

Abstract
The coordinate modulation of the cellular functions of cadherins and integrins plays an essential role in fundamental physiological and pathological processes, including morphogenesis, tissue differentiation and renewal, wound healing, immune surveillance, inflammatory response, tumor progression, and metastasis. However, the molecular mechanisms underlying the fine-tuned functional communication between cadherins and integrins are still elusive. This paper focuses on recent findings towards the involvement of reactive oxygen species (ROS) in the regulation of cell adhesion and signal transduction functions of integrins and cadherins, pointing to ROS as emerging strong candidates for modulating the molecular crosstalk between cell-matrix and cell-cell adhesion receptors.
1. Introduction
The communication between signaling pathways, the so-called molecular crosstalk, plays a central role in cell biology, enabling the cell to couple the molecular functions of either near neighbors or distant cell components, with resulting synergistic or antagonistic effects and eventually appropriate biological outcomes.
Among the most important cellular crosstalk events is the signaling network that couples the molecular functions of adhesion receptors of the integrin and cadherin families. Indeed, acting in concert with growth factor receptor signaling pathways, this regulatory network is fundamentally important during the entire life of all metazoans, whereas its dysfunction almost invariably leads to developmental defects and/or diseases, including genetic diseases and cancer [1].
Integrins and cadherins are the major cell-extracellular matrix (ECM) and cell-cell adhesion receptors, respectively, and represent critical determinants of tissue architecture and function both in developing and adult organisms [2, 3].
Integrins are heterodimeric transmembrane glycoproteins composed of noncovalently linked α and β subunits, which are endowed with both structural and regulatory functions. They link the ECM to several distinct cytoplasmic proteins and the actin cytoskeleton at focal adhesions, thus serving as organizing centers for the assembly of structural and regulatory protein complexes at discrete cell-matrix adhesion sites and providing a mechanically sensitive system for mechanotransduction [4]. Furthermore, often acting in concert with growth factor receptors, they provide both outside-in and inside-out transmission of signals across the plasma membrane that control a number of critical cellular processes, including adhesion, cytoskeleton remodeling, migration, proliferation, differentiation, apoptosis, and gene expression [2, 5]. Specifically, integrin-mediated outside-in signaling stimulates tyrosine phosphorylation and activation of several proteins, including major components of focal adhesions, such as Src and FAK nonreceptor tyrosine kinases (PTK), and paxillin, as well as receptor tyrosine kinases (RPTK). In turn, these proteins are antagonized by nonreceptor (PTP) and receptor tyrosine phosphatases (RPTP), and entwined in a dynamic interplay with small GTPases and components of specialized plasma membrane and endosome microdomains, including caveolin-1, to form compartmentalized signaling platforms that allow for temporal and spatial coordination of specific downstream signaling events [6, 7].
Cadherins are single-pass transmembrane glycoproteins that support calcium-dependent, homophilic cell-cell adhesion. Together with their cytoplasmic domain interactors, such as β-catenin and p120ctn, they constitute the core components of adherens junctions (AJs). These specialized adhesive structures link the cadherin homophilic adhesion to the actin cytoskeleton and are required for formation and maintenance of stable cell-cell adhesion and differentiated phenotype in all solid tissues [3, 8, 9]. Cadherin endocytosis and endosome-mediated trafficking has emerged as a major mechanism for controlling AJ remodeling [10–17]. Moreover, growing evidence demonstrates that cadherins can modulate the signaling activity of several proteins, including β-catenin, Ras and Rho family GTPases, PTK, RPTK, PTP, and RPTP, as well as mechanotransduction pathways that affect membrane and actin cytoskeleton dynamics [3, 18–21].
Although there is a large body of evidence supporting the existence of a fine-tuned crosstalk between members of these two adhesive receptor families, which influences their expression, turnover, positioning, and/or functions, and may enhance or suppress adhesion depending on the cellular and environmental context [1, 10, 17, 22–41], the molecules and molecular mechanisms involved in such important phenomenon are not completely defined. To clarify how this crosstalk is regulated remains therefore a fundamental challenge for basic and translational research, including research on tumor and vascular disease progression.
Multiple molecules and regulatory mechanisms have been placed at the heart of the molecular crosstalk between integrins and cadherins, including small GTPases of the Ras and the Rho families [10, 17, 42–45], nonreceptor kinases such as Src, FAK, Fer, and PI3K [27, 34, 46, 47], cell surface receptor-mediated pathways [48–50], and adhesion-dependent actomyosin traction forces [26, 34, 51].
Previously, we reported the pivotal role of the small GTPase Rap1 in regulating the crosstalk between cadherins and integrins, suggesting a model where Rap1 acts as a turnabout for endosome signaling and membrane trafficking pathways to orchestrate the delivery of cadherins and integrins to specific cell-cell and cell-matrix landmarks at the plasma membrane, respectively [10, 17].
Intriguingly, recent growing evidence suggests that reactive oxygen species (ROS) play an important role in both integrins, small GTPases, and cadherins functions, raising the possibility that ROS may contribute to the modulation of the molecular crosstalk between integrins and cadherins.
In this paper, we discuss the most recent advances on the role of ROS in outside-in and inside-out signal transduction events implicating integrins and cadherins, providing building bloks for the hypothesis that ROS constitute important players in the molecular crosstalk between these cell adhesion receptors.
2. ROS Metabolism and Signaling
ROS are a highly reactive group of oxygen-containing molecules, including free radicals and peroxides, such as superoxide anion (O2 ∙−) and hydrogen peroxide (H2O2), which are generated constitutively, as common by-products of oxidative metabolism, or in response to the activation of several oxidative enzyme complexes [52–55].
The superoxide anion (O2 ∙−) is the key determinant of the overall effects of ROS. Indeed, even though it has a short half-life, O2 ∙− is the precursor of all other major reactive oxygen species found in biological systems, including the powerful oxidants hydroxyl radical (∙OH), hydrogen peroxide (H2O2), and peroxynitrite (OONO−) [52–55]. It is generated by a number of sources located throughout the cell via the incomplete, one-electron reduction of molecular oxygen (O2). Specifically, under physiological conditions the redox complexes I (NADH/ubiquinone oxidoreductase) and III (ubiquinol/cytochrome c oxidoreductase) of the mitochondrial electron transport chain are the major constitutive source, converting up to 5% of molecular O2 to O2 ∙− [56]. In addition, O2 ∙− is produced by the activity of NAD(P)H oxidases, xanthine oxidases, cytochrome P450 monooxygenases, uncoupled NO synthase (NOS), myeloperoxidases, lipoxygenases, and cyclooxygenases [52–55], which can be induced by a variety of chemical and physical stimuli, including integrin ligands, growth factors, G-protein coupled receptor agonists, cytokines, neurotransmitters, metabolic factors, shear stress, ischemia/reperfusion, chemotherapeutics, and ionizing radiations, as well as aging [52–54, 57, 58]. Conversely, O2 ∙− is rapidly removed by distinct superoxide dismutase (SOD) isoenzymes, located in the mitochondria (SOD2), cytoplasm (SOD1), and extracellular (SOD3) compartments, which catalyze the dismutation of O2 ∙− into H2O2 and O2. In turn, H2O2 is reduced to H2O by the catalase and glutathione peroxidase enzymes. In addition, O2 ∙− can be converted to hydroxyl radical (∙OH) by the Fenton or Haber-Weiss reactions, or to peroxynitrite (OONO−) by reacting with nitric oxide (NO) [59] (Figure 1).
Figure 1.
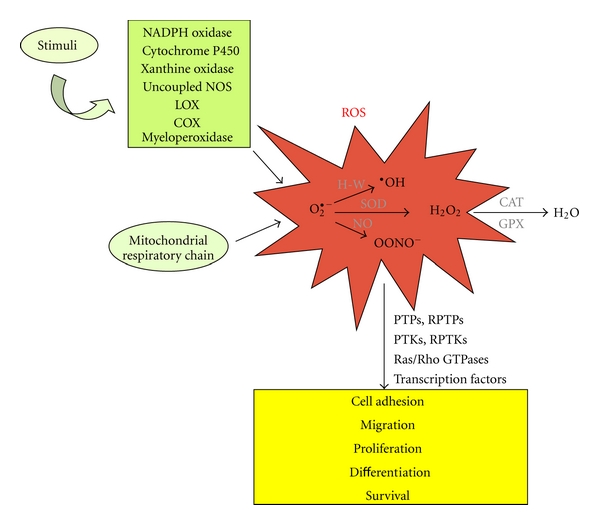
Schematic representation of ROS metabolism and signaling. The superoxide anion (O2 ∙−) is a key determinant of oxidative effects as well as the precursor of all other major reactive oxygen species, including hydroxyl radical (∙OH), hydrogen peroxide (H2O2), and peroxynitrite (OONO−). It is generated constitutively as by-product of oxidative metabolism, as well as upon stimuli triggering the activation of oxidative enzymes, including NADPH oxidases, xanthine oxidases, cytochrome P450 monooxygenases, uncoupled NO synthase (NOS), myeloperoxidases, lipoxygenases (LOX), and cyclooxygenases (COX). Conversely, O2 ∙− is removed by superoxide dismutase (SOD) enzymes, which catalyze the dismutation of O2 ∙− into H2O2 and O2. In turn, H2O2 is reduced to H2O by the catalase (CAT) and glutathione peroxidase (GPX) enzymes. At physiologic concentrations, ROS are endowed with essential signaling properties, being involved in the redox-dependent regulation of multiple signal transduction pathways to fulfill a wide range of essential biological processes, including cell adhesion, migration, proliferation, differentiation, and survival. However, at high levels, ROS exert very damaging effects through oxidative stress. H-W: Haber-Weiss reaction; NO: nitric oxide.
It is now well established that physiologic concentrations of ROS are endowed with essential signaling properties, which are mainly due to the reversible oxidation of redox-sensitive molecular targets, thereby functioning as signaling molecules. Accordingly, it has been clearly demonstrated that ROS are involved in the redox-dependent regulation of multiple signal transduction pathways to fulfill a wide range of essential biological processes, including cell adhesion, migration, proliferation, differentiation, and survival [52–55]. However, at high levels, ROS are known to exert very damaging effects through oxidative stress. This is caused by an imbalance between the production of ROS and the ability of cellular antioxidant mechanisms to readily detoxify the reactive intermediates. Importantly, because O2 ∙− can spontaneously react with NO to form OONO− at a rate 3 times faster than O2 ∙− dismutation by SOD, modest increases of O2 ∙− can result in a great reduction of NO bioavailability and increased formation of OONO−, a very strong oxidant with the potential to produce multiple cytotoxic effects [60, 61]. In addition, OONO− can also trigger feedforward mechanisms that further amplify O2 ∙− generation and oxidative stress, including the uncoupling of NO synthase (NOS) which produces O2 ∙− instead of NO, thus amplifying the risk of cellular dysfunction and oxidative injury [52]. The maintenance of highly regulated mechanisms to control ROS levels and functional specificity is therefore essential for normal cellular homeostasis and proper response to environmental stimuli.
Among the major source of ROS, NADPH oxidases have been demonstrated to play a fundamental role in the compartmentalization of ROS production and redox signaling [7].
The NADPH oxidase (NOX) complex was originally identified in phagocytic leukocytes as an enzymatic defense system against infections required for the oxidative burst-dependent microbial killing [62, 63]. It is composed of membrane-associated and cytosolic components, which assembly to form the active NOX enzymatic complex in response to appropriate stimuli. Specifically, this complex consists of membrane-associated cytochrome b558, comprising the catalytic gp91phox (also known as NOX2) and regulatory p22phox subunits, and four cytosolic regulatory components, including p40phox, p47phox, p67phox, and the small GTPase Rac1 [63]. Subsequently, NADPH oxidase complexes were also found in nonphagocytic cells, where several isoforms of the catalytic NOX2 protein were identified, including NOX1, NOX3, NOX4, and NOX5, and shown to localize in proximity of specific redox-sensitive molecular targets within discrete subcellular compartments, thereby facilitating the compartmentalization of redox signaling [7]. Indeed, NADPH oxidases can be targeted and activated within caveolae/lipid rafts, focal adhesions, cell-cell contacts, lamellipodial leading edges and membrane ruffles, endosomes, and the nucleus, allowing spatiotemporally confined ROS production and activation of specific redox signaling events [7].
Besides NADPH oxidase, an important role in the spatio-temporal regulation of ROS production is also played by enzymes involved in arachidonic acid (AA) metabolism, such as phospholipase A2(PLA2), lipooxygenases (LOX), and cyclooxygenases (COX), suggesting that a complex regulatory network may take place for proper modulation of redox signaling [64].
Accumulating evidence points to PTPs as the major redox-sensitive molecular targets of ROS [65]. This protein family is indeed characterized by the presence in the active site of a highly conserved sequence motif containing a Cys residue that is essential for catalysis and very susceptible to reversible inactivating oxidation by ROS. In turn, oxidative inactivation of PTPs promotes phosphorylation-dependent downstream signaling events. In addition to PTPs, other important signaling proteins have been shown to act as endogenous redox sensors for mediating ROS signaling, including RPTKs, cytoplasmic kinases, small GTPases of the Ras and Rho families, and transcription factors [7, 65, 66] (Figure 1). Conversely, protein oxidation can be reversed by thiol donors such as glutathione [67].
Remarkably, the activation of redox signaling complexes at integrin-mediated cell-matrix adhesion sites and cadherin-mediated cell-cell junctions induces opposite effects, leading to the assembly of focal adhesions and the disassembly of adherens junctions, respectively [7].
3. ROS and Integrins
It is now well established that ROS are implicated in regulating many integrin-mediated cellular responses, including adhesion, cytoskeleton organization, migration, proliferation, differentiation, and survival. Indeed, a large body of evidence demonstrates that integrin activation triggers a transient and localized burst of ROS, either independently or in cooperation with growth factor receptors, which is essential to the proper transduction of outside-in signaling pathways [7, 47, 68]. Specifically, although the underlying molecular mechanisms remain to be precisely defined, there is clear evidence that integrin engagement with antibodies or extracellular matrix proteins triggers ROS production by promoting changes in mitochondrial metabolic/redox function [69–71], and activation of distinct oxidases, including NADPH-oxidases [47, 72, 73], and the AA-metabolizing enzymes 5-lipoxygenase (5-LOX) [70, 72] and cyclooxygenase-2 (COX-2) [74]. Conversely, there is evidence that ROS can also influence integrin-mediated inside-out signaling by inducing the conformational change required for integrin activation [64]. Remarkably, the small GTPase Rac1 has emerged as a crucial, common upstream mediator of ROS production in integrin-mediated outside-in signaling [64, 69–72]. Consistently, Rac1 acts upstream of both NADPH oxidase [7] and AA-metabolizing enzymes, such as PLA2 [75, 76], 5-LOX [70, 72, 76], and COX-2 [77], whereas many reports show that AA metabolism modulates NADPH oxidase and mitochondrial ROS production, as well as the existence of a bidirectional signaling crosstalk between mitochondria, and NADPH oxidase, suggesting that Rac1 can orchestrate a complex web of regulation for ROS production [64, 78] (Figure 2). In addition, it is becoming evident that the formation of focal adhesions promotes the assembly of redox signaling platforms, involving integrins, growth factor receptors, and NADPH oxidases, which are essential for localized ROS production and activation of specific redox signaling pathways that mediate adhesion-dependent cell functions [7]. Furthermore, there is evidence that fine-tuned sequential compartmentalization and kinetics of ROS production can account for the modulation of distinct subsets of redox-sensitive signaling molecules involved in early and late phases of cell adhesion, leading to distinct outcomes [70, 79].
Figure 2.

ROS mediate integrin outside-in signaling. Integrin engagement with extracellular matrix (ECM) proteins triggers a transient and localized burst of ROS, either independently or in cooperation with growth factor receptors (GFR), which is essential to the proper transduction of outside-in signaling pathways. The small GTPase Rac1 acts as a crucial upstream regulator of ROS production, orchestrating integrin outside-in signaling-mediated changes in mitochondrial metabolic/redox function, and activation of distinct oxidases, including NADPH-oxidases (NOX), 5-lipoxygenase (5-LOX), and cyclooxygenase-2 (COX-2). The signaling properties of integrin-induced ROS are largely due to the reversible oxidation of specific subsets of redox-sensitive proteins, including oxidative inhibition of PTPs, and activation of PTKs, RPTKs, small GTPases of the Ras and Rho families, and transcription factors (TF) such as AP-1 and NF-κB.
The signaling properties of integrin-induced ROS are largely due to the reversible oxidation of target proteins, and especially of PTPs, as the activity of these proteins is dependent on reactive cysteine residues (Cys-SH) at their active site that are readily susceptible to reversible oxidation [7, 65]. Indeed, ROS produced locally by the synergistic action of integrins and growth factor receptors on NADPH oxidase, as well as on mitochondria and 5-LOX, have been shown to induce oxidative inactivation of distinct PTPs, including the low-molecular-weight protein tyrosine phosphatase (LMW-PTP), PTP1B, PTEN, and SHP2, preventing these enzymes from dephosphorylating and inactivating specific targets, and thereby promoting downstream adhesion-related signaling events (Figure 2). Consistently, integrin-mediated adhesive and signaling functions are significantly mimicked by PTP inhibition [80]. However, ROS generated by integrin activation can also activate PTKs and RPTKs through either direct oxidation of susceptible cysteine residues or indirect inhibition of negative regulatory PTPs [68], whereas the synergistic cooperation between integrins and growth factor receptors expands enormously the plethora of ROS-regulated target proteins to include redox-sensitive small GTPases of the Ras superfamily and transcription factors such as AP-1 and NF-κB [7, 81–83] (Figure 2).
Remarkably, ROS production has often a dual role in small GTPase regulation, leading to either inhibition or activation under different conditions [83–88]. In particular, the inactivation of RhoA has been shown to occur indirectly through a signaling cascade involving the Rac-stimulated release of O2 ∙− from NADPH oxidase, which in turn inhibits LMW-PTP. Because p190Rho-GTPase-activating protein (p190RhoGAP) is a substrate of LMW-PTP, inactivation of LMW-PTP results in accumulation of the active phosphorylated form of p190RhoGAP, which stimulates the hydrolysis of bound GTP to produce inactive GDP-bound RhoA, thereby determining well-characterized readouts, including decreased cell contractility and stabilization of cell-cell junctions [85, 88]. Conversely, RhoA can be directly activated by ROS-mediated reversible oxidation of two critical cysteine residues located in a unique redox-sensitive motif within the phosphoryl binding loop, leading to characteristic outcomes, including stress fiber and focal adhesion formation and cell-cell junction weakening [84, 89, 90]. On the other hand, there is evidence that ROS can activate Rap1 [91], whose signaling is in turn required for suppression of Ras-generated ROS and protection against oxidative stress and consequent cell dysfunctions [92–94].
4. ROS and Cadherins
Growing evidence demonstrates that ROS play a major role in either stabilization or destabilization of cell-cell junctions mediated by distinct cadherins, including E-, N-, and VE-cadherin [81, 95–99]. In particular, it has been reported that Rac1-mediated ROS production is an essential component in signaling cascades that promote p190RhoGAP translocation to the AJs and the consequent inhibition of local RhoA activity, thus favoring the stabilization of cell-cell contacts [99]. Conversely, clear evidence shows that Rac1-induced ROS function as signaling molecules to disrupt VE-cadherin-based cell-cell adhesion leading to various biological responses, including endothelial barrier dysfunction, enhanced microvascular permeability, and endothelial migration and proliferation involved in angiogenesis [7, 96, 98, 100, 101] (Figure 3). Intriguingly, the apparent contrast between the positive and negative roles of ROS in the maintenance of cadherin-mediated cell-cell junctions correlates with similar features of small GTPases involved in this process, including Rac1, RhoA, and Rap1. Indeed, depending on the extracellular and intracellular context, the activities of Rac1, RhoA, and Rap1 may be not only involved in regulating AJs and endothelial barrier maintenance, but also in active enforcement or disruption of AJs and endothelial barrier integrity, suggesting that the location and duration of the activities of these small GTPases affect the choice of downstream targets, thereby determining distinct biological outcomes [81]. Indeed, while under basal conditions Rac1 enforces the junctions that form the endothelial barrier by promoting ROS-mediated p190RhoGAP recruitment to AJs and the consequent inhibition of local RhoA activity, upon certain growth factor stimuli, including VEGF, it becomes part of a barrier-disturbing mechanism by inducing ROS-mediated phosphorylation of VE-cadherin at Tyr658 and Tyr731, and β-catenin at Tyr654, which lead to the disassembly of AJs [81, 96]. Whether ROS act directly on growth factor receptor kinase activity or, more likely, inhibit VE-cadherin-associated tyrosine phosphatases has still to be clarified. In addition, there is evidence for the involvement of the Pyk2 and Src redox-sensitive kinases in the phosphorylation of AJ proteins, including β-catenin and p120ctn, and the resulting loss of cell-cell adhesion mediated by the Rac1-ROS signaling pathway [95, 100, 102] (Figure 3). Notably, it has been reported that antioxidant compounds can inhibit VEGF-induced angiogenesis through disruption of ROS-dependent Src kinase activation and the subsequent VE-cadherin tyrosine phosphorylation, resulting in the retention of VE-cadherin at cell-cell contacts [100]. Conversely, the cell-cell contact-dependent inhibition of cell growth and stimulation of PTP activity [103] have been associated with a decrease in the steady-state levels of intracellular ROS and the consequent impairment of redox signaling mediated by growth factor receptors [104].
Figure 3.
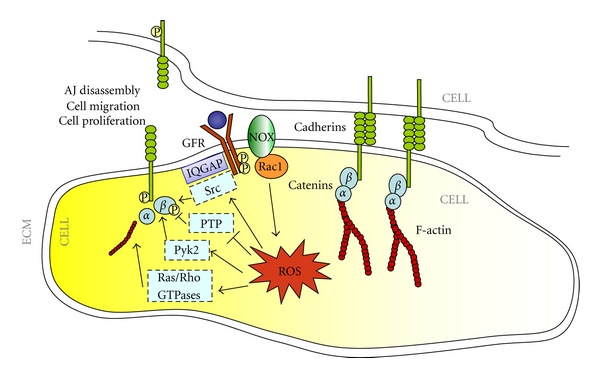
ROS modulate cadherin-mediated cell-cell junctions. Rac1-induced ROS may function as signaling molecules to disrupt cadherin-based cell-cell adhesion through either inhibition or activation of regulatory tyrosine phosphatases and kinases, respectively, as well as by localized activation of IQGAP and small GTPases, leading to various biological responses, including cell migration and proliferation.
Another component of the Rac1-ROS signaling pathway that plays an important role in the regulation of cadherin adhesive functions is IQGAP, a scaffold protein involved in cellular motility and morphogenesis [105]. IQGAP has been shown to be required for the establishment of VE-cadherin-based cell-cell contacts, and to colocalize and form a complex with VE-cadherin at cell-cell contact sites in quiescent endothelial cells [105]. It may act as a downstream effector of Rac1, as well as an inhibitor of its GTPase activity through a RasGAP-related domain [106–108]. Furthermore, it can facilitate localized ROS production through compartmentalization of Nox2 [109]. Indeed, there is evidence that IQGAP1 plays an essential role in VEGF-stimulated ROS production and VEGFR2-mediated endothelial cell migration and proliferation, suggesting that IQGAP1 may function as a scaffold protein to link VEGFR2 to the VE-cadherin/β-catenin complex at AJs, thereby promoting VEGF-stimulated ROS-dependent tyrosine phosphorylation of VE-cadherin, which may contribute to AJ weakening and angiogenesis [105, 110] (Figure 3).
Besides biochemical modification of AJ molecules, the ROS-dependent regulation of cadherins may be also driven by epigenetic events, as a ROS-induced hypermethylation of E-cadherin promoter, due to the upregulation of the transcriptional factor Snail and the recruitment of the DNA methyltransferase-1, and the consequent downregulation of cadherin expression have been reported [111].
5. ROS as Potential Pivotal Players in the Crosstalk between Integrins and Cadherins
A number of experimental reports have shown that the engagement of integrins with ECM proteins can affect cadherin-containing adherens junctions via multiple mechanisms, including the activation of signaling pathways mediated by small GTPases [10, 17, 42–45], cell surface receptors [48–50], and nonreceptor kinases [22, 27, 34, 47, 112], and the modulation of the actin network [26, 34, 51, 112]. Conversely, there are relatively fewer examples where cadherins have been shown to regulate integrin function [40, 113], but this may be due to the fact that crosstalk in this direction has been explored less extensively. In this context, we have previously reported that the small GTPase Rap1 plays a pivotal role in regulating the crosstalk between cadherins and integrins, suggesting a model where Rap1 acts as a turnabout for endosome signaling and membrane trafficking pathways to orchestrate the control of cadherin and integrin adhesive and signaling functions [10, 17].
Intriguingly, despite the lack of direct experimental evidence, the large number of studies implicating ROS as major modulators of integrin and cadherin adhesive and signaling functions strikingly supports the thought-provoking hypothesis that ROS play a crucial role in the crosstalk between integrins and cadherins (Figure 4). In particular, there is clear evidence that the assembly of integrin-mediated focal adhesions and the disassembly of cadherin-mediated adherens junctions require the activation of redox signaling complexes involving common regulatory proteins and mechanisms, including redox-sensitive small GTPases and the oxidative inactivation of PTPs [7]. Consistently, both focal adhesions assembly and adherens junctions disassembly are significantly mimicked by oxidative inhibitors of PTPs [10, 80], and prevented by ROS scavengers [95, 96].
Figure 4.
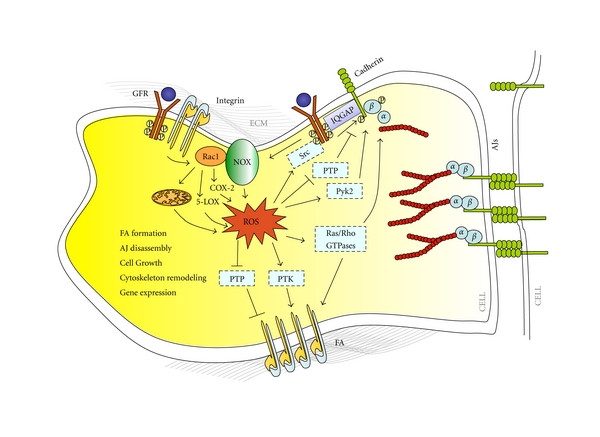
ROS in the crosstalk between integrins and cadherins. ROS generated by integrin activation may influence cadherin adhesive functions by various mechanisms, including inhibition of PTPs and/or activation of PTKs, RPTKs, and IQGAP acting at adherens junctions, as well as spatiotemporal modulation of the activity of redox-sensitive small GTPases and signaling endosomes.
Remarkably, both integrin- and cadherin-related redox signaling pathways involve Rac1 as a key mediator, which is in turn implicated in intimately intertwined functional relationships with other small GTPases, including Ras, RhoA, and Rap1 [7, 64, 69–72, 96, 98, 100, 101, 114].
Furthermore and importantly, recent evidence shows that Rap1 activation by Epac1, a Rap1-GEF involved in the Rap1-dependent regulation of cadherins, may be stimulated by ROS and inhibited by ROS scavengers, indicating that ROS production can trigger Rap1 activation [91]. Conversely, Rap1 signaling has been shown to be required for suppression of Ras-generated ROS and protection against oxidative stress and consequent cell dysfunctions [92–94]. Taking together, these data suggest that the role of Rap1 as pivotal regulator in the crosstalk between cadherins and integrins [10, 17] may underlie feedback mechanisms involving spatially and temporally regulated ROS production and scavenging. Consistently, KRIT1, a Rap1 effector whose loss-of-function mutations are implicated in endothelial cell-cell junction dysfunction and enhanced microvascular permeability underlying the Cerebral Cavernous Malformation disease, has been recently shown to play a role in molecular mechanisms involved in the maintenance of the intracellular ROS homeostasis to prevent oxidative cellular damage [115].
Finally, ROS generated by integrin activation could influence cadherin adhesive functions through the activation of either PTKs and RPTKs, including Src and growth factor receptors [68], or IQGAP, a component of the Rac1-ROS signaling pathway implicated in the modulation of AJ dynamics [105, 110] as well as in signaling downstream from both integrins and RPTKs [116], suggesting a further crosstalk mechanisms (Figure 4).
6. Concluding Remarks
It is well established that, besides their structural roles, both integrins and cadherins can provide bidirectional transmission of signals across topographically discrete regions of the plasma membrane. In addition, there is growing evidence supporting the existence of a fine-tuned, bidirectional crosstalk between these adhesion molecules, which may enhance or suppress their adhesive and signaling functions depending on the cellular and environmental context. Indeed, the integrin-cadherin crosstalk is involved in the epithelial-mesenchymal transition (EMT) underlying fundamental physiological and pathological processes, including embryonic development and cancer [22, 25–27, 33, 39].
This paper highlights recent growing evidence supporting a major role of reactive oxygen species (ROS) in both outside-in and inside-out signaling of integrins and cadherins, raising the possibility that ROS constitute master regulators of the crosstalk between these fundamental cell adhesion receptors.
Indeed, over the past few years, it has clearly emerged that outside-in integrin signaling triggers ROS production by several distinct mechanisms, such as changes in mitochondrial metabolic/redox function [69–71] and activation of distinct oxidases, including NADPH oxidase [47, 70, 72–74]. On the other hand, growing evidence demonstrates that ROS play a major role in the regulation of cadherin adhesive and signaling functions by mechanisms involving either biochemical modifications (e.g., phosphorylation) of AJ proteins, including cadherins and catenins, epigenetic modifications of the cadherin promoter, or modulation of small GTPases regulating cadherin-dependent cell-cell adhesion [7, 81, 95–101].
Remarkably, whereas emerging data show that integrin and cadherin redox signaling involves shared regulatory proteins, accumulating evidence suggests that discrete subcellular compartmentalization of ROS constitutes a major mechanism of localizing activation of downstream redox signaling events, thereby playing a critical role in transmitting cell signals in response to various environmental stimuli to regulate distinct cell functions, including cell-matrix and cell-cell adhesion [7, 117]. In particular, ROS production may be localized through interactions of NADPH oxidase with signaling platforms associated with lipid rafts and caveolae, as well as with endosomes [7, 118]. Furthermore, there is evidence that growth factor receptors mediate signaling through a subset of signaling endosomes termed redoxosomes (redox-active endosomes), which are uniquely equipped with redox-processing proteins capable of transmitting ROS signals from the endosome interior to redox-sensitive effectors on the endosomal surface, thereby controlling redox-dependent effector functions through the spatial and temporal regulation of ROS as second messengers [117].
Taken together with the well-established roles of growth factor receptors, small GTPases and endosome signaling in the functional relationship between integrins and cadherins [17], the experimental evidence and observation discussed in this paper point to a novel hypothetical mechanism whereby the spatial and temporal regulation of ROS may contribute significantly to the modulation of the molecular crosstalk between these cell adhesion receptors, thus opening a novel research avenue.
Furthermore, as the impairment of the integrin-cadherin crosstalk is involved in the development of serious pathological processes, including abnormal angiogenesis, tumor invasion, and metastasis, strategies aimed at controlling ROS homeostasis to preserve the coordinated adhesive and signaling functions of integrins and cadherins might harbor important therapeutic potential for human health.
Acknowledgments
The authors are grateful to Giosuè Boscolo for helping in figure drawing and to Salvatore Bozzaro, Stefano Braggion, Valentina Cutano, Chiara Martino, Elisa Ciglieri, Alessandro Morina, and Santina Barbaro for critical reading of the paper and helpful discussion. This work was supported by grants to S. F. Retta from the Fondazione Telethon (grant N° GGP06222) and the Ministero dell'Istruzione, dell'Università e della Ricerca (MIUR - PRIN 2008).
References
- 1.Thiery JP, Acloque H, Huang RYJ, Nieto MA. Epithelial-mesenchymal transitions in development and disease. Cell. 2009;139(5):871–890. doi: 10.1016/j.cell.2009.11.007. [DOI] [PubMed] [Google Scholar]
- 2.Hynes RO. Integrins: bidirectional, allosteric signaling machines. Cell. 2002;110(6):673–687. doi: 10.1016/s0092-8674(02)00971-6. [DOI] [PubMed] [Google Scholar]
- 3.Wheelock MJ, Johnson KR. Cadherins as modulators of cellular phenotype. Annual Review of Cell and Developmental Biology. 2003;19:207–235. doi: 10.1146/annurev.cellbio.19.011102.111135. [DOI] [PubMed] [Google Scholar]
- 4.Schwartz MA, DeSimone DW. Cell adhesion receptors in mechanotransduction. Current Opinion in Cell Biology. 2008;20(5):551–556. doi: 10.1016/j.ceb.2008.05.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Yamada KM, Even-Ram S. Integrin regulation of growth factor receptors. Nature Cell Biology. 2002;4(4):E75–E76. doi: 10.1038/ncb0402-e75. [DOI] [PubMed] [Google Scholar]
- 6.del Pozo MA, Balasubramanian N, Alderson NB, et al. Phospho-caveolin-1 mediates integrin-regulated membrane domain internalization. Nature Cell Biology. 2005;7(9):901–908. doi: 10.1038/ncb1293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Ushio-Fukai M. Compartmentalization of redox signaling through NaDPH oxidase-derived rOS. Antioxidants and Redox Signaling. 2009;11(6):1289–1299. doi: 10.1089/ars.2008.2333. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Dejana E. Endothelial cell-cell junctions: happy together. Nature Reviews Molecular Cell Biology. 2004;5(4):261–270. doi: 10.1038/nrm1357. [DOI] [PubMed] [Google Scholar]
- 9.Peifer M, Yap AS. Traffic control: p120-catenin acts as a gatekeeper to control the fate of classical cadherins in mammalian cells. Journal of Cell Biology. 2003;163(3):437–440. doi: 10.1083/jcb.200310090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Balzac F, Avolio M, Degani S, et al. E-cadherin endocytosis regulates the activity of Rap1: a traffic light GTPase at the crossroads between cadherin and integrin function. Journal of Cell Science. 2005;118(20):4765–4783. doi: 10.1242/jcs.02584. [DOI] [PubMed] [Google Scholar]
- 11.Fujita Y, Krause G, Scheffner M, et al. Hakai, a c-Cbl-like protein, ubiquitinates and induces endocytosis of the E-cadherin complex. Nature Cell Biology. 2002;4(3):222–231. doi: 10.1038/ncb758. [DOI] [PubMed] [Google Scholar]
- 12.Harris TJC, Tepass U. Adherens junctions: from molecules to morphogenesis. Nature Reviews Molecular Cell Biology. 2010;11(7):502–514. doi: 10.1038/nrm2927. [DOI] [PubMed] [Google Scholar]
- 13.Le TL, Yap AS, Stow JL. Recycling of E-cadherin: a potential mechanism for regulating cadherin dynamics. Journal of Cell Biology. 1999;146(1):219–232. [PMC free article] [PubMed] [Google Scholar]
- 14.Palacios F, Price L, Schweitzer J, Collard JG, D’Souza-Schorey C. An essential role for ARF6-regulated membrane traffic in adherens junction turnover and epithelial cell migration. EMBO Journal. 2001;20(17):4973–4986. doi: 10.1093/emboj/20.17.4973. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Palovuori R, Sormunen R, Eskelinen S. Src-induced disintegration of adherens junctions of madin-darby canine kidney cells is dependent on endocytosis of cadherin and antagonized by Tiam-1. Laboratory Investigation. 2003;83(12):1901–1915. doi: 10.1097/01.lab.0000107009.75152.03. [DOI] [PubMed] [Google Scholar]
- 16.Pece S, Gutkind JS. E-cadherin and Hakai: signalling, remodeling or destruction? Nature Cell Biology. 2002;4(4):E72–E74. doi: 10.1038/ncb0402-e72. [DOI] [PubMed] [Google Scholar]
- 17.Retta SF, Balzac F, Avolio M. Rap1: a turnabout for the crosstalk between cadherins and integrins. European Journal of Cell Biology. 2006;85(3-4):283–293. doi: 10.1016/j.ejcb.2005.09.007. [DOI] [PubMed] [Google Scholar]
- 18.Perez TD, Tamada M, Sheetz MP, Nelson WJ. Immediate-early signaling induced by E-cadherin engagement and adhesion. Journal of Biological Chemistry. 2008;283(8):5014–5022. doi: 10.1074/jbc.M705209200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Smutny M, Yap AS. Neighborly relations: cadherins and mechanotransduction. Journal of Cell Biology. 2010;189(7):1075–1077. doi: 10.1083/jcb.201005151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Vestweber D, Broermann A, Schulte D. Control of endothelial barrier function by regulating vascular endothelial-cadherin. Current Opinion in Hematology. 2010;17(3):230–236. doi: 10.1097/MOH.0b013e328338664b. [DOI] [PubMed] [Google Scholar]
- 21.Yap AS, Kovacs EM. Direct cadherin-activated cell signaling: a view from the plasma membrane. Journal of Cell Biology. 2003;160(1):11–16. doi: 10.1083/jcb.200208156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Avizienyte E, Frame MC. Src and FAK signalling controls adhesion fate and the epithelial-to- mesenchymal transition. Current Opinion in Cell Biology. 2005;17(5):542–547. doi: 10.1016/j.ceb.2005.08.007. [DOI] [PubMed] [Google Scholar]
- 23.Avizienyte E, Wyke AW, Jones RJ, et al. Src-induced de-regulation of E-cadherin in colon cancer cells requires integrin signalling. Nature Cell Biology. 2002;4(8):632–638. doi: 10.1038/ncb829. [DOI] [PubMed] [Google Scholar]
- 24.Borghi N, Lowndes M, Maruthamuthu V, Gardel ML, Nelson WJ. Regulation of cell motile behavior by crosstalk between cadherin- and integrin-mediated adhesions. Proceedings of the National Academy of Sciences of the United States of America. 2010;107(30):13324–13329. doi: 10.1073/pnas.1002662107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Chen X, Gumbiner BM. Crosstalk between different adhesion molecules. Current Opinion in Cell Biology. 2006;18(5):572–578. doi: 10.1016/j.ceb.2006.07.002. [DOI] [PubMed] [Google Scholar]
- 26.De Rooij J, Kerstens A, Danuser G, Schwartz MA, Waterman-Storer CM. Integrin-dependent actomyosin contraction regulates epithelial cell scattering. Journal of Cell Biology. 2005;171(1):153–164. doi: 10.1083/jcb.200506152. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Genda T, Sakamoto M, Ichida T, Asakura H, Hirohashi S. Loss of cell-cell contact is induced by integrin-mediated cell-substratum adhesion in highly-motile and highly-metastatic hepatocellular carcinoma cells. Laboratory Investigation. 2000;80(3):387–394. doi: 10.1038/labinvest.3780043. [DOI] [PubMed] [Google Scholar]
- 28.Gimond C, Van Der Flier A, Van Delft S, et al. Induction of cell scattering by expression of β1 integrins in β1- deficient epithelial cells requires activation of members of the Rho family of GTPases and downregulation of cadherin and catenin function. Journal of Cell Biology. 1999;147(6):1325–1340. doi: 10.1083/jcb.147.6.1325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Hintermann E, Yang N, O’Sullivan D, Higgins JMG, Quaranta V. Integrin α6β4-erbB2 complex inhibits haptotaxis by up-regulating E-cadherin cell-cell junctions in keratinocytes. Journal of Biological Chemistry. 2005;280(9):8004–8015. doi: 10.1074/jbc.M406301200. [DOI] [PubMed] [Google Scholar]
- 30.Hodivala KJ, Watt FM. Evidence that cadherins play a role in the downregulation of integrin expression that occurs during keratinocyte terminal differentiation. Journal of Cell Biology. 1994;124(4):589–600. doi: 10.1083/jcb.124.4.589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Huttenlocher A, Lakonishok M, Kinder M, et al. Integrin and cadherin synergy regulates contact inhibition of migration and motile activity. Journal of Cell Biology. 1998;141(2):515–526. doi: 10.1083/jcb.141.2.515. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Lu Q, Paredes M, Zhang J, Kosik KS. Basal extracellular signal-regulated kinase activity modulates cell-cell and cell-matrix interactions. Molecular and Cellular Biology. 1998;18(6):3257–3265. doi: 10.1128/mcb.18.6.3257. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Marsden M, DeSimone DW. Integrin-ECM interactions regulate cadherin-dependent cell adhesion and are required for convergent extension in Xenopus. Current Biology. 2003;13(14):1182–1191. doi: 10.1016/s0960-9822(03)00433-0. [DOI] [PubMed] [Google Scholar]
- 34.Martinez-Rico C, Pincet F, Thiery JP, Dufour S. Integrins stimulate E-cadherin-mediated intercellular adhesion by regulating Src-kinase activation and actomyosin contractility. Journal of Cell Science. 2010;123(5):712–722. doi: 10.1242/jcs.047878. [DOI] [PubMed] [Google Scholar]
- 35.Monier-Gavelle F, Duband JL. Cross talk between adhesion molecules: control of N-cadherin activity by intracellular signals elicited by β1 and β3 integrins in migrating neural crest cells. Journal of Cell Biology. 1997;137(7):1663–1681. doi: 10.1083/jcb.137.7.1663. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Nelson CM, Pirone DM, Tan JL, Chen CS. Vascular endothelial-cadherin regulates cytoskeletal tension, cell spreading, and focal adhesions by stimulating RhoA. Molecular Biology of the Cell. 2004;15(6):2943–2953. doi: 10.1091/mbc.E03-10-0745. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Retta SF, Cassarà G, D’Amato M, et al. Cross talk between β1 and αV integrins: β1 affects β3 mRNA stability. Molecular Biology of the Cell. 2001;12(10):3126–3138. doi: 10.1091/mbc.12.10.3126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Schreider C, Peignon G, Thenet S, Chambaz J, Pinçon-Raymond M. Integrin-mediated functional polarization of Caco-2 cells through E-cadherin-actin complexes. Journal of Cell Science. 2002;115(3):543–552. doi: 10.1242/jcs.115.3.543. [DOI] [PubMed] [Google Scholar]
- 39.Thiery JP, Sleeman JP. Complex networks orchestrate epithelial-mesenchymal transitions. Nature Reviews Molecular Cell Biology. 2006;7(2):131–142. doi: 10.1038/nrm1835. [DOI] [PubMed] [Google Scholar]
- 40.Von Schlippe M, Marshall JF, Perry P, Stone M, Zhu AJ, Hart IR. Functional interaction between E-cadherin and αv-containing integrins in carcinoma cells. Journal of Cell Science. 2000;113(3):425–437. doi: 10.1242/jcs.113.3.425. [DOI] [PubMed] [Google Scholar]
- 41.Yano H, Mazaki Y, Kurokawa K, Hanks SK, Matsuda M, Sabe H. Roles played by a subset of integrin signaling molecules in cadherin-based cell-cell adhesion. Journal of Cell Biology. 2004;166(2):283–295. doi: 10.1083/jcb.200312013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Arthur WT, Noren NK, Burridge K. Regulation of Rho family GTPases by cell-cell and cell-matrix adhesion. Biological Research. 2002;35(2):239–246. doi: 10.4067/s0716-97602002000200016. [DOI] [PubMed] [Google Scholar]
- 43.Kümper S, Ridley AJ. P120ctn and P-cadherin but not E-cadherin regulate cell motility and invasion of DU145 prostate cancer cells. PLoS One. 2010;5(7) doi: 10.1371/journal.pone.0011801. Article ID e11801. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Lozano E, Betson M, Braga VMM. Tumor progression: small GTpases and loss of cell-cell adhesion. BioEssays. 2003;25(5):452–463. doi: 10.1002/bies.10262. [DOI] [PubMed] [Google Scholar]
- 45.Tsygankova OM, Ma C, Tang W, et al. Downregulation of Rap1GAP in human tumor cells alters cell/matrix and cell/cell adhesion. Molecular and Cellular Biology. 2010;30(13):3262–3274. doi: 10.1128/MCB.01345-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Arregui C, Pathre P, Lilien J, Balsamo J. The nonreceptor tyrosine kinase Fer mediates cross-talk between N-cadherin and β1-integrins. Journal of Cell Biology. 2000;149(6):1263–1273. doi: 10.1083/jcb.149.6.1263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Sangrar W, Gao Y, Scott M, Truesdell P, Greer PA. Fer-mediated cortactin phosphorylation is associated with efficient fibroblast migration and is dependent on reactive oxygen species generation during integrin-mediated cell adhesion. Molecular and Cellular Biology. 2007;27(17):6140–6152. doi: 10.1128/MCB.01744-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Borok Z. Role for α3 integrin in EMT and pulmonary fibrosis. Journal of Clinical Investigation. 2009;119(1):7–10. doi: 10.1172/JCI38084. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Kim Y, Kugler MC, Wei Y, et al. Integrin α3β1-dependent β-catenin phosphorylation links epithelial smad signaling to cell contacts. Journal of Cell Biology. 2009;184(2):309–322. doi: 10.1083/jcb.200806067. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Ogita H, Takai Y. Cross-talk among integrin, cadherin, and growth factor receptor: roles of nectin and nectin-like molecule. International Review of Cytology. 2008;265:1–54. doi: 10.1016/S0074-7696(07)65001-3. [DOI] [PubMed] [Google Scholar]
- 51.Tsai J, Kam L. Rigidity-dependent cross talk between integrin and cadherin signaling. Biophysical Journal. 2009;96(6):L39–L41. doi: 10.1016/j.bpj.2009.01.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Chrissobolis S, Faraci FM. The role of oxidative stress and NADPH oxidase in cerebrovascular disease. Trends in Molecular Medicine. 2008;14(11):495–502. doi: 10.1016/j.molmed.2008.09.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Fortuño A, San José G, Moreno MU, Díez J, Zalba G. Oxidative stress and vascular remodelling. Experimental Physiology. 2005;90(4):457–462. doi: 10.1113/expphysiol.2005.030098. [DOI] [PubMed] [Google Scholar]
- 54.Miller AA, Drummond GR, Sobey CG. Reactive oxygen species in the cerebral circulation: are they all bad? Antioxidants and Redox Signaling. 2006;8(7-8):1113–1120. doi: 10.1089/ars.2006.8.1113. [DOI] [PubMed] [Google Scholar]
- 55.Valko M, Leibfritz D, Moncol J, Cronin MTD, Mazur M, Telser J. Free radicals and antioxidants in normal physiological functions and human disease. International Journal of Biochemistry and Cell Biology. 2007;39(1):44–84. doi: 10.1016/j.biocel.2006.07.001. [DOI] [PubMed] [Google Scholar]
- 56.Turrens JF. Mitochondrial formation of reactive oxygen species. Journal of Physiology. 2003;552(2):335–344. doi: 10.1113/jphysiol.2003.049478. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Girouard H, Iadecola C. Neurovascular coupling in the normal brain and in hypertension, stroke, and Alzheimer disease. Journal of Applied Physiology. 2006;100(1):328–335. doi: 10.1152/japplphysiol.00966.2005. [DOI] [PubMed] [Google Scholar]
- 58.Iadecola C, Park L, Capone C. Threats to the mind: aging, amyloid, and hypertension. Stroke. 2009;40(3):S40–S44. doi: 10.1161/STROKEAHA.108.533638. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Faraci FM, Didion SP. Vascular protection: superoxide dismutase isoforms in the vessel wall. Arteriosclerosis, Thrombosis, and Vascular Biology. 2004;24(8):1367–1373. doi: 10.1161/01.ATV.0000133604.20182.cf. [DOI] [PubMed] [Google Scholar]
- 60.Girouard H, Park L, Anrather J, Zhou P, Iadecola C. Cerebrovascular nitrosative stress mediates neurovascular and endothelial dysfunction induced by angiotensin II. Arteriosclerosis, Thrombosis, and Vascular Biology. 2007;27(2):303–309. doi: 10.1161/01.ATV.0000253885.41509.25. [DOI] [PubMed] [Google Scholar]
- 61.Pacher P, Beckman JS, Liaudet L. Nitric oxide and peroxynitrite in health and disease. Physiological Reviews. 2007;87(1):315–424. doi: 10.1152/physrev.00029.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Babior BM. Oxygen dependent microbial killing by phagocytes. (Second of two parts) New England Journal of Medicine. 1978;298(13):721–725. doi: 10.1056/NEJM197803302981305. [DOI] [PubMed] [Google Scholar]
- 63.Babior BM. NADPH oxidase. Current Opinion in Immunology. 2004;16(1):42–47. doi: 10.1016/j.coi.2003.12.001. [DOI] [PubMed] [Google Scholar]
- 64.Gregg D, De Carvalho DD, Kovacic H. Integrins and coagulation: a role for ROS/Redox signaling? Antioxidants and Redox Signaling. 2004;6(4):757–764. doi: 10.1089/1523086041361604. [DOI] [PubMed] [Google Scholar]
- 65.Monteiro HP, Arai RJ, Travassos LR. Protein tyrosine phosphorylation and protein tyrosine nitration in redox signaling. Antioxidants and Redox Signaling. 2008;10(5):843–889. doi: 10.1089/ars.2007.1853. [DOI] [PubMed] [Google Scholar]
- 66.Liu H, Colavitti R, Rovira II, Finkel T. Redox-dependent transcriptional regulation. Circulation Research. 2005;97(10):967–974. doi: 10.1161/01.RES.0000188210.72062.10. [DOI] [PubMed] [Google Scholar]
- 67.Forman HJ, Fukuto JM, Torres M. Redox signaling: thiol chemistry defines which reactive oxygen and nitrogen species can act as second messengers. American Journal of Physiology. 2004;287(2):C246–C256. doi: 10.1152/ajpcell.00516.2003. [DOI] [PubMed] [Google Scholar]
- 68.Chiarugi P, Fiaschi T. Redox signalling in anchorage-dependent cell growth. Cellular Signalling. 2007;19(4):672–682. doi: 10.1016/j.cellsig.2006.11.009. [DOI] [PubMed] [Google Scholar]
- 69.Kheradmand F, Werner E, Tremble P, Symons M, Werb Z. Role of rac1 and oxygen radicals in collagenase-1 expression induced by cell shape change. Science. 1998;280(5365):898–902. doi: 10.1126/science.280.5365.898. [DOI] [PubMed] [Google Scholar]
- 70.Taddei ML, Parri M, Mello T, et al. Integrin-mediated cell adhesion and spreading engage different sources of reactive oxygen species. Antioxidants and Redox Signaling. 2007;9(4):469–481. doi: 10.1089/ars.2006.1392. [DOI] [PubMed] [Google Scholar]
- 71.Werner E, Werb Z. Integrins engage mitochondrial function for signal transduction by a mechanism dependent on Rho GTPases. Journal of Cell Biology. 2002;158(2):357–368. doi: 10.1083/jcb.200111028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Chiarugi P, Pani G, Giannoni E, et al. Reactive oxygen species as essential mediators of cell adhesion: the oxidative inhibition of a FAK tyrosine phosphatase is required for cell adhesion. Journal of Cell Biology. 2003;161(5):933–944. doi: 10.1083/jcb.200211118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Honoré S, Kovacic H, Pichard V, Briand C, Rognoni JB. α2β1-Integrin signaling by itself controls G1/S transition in a human adenocarcinoma cell line (Caco-2): implication of NADPH oxidase-dependent production of ROS. Experimental Cell Research. 2003;285(1):59–71. doi: 10.1016/s0014-4827(02)00038-1. [DOI] [PubMed] [Google Scholar]
- 74.Broom OJ, Massoumi R, Sjölander A. α2β1 integrin signalling enhances cyclooxygenase-2 expression in intestinal epithelial cells. Journal of Cellular Physiology. 2006;209(3):950–958. doi: 10.1002/jcp.20796. [DOI] [PubMed] [Google Scholar]
- 75.Peppelenbosch MP, Qiu RG, De Vries-Smits AMM, et al. Rac mediates growth factor-induced arachidonic acid release. Cell. 1995;81(6):849–856. doi: 10.1016/0092-8674(95)90005-5. [DOI] [PubMed] [Google Scholar]
- 76.Woo CH, Eom YW, Yoo MH, et al. Tumor necrosis factor-α generates reactive oxygen species via a cytosolic phospholipase A2-linked cascade. Journal of Biological Chemistry. 2000;275(41):32357–32362. doi: 10.1074/jbc.M005638200. [DOI] [PubMed] [Google Scholar]
- 77.Wu R, Coniglio SJ, Chan A, Symons MH, Steinberg BM. Up-regulation of Rac1 by epidermal growth factor mediates COX-2 expression in recurrent respiratory papillomas. Molecular Medicine. 2007;13(3-4):143–150. doi: 10.2119/2007-00005.Wu. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Seung BL, In HB, Yun SB, Um HD. Link between mitochondria and NADPH oxidase 1 isozyme for the sustained production of reactive oxygen species and cell death. Journal of Biological Chemistry. 2006;281(47):36228–36235. doi: 10.1074/jbc.M606702200. [DOI] [PubMed] [Google Scholar]
- 79.Chiarugi P. From anchorage dependent proliferation to survival: lessons from redox signalling. IUBMB Life. 2008;60(5):301–307. doi: 10.1002/iub.45. [DOI] [PubMed] [Google Scholar]
- 80.Retta SF, Barry ST, Critchley DR, Defilippi P, Silengo L, Tarone G. Focal adhesion and stress fiber formation is regulated by tyrosine phosphatase activity. Experimental Cell Research. 1996;229(2):307–317. doi: 10.1006/excr.1996.0376. [DOI] [PubMed] [Google Scholar]
- 81.Beckers CML, Van Hinsbergh VWM, Van Nieuw Amerongen GP. Driving Rho GTPase activity in endothelial cells regulates barrier integrity. Thrombosis and Haemostasis. 2010;103(1):40–55. doi: 10.1160/TH09-06-0403. [DOI] [PubMed] [Google Scholar]
- 82.Clempus RE, Griendling KK. Reactive oxygen species signaling in vascular smooth muscle cells. Cardiovascular Research. 2006;71(2):216–225. doi: 10.1016/j.cardiores.2006.02.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Heo J. Redox control of GTPases: from molecular mechanisms to functional significance in health and disease. Antioxidants and Redox Signaling. 2011;14(4):689–724. doi: 10.1089/ars.2009.2984. [DOI] [PubMed] [Google Scholar]
- 84.Aghajanian A, Wittchen ES, Campbell SL, Burridge K. Direct activation of RhoA by reactive oxygen species requires a redox-sensitive motif. PloS One. 2009;4(11, article e8045) doi: 10.1371/journal.pone.0008045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Caron E. Rac signalling: a radical view. Nature Cell Biology. 2003;5(3):185–187. doi: 10.1038/ncb0303-185. [DOI] [PubMed] [Google Scholar]
- 86.Chen K, Craige SE, Keaney JF., Jr. Downstream targets and intracellular compartmentalization in Nox signaling. Antioxidants and Redox Signaling. 2009;11(10):2467–2480. doi: 10.1089/ars.2009.2594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Heo J, Raines KW, Mocanu V, Campbell SL. Redox regulation of RhoA. Biochemistry. 2006;45(48):14481–14489. doi: 10.1021/bi0610101. [DOI] [PubMed] [Google Scholar]
- 88.Nimnual AS, Taylor LJ, Bar-Sagi D. Redox-dependent downregulation of Rho by Rac. Nature Cell Biology. 2003;5(3):236–241. doi: 10.1038/ncb938. [DOI] [PubMed] [Google Scholar]
- 89.Heo J, Campbell SL. Mechanism of redox-mediated guanine nucleotide exchange on redox-active Rho GTPases. Journal of Biological Chemistry. 2005;280(35):31003–31010. doi: 10.1074/jbc.M504768200. [DOI] [PubMed] [Google Scholar]
- 90.Jin L, Ying Z, Webb RC. Activation of Rho/Rho kinase signaling pathway by reactive oxygen species in rat aorta. American Journal of Physiology. 2004;287(4):H1495–H1500. doi: 10.1152/ajpheart.01006.2003. [DOI] [PubMed] [Google Scholar]
- 91.Moon E-Y, Lee J-H, Lee J-W, Song J-H, Pyo S. ROS/Epac1-mediated Rap1/NF-kappaB activation is required for the expression of BAFF in Raw264.7 murine macrophages. Cellular Signalling. 2011;23(9):1479–1488. doi: 10.1016/j.cellsig.2011.05.001. [DOI] [PubMed] [Google Scholar]
- 92.Han KH, Lim S, Ryu J, et al. CB1 and CB2 cannabinoid receptors differentially regulate the production of reactive oxygen species by macrophages. Cardiovascular Research. 2009;84(3):378–386. doi: 10.1093/cvr/cvp240. [DOI] [PubMed] [Google Scholar]
- 93.Remans PHJ, Gringhuis SI, Van Laar JM, et al. Rap1 signaling is required for suppression of Ras-generated reactive oxygen species and protection against oxidative stress in T lymphocytes. Journal of Immunology. 2004;173(2):920–931. doi: 10.4049/jimmunol.173.2.920. [DOI] [PubMed] [Google Scholar]
- 94.Remans PHJ, Wijbrandts CA, Sanders ME, et al. CTLA-4Ig suppresses reactive oxygen species by preventing synovial adherent cell-induced inactivation of Rap1, a Ras family GTPase mediator of oxidative stress in rheumatoid arthritis T cells. Arthritis and Rheumatism. 2006;54(10):3135–3143. doi: 10.1002/art.22139. [DOI] [PubMed] [Google Scholar]
- 95.Inumaru J, Nagano O, Takahashi E, et al. Molecular mechanisms regulating dissociation of cell-cell junction of epithelial cells by oxidative stress. Genes to Cells. 2009;14(6):703–716. doi: 10.1111/j.1365-2443.2009.01303.x. [DOI] [PubMed] [Google Scholar]
- 96.Monaghan-Benson E, Burridge K. The regulation of vascular endothelial growth factor-induced microvascular permeability requires Rac and reactive oxygen species. Journal of Biological Chemistry. 2009;284(38):25602–25611. doi: 10.1074/jbc.M109.009894. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Thews O, Lambert C, Kelleher DK, Biesalski HK, Vaupel P, Frank J. Impact of reactive oxygen species on the expression of adhesion molecules in vivo. Advances in experimental medicine and biology. 2009;645:95–100. doi: 10.1007/978-0-387-85998-9_15. [DOI] [PubMed] [Google Scholar]
- 98.van Wetering S, van Buul JD, Quik S, et al. Reactive oxygen species mediate Rac-induced loss of cell-cell adhesion in primary human endothelial cells. Journal of Cell Science. 2002;115(9):1837–1846. doi: 10.1242/jcs.115.9.1837. [DOI] [PubMed] [Google Scholar]
- 99.Wildenberg GA, Dohn MR, Carnahan RH, et al. p120-catenin and p190RhoGAP regulate cell-cell adhesion by coordinating antagonism between Rac and Rho. Cell. 2006;127(5):1027–1039. doi: 10.1016/j.cell.2006.09.046. [DOI] [PubMed] [Google Scholar]
- 100.Lin MT, Yen MI, Lin CY, Kuo ML. Inhibition of vascular endothelial growth factor-induced angiogenesis by resveratrol through interruption of Src-dependent vascular endothelial cadherin tyrosine phosphorylation. Molecular Pharmacology. 2003;64(5):1029–1036. doi: 10.1124/mol.64.5.1029. [DOI] [PubMed] [Google Scholar]
- 101.Nwariaku FE, Liu Z, Zhu X, et al. NADPH oxidase mediates vascular endothelial cadherin phosphorylation and endothelial dysfunction. Blood. 2004;104(10):3214–3220. doi: 10.1182/blood-2004-05-1868. [DOI] [PubMed] [Google Scholar]
- 102.Van Buul JD, Anthony EC, Fernandez-Borja M, Burridge K, Hordijk PL. Proline-rich tyrosine kinase 2 (Pyk2) mediates vascular endothelial-cadherin-based cell-cell adhesion by regulating β-catenin tyrosine phosphorylation. Journal of Biological Chemistry. 2005;280(22):21129–21136. doi: 10.1074/jbc.M500898200. [DOI] [PubMed] [Google Scholar]
- 103.Sörby M, Östman A. Protein-tyrosine phosphatase-mediated decrease of epidermal growth factor and platelet-derived growth factor receptor tyrosine phosphorylation in high cell density cultures. Journal of Biological Chemistry. 1996;271(18):10963–10966. doi: 10.1074/jbc.271.18.10963. [DOI] [PubMed] [Google Scholar]
- 104.Pani G, Colavitti R, Bedogni B, Anzevino R, Borrello S, Galeotti T. A redox signaling mechanism for density-dependent inhibition of cell growth. Journal of Biological Chemistry. 2000;275(49):38891–38899. doi: 10.1074/jbc.M007319200. [DOI] [PubMed] [Google Scholar]
- 105.Yamaoka-Tojo M, Tojo T, Kim HW, et al. IQGAP1 mediates VE-cadherin-based cell-cell contacts and VEGF signaling at adherence junctions linked to angiogenesis. Arteriosclerosis, Thrombosis, and Vascular Biology. 2006;26(9):1991–1997. doi: 10.1161/01.ATV.0000231524.14873.e7. [DOI] [PubMed] [Google Scholar]
- 106.Hart MJ, Callow MG, Souza B, Polakis P. IQGAP1, a calmodulin-binding protein with a rasGAP-related domain, is a potential effector for cdc42Hs. EMBO Journal. 1996;15(12):2997–3005. [PMC free article] [PubMed] [Google Scholar]
- 107.Kuroda S, Fukata M, Kobayashi K, et al. Identification of IQGAP as a putative target for the small GTPases, Cdc42 and Rac1. Journal of Biological Chemistry. 1996;271(38):23363–23367. doi: 10.1074/jbc.271.38.23363. [DOI] [PubMed] [Google Scholar]
- 108.Mataraza JM, Briggs MW, Li Z, Entwistle A, Ridley AJ, Sacks DB. IQGAP1 promotes cell motility and invasion. Journal of Biological Chemistry. 2003;278(42):41237–41245. doi: 10.1074/jbc.M304838200. [DOI] [PubMed] [Google Scholar]
- 109.Ikeda S, Yamaoka-Tojo M, Hilenski L, et al. IQGAP1 regulates reactive oxygen species-dependent endothelial cell migration through interacting with Nox2. Arteriosclerosis, Thrombosis, and Vascular Biology. 2005;25(11):2295–2300. doi: 10.1161/01.ATV.0000187472.55437.af. [DOI] [PubMed] [Google Scholar]
- 110.Yamaoka-Tojo M, Ushio-Fukai M, Hilenski L, et al. IQGAP1, a novel vascular endothelial growth factor receptor binding protein, is involved in reactive oxygen species-dependent endothelial migration and proliferation. Circulation Research. 2004;95(3):276–283. doi: 10.1161/01.RES.0000136522.58649.60. [DOI] [PubMed] [Google Scholar]
- 111.Lim SO, Gu JM, Kim MS, et al. Epigenetic changes induced by reactive oxygen species in hepatocellular carcinoma: methylation of the E-cadherin promoter. Gastroenterology. 2008;135(6):2128–2140.e8. doi: 10.1053/j.gastro.2008.07.027. [DOI] [PubMed] [Google Scholar]
- 112.Wang Y, Jin G, Miao H, Li JYS, Usami S, Chien S. Integrins regulate VE-cadherin and catenins: dependence of this regulation on Src, but not on Ras. Proceedings of the National Academy of Sciences of the United States of America. 2006;103(6):1774–1779. doi: 10.1073/pnas.0510774103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Li H, Leung TC, Hoffman S, Balsamo J, Lilien J. Coordinate regulation of cadherin and integrin function by the chondroitin sulfate proteoglycan neurocan. Journal of Cell Biology. 2000;149(6):1275–1288. doi: 10.1083/jcb.149.6.1275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Yeh LH, Park YJ, Hansalia RJ, et al. Shear-induced tyrosine phosphorylation in endothelial cells requires Rac1-dependent production of ROS. American Journal of Physiology. 1999;276(4):C838–C847. doi: 10.1152/ajpcell.1999.276.4.C838. [DOI] [PubMed] [Google Scholar]
- 115.Goitre L, Balzac F, Degani S, et al. KRIT1 regulates the homeostasis of intracellular reactive oxygen species. PLoS One. 2010;5(7) doi: 10.1371/journal.pone.0011786. Article ID e11786. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Brandt DT, Grosse R. Get to grips: steering local actin dynamics with IQGAPs. EMBO Reports. 2007;8(11):1019–1023. doi: 10.1038/sj.embor.7401089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Oakley FD, Abbott D, Li Q, Engelhardt JF. Signaling components of redox active endosomes: the redoxosomes. Antioxidants and Redox Signaling. 2009;11(6):1313–1333. doi: 10.1089/ars.2008.2363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Ushio-Fukai M. Localizing NADPH oxidase-derived ROS. Science’s STKE. 2006;2006(349):p. re8. doi: 10.1126/stke.3492006re8. [DOI] [PubMed] [Google Scholar]