

Abstract
Chapter summary
This paper presents a brief review of several lines of evidence suggesting that chemokine receptors on dendritic cells play an important role in breaking tolerance to self and in inducing autoimmunity. First, we have shown that an idiotypic self-antigen obtained from malignant murine lymphomas, when covalently linked to selected chemokines or defensins that interact with receptors on immature dendritic cells (iDCs), has the capacity to break tolerance to self and induce humoral or cell-mediated anti-tumor responses. Since unlinked antigens mixed with the same chemokines or defensins or antigens fused with a mutant ligand deficient in receptor-binding capacity were not immunogenic, we propose that delivery of an antigen coupled to a ligand for receptors on iDCs promotes the processing and subsequent presentation of the antigen, resulting in immunoadjuvant effects. In a second study, we observed that two of five aminoacyl tRNA synthetases (aaRSs) – which act as autoantigens to which some patients with myositis have autoantibodies – were chemotactic for activated monocytes, T cells, and iDCs. These aaRSs interacted with either CC chemokine receptor (CCR)5 or CCR3, as was shown by desensitization with chemokines and the response of cell lines transfected with the chemokine receptor. Presumably, these autoantigens therefore have the capacity to attract inflammatory cells, including iDCs, to infiltrate affected muscle cells. These observations suggest the hypothesis that antigens delivered to receptors on iDCs are potent immunogens capable of breaking self-tolerance to tumor antigens to induce autoimmune diseases.
Keywords: aminoacyl + RNA synthetase, autoimmune myositis, chemokine receptors, dendritic cells, idiotypic lymphoma antigen
Introduction: characteristics of dendritic cells
It is now well established that dendritic cells (DCs) are pivotal antigen-presenting cells with the unique capacity to initiate primary as well as secondary immune responses. 'Immature' dendritic cells (iDCs) can be generated in vitro from CD34+ hematopoietic stem cells or from CD14+ monocytes by incubation with granulocyte/monocyte-colony-stimulating factor plus tumor necrosis factor-α or IL-4 for 7 days. In the course of this differentiation process, the DC precursors lose the CD34 and CD14 markers and express a number of novel phenotypic markers and cell-surface receptors [1-4]. After an additional 2 days of CD40 ligation or incubation with proinflammatory agents such as bacterial lipopolysaccharide or cytokines such as tumor necrosis factor-α, the iDCs become mature DCs (mDCs), with consistent changes in their phenotypic markers as shown in Fig. 1.
Figure 1.

In vitro stages of differentiation and maturation of dendritic cells. The phenotypic and functional characteristics of immature and mature dendritic cells are compared. APC, antigen-presenting cell; DC, dendritic cell; GM-CSF, granulocyte/macrophage-colony-stimulating factor; HLA-DR, histocompatibility leukocyte antigen; LPS, lipopolysaccharide; MLR, mixed leukocyte reaction; TNF, tumor necrosis factor.
The differences in the functional capabilities of iDCs and mDCs are relevant to understanding their respective roles in adaptive immunity and autoimmunity [2]. The iDCs can phagocytose particulate and larger soluble antigens and can enzymatically process them to generate small peptide fragments that can be presented to T cells on cell-surface MHC (major histocompatibility complex) antigens. However, the expression of MHC class II and costimulatory proteins by iDCs is low and hence these cells have little or no capacity to present antigens until they differentiate into mDCs. The mDCs lose their capacity to phagocytose particles and process antigens, but are induced by proinflammatory stimulants to express high levels of MHC class II alloantigens on their surface, along with processed antigenic peptides and costimulatory surface markers such as CD40, CD80, and CD86 (see Fig. 1). Thus, by expressing costimulatory molecules, mDCs can induce a second signal in T cells that, together with the first signal generated by TