

Abstract
The major cell wall constituent of Ganoderma lucidum (G. lucidum) is β-1,3-glucan. This study examined the polysaccharide from the residues of alkaline-extracted fruiting bodies using high-performance anion-exchange chromatography (HPAEC), and it employed nuclear magnetic resonance (NMR) and mass spectrometry (MS) to confirm the structures. We have successfully isolated low-molecular-weight β-1,3-glucan (LMG), in high yields, from the waste residue of extracted fruiting bodies of G. lucidum. The 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) assay evaluated the capability of LMG to suppress H2O2-induced cell death in RAW264.7 cells, identifying that LMG protected cells from H2O2-induced damage. LMG treatment decreased H2O2-induced intracellular reactive oxygen species (ROS) production. LMG also influenced sphingomyelinase (SMase) activity, stimulated by cell death to induce ceramide formation, and then increase cell ROS production. Estimation of the activities of neutral and acid SMases in vitro showed that LMG suppressed the activities of both neutral and acid SMases in a concentration-dependent manner. These results suggest that LMG, a water-soluble β-1,3-glucan recycled from extracted residue of G. lucidum, possesses antioxidant capability against H2O2-induced cell death by attenuating intracellular ROS and inhibiting SMase activity.
1. Introduction
Beta-1,3-D-glucans are constituents of fungi, algae, and higher plants. These glucans exhibit different solubility in water-based solutions [1, 2]. G. lucidum is the most popular medicinal mushroom, with immunomodulating activity, applied in the treatment of a variety of diseases, including cancer, in numerous Asian countries [3–5]. Previous research has isolated several major substances with potent bioactivities, such as polysaccharides (in particular, β-1,3-glucan) or protein-conjugated β-1,3-glucan [1, 6–8]. Polysaccharides from G. lucidum are not linear in structure, comprising β-(1→6)-D-glucosyl-linked side chains on a β-1,3-D-glucan backbone. Numerous studies have described their functions [9–12].
Beta-1,3-D-glucans in G. lucidum have high molecular weights and low solubility in neutral water solutions. This creates the need to degrade glucan into smaller sizes for further application. Polysaccharide extraction from G. lucidum produces a large amount of residue as waste material. Therefore, the development of a “green” method of production, which recycles used materials and low volumes of waste, is of high importance. The present study's methods for isolating β-1,3-D-glucan are efficient and low cost when compared with other extraction methods, providing high-quality products from the residue of G. lucidum, which may be useful in commercial applications.
Oxidative stress is implicated in the production of superoxide radicals in biological systems. ROS released from phagocytic cells initiates a wide range of toxic oxidative reactions and excessive formation of ROS causes cell apoptosis. Administration of antioxidants, such as medicinal herbs, can protect cells by attenuating injury caused by oxidative stress [13]. Almost all cell types express sphingomyelinase (SMase). Intracellular ceramide production increases the hydrolytic activity of SMases [14]. Ceramide is an intracellular lipid mediator that responds to various stimuli including stress. The inflammatory oxidant, H2O2, induces rapid increases in ceramide levels because of hydrolysis of sphingomyelin (SM) in plasma membranes, which contributes to the induction of various types of cell death. This study demonstrates that β-1,3-glucan activity is capable of interfering with SMases, and identifies β-1,3-glucan deriving from G. lucidum as a potential antioxidant source.
This study expanded the application of G. lucidum, reusing the waste materials from the extracted residue of fruiting bodies to obtain high-purity β-1,3-glucan in high yields. We also explored the biofunctions of low-molecular-weight-G. lucidum β-1,3-glucan; an efficient antioxidative agent that eliminates ROS in H2O2-treated cells by interfering with SMases. Results indicated that low-molecular-weight β-1,3-glucan has potential future use as a biomedicine. The technology used during polysaccharide degradation might also have potential application on other medicinal herbs to release various glycans for biological assay.
2. Materials and Methods
2.1. Recycling and Isolation of β-1,3-Glucans from Extracted Residue of G. lucidum
The dried fruiting bodies of G. lucidum were ground and extracted using dilute NaOH solution at 50°C for 12 h. The extract was centrifuged to provide water-insoluble part (waste residue, 80 to 89%) and water-soluble part (soluble heteroglycan extracts, 11 to 20%) [15]. The waste residue was further degraded by 2 N HCl at 50°C for 4 h to provide hydrolysate (45%) and insoluble residue (55%). This glucan-containing hydrolysate underwent purification by gel-filtration chromatography (Sephadex G-15) to obtain three fractions; named high-molecular-weight-glucan (HMG) in 18%, low-molecular-weight-glucan (LMG) in 60% and oligosaccharide of glucose (OSG) in 22% yield, respectively. Figure 1 displays the chromatographic diagram of recycled β-1,3-glucan from the waste residue of G. lucidum.
Figure 1.
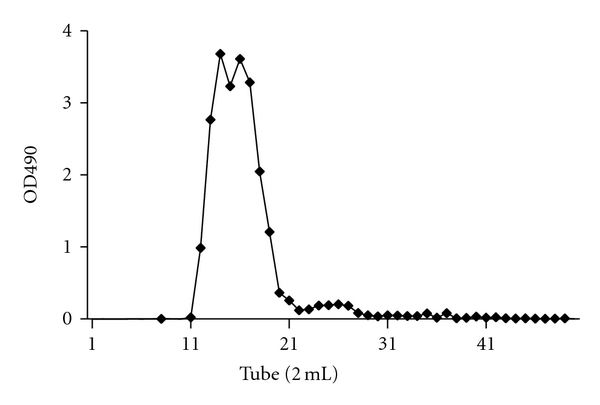
Size exclusion chromatography diagram of recycled β-1,3-glucan from the waste residue of G. lucidum. Samples were isolated using a Sephadex G-15 column (HMG: 11–13; LMG: 14–20; OSG: 23–29 tubes), and the sugar content of each tube was determined using phenol-sulfuric acid analysis (OD490 nm).
2.2. Instruments in Structural Determination of β-1,3-Glucan
High-performance anion-exchange chromatography (HPAEC) was used for the composition analysis of β-1,3-glucans. Glucose released by the acidic hydrolysis of LMG (4 N TFA 113°C 6 h) and characterized using HPAEC-PAD with known monosaccharides as standard, a Dionex BioLC system (Dionex, Sunnyvale, CA, USA) with a CarboPac PA10 analytical column (4.5 mm × 250 mm) and a CarboPac PA10 guard column (4 mm × 50 mm) was used. Gas chromatography-mass spectrometer (GC-MS) was used for analysis of saccharide composition and linkages according to the methods described by Hakomori et al. [16]. The alditol acetates and partially methylated alditol acetates were separated by GC-MS using a HP1 capillary column (dimethyl siloxane, 30 cm × 200 mm from Hewlett Packard) and a BPX-70 column (SGE Analytical Science Pty Ltd., Victoria 3134, Australia) on PolarisQ Ion Trap GC-MS/MS System (Thermo Fisher Scientific, Inc., Waltham, MA 024253, USA). The oven temperature was increased from 38°C to 150°C at 50°C per min, then to 230°C at 3°C per min and to 260°C for 5 min, with carrier gas (He) at a flow rate of 1 mL/min. NMR studies, 1H/13C NMR, and other 1D and 2D experiments were performed on a Bruker Fourier transform spectrometer (AV-600) equipped with a 5 mm DCI dual cryoprobe. Spectra were obtained at 298 K with solutions of polysaccharide in D2O. The HOD signal was achieved by a presaturation for 0.5 s. NMR diffusion ordered spectroscopy (DOSY) experiments were carried out using the standard pulse sequence on the same AV-600 NB spectrometer for determination of molecular weight. The equation for DOSY experiments was as follows [17];
| (1) |
Mass spectrometry was used for analyses of molecular weight and linkages. The molecular weights of the sodium adducts of oligosaccharides [M + Na]+ were determined using MALDI-TOF MS (Applied Biosystems, Foster City, CA. USA). The glycan was mixed with 10 mg/mL of 2, 5-dihydroxybenzoic acid (2,5-DHB) and 10 mM NaCl in the ratio of 5 : 5 : 3. In MS mode, the spectra were accumulated at an average of 1,000 to 2,000 shuts at a sample concentration of 1 mg/mL.
2.3. Cell Culture
The mouse monocyte-macrophage cell line, RAW 264.7, was cultured in DMEM supplemented with 10% FBS, 100 U/mL penicillin, and 100 mg/mL streptomycin. Conditions were maintained in a humidified incubator (95% air with 5% CO2) at 37°C.
2.4. Cell Viability Assay
MTT assay was conducted to assay cytotoxicity and cell viability, as described previously [18], dependent on the conversion of yellow tetrazolium salt to purple formazan product. Cells (1 × 105 cells/mL) were grown on a 96-well plate. Cells were treated with LMG (0–200 μg/mL) or LMG (0–150 μg/mL) combined with 200 μM H2O2 for 24 h, after which an MTT solution (1 mg/mL in PBS) was added, and cells were incubated for 2 h. After the supernatant was discarded and DMSO was added to the well, the absorbance at 570 nm was measured using a spectrophotometer (Thermo Varioskan Flash, Vantaa, Finland).
2.5. Detection of Intracellular ROS Formation
Levels of cellular oxidative stress were detected using the fluorescent probe, DCFDA, as described previously [19]. After being treated with 20 μM H2O2 in the absence or presence of LMG (100 μg/mL), RAW264.7 cells were treated with 20 μM DCFDA for 30 min and then washed in PBS. DCFDA is mainly trapped in the cytoplasm and is oxidized to the highly fluorescent dichlorofluorescein (DCF) by intracellular ROS. DCF fluorescence intensities in cells were observed using fluorescence microscopy (Olympus IX71, Tokyo, Japan).
2.6. Suppression of Neutral and Acid SMase Activities by LMG
Neutral SMase (nSMase) activity was estimated using an Amplex Red SMase assay kit (Invitrogen), as described previously [20]. Briefly, the assay mixture contained the following components in a total volume of 200 μL. The enzyme source (0.04 U/mL SMase, 1 U/mL HRP, 0.2 U/mL choline oxidase, 8 U/mL alkaline phosphatase, and 500 μM SM at pH 7.4) in the absence or presence of LMG (0–200 μg/mL) was prepared. For the nSMase assay, assay mixtures were incubated at 37°C for 30 min. Assay of acid SMase (aSMase) activity was performed at pH 5.0. The reaction mixture contained 0.4 U/mL SMase, LMG (0–200 μg/mL), and 500 μM SM in 100 μL of 50 mM sodium acetate, pH 5.0, and cell samples were incubated at 37°C for 30 min. A 100 μL working solution (consisting of 100 μM Amplex Red, 1 U/mL HRP, 0.2 U/mL choline oxidase, and 8 U/mL alkaline phosphatase in 100 mM Tris-HCl, pH 8.0) was added, and cells were incubated at 37°C for 30 min. Fluorescence was measured using a microplate reader (Thermo Varioskan Flash) with excitation at 545 nm and emission at 590 nm.
3. Results and Discussion
3.1. Isolation of β-1,3-Glucans from Extracted Residue of G. Lucidum
This study's aim was to obtain LMG from the extracted residue of G. lucidum and to examine its antioxidative activities. Alkaline extracted the ground fruiting bodies of G. lucidum to release a water-soluble component (20%) and a water-insoluble residue (80%). The water-soluble component contained heteroglycan, glycoprotein and others, and earlier studies have reported its immunomodulating function [3, 6, 10]. The recycling of β-1,3-glucan from the residue of G. lucidum was of interest due to the abundance of β-1,3-glucan in the cell walls of fungi and wide application in medicinal food [1, 2, 4]. Hydrolysis of the waste material, a water-insoluble residue, by strong acidic degradation (2 M HCl, 50°C, 4 h) obtained LMG as low molecular weight glucans. This process provided smaller sized beta-glucans by narrowing down the high-molecular-weight-polyglycans, with the hydrolyzed glucans becoming water soluble. Further purification of the hydrolysate using size-exclusion chromatography (Sephadex G-15) provided three fractions; named HMG, LMG, and OSG. The isolated process is shown in Figure 1. The LMG is also obtained from the hydrolysate by ethanol precipitation (EtOH/H2O = 5/1) or by ultrafiltration (UF) with a molecular cutoff 500 Da to remove low-molecular-weight-components for large-scale isolation. Structural analyses and biological assays used the prepared LMG following collection and lyophilization.
3.2. Structural Analysis of LMG
The isolated β-1,3-glucan from the residue of extracted G. lucidum fruiting bodies was a white powder and relatively pure, despite containing compounds of various sizes due to chemical degradation producing a variety of molecular weights. For composition analysis, acidic hydrolysis (4 N TFA, 110°C, 4 h) further degraded LMG and determined the monosaccharide construct in HPAEC. Procedures identified the only glucose present in the LMG fraction, suggesting that LMG is a glucan from the cell wall of G. lucidum. Linkages analysis used GC-MS according to the methods described by Hakomori et al. [16]. Results showed that LMG contains a high ratio of 2,4,6-trimethyl acetyl glucitol (1→3 linked glucose) and trace amounts of 2,3,4,6-tetramethyl acetyl glucitol (terminal glucose) and 2,4-dimethyl acetyl glucitol (1→6 linked glucose). NMR (Figure 2), at 298 K, elucidated the structure of LMG is β-1,3-glucan. The 1H NMR spectrum of LMG protons H1–H6 appeared at 4.82, 3.59, 3.82, 3.55, 3.50, 3.976a, and 3.806b ppms, and its 13C NMR spectrum appeared at 103.0 (C-1), 73.2 (C-2), 85.2 (C-3), 68.4 (C-4), 76.3 (C-5), and 60.9 (C-6) ppms, respectively. These signals are in agreement with results from previous analyses [11, 21]. As shown in Figure 2, the tiny signals of β-1,6-glucan appeared at 102.2 (C-1), 73.9 (C-2), 76.5 (C-3), 70.4 (C-4), 76.8 (C-5), and 69.7 (C-6) ppms and the protons H1–H6 at 4.70, 3.40, 4.15, 4.05, 3.90, 4.606a, and 3.786b ppms [22].
Figure 2.
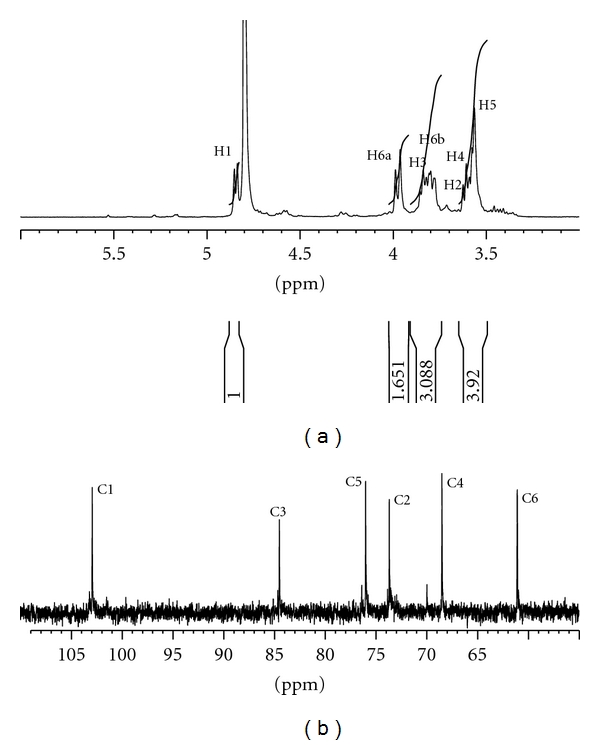
The 1H NMR spectrum (a) and 13C NMR spectrum (b) of LMG. These signals were in agreement with those of β-1,3-glucan from previously results [11].
3.3. Molecular Weights of LMG as Determined Using NMR and MALDI-TOF MS
Recently, investigators developed NMR DOSY as a useful tool for measurement of molecular weights, including those of polysaccharides [17, 23]. Using the equation in the experimental section, the estimated molecular weight of LMG is 3979 Da. The signal at 4.8 ppm, log D = −9.0, is H2O with molecular weight = 18 (Figure 3). The signal of anomeric protons on LMG merged in the HOD at 4.8 ppm. The convergent dots in the same horizontal, at log D = −10.0, indicated that LMG has a narrow range in terms of size. Excepting the HOD signal, no dots appeared in the parallel line in the DOSY spectrum indicating that protons in LMG are connected, and LMG is a homogeneous glucan with differences in narrow range sizes.
Figure 3.
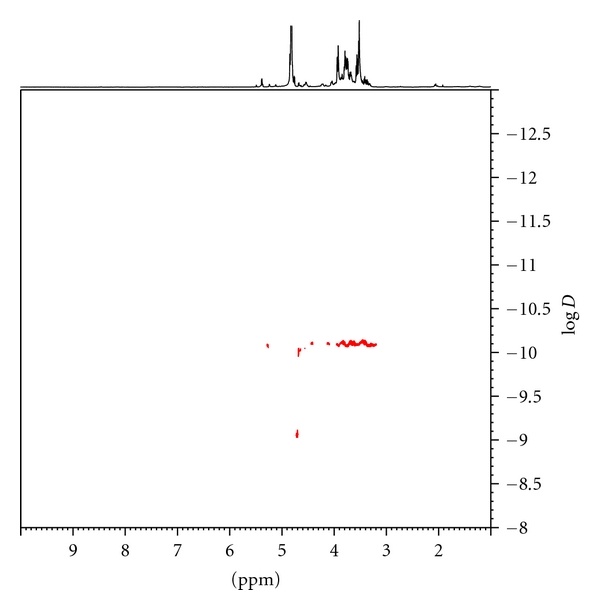
The 1D NMR DOSY experiment of LMG. Based on the equation, the calculated molecular weight of LMG is 3979 Da. The signal of anomeric protons merged in the HOD at 4.8 ppm and the signal, at log D = −9.0, is H2O with molecular weight = 18.
MALDI-TOF MS analysis confirmed the presence of β-1,3-glucan in LMG. Figure 4 shows the molecular mass of LMG with gaps as 162 Da (one hexose unit) between peaks to peaks. Use of 2,5-DHB as a matrix substantially enhanced polysaccharide signals in the MALDI mass spectrometry [24]. This study obtained positive ions of glucan with unambiguous signals, observing LMG as sodiated ions ([Glcn + Na]+) where n is the number of glucose units (6∼15). The mass region of ion peaks had a peak-to-peak mass difference of 162.1 Da, consistent with the repeating unit of the β-(1→3)-glucan. For example, when determining the degree of polymerization (DP) of LMG using MALDI-TOF, the average LMG mass appeared at DP = 9 (1499.1 Da, [Glc9 + Na]+) as a base peak. The molecular weights of LMG showed an average of 1,500 Da in MALDI-TOF MS, with slight differences from those obtained using the DOSY experiment. Based on these results, this study aimed to recover low molecular weight β-1,3-glucan from waste materials of G. lucidum residue. These recycled β-1,3-glucans might have potential commercial application as low-cost medicinal food.
Figure 4.
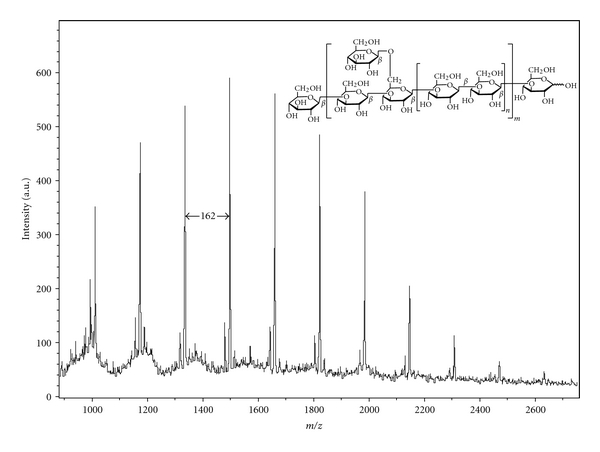
MALDI-TOF with 2,5-DHB as matrix, determined the molecular mass of LMG. The gap with 162 Da between peaks represents a hexose unit in LMG. Sodiated ions ([Glcn + Na]+) appeared at DP = 6 to 15 with 1013.1, 1175.1, 1337.1, 1499.1, 1661.2, 1823.2, 1985.2, 2147.2, 2309.2, and 2471.3 Da, respectively.
Although some β-D-glucans are commercially available, they have different molecular weights, biological activities, and structures. Previous studies have identified the biological activities of β-1,3-glucan containing G. lucidum [5–8]. However, the antioxidative activities of low molecular weight glucans have yet to be fully understood. The structure-activity relationship (SAR) of LMG when exerting antioxidative effects was of interest. Using intracellular ceramide as a marker to determine the hydrolytic activity of SMases evaluated the antioxidative activity of isolated LMG, and also indicated if LMGs exerts protective effects on cells by attenuating injury under oxidative stress. Inflammatory oxidants cause this injury by inducing rapid increases in ceramide levels due to the hydrolysis of SM in plasma membranes. The study findings showed that LMG is capable of protecting cells against H2O2-induced apoptosis. LMG inhibited the activity of SMases to decrease the levels of intracellular ceramides, which may provide a useful indicator of the antioxidative activity of β-(1→3)-D-glucan.
3.4. Protective Effects of LMG against H2O2-Induced Macrophage Death
Beta-(1→3)-D-glucan is the one of the major components of polysaccharide from G. lucidum, and has anti-inflammatory properties [5]. This study examined the antioxidant effects of LMG. MTT assay evaluated the cytotoxicity of LMG to RAW264.7 cells (Figure 5(a)), indicating that LMG is not meaningfully cytotoxic to RAW264.7 macrophages when LMG was added at 0–200 μg/mL for 24 h. Procedures also evaluated the protective ability of LMG against H2O2-induced injury. After 24 h H2O2 (20 μM) treatment, MTT assay revealed 40% cell viability, which increased to over 80% after cotreatment of cells with LMG (100 μg/mL; Figure 5(b)). Results showed that LMG exerts dose-dependent protective effects against H2O2-induced cell death in RAW264.7 cells. The antioxidative activities of the other acidic hydrolyzed fractions, HMG and LMG, showed similar potency, but OSG (wich contains monosaccharide) demonstrated reduced antioxidative activity. This finding suggests that the molecular weights of beta-1,3-glucan plays important roles in determining its biological activity.
Figure 5.
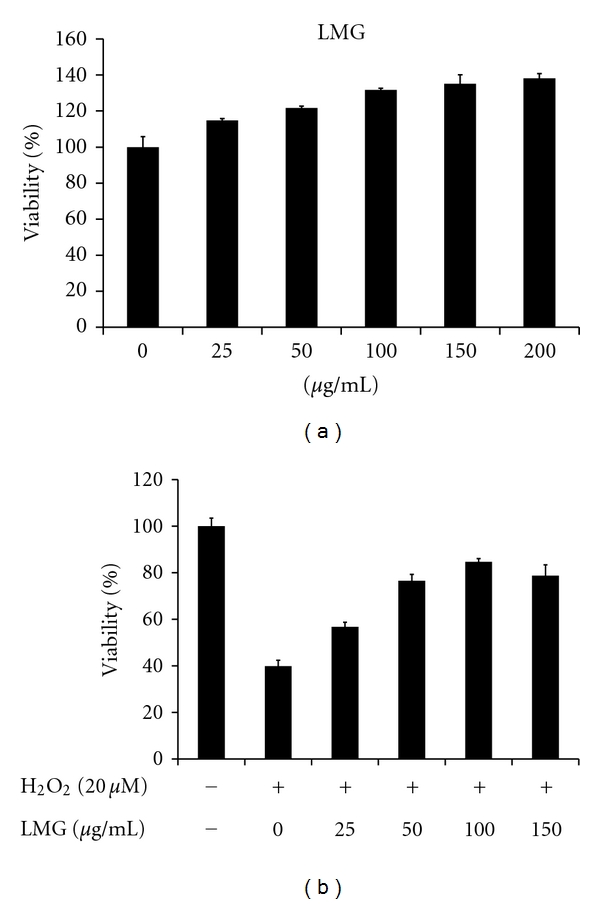
Protective abilities of LMG against H2O2-induced cytotoxicity in RAW264.7 cells. (a) MTT assay evaluated the cytotoxicity of LMG to RAW264.7 cells. Cells were treated with LMG (at 0, 25, 50, 100, 150, and 200 μg/mL) for 24 h. Results are presented as means ± SD for n = 5 replicates. (b) The protective ability of LMG from H2O2-induced cytotoxicity in RAW264.7 cells. Cells were cotreated with H2O2 (20 μM) and LMG (0, 25, 50, 100, and 150 μg/mL) for 24 h. Results are presented as the means ± SD for n = 5 replicates.
3.5. LMG Reduced H2O2-Induced Intracellular ROS Levels
H2O2 medication induces intracellular oxidative stress by increasing ROS production. This study estimated intracellular ROS in exogenous H2O2-injured (20 μM) macrophage cells. Fluorescence microscopy with DCFDA, a redox-sensitive fluorescent dye, visualized ROS production. In cells treated with H2O2 alone (Figure 6(b)), the oxidized DCF fluorescent intensity was significantly greater than in untreated control RAW264.7 cells (Figure 6(a)). Cotreatment with LMG (100 μg/mL) provided markedly reduced DCF fluorescence compared with fluorescence after addition of H2O2 only (Figure 6(c)). Increasing LMG decreased ROS production induced by addition of H2O2 to RAW264.7 cells.
Figure 6.
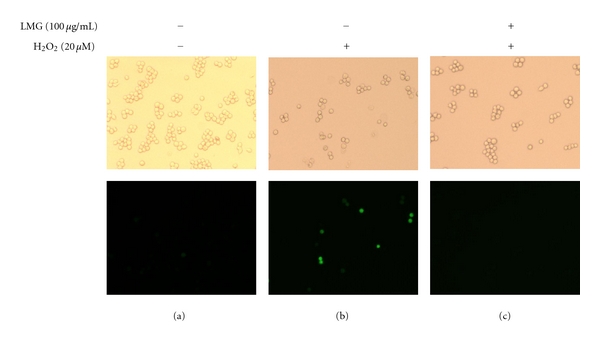
LMG reduced H2O2-induced intracellular ROS levels. RAW264.7 cells were treated with H2O2 (20 μM) and LMG (100 μg/mL) for 30 min, and intracellular ROS levels were measured using DCFDA fluorescent staining and fluorescence microscopy. Left panels, untreated control; middle panels, H2O2 added alone; right panels, co-treatment with H2O2 and LMG. Images in the upper panels were captured using bright-field microscopy (original magnification 200x), and those in the lower panels were captured using fluorescence microscopy.
3.6. LMG Inhibited SMase Activity
In previous studies, factors of inflammation, including TNF-α, ROS, LPS, amyloid-β, IL-1β, and LDL, stimulated SMase activation activity, which then produced ceramide and initiated ceramide signal pathways to transfer death signaling [25–27]. This study evaluated the effects of LMG on bacterial SMase activity in vitro. Increasing LMG (0–200 μg/mL) reduced the activity of nSMase, with an IC50 value of approximately 120 μg/mL (Figure 7(a)). LMG also had inhibitory effects on aSMase activity (Figure 7(b)). These data suggest that LMG has the potential to act as a SMase inhibitor. In Our previous research identified that a polysaccharide from Cordyceps mycelia possessed antioxidative ability and associated with SMase inhibitory effects, thus reducing levels of ceramides produced from hydrolysis of sphingomyelin [20]. Although findings did not fully elucidate the mechanism by which LMG exerts antioxidative effects, LMG's potential as a SMase inhibitor may be of importance for its future applications.
Figure 7.
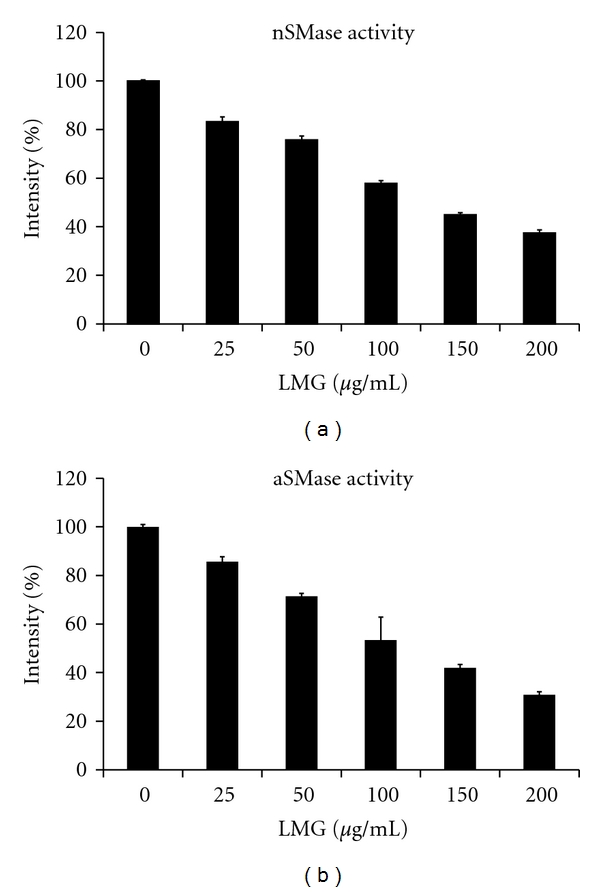
Amplex red assay evaluated LMG's SMase inhibitory activity. The relative activities of nSMase (a) and aSMase (b) were detected after treatment with 0, 25, 50, 100, and 200 μg/mL of LMG for 30 min. The relative activity is presented as the means ± SD for n = 3 replicates.
