and exhibited reduced emotional threshold. They displayed a tendency towards oral behaviour such as attempting to ingest inedible objects. Ablated monkeys also demonstrated hypersexualized behaviour by mounting females of the same and different species. Finally, these animals revealed a reduced threshold to visual stimuli and were, therefore, unable to recognize hitherto familiar objects [60–62]. At that time, this syndrome was designated by its proponents as “the temporal lobe syndrome.” Nowadays, this syndrome is referred to as the Klüver-Bucy syndrome.
It has been subsequently demonstrated that some of these characteristics are accounted for by a form of visual agnosia caused by lesions in the temporal cortex, although the change in reactivity to aversive and reinforcing stimuli is caused by lesions in the amygdala. Similarly, humans submitted to bilateral amygdalectomy as well as to ablation of the amygdala's cortical connections present a Klüver-Bucy-like syndrome with additional mnemonic deficits [60–65].
On the account of a series of experiments conducted in the beginning of the twentieth century, Papez and MacLean correlated the limbic system with emotional patterns observed in humans [11]. In 1937, the American neuroanatomist Papez described a circuit of cerebral connections—the Circuit of Papez—which generates emotion as a result of the flow of information that travels via reciprocal anatomical networks between the hypothalamus and the mesial cerebral cortex [6, 66]. According to Papez, two integration forms of neural information occur in the emotional process: one via the hippocampus and cingulate cortex, which are directly involved with hypothalamic activity, the other via the lateral cortex, which is involved in nonspecific sensory activities mediated by the dorsal thalamus. The Circuit of Papez also includes other cerebral regions with locomotor, mnemic and associative functions [66]. In order for a piece of information to be recorded as a long-term memory, it must pass through Papez's circuit, in that injuries to this circuit can result in memory loss.
It was originally believed that the limbic structures were uniquely associated with the sense of smell. In fact, the olfactory bulb and supplementary olfactory pathways conduct perceptions of smell to limbic structures, including the amygdala, periamygdaloid, and prepyriform cortex. These interconnections confirm an association between olfaction and emotional mechanisms [67]. This also was proposed originally by Papez, who reported emotional disorders in some of his patients who presented with lesions in the hippocampus and in the cingulate gyrus [66]. The hypothesis of Papez states that afferent sensory stimuli are subdivided in the thalamic region into thought flow and emotional flow. Through sensory nervous stimuli from the thalamus to the lateral regions of the neocortex, sensations would be generated. According to Papez, through this flow of nervous stimuli sensations would be transformed into perceptions, thoughts and memories. The flow of feelings would also extend to the thalamus and its connections, although at thalamic level a direct retransmission to the hypothalamus would occur, initiating in this way—according to Cannon—the emotional process [6, 55, 58, 66]. Papez maintained, though, that the participation of the cerebral cortex in the subjective process of emotions was essential [66].
5.2. Defining the Anatomic Dimensions of the Limbic System
The concept of the limbic system was later redefined and expanded by the American physician and neuroscientist MacLean, who reintroduced the term “limbic,” not exclusively with neuroanatomical connotations, but at first as a descriptor of a complex system related to emotional functioning [68, 69]. To come to these conclusions, he correlated the theories of Papez and Cannon-Bard with the discoveries of Kluver and Bucy. MacLean noted that the stimulation of rhinencephalic regions generated autonomic reactions, such as changes in the respiratory pattern, blood pressure, and heart rate. As a result, he referred to the rhinencephalon as the “visceral brain” and stated that it could correlate with disorders such as hypertension, asthma, and peptic ulcer [6, 69]. To MacLean, the hippocampus was the nucleus of the visceral brain. Subsequently he coined the term “limbic system” to designate the visceral brain, with the intention of characterizing a functional system instead of an exclusively anatomic system [68]. This way he added the amygdala, septum, and pre-frontal cortex to the previously described Papez's Circuit structures such as the thalamus, hypothalamus, hippocampus, and cingulate cortex. MacLean postulated that the expression of social behaviours has a central function in the evolutionary development of the neocortex. MacLean initially considered the olfactory system as not essential to the development of the human species, despite the emphasis given by Broca to the association between limbic and olfactory structures [70, 71]. He understood that the Papez Circuit represented a substrate for emotional behaviour. MacLean also perceived that the limbic structures are strategically displayed among areas responsible for somatic functions and those that involve the visceral nervous system [67, 72].
One of the essential ideas promulgated by MacLean was that emotional experiences were formed by the sum of external perceptions and internal bodily sensations [6, 68–70]. In fact, the limbic system also integrates autonomic activities, as well as somatic phenomena via the neocortex, with its projections to the thalamus, midbrain, and spinal cord. Yet, an “extensive overlapping” between these structures in the integration of autonomic and somatic spheres was reported. Both autonomic and somatic spheres' reactions result from localized electrical and chemical stimulation of limbic structures [73].
The limbic structures, therefore, are capable of integrating internal and external sensations, considering that the limbic system includes not only cortical structures, but also subcortical areas (Figure 3) [67]. In addition, MacLean investigated the corticocortical relationships of the frontotemporal region with the limbic system [68]. The entire frontotemporal region is connected to the amygdala and to the rostral part of the hippocampus, and there is also a dense projection to the hypothalamus, to the septal region, and towards some regions of the basal ganglia [68, 74, 75].
Figure 3.
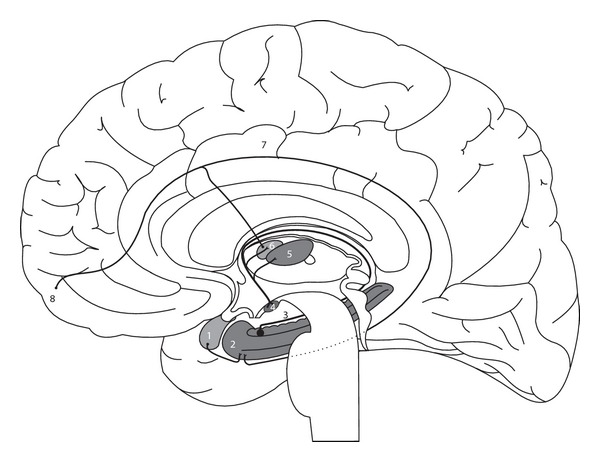
Anatomical representation of the neural pathways involving the main limbic structures in the human brain, adapted from Nieuwenhuys et al. [4]. It includes (1) amygdala, (2) hippocampus, (3) fornix, (4) mammillary body, (5) mediodorsal thalamic nucleus, (6) anterior nucleus of thalamus, (7) cingulate gyrus, and (8) prefrontal cortex.
5.3. From the “Triune Brain” to the Classic Conditioned Reflexes
MacLean proposed the concept of the “triune brain” for the first time in 1969 [70, 76]. According to this evolutionary theory, Man possesses essentially three brains inside one sole brain structure. The oldest is the “reptilian brain,” designated “R-complex,” which consists of the ventral striatum and the basal ganglia. The R-complex is the definition given by MacLean to structures that evolved from the floor of the forebrain during development. He understood that “the reptilian brain” was responsible for typical instinctual behaviours, including territoriality, dominance, and aggression.
The second brain originates from the first and less complex mammals and is designated as the visceral brain or, preferably, limbic system. The third brain, from an evolutionary perspective, is the neocortex, which belongs to nonhuman primates and humans. This is the central structure that makes it possible for humans to acquire the command of symbolic language. These three evolutionally distinct components, according to MacLean, possess the capacity to produce different patterns of behaviours [76, 77]. According to the theory of the “triune brain,” the R-complex is essential for the integration of behavioural patterns involved in self-preservation and in the preservation of the species, as well as for the continuity of nonverbal communication. Furthermore, MacLean stated that epilepsy allows the R-complex to manifest itself physically, in the form of tonic-clonic seizures.
MacLean also conducted observational studies concerning the behaviour of apes after traumatic ablation of temporal regions of the limbic system. He concluded that these animals could not discriminate between different types of food anymore due to damage to the system that promotes the trigger for feeding behavior [76]. The innovative perspective of MacLean, as well as his appreciation of the evolutionary significance and complexity of social systems, offers valuable insights into contemporary neuroscience and psychiatry [37, 70].
Another great researcher of behaviour in animals was the Russian physiologist Ivan P. Pavlov, who, at the beginning of the twentieth century, discovered through painstaking experimentation the classic conditioned reflexes. These reflexes consist of alteration of a response to a given stimulus due to its temporal association with another stimulus; this way, a dog learns to salivate in response to a sound after this has been associated with food [63, 78]. Pavlov also described the instrumental conditioned reflexes, in which the animal uses the emission or the omission of a response as an instrument to receive or avoid the second stimulus [63, 78].
5.4. The Quest for Authenticity
Prior to Broca, elements pertaining to the limbic system had already been described with anatomic precision [79]. In 1958, the concept of a limbic system was expanded by Walle Nauta. The limbic system had hitherto encompassed mainly telencephalic and diencephalic structures. Nauta also was the first to describe the ganglions of the habenulas within the mesencephalon as the integrating part of the system responsible for the emotions [80].
As previously described, the components of the limbic cortex vary according to different neuroanatomical descriptions. Nauta considered as limbic the telencephalic cortical areas, which are reciprocally connected to subcortical areas that spread rostrally from the septum and caudally towards the encephalic trunk [81]. Swanson, in a critical review about the concept of the limbic system, stated that it is undeniable that the hippocampal formation (Ammon's horn, dentate gyrus, and subiculum), together with the cingulate gyrus, pre-frontal region, and discrete subcortical nuclei, are highly interconnected and, therefore, should be considered as a system involved with hypothalamus-related functions [82].
The neuroscientist LeDoux explored several issues, which prevented him from characterizing the limbic system as the only cerebral system in which emotions are generated [6, 83]. The author pointed to several factors to substantiate his claim, such as the absence of structural and functional criteria for the concept of the limbic system and the fact that several limbic regions have not been implicated in emotional processes according to recent functional neuroimaging studies. In addition, he asserted that, whilst various limbic regions participate in emotional processes, they do not function as an integrated and specialized system for the mediation of these processes. Finally, the emotions do not constitute the sole, or even the main, functions of the various limbic areas, many of which are related to cognitive processes—such as the hippocampus being related to mnemonic processes [83].
The multitude of scientific investigations into the limbic system has facilitated its understanding as well as its neuroanatomical identification. The evident association between the limbic system and human emotional processes probably adds to the difficulty in establishing its concept [6, 83, 84]. Conceptually, the idea of the limbic system has evolved anatomically from the Great Limbic Lobe of Broca, the Papez's Circuit, the limbic system of MacLean, and the ideas of Nauta. These terms and concepts of the limbic brain were examined as a hemispheric and independent domain [12].
6. CONCLUSION
Human emotions are essential and intrinsic to human behaviour. Despite our current knowledge about a wide range of human emotions, there is no consensus in the scientific community about how to define the emotions and on which ones are elemental. Throughout the centuries, various scientists have attempted to elucidate the neural systems that control human emotions and behaviour. Research into human emotions and behaviour is vast and is leading exponentially to more questions, which in turn require solutions. For that to happen successfully, efforts should be put into studies based on behavioural genetics, functional neuroimaging investigations, psychopharmacology, and the emerging field of behavioural neuroscience.
DISCLOSURE
The authors hereby confirm that this paper is an original work. It has not been published elsewhere in whole or part. This submission does not contain any material that is libelous, defamatory, or otherwise unlawful. This submission does not contain any material that invades the right of privacy or any proprietary right. This study was not funded by any specific research grant. The authors disclose no conflict of interest. The figures in this paper were produced by one of the authors.
References
- 1.LeDoux JE. Emotion. In: Plum F, editor. Handbook of Physiology, Section 1: The Nervous System. Vol. V. Higher Functions of the Brain. Bethesda, Md, USA: American Physiological Society; 1987. [Google Scholar]
- 2.Rolls ET. International Encyclopedia of the Social & Behavioral Science. Elsevier Science; 2001. The neural basis of emotion; pp. 4444–4449. [Google Scholar]
- 3.Davidson RJ. Darwin and the neural bases of emotion and affective style. Annals of the New York Academy of Sciences. 2003;1000:316–336. doi: 10.1196/annals.1280.014. [DOI] [PubMed] [Google Scholar]
- 4.Nieuwenhuys R, Voogd J, van Huijzen C. The Human Central Nervous System. Berlin, Germany: Springer; 1996. [Google Scholar]
- 5.Rolls ET. The Brain and Emotion. Oxford, UK: Oxford University Press; 1999. [Google Scholar]
- 6.LeDoux J. Emotional networks and motor control: a fearful view. Progress in Brain Research. 1996;107:437–446. doi: 10.1016/s0079-6123(08)61880-4. [DOI] [PubMed] [Google Scholar]
- 7.Aggleton JP. The Amygdala: Neurobiological Aspects of Emotion, Memory and Mental Dysfunction. New York, NY, USA: Wiley-Liss; 1992. [Google Scholar]
- 8.LeDoux JE. A neurobiological view of the psychology of emotion. In: Hirst W, editor. Mind and Brain. New York, NY, USA: Cambridge University Press; 1986. pp. 355–358. [Google Scholar]
- 9.Isaacson RL. The Limbic System. New York, NY, USA: Plenum; 1982. [Google Scholar]
- 10.Brodal A. Neurological Anatomy. New York, NY, USA: Oxford University Press; 1982. [Google Scholar]
- 11.Houssay AB, Cingolani HE. Fisiologia Humana. Porto Alegre, Brazil: Artmed; 2004. [Google Scholar]
- 12.Lautin A. The Limbic Brain. New York, NY, USA: Kluwer Academic Publishers; 2002. [Google Scholar]
- 13.Carlino A. Books of the Body: Anatomical Ritual and Renaissance Learning. Chicago, Ill, USA: University of Chicago Press; 1999. [Google Scholar]
- 14.Green CD. Where did the ventricular localization of mental faculties come from? Journal of the History of the Behavioral Sciences. 2003;39(2):131–142. doi: 10.1002/jhbs.10088. [DOI] [PubMed] [Google Scholar]
- 15.Tessman PA, Suarez JI. Influence of early printmaking on the development of neuroanatomy and neurology. Archives of Neurology. 2002;59(12):1964–1969. doi: 10.1001/archneur.59.12.1964. [DOI] [PubMed] [Google Scholar]
- 16.Del Maestro RF. Leonardo da Vinci: the search for the soul. Journal of Neurosurgery. 1998;89(5):874–887. doi: 10.3171/jns.1998.89.5.0874. [DOI] [PubMed] [Google Scholar]
- 17.Pevsner J. Leonardo da Vinci’s contributions to neuroscience. Trends in Neurosciences. 2002;25(4):217–220. doi: 10.1016/s0166-2236(00)02121-4. [DOI] [PubMed] [Google Scholar]
- 18.Febvre LPV, Martin H-J. The Coming of the Book: The Impact of Printing 1450–1800. London, UK: Verso; 1976. [Google Scholar]
- 19.McLachlan JC, Patten D. Anatomy teaching: ghosts of the past, present and future. Medical Education. 2006;40(3):243–253. doi: 10.1111/j.1365-2929.2006.02401.x. [DOI] [PubMed] [Google Scholar]
- 20.Sourkes TL. The origins of neurochemistry: the chemical study of the brain in France at the end of the eighteenth century. Journal of the History of Medicine and Allied Sciences. 1992;47(3):322–339. doi: 10.1093/jhmas/47.3.322. [DOI] [PubMed] [Google Scholar]
- 21.Sourkes TL. Social and medical origins of neurochemistry. Progress in Neuro-Psychopharmacology & Biological Psychiatry. 2004;28(5):885–890. doi: 10.1016/j.pnpbp.2004.05.030. [DOI] [PubMed] [Google Scholar]
- 22.Turgeon Y, Whitaker HA. Pierre Jean Georges Cabanis (1757–1808): an early nineteenth century source for the concept of nervous energy in European behavioral neurosciences. Brain and Cognition. 2000;43(1-3):412–417. [PubMed] [Google Scholar]
- 23.Greenblatt SH. Phrenology in the science and culture of the 19th century. Neurosurgery. 1995;37(4):790–805. doi: 10.1227/00006123-199510000-00025. [DOI] [PubMed] [Google Scholar]
- 24.Finger S. Paul Broca (1824–1880) Journal of Neurology. 2004;251(6):769–770. doi: 10.1007/s00415-004-0456-6. [DOI] [PubMed] [Google Scholar]
- 25.Finger S. Minds behind the Brain. New York City, NY, USA: Oxford University Press; 2000. Paul Broca; pp. 137–154. [Google Scholar]
- 26.Finger S. Origins of Neuroscience. New York City, NY, USA: Oxford University Press; 1994. [Google Scholar]
- 27.Monod-Broca P. Paul Broca: 1824–1880. Annales de Chirurgie. 2001;126(8):801–807. doi: 10.1016/s0003-3944(01)00600-9. [DOI] [PubMed] [Google Scholar]
- 28.Cubelli R, Montagna CG. A reappraisal of the controversy of Dax and Broca. Journal of the History of the Neurosciences. 1994;3(4):215–226. doi: 10.1080/09647049409525614. [DOI] [PubMed] [Google Scholar]
- 29.Greenblatt SH. The multiple roles of Broca’s discovery in the development of the modern neurosciences. Brain and Cognition. 1984;3(3):249–258. doi: 10.1016/0278-2626(84)90020-4. [DOI] [PubMed] [Google Scholar]
- 30.Roe D, Finger S. Gustave Dax and his fight for recognition: an overlooked chapter in the early history of cerebral dominance. Journal of the History of the Neurosciences. 1996;5(3):228–240. doi: 10.1080/09647049609525672. [DOI] [PubMed] [Google Scholar]
- 31.Stone JL. Paul Broca and the first craniotomy based on cerebral localization. Journal of Neurosurgery. 1991;75(1):154–159. doi: 10.3171/jns.1991.75.1.0154. [DOI] [PubMed] [Google Scholar]
- 32.Damasio AR. Descarte's Error. New York, NY, USA: Putnam; 1994. [Google Scholar]
- 33.Harlow JM. Recovery of the passage of an iron bar through the head. Publication of Massachusetts Medical Society. 1868;2:327–334. [Google Scholar]
- 34.Macmillan M. Inhibition and the control of behavior. From Gall to Freud via Phineas Gage and the frontal lobes. Brain and Cognition. 1992;19(1):72–104. doi: 10.1016/0278-2626(92)90038-n. [DOI] [PubMed] [Google Scholar]
- 35.Macmillan M. Nineteenth-century inhibitory theories of thinking: Bain, Ferrier, Freud (and Phineas Gage) History of Psychology. 2000;3(3):187–217. doi: 10.1037/1093-4510.3.3.187. [DOI] [PubMed] [Google Scholar]
- 36.Ratiu P, Talos IF. Images in clinical medicine. The tale of Phineas Gage, digitally remastered. The New England Journal of Medicine. 2004;351(23):p. e21. doi: 10.1056/NEJMicm031024. [DOI] [PubMed] [Google Scholar]
- 37.Dalgleish T. The emotional brain. Nature Reviews Neuroscience. 2004;5(7):582–589. doi: 10.1038/nrn1432. [DOI] [PubMed] [Google Scholar]
- 38.Darwin C. The Expression of the Emotions in Man and Animals. Chicago, Ill, USA: Chicago University Press; 1872-1965. [Google Scholar]
- 39.Ekman P. Darwin and Facial Expression: A Century of Research in Review. New York, NY, USA: Academic; 1973. [Google Scholar]
- 40.James W. What is an emotion? Mind. 1884;9:188–205. [Google Scholar]
- 41.Ellsworth PC. William James and emotion: is a century of fame worth a century of misunderstanding? Psychological Review. 1994;101(2):222–229. doi: 10.1037/0033-295x.101.2.222. [DOI] [PubMed] [Google Scholar]
- 42.Lange C. The emotions. In: Dunlap E, editor. The Emotions. Baltimore, Md, USA: Williams & Wilkins; 1885. pp. 33–92. [Google Scholar]
- 43.Peper M, Markowitsch HJ. Pioneers of affective neuroscience and early concepts of the emotional brain. Journal of the History of the Neurosciences. 2001;10(1):58–66. doi: 10.1076/jhin.10.1.58.5628. [DOI] [PubMed] [Google Scholar]
- 44.Panksepp J. Affective Neuroscience. Oxford, UK: University Press; 1998. [Google Scholar]
- 45.Macmillan M. The concept of inhibition in some nineteenth century theories of thinking. Brain and Cognition. 1996;30(1):4–19. doi: 10.1006/brcg.1996.0002. [DOI] [PubMed] [Google Scholar]
- 46.Bachrach AJ. Experimental Foundations of Clinical Psychology. New York, NY, USA: Basic Books; 1962. [Google Scholar]
- 47.Dror OE. Techniques of the brain and the paradox of emotions, 1880–1930. Science in Context. 2001;14(4):643–660. [PubMed] [Google Scholar]
- 48.Bulatao E, Cannon WB. The role of the adrenal medulla in pseudaffective hyperglycemia. American Journal of Physiology. 1925;72:295–313. [Google Scholar]
- 49.Cannon WB, Britton SW. Studies on the conditions of activity in endocrine glands. American Journal of Physiology. 1925;72:283–294. [Google Scholar]
- 50.Anon. Students measure fear by a pupilometer: kick subject’s shins to experiment on anger. The New York Times. 1925 Nov;:p. 27:4. [Google Scholar]
- 51.Blatz W. A Physiological Study of the Emotion of Fear. Chicago, Ill, USA: University of Chicago; 1924. [Google Scholar]
- 52.Brunswick D. The effect of emotional stimuli on the gastro-intestinal tone. I. Methods and technique. Journal of Comparative Psychology. 1924;4(1):19–79. [Google Scholar]
- 53.Lombard JS. Experimental Researches on the Regional Temperature of the Head under Conditions of Rest, Intellectual Activity, and Emotion. London, UK: H. K. Lewis; 1879. [Google Scholar]
- 54.Lund FH. Emotions: Their Psychological, Physiological and Educative Implications. New York, NY, USA: Ronald Press; 1939. [Google Scholar]
- 55.Cannon WB. The James-Lange theory of emotions: a critical examination and an alternative theory. By Walter B. Cannon, 1927. The American Journal of Psychology. 1987;100(3-4):567–586. [PubMed] [Google Scholar]
- 56.Bard PA. A diencephalic mechanism for the expression of rage with special reference to the central nervous system. American Journal of Physiology. 1928;84:490–513. [Google Scholar]
- 57.Bard PA, Rioch DM. A study of four cats deprived of neocortex and additional portions of the forebrain. John Hopkins Medical Journal. 1937;60:73–153. [Google Scholar]
- 58.Cannon WB. Again the James-Lange and the thalamic theories of emotion. Psychological Review. 1931;38(4):281–295. [Google Scholar]
- 59.Kluver H, Bucy PC. “Psychic blindness” and other symptoms following bilateral temporal lobectomy. American Journal of Physiology. 1937;119:254–284. [Google Scholar]
- 60.Hayman LA, Rexer JL, Pavol MA, Strite D, Meyers CA. Kluver-Bucy syndrome after bilateral selective damage of amygdala and its cortical connections. Journal of Neuropsychiatry and Clinical Neurosciences. 1998;10(3):354–358. doi: 10.1176/jnp.10.3.354. [DOI] [PubMed] [Google Scholar]
- 61.Nahm FK. Heinrich Klüver and the temporal lobe syndrome. Journal of the History of the Neurosciences. 1997;6(2):193–208. doi: 10.1080/09647049709525702. [DOI] [PubMed] [Google Scholar]
- 62.Lilly R, Cummings JL, Benson DF, Frankel M. The human Kluver-Bucy syndrome. Neurology. 1983;33(9):1141–1145. doi: 10.1212/wnl.33.9.1141. [DOI] [PubMed] [Google Scholar]
- 63.Guyton AC, Hall JE. Behavioral and motivational mechanisms of the brain—the limbic system and the hypothalamus. In: Guyton AC, Hall JE, editors. Textbook of Medical Physiology. Philadelphia, Pa, USA: W. B. Saunders; 2001. pp. 678–687. [Google Scholar]
- 64.Trimble MR, Mendez MF, Cummings JL. Neuropsychiatric symptoms from the temporolimbic lobes. Journal of Neuropsychiatry and Clinical Neurosciences. 1997;9(3):429–438. doi: 10.1176/jnp.9.3.429. [DOI] [PubMed] [Google Scholar]
- 65.Ghika-Schmid F, Assal G, De Tribolet N, Regli F. Kluver-Bucy syndrome after left anterior temporal resection. Neuropsychologia. 1995;33(1):101–113. doi: 10.1016/0028-3932(94)00097-9. [DOI] [PubMed] [Google Scholar]
- 66.Papez JW. A proposed mechanism of emotion. Archives of Neurology and Psychiatry. 1937;38:725–743. [Google Scholar]
- 67.Lopes da Silva FH, Witter MP, Boeijinga PH, Lohman AHM. Anatomic organization and physiology of the limbic cortex. Physiological Reviews. 1990;70(2):453–511. doi: 10.1152/physrev.1990.70.2.453. [DOI] [PubMed] [Google Scholar]
- 68.MacLean PD. Some psychiatric implications of physiological studies on frontotemporal portion of limbic system (Visceral brain) Electroencephalography and Clinical Neurophysiology. 1952;4(4):407–418. doi: 10.1016/0013-4694(52)90073-4. [DOI] [PubMed] [Google Scholar]
- 69.MacLean PD. Psychosomatic disease and the visceral brain; recent developments bearing on the Papez theory of emotion. Psychosomatic Medicine. 1949;11(6):338–353. doi: 10.1097/00006842-194911000-00003. [DOI] [PubMed] [Google Scholar]
- 70.Lambert KG. The life and career of Paul MacLean: a journey toward neurobiological and social harmony. Physiology & Behavior. 2003;79(3):343–349. doi: 10.1016/s0031-9384(03)00147-1. [DOI] [PubMed] [Google Scholar]
- 71.MacLean PD. The Triune Brain in Evolution: Role in Paleocerebral Functions. New York, NY, USA: Plenum; 1990. [DOI] [PubMed] [Google Scholar]
- 72.Lambert KG, Gerlai R. The neurobiological relevance of social behavior: Paul MacLean’s legacy. Physiology & Behavior. 2003;79(3):341–342. doi: 10.1016/s0031-9384(03)00146-x. [DOI] [PubMed] [Google Scholar]
- 73.Fulton JF. Somatic functions of the central nervous system. Annual Review of Physiology. 1953;15:305–328. doi: 10.1146/annurev.ph.15.030153.001513. [DOI] [PubMed] [Google Scholar]
- 74.MacLean PD, Pribram KH. A neuronographic analysis of the medial and basal cerebral cortex, comparing cat and monkey. Journal of Neurophysiology. 1952;16:324–340. doi: 10.1152/jn.1953.16.3.324. [DOI] [PubMed] [Google Scholar]
- 75.Pribram KH, Lennox MA, Dunsmore RH. Some connections of the orbito-fronto-temporal, limbic and hippocampal areas of Macaca mulatta. Journal of Neurophysiology. 1950;13:127–135. doi: 10.1152/jn.1950.13.5.383. [DOI] [PubMed] [Google Scholar]
- 76.Holden C. Paul MacLean and the triune brain. Science. 1979;204(4397):1066–1068. doi: 10.1126/science.377485. [DOI] [PubMed] [Google Scholar]
- 77.MacLean PD. Man and his animal brains. Modern Medicine. 1964:95–106. [Google Scholar]
- 78.Windholz G, Grimsley DL. Pavlov’s position on the effects of decortication on conditional reflexes. Integrative Physiological and Behavioral Science. 1992;27(2):170–181. doi: 10.1007/BF02698505. [DOI] [PubMed] [Google Scholar]
- 79.Huxley TH. On the brain of ateles paniscus. Proceedings of the Zoological Society of London. 1861;17:247–260. [Google Scholar]
- 80.Nauta WJH. Hippocampal projections and related neural pathways to the mid-brain in the cat. Brain. 1958;81(3):319–340. doi: 10.1093/brain/81.3.319. [DOI] [PubMed] [Google Scholar]
- 81.Nauta WJH, Feirtag M. Fundamental Neuroanatomy. New York, NY, USA: Freeman; 1986. [Google Scholar]
- 82.Swanson LW. The hippocampus and the concept of the limbic system. In: Seifert W, editor. Neurobiology of the Hippocampus. London, UK: Academic; 1983. pp. 3–20. [Google Scholar]
- 83.LeDoux JE. Emotion and the limbic system concept. Concepts in Neuroscience. 1991;2(2):169–199. [Google Scholar]
- 84.Olry R, Nicolay X. De Paul Broca à la potentiation à long terme: les aléas de l'affirmation d'une identité limbique. Historie des Sciences Medicales. 1994;28(3):199–203. [PubMed] [Google Scholar]