

Abstract
A recombinant Escherichia coli K-12 strain, transformed with a modified bacterial luciferase gene (luxABCDE) from Photorhabdus luminescens, was constructed in order to monitor the activity of various antimicrobial agents on a real-time basis. This E. coli-lux emitted, without any addition of substrate, constitutive bioluminescence (BL), which correlated to the number of viable bacterial cells. The decrease in BL signal correlated to the number of killed bacterial cells. Antimicrobial activity of hydrogen peroxide (H2O2) and myeloperoxidase (MPO) was assessed. In high concentrations, H2O2 alone had a bacteriocidic function and MPO enhanced this killing by forming hypochlorous acid (HOCl). Taurine, the known HOCl scavenger, blocked the killing by MPO. When E. coli-lux was incubated with neutrophils, similar killing kinetics was recorded as in H2O2/MPO experiments. The opsonization of bacteria enhanced the killing, and the maximum rate of the MPO release from lysosomes coincided with the onset of the killing.
Keywords: phagocytosis, neutrophil, myeloperoxidase, hydrogen, peroxide, hypochlorous, acid, bacterial, luciferase, bioluminescence
1. INTRODUCTION
Polymorphonuclear neutrophils are the most abundant type of white blood cells in mammals; they are the key effector cells in the host defense response to microbial invasion providing the rapid deployment and effector arm of the immune system [1–6]. Neutrophils are professional phagocytes capable for ingesting microbes and particles into intracellular compartments called phagolysosomes.
An arsenal of cytotoxic agents in phagolysosomes is directed to degrade the phagocytosed material [7–10]. The process of opsonization is a means of identifying the invading microbes by phagocytes. The nonspecific binding of the complement component C3b and C3bi and the specific binding of the serum antibodies to the invading pathogen are normally required for the successful recognition and destruction of this pathogen by phagocytic cell [11–16].
The ingestion of the target leads to enhanced consumption of oxygen through the activity of NADPH oxidase that generates superoxide anion (O2 −) which is further dismutated forming hydrogen peroxide (H2O2) [6–10]. This leads to the formation of further reactive oxidants such as hypochlorous acid (HOCl) catalyzed by myeloperoxidase (MPO) [6–9, 17]. MPO is a peroxidase enzyme having a fundamental role in the oxidant production and thus in antimicrobial activity of neutrophils. It is a lysosomal protein stored in azurophilic granules of the neutrophils [6–9]. MPO is released by fusing these granules with phagosome leading to the formation of the phagolysosome [18].
The assessment of the antimicrobial activities of neutrophils is generally made by flow cytometry (FC), chemiluminescence (CL) assay, or microscopy [13, 19–23]. Moreover, the detection of MPO is mainly based on the quantification of the enzyme protein by immunoassays without any knowledge of MPO activity. We have previously used luminol-amplified CL and FC to analyze the binding, ingestion and respiratory burst activities of phagocytes [24–28], and there is a strict correlation between the number of ingested particles and luminol-amplified CL response of neutrophils [23, 24], provided that the CL of adhesion events is excluded [23, 24]. Luminol-amplified CL has been shown to be almost completely dependent on the release of MPO from azurophilic granules.
These are reliable methods, but they do not reveal anything about the killing of the microbes. Conventionally, viability assessment is made by plate counting. The kinetic measurement of killing by this method is troublesome, and the results are not obtained on a real time basis, since the plates demand a long incubation period. Optical density (OD) measurement also provides a real-time assay, but the high cell density required for the turbidity measurements and an inability to distinguish between live and dead bacteria restrict the application of this method [29].
In this study, we describe an approach in which Escherichia coli K-12 pEGFPluxABCDEAmp (E. coli-lux) was utilized for the assessment of the killing by neutrophil-derived oxidants. We have previously shown that the bioluminescence (BL) signal of E. coli-lux was directly related to the number of viable bacterial cells and the diminishment of the signal, caused by the addition of antimicrobial agent, correlates to the number of killed E. coli-lux cells [29]. We can monitor this killing reaction quantitatively on a real-time basis by measuring the BL signal continuously during the incubation [29].
2. MATERIALS AND METHODS
2.1. Materials
Agar, tryptone, and yeast extract were obtained from Difco laboratories (Detroit, Mich). Disodium phosphate (Na2HPO4 · 2H2O) and monopotassium phosphate (KH2PO4) were purchased from J. T. Baker (Deventer, Holland). Ampicillin sodium salt, glycerol, H2O2, luminol, sodium chloride (NaCl), and taurine were obtained from Sigma-Aldrich (St. Louis, Mo, USA). MPO was purchased from Planta Natural Products (Vienna, Austria). All reagents were at least of analytical grade.
2.2. Bacterial Preparation and Cultivation
E. coli-lux was precultivated in 5 mL of Luria Bertani Broth (LB) (10 g tryptone, 5 g of yeast extract, 5 g NaCl, and pH 7.4) and incubated in a shaker (250 rpm) at 37°C overnight. Bacterial cultivation was then suspended in 100 mL LB medium and incubated in a shaker (250 rpm) at 37°C until OD620 nm was 0.25, measured with Shimadzu UV-1601 photometer (Shimadzu Corporation, Japan). At this OD, the cells were in logarithmic growth, and the cultures contained approximately 3.5 × 107 bacterial cells/mL. The cells were harvested by centrifugation at 2500 ×g, resuspended in the mixture of 20 mL of LB (containing 25% glycerol) for freezer stock to be stored at −70°C. All E. coli media contained 100 μg/mL ampicillin in order to maintain the selection pressure.
Before killing experiments, E. coli-lux was first cultivated by adding 50 μL of freezer stock to 5 mL LB medium and then incubated in a shaker (250 rpm) at 37°C until OD620 nm was 0.25. After incubation bacterial cells were harvested by centrifugation at 2500 ×g, washed two times, and then resuspended either in 67 mM phosphate buffer (pH 7.4) with 0.9% sodium chloride (NaCl) or Hanks Balanced Salt Solution (HBSS) with gelatin (1 mg/mL) (gHBSS) for phagocytosis analysis.
2.3. Serum Handling and Leukocyte Separation
Peripheral human blood was collected from a healthy volunteer into Venosafe 3 mL EDTA tube and one Venosafe 4 mL serum tube (Terumo Corporation, Leuven, Belgium). Serum was separated and used for the opsonization. 1 mL of EDTA blood was mixed with 10 mL of 0.83% NH2Cl including 370 mg/l EDTA in order to lyse the red blood cells (RB). The suspension was kept at room temperature for 15 min, after which leukocytes were separated by centrifugation for 10 min at 400 ×g. The lysed RB debris was sucked away, and leukocytes were resuspended in 1 mL of gHBSS. To avoid unnecessary stress to cells additional purification steps were refrained. Leukocytes were calculated with microscope by using Bürker counting chamber, and isolated leukocyte suspension contained approximately 2.5 × 106 neutrophils/mL, representing 85% of the phagocyte population (monocytes 12% and eosinophils 3%). We have previously shown that the luminol-amplified CL response of monocytes and eosinophils is only 50% of that of neutrophils and, thus, practically, the entire CL-signal per blood volume originates from neutrophils [25].
2.4. Antimicrobial Reaction of H2O2 and MPO
Killing assays were performed by adding 140 μL of cultivated E. coli-lux bacteria suspended in phosphate buffer (pH 7.4 with 0.9% NaCl), into the wells of a white clear bottom 96-well microtiter plate (Greiner Bio-One, Düsseldorf, Germany), containing 3 × 105 or 3 × 106 bacterial cells/well. MPO (1 or 2 μg/well), H2O2 (30 μM, 125 μM, 500 μM, 2 mM, or 20 mM/well) and taurine (50 mM/well) were all added separately in 20 μL of phosphate buffer. The final reaction volume was 200 μL/well, and the reaction was started by adding H2O2.
2.5. Killing Activity of Neutrophils
Neutrophils-killing activity assay was performed by pipetting 100 μL of cultivated bacteria suspended in gHBSS, containing 1 × 106 bacterial cells/well. Neutrophils were added by pipetting 2.5 × 105 neutrophils in 100 μL of gHBSS into the wells. Opsonization was performed by adding 40 μL of 2.5% serum solution in gHBSS to the reaction mixture, final serum concentration being 0.4%. The final reaction volume was 250 μL/well.
Luminol-amplified CL assay was made by adding 100 μL of bacterial suspension (1 × 106 E. coli-lux cells/well) in gHBSS, 100 μL of leukocyte suspension (2.5 × 105 neutrophils/well) in gHBSS, 20 μL of luminol (10 mM in borate buffer, pH 9.0), and 20 μL of human serum solution in gHBSS to the microtiter plate wells (final serum concentration 0.4%). The final reaction volume was 250 μL/well.
2.6. Multimode Readers
BL and OD (620 nm) of E. coli-lux was measured in microtiter plate wells at 37°C in Mithras LB 940 microplate reader (Berthold technologies, Bad Wildbad, Germany). Luminol-amplified CL signal was measured in Chameleon microplate reader (Hidex, Turku, Finland), both controlled by MicroWin 2000 (Mikrotek). Three parallel wells were prepared from every reaction mixture. Readers were programmed to measure luminescence and OD continuously 0.5 sec/well, at 60 or 240 sec interval. Background signal was measured from a well containing only the buffer, and this reading was subtracted from the readings of the experimental wells. Results are shown as the mean ± SD of three parallel wells.
2.7. Colony Forming Unit (CFU) Measurement
A duplicate microplate, similar to that placed into the multimode reader in H2O2/MPO and neutrophil analysis, was placed into a laminar hood and incubated at 37°C. At 30 min intervals, 100 μL of every reaction suspension was diluted 101–107-fold, and 100 μL and 20 μL from every dilution were pipetted into Petri dishes containing LA agar with 100 μg/mL ampicillin. Colonies were counted after overnight incubation at 37°C. Results are shown as CFU/reaction volume.
2.8. Data Handling
Raw data was analyzed in Excel version 2007 (Microsoft), and all graphs were prepared with Origin version 8 (Microcal).
3. RESULTS
3.1. H2O2 and MPO Activity
H2O2 and MPO were added into the E. coli-lux suspension (3 × 105 cells/well), and BL and CFU were recorded (Figure 1). H2O2 alone, in the absence of MPO, had a capacity of killing, and more than 99% of bacterial cells were killed by 500 μM H2O2 after 90 min of incubation (Figures 1(a) and 1(b)). Addition of MPO enhanced the killing substantially, since the addition of 1 μg of MPO into 500 μM H2O2 killed more than 99% of bacterial cells already after one minute of reaction (Figures 1(a) and 1(b)). This enhancement was due to the HOCl generated by MPO reaction [7–10]. This was confirmed by adding 50 mM taurine, the known HOCl scavenger [22, 30], which blocked the killing by MPO reaction nearly totally (Figure 1(a)). That the BL measurement and plate counting were in accord with each other is shown in Table 1.
Figure 1.
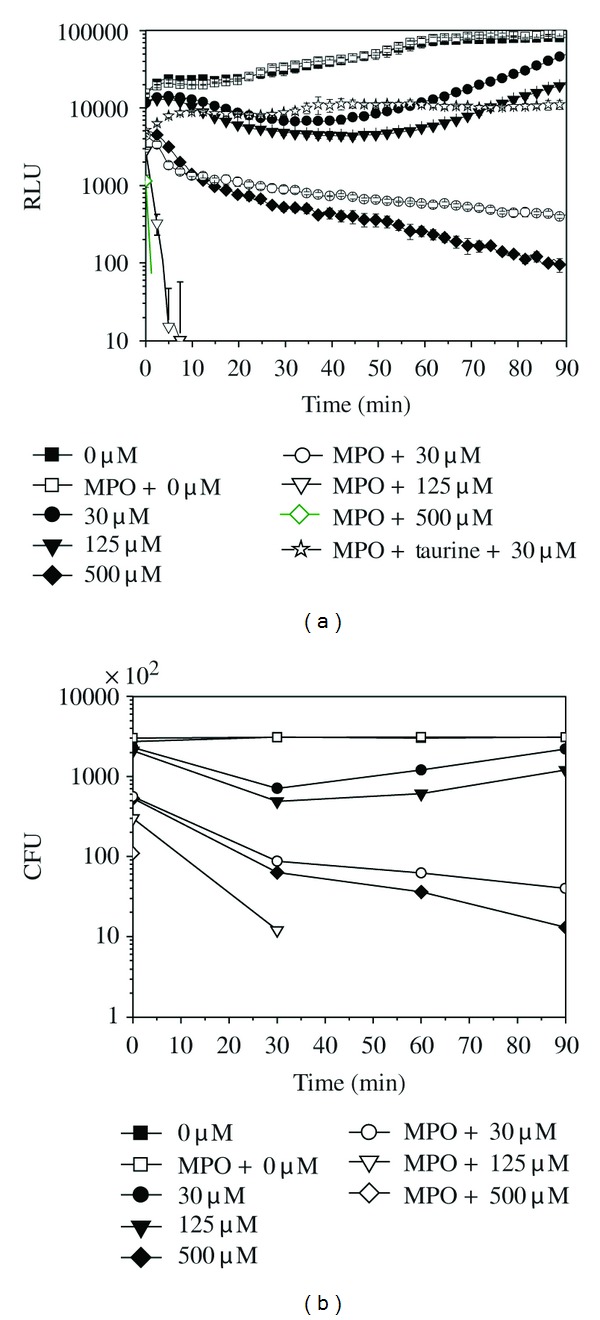
The bioluminescence signal (a) and colony forming units (CFU)/200 μL (b) of E. coli-lux (3 × 105 cells) incubated in the presence of various amounts of H2O2 (μM) and 1 μg/well MPO in phosphate buffer at 37°C. (■) 0 μM, (●) 30 μM, (▾) 125 μM, (♦) 500 μM, (□) MPO + 0 μM, (○) MPO + 30 μM, (∇) MPO + 125 μM, (♢) MPO + 500 μM, and (□) MPO + 50 mM of taurine + 30 μM. Relative luminescence unit (RLU) values are shown as the mean ± SD of measurements from three parallel wells.
Table 1.
The killing capacity of the H2O2/MPO system. The killing capacity of the H2O2/MPO system, both from the RLU and CFU analysis, was calculated by setting the values from the bacterial well as a 0% of killing.
| The killing capacity of the H2O2/MPO system | Killing (%) | |||||||
|---|---|---|---|---|---|---|---|---|
| 0 min | 30 min | 60 min | 90 min | |||||
| RLU | CFU | RLU | CFU | RLU | CFU | RLU | CFU | |
| MPO 1 μg/well | 0 | 0 | 0 | 1.3 | 0 | 0 | 0 | 0.6 |
| H2O2 30 μM | 29.4 | 15.8 | 81.1 | 77.2 | 82.9 | 60.3 | 46.0 | 29.3 |
| H2O2 125 μM | 29.4 | 23.1 | 86.5 | 84.2 | 91.4 | 79.8 | 78.0 | 61.4 |
| H2O2 500 μM | 69.1 | 80.6 | 98.6 | 98.0 | 99.5 | 98.8 | 99.9 | 99.6 |
| H2O2 30 μM + MPO 1 μg/well | 72.9 | 79.5 | 97.6 | 97.2 | 99.1 | 98.0 | 99.6 | 98.7 |
| H2O2 125 μM + MPO 1 μg/well | 82.4 | 89.0 | 100 | 99.6 | 100 | 100 | 100 | 100 |
| H2O2 500 μM + MPO 1 μg/well | 93.2 | 96.0 | 100 | 100 | 100 | 100 | 100 | 100 |
Bacteriolytic effect of the antimicrobial agent is detectable only by assessing the decrease in the OD signal [31–33]. The OD detection limit of the used plate reader, for the bacterial suspension, was approximately 1 × 106 cells/well corresponding to 0.030 OD in 200 μL, and therefore 10-fold higher bacterial cell number (3 × 106 cells/well) had to be used in killing experiments. The H2O2 concentrations used in previous experiments were not high enough to show any decrease in OD. Only 2 mM H2O2 in the presence of 2 μg of MPO and 20 mM H2O2 in the absence of MPO showed a decrease in OD, that is, bacteriolysis (Figure 2). In summary, the antimicrobial agents of neutrophils, H2O2, and HOCl showed significant killing capacity, which was easily measurable using E. coli-lux. These agents in high concentrations were even able to lyse the bacterial cells.
Figure 2.
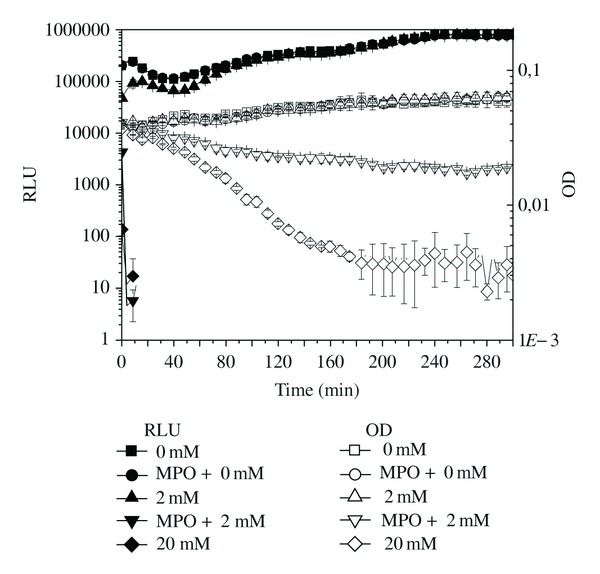
The bioluminescence signal and optical density (OD) of E. coli-lux (3 × 106 cells) incubated in the presence of various amounts of H2O2 (mM) and 2 μg/well MPO in phosphate buffer at 37°C. RLU: (■) 0 mM, (●) MPO + 0 mM, (▲) 2 mM, (▾) MPO + 2 mM, and (♦) 20 mM; OD620 nm: (□) 0 mM, (○) MPO + 0 mM, (∆) 2 mM, (∇) MPO + 2 mM, and (♢) 20 mM of H2O2. Relative luminescence unit (RLU) and OD values are shown as the mean ± SD of measurements from three parallel wells.
3.2. Killing Activity of Neutrophils
When neutrophils, instead of H2O2 and MPO, were added to the reaction, similar killing curves (Figure 3) were recorded as in previous experiments. It is noteworthy that neutrophils quenched partially BL since the initial relative luminescence unit (RLU) value of bacteria was reduced by 40% instantly after adding 2.5 × 105 neutrophils into the mixture. This was not killing since CFU values did not instantly decrease.
Figure 3.

The bioluminescence signal (a) and colony forming units (CFU)/250 μL (b) of E. coli-lux (1 × 106 cells) incubated in the presence of 2.5 × 105 neutrophils and 0.4% serum for opsonization in gHBSS medium at 37°C: (■) bacteria, (●) bacteria + serum, (▲) bacteria + neutrophils and (▾) bacteria + neutrophils + serum. Relative luminescence unit (RLU) values are shown as the mean ± S.D. of measurements from three parallel wells. Arrows point out the onset of the killing.
The actual killing of the microbes started approximately after 50–60 min of incubation when E. coli-lux was opsonized with 0.4% serum (OPS) and after 90–100 min in the absence of opsonins (NOPS) (Figures 3(a) and 3(b)).
Killing rate after 180 min of measurement was 41% for NOPS and 96% for OPS measured by RLU (Figure 3(a)) and 49% for NOPS and 92% for OPS measured by CFU (Figure 3(b)). Leukocytes were disrupted to find out whether the ingested bacteria were killed. Since there was no difference between the disrupted samples and normal samples in RLU and CFU (data not shown), it was concluded that all the ingested bacteria were killed.
Without neutrophils, the RLU signal was reduced by 4% in the presence of 0.4% serum (Figure 3(a)). The same reduction in CFU was 75% (Figure 3(b)). The discrepancy between these two values is thoroughly studied in another paper [29].
When luminol-amplified CL was recorded in the same reaction conditions as BL and CFU, the time to reach the maximum CL response (Figure 4) was the same as the time for the onset of killing (Figure 3).
Figure 4.
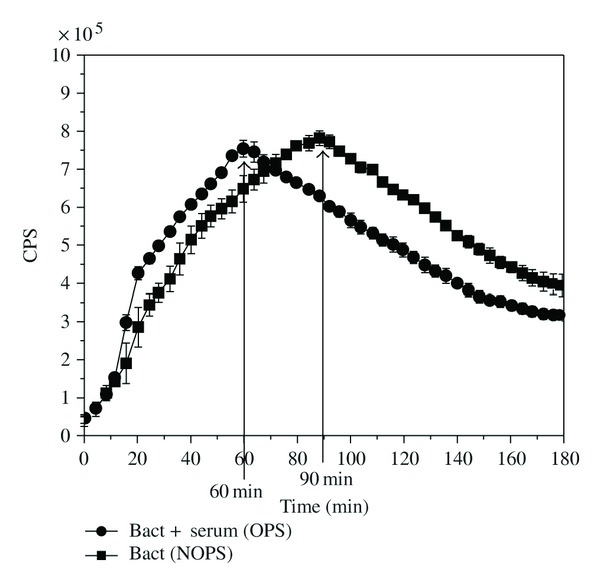
The luminol-amplified chemiluminescence (CL) signal of E. coli-lux (1 × 106 cells) with 2.5 × 105 neutrophils and 0.4% serum in gHBSS medium at 37°C. (●) bacteria + serum (OPS) and (■) bacteria (NOPS). Counts per second (CPS) values are shown as the mean ± SD of measurements from four parallel wells. Arrows point out the peak times of the CL signals.
4. DISCUSSION
E. coli-lux-based real-time BL measurement provides a new approach for the bacterial killing by neutrophils and also for the antimicrobial activity of MPO. The kinetics of the killing activity of the antimicrobial agents can be continuously recorded; hence, the conventional kinetic parameters such as Vmax and Km can be obtained. From the kinetic curves, even the absolute number of the killed bacterial cells can be derived. It is obvious that the mechanisms of various scavenger and inhibitor functions, like taurine function in this study, can be explored.
In the present work, RLU and CFU values were in general in good correlation. Two exceptions were noticed. First, there was an initial reduction of 40% in RLU after the addition of neutrophils. This reduction was not present in CFU measurements. We have noticed that the extent of this reduction is dependent on the number of neutrophils added, even if they were heat inactivated (data not shown). The conclusion is that neutrophils somehow quench the BL of E. coli-lux either by absorbing the emission or by binding the bacteria, thus altering the BL properties.
The second exception was the reduction of CFU values caused by 0.4% serum (Figure 3(b)). Complement compounds present in this low serum concentration were transferred with the E. coli-lux cells into the plates and continued their activity during the overnight incubation, causing a considerable reduction in the bacterial viability in the plates. We have described this phenomenon earlier [29].
The lysis of the target E. coli-lux cells in the presence of H2O2 and/or HOCl was observed as a reduction in OD signal (Figure 2). These agents caused a rapid loss in viability (RLU values in Figures 1(a) and 2 and CFU values in Figure 1(b)) followed by much slower gradual lysis of the E. coli-lux (Figure 2). We will further study what are the mechanisms lying behind H2O2- and HOCl-induced lysis.
When the luminol-amplified CL assay (MPO release) was compared with viability assays, the maximum rate of the MPO release (the peak CL) coincided with the onset of the killing, suggesting that a certain threshold concentration of HOCl must be achieved before killing starts.
The results of this study suggest that the viability of E. coli-lux can be assessed on a real-time basis quickly and efficiently without tedious and time-consuming plate counting.
References
- 1.Anderson KL, Smith KA, Pio F, Torbett BE, Maki RA. Neutrophils deficient in PU.1 do not terminally differentiate or become functionally competent. Blood. 1998;92(5):1576–1585. [PubMed] [Google Scholar]
- 2.Lieber JG, Webb S, Suratt BT, et al. The in vitro production and characterization of neutrophils from embryonic stem cells. Blood. 2004;103(3):852–859. doi: 10.1182/blood-2003-04-1030. [DOI] [PubMed] [Google Scholar]
- 3.Hayashi F, Means TK, Luster AD. Toll-like receptors stimulate human neutrophil function. Blood. 2003;102(7):2660–2669. doi: 10.1182/blood-2003-04-1078. [DOI] [PubMed] [Google Scholar]
- 4.Cassatella MA, Locati M, Mantovani A. Never underestimate the power of a neutrophil. Immunity. 2009;31(5):698–700. doi: 10.1016/j.immuni.2009.10.003. [DOI] [PubMed] [Google Scholar]
- 5.Dale DC, Boxer L, Liles WC. The phagocytes: neutrophils and monocytes. Blood. 2008;112(4):935–945. doi: 10.1182/blood-2007-12-077917. [DOI] [PubMed] [Google Scholar]
- 6.Klebanoff SJ. Myeloperoxidase: friend and foe. Journal of Leukocyte Biology. 2005;77(5):598–625. doi: 10.1189/jlb.1204697. [DOI] [PubMed] [Google Scholar]
- 7.Klebanoff SJ. Myeloperoxidase-halide-hydrogen peroxide antibacterial system. Journal of Bacteriology. 1968;95(6):2131–2138. doi: 10.1128/jb.95.6.2131-2138.1968. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Kurutas EB, Arican O, Sasmaz S. Superoxide dismutase and myeloperoxidase activities in polymorphonuclear leukocytes in acne vulgaris. Acta Dermatovenerologica Alpina, Pannonica et Adriatica. 2005;14(2):39–42. [PubMed] [Google Scholar]
- 9.Mullane KM, Kraemer R, Smith B. Myeloperoxidase activity as a quantitative assessment of neutrophil infiltration into ischemic myocardium. Journal of Pharmacological Methods. 1985;14(3):157–167. doi: 10.1016/0160-5402(85)90029-4. [DOI] [PubMed] [Google Scholar]
- 10.Kumar P, Pai K, Pandey HP, Sundar S. NADH-oxidase, NADPH-oxidase and myeloperoxidase activity of visceral leishmaniasis patients. Journal of Medical Microbiology. 2002;51(10):832–836. doi: 10.1099/0022-1317-51-10-832. [DOI] [PubMed] [Google Scholar]
- 11.Nuutila J, Hohenthal U, Laitinen I, et al. Quantitative analysis of complement receptors, CR1 (CD35) and CR3 (CD11b), on neutrophils improves distinction between bacterial and viral infections in febrile patients: comparison with standard clinical laboratory data. Journal of Immunological Methods. 2006;315(1-2):191–201. doi: 10.1016/j.jim.2006.07.021. [DOI] [PubMed] [Google Scholar]
- 12.Nuutila J, Jalava-Karvinen P, Hohenthal U, et al. CRP/CD11b ratio: a novel parameter for detecting gram-positive sepsis. Human Immunology. 2009;70(4):237–243. doi: 10.1016/j.humimm.2009.01.009. [DOI] [PubMed] [Google Scholar]
- 13.Nuutila J, Lilius EM. Flow cytometric quantitative determination of ingestion by phagocytes needs the distinguishing of overlapping populations of binding and ingesting cells. Cytometry Part A. 2005;65(2):93–102. doi: 10.1002/cyto.a.20139. [DOI] [PubMed] [Google Scholar]
- 14.Carpentier JL, Lew DP, Paccaud JP, et al. Internalization pathway of C3b receptors in human neutrophils and its transmodulation by chemoattractant receptors stimulation. Cell Regulation. 1991;2(1):41–55. doi: 10.1091/mbc.2.1.41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Kemp AS, Turner MW. The role of opsonins in vacuolar sealing and the ingestion of zymosan by human neutrophils. Immunology. 1986;59(1):69–74. [PMC free article] [PubMed] [Google Scholar]
- 16.Gaither TA, Hammer CH, Gadek JE. Cleavage of membrane-bound C3b and C3bi by viable human neutrophils (PMN) Molecular Immunology. 1983;20(6):623–635. doi: 10.1016/0161-5890(83)90007-x. [DOI] [PubMed] [Google Scholar]
- 17.Asselbergs FW, Tervaert JWC, Tio RA, et al. Prognostic value of myeloperoxidase in patients with chest pain. The New England Journal of Medicine. 2004;350(5):516–518. [PubMed] [Google Scholar]
- 18.Cougoule C, Constant P, Etienne G, Daffé M, Maridonneau-Parini I. Lack of fusion of azurophil granules with phagosomes during phagocytosis of Mycobacterium smegmatis by human neutrophils is not actively controlled by the bacterium. Infection and Immunity. 2002;70(3):1591–1598. doi: 10.1128/IAI.70.3.1591-1598.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Nuutila J, Jalava-Karvinen P, Hohenthal U, et al. Comparison of degranulation of easily mobilizable intracellular granules by human phagocytes in healthy subjects and patients with infectious diseases. Human Immunology. 2009;70(10):813–819. doi: 10.1016/j.humimm.2009.06.017. [DOI] [PubMed] [Google Scholar]
- 20.Ihalin R, Nuutila J, Loimaranta V, Lenander M, Tenovuo J, Lilius EM. Susceptibility of Fusobacterium nucleatum to killing by peroxidase-iodide-hydrogen peroxide combination in buffer solution and in human whole saliva. Anaerobe. 2003;9(1):23–30. doi: 10.1016/S1075-9964(03)00005-2. [DOI] [PubMed] [Google Scholar]
- 21.Cassatella MA, Mosna F, Micheletti A, et al. Toll-like receptor-3-activated human mesenchymal stromal cells significantly prolong the survival and function of neutrophils. Stem Cells. 2011;29(6):1001–1011. doi: 10.1002/stem.651. [DOI] [PubMed] [Google Scholar]
- 22.Selloum L, Djelili H, Sebihi L, Arnhold J. Scavenger effect of flavonols on HOCl-induced luminol chemiluminescence. Luminescence. 2004;19(4):199–204. doi: 10.1002/bio.772. [DOI] [PubMed] [Google Scholar]
- 23.Loimaranta V, Nuutila J, Marnila P, Tenovuo J, Korhonen H, Lilius EM. Colostral proteins from cows immunised with Streptococcus mutans/S. sobrinus support the phagocytosis and killing of mutans streptococci by human leucocytes. Journal of Medical Microbiology. 1999;48(10):917–926. doi: 10.1099/00222615-48-10-917. [DOI] [PubMed] [Google Scholar]
- 24.Nikoskelainen S, Verho S, Airas K, Lilius EM. Adhesion and ingestion activities of fish phagocytes induced by bacterium Aeromonas salmonicida can be distinguished and directly measured from highly diluted whole blood of fish. Developmental and Comparative Immunology. 2005;29(6):525–537. doi: 10.1016/j.dci.2004.10.006. [DOI] [PubMed] [Google Scholar]
- 25.Lilius EME, Nuutila JTJ. Particle-induced myeloperoxidase release in serially diluted whole blood quantifies the number and the phagocytic activity of blood neutrophils and opsonization capacity of plasma. Luminescence. 2006;21(3):148–158. doi: 10.1002/bio.899. [DOI] [PubMed] [Google Scholar]
- 26.Lojek A, Ciz M, Marnila P, Duskova M, Lilius EM. Measurement of whole blood phagocyte chemiluminescence in the Wistar rat. Journal of Bioluminescence and Chemiluminescence. 1997;12(5):225–231. doi: 10.1002/(SICI)1099-1271(199709/10)12:5<225::AID-BIO448>3.0.CO;2-X. [DOI] [PubMed] [Google Scholar]
- 27.Roman RM, Camargo PV, Borges FK, Rossini AP, Polanczyk CA. Prognostic value of myeloperoxidase in coronary artery disease: comparison of unstable and stable angina patients. Coronary Artery Disease. 2010;21(3):129–136. doi: 10.1097/MCA.0b013e328333f50d. [DOI] [PubMed] [Google Scholar]
- 28.Cavusoglu E, Ruwende C, Eng C, et al. Usefulness of baseline plasma myeloperoxidase levels as an independent predictor of myocardial infarction at two years in patients presenting with acute coronary syndrome. American Journal of Cardiology. 2007;99(10):1364–1368. doi: 10.1016/j.amjcard.2006.12.060. [DOI] [PubMed] [Google Scholar]
- 29.Atosuo J, Lehtinen J, Vojtek L, Lilius E-M. Escherichia coli K-12 (pEGFPluxABCDEamp). A tool for analysis of bacterial killing by antibacterial agents and human complement activities on a real-time basis. doi: 10.1002/bio.2435. submitted to International Journal of Antimicrobial Agents. [DOI] [PubMed] [Google Scholar]
- 30.Kearns S, Dawson R., Jr. Cytoprotective effect of taurine against hypochlorous acid toxicity to PC12 cells. Advances in Experimental Medicine and Biology. 2000;483:563–570. doi: 10.1007/0-306-46838-7_60. [DOI] [PubMed] [Google Scholar]
- 31.Rozenberg-Arska M, Porsius JC, Jaarsma EY, Verhoef J. Bactericidal, bacteriolytic and opsonic activity of human serum against Escherichia coli. Journal of Medical Microbiology. 1986;22(2):143–149. doi: 10.1099/00222615-22-2-143. [DOI] [PubMed] [Google Scholar]
- 32.Taylor PW. Sensitivity of some smooth strains of Escherichia coli to the bactericidal action of normal human serum. Journal of Clinical Pathology. 1974;27(8):626–629. doi: 10.1136/jcp.27.8.626. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Taylor PW. Bactericidal and bacteriolytic activity of serum against gram-negative bacteria. Microbiological Reviews. 1983;47(1):46–83. doi: 10.1128/mr.47.1.46-83.1983. [DOI] [PMC free article] [PubMed] [Google Scholar]