4. Conclusion
This study successfully isolated LMG, a low molecular weight β-1,3-glucan fraction, from the extracted residue of G. lucidum in high yields. A chemical method degraded the high-molecular-weight insoluble cell wall glucans into smaller sized compounds. This study also investigated the antioxidant activity of isolated LMG β-1,3-glucan to indicate the potential commercial value of G. lucidum glucans. LMG can protect cells by attenuating injury under oxidative stress caused by inflammatory oxidants. The study finding show that LMG, a water-soluble β-1,3-glucan with low molecular weight, recycled from extracted residue of G. lucidum, has activity against H2O2-induced apoptosis. LMG, therefore, is capable of exerting antioxidant effects by inhibiting SMases. The recycling of β-1,3-glucan from the remains of fruiting bodies could potentially provide a food supplement source of antioxidants for maintenance of body health. This is the first study to report the antioxidative effects of low molecular weight β-1,3-glucan from G. lucidum. The methods used have potential application in the isolation of antioxidative polysaccharides from other medicinal herbs.
Acknowledgments
This paper was supported by a Grant-in-Aid for scientific research from Genomics Research Center, Academia Sinica, Taiwan R.O.C., and a grant from Wan Fang Hospital, Taipei, Taiwan R.O.C. (100TMU-WFH-01-3). P. F. Kao and S. H. wang are contributed equally to this work.
References
- 1.Novak M, Vetvicka V. β-glucans, history, and the present: immunomodulatory aspects and mechanisms of action. Journal of Immunotoxicology. 2008;5(1):47–57. doi: 10.1080/15476910802019045. [DOI] [PubMed] [Google Scholar]
- 2.Laroche C, Michaud P. New developments and prospective applications for beta (1,3) glucans. Recent Patents on Biotechnology. 2007;1(1):59–73. doi: 10.2174/187220807779813938. [DOI] [PubMed] [Google Scholar]
- 3.Sliva D. Ganoderma lucidum in cancer research. Leukemia Research. 2006;30(7):767–768. doi: 10.1016/j.leukres.2005.12.015. [DOI] [PubMed] [Google Scholar]
- 4.Chan GC, Chan WK, Sze DM. The effects of beta-glucan on human immune and cancer cells. Journal of Hematology & Oncology. 2009;2(25):1–11. doi: 10.1186/1756-8722-2-25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Paterson RRM. Ganoderma—a therapeutic fungal biofactory. Phytochemistry. 2006;67(18):1985–2001. doi: 10.1016/j.phytochem.2006.07.004. [DOI] [PubMed] [Google Scholar]
- 6.Lin K-I, Kao Y-Y, Kuo H-K, et al. Reishi polysaccharides induce immunoglobulin production through the TLR4/TLR2-mediated induction of transcription factor Blimp-1. Journal of Biological Chemistry. 2006;281(34):24111–24123. doi: 10.1074/jbc.M601106200. [DOI] [PubMed] [Google Scholar]
- 7.Aydin S, Aytac E, Uzun H, et al. Effects of Ganoderma lucidum on obstructive jaundice-induced oxidative stress. Asian Journal of Surgery. 2010;33(4):173–180. doi: 10.1016/S1015-9584(11)60003-1. [DOI] [PubMed] [Google Scholar]
- 8.Li W-J, Nie S-P, Xie M-Y, Yu Q, Chen Y, He M. Ganoderma atrum polysaccharide attenuates oxidative stress induced by D-galactose in mouse brain. Life Sciences. 2011;88(15):713–718. doi: 10.1016/j.lfs.2011.02.010. [DOI] [PubMed] [Google Scholar]
- 9.Hung W-T, Wang S-H, Chen C-H, Yang W-B. Structure determination of β-glucans from Ganoderma lucidum with matrix-assisted laser desorption/ionization (MALDI) mass spectrometry. Molecules. 2008;13(8):1538–1550. doi: 10.3390/molecules13081538. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Miyazaki T, Nishijima M. Structural examination of an alkali-extracted, water-soluble heteroglycan of the fungus Ganoderma lucidum. Carbohydrate Research. 1982;109(1):290–294. [Google Scholar]
- 11.Bao X, Liu C, Fang J, Li X. Structural and immunological studies of a major polysaccharide from spores of Ganoderma lucidum (Fr.) Karst. Carbohydrate Research. 2001;332(1):67–74. doi: 10.1016/s0008-6215(01)00075-1. [DOI] [PubMed] [Google Scholar]
- 12.Bao X-F, Wang X-S, Dong Q, Fang J-N, Li X-Y. Structural features of immunologically active polysaccharides from Ganoderma lucidum. Phytochemistry. 2002;59(2):175–181. doi: 10.1016/s0031-9422(01)00450-2. [DOI] [PubMed] [Google Scholar]
- 13.Lee SE, Hwang HJ, Ha J-S, Jeong HS, Kim JH. Screening of medicinal plant extracts for antioxidant activity. Life Sciences. 2003;73(2):167–179. doi: 10.1016/s0024-3205(03)00259-5. [DOI] [PubMed] [Google Scholar]
- 14.Kolesnick RN, Krönke M. Regulation of ceramide production and apoptosis. Annual Review of Physiology. 1998;60:643–665. doi: 10.1146/annurev.physiol.60.1.643. [DOI] [PubMed] [Google Scholar]
- 15.Wang Y-Y, Khoo K-H, Chen S-T, Lin C-C, Wong C-H, Lin C-H. Studies on the immuno-modulating and antitumor activities of Ganoderma lucidum (Reishi) polysaccharides: functional and proteomic analyses of a fucose-containing glycoprotein fraction responsible for the activities. Bioorganic & Medicinal Chemistry. 2002;10(4):1057–1062. doi: 10.1016/s0968-0896(01)00377-7. [DOI] [PubMed] [Google Scholar]
- 16.Stellner K, Saito H, Hakomori S-I. Determination of aminosugar linkages in glycolipids by methylation: aminosugar linkages of ceramide pentasaccharides of rabbit erythrocytes and of Forssman antigen. Archives of Biochemistry and Biophysics. 1973;155(2):464–472. doi: 10.1016/0003-9861(73)90138-0. [DOI] [PubMed] [Google Scholar]
- 17.Suárez ER, Syvitski R, Kralovec JA, et al. Immunostimulatory polysaccharides from chlorella pyrenoidosa. A new galactofuranan. Measurement of molecular weight and molecular weight dispersion by DOSY NMR. Biomacromolecules. 2006;7(8):2368–2376. doi: 10.1021/bm060365x. [DOI] [PubMed] [Google Scholar]
- 18.Lin H-C, Tsai S-H, Chen C-S, et al. Structure-activity relationship of coumarin derivatives on xanthine oxidase-inhibiting and free radical-scavenging activities. Biochemical Pharmacology. 2008;75(6):1416–1425. doi: 10.1016/j.bcp.2007.11.023. [DOI] [PubMed] [Google Scholar]
- 19.Huang S-H, Lin C-M, Chiang B-H. Protective effects of Angelica sinensis extract on amyloid [beta]-peptide-induced neurotoxicity. Phytomedicine. 2008;15(9):710–721. doi: 10.1016/j.phymed.2008.02.022. [DOI] [PubMed] [Google Scholar]
- 20.Wang SH, Yang W-B, Liu Y-C, et al. A potent sphingomyelinase inhibitor from Cordyceps mycelia contributes to its cytoprotective effect against oxidative stress in macrophages. Journal of Lipid Research. 2011;52:471–479. doi: 10.1194/jlr.M011015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Kim Y-T, Kim E-H, Cheong C, Williams DL, Kim C-W, Lim S-T. Structural characterization of [beta]-D-(1 → 3, 1 → 6)-linked glucans using NMR spectroscopy. Carbohydrate Research. 2000;328(3):331–341. doi: 10.1016/s0008-6215(00)00105-1. [DOI] [PubMed] [Google Scholar]
- 22.Kollár R, Reinhold BB, Petráková E, et al. Architecture of the yeast cell wall. Journal of Biological Chemistry. 1997;272(28):17762–17775. doi: 10.1074/jbc.272.28.17762. [DOI] [PubMed] [Google Scholar]
- 23.Politi M, Groves P, Chávez MI, Cañada FJ, Jiménez-Barbero J. Useful applications of DOSY experiments for the study of mushroom polysaccharides. Carbohydrate Research. 2006;341(1):84–89. doi: 10.1016/j.carres.2005.11.008. [DOI] [PubMed] [Google Scholar]
- 24.Chang WC, Huang LCL, Wang Y-S, et al. Matrix-assisted laser desorption/ionization (MALDI) mechanism revisited. Analytica Chimica Acta. 2007;582(1):1–9. doi: 10.1016/j.aca.2006.08.062. [DOI] [PubMed] [Google Scholar]
- 25.Stancevic B, Kolesnick R. Ceramide-rich platforms in transmembrane signaling. FEBS Letters. 2010;584(9):1728–1740. doi: 10.1016/j.febslet.2010.02.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Jana A, Pahan K. Fibrillar amyloid-β-activated human astroglia kill primary human neurons via neutral sphingomyelinase: implications for Alzheimer’s disease. The Journal of Neuroscience. 2010;30(38):12676–12689. doi: 10.1523/JNEUROSCI.1243-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Jenkins RW, Canals D, Idkowiak-Baldys J, et al. Regulated secretion of acid sphingomyelinase. Journal of Biological Chemistry. 2010;285(46):35706–35718. doi: 10.1074/jbc.M110.125609. [DOI] [PMC free article] [PubMed] [Google Scholar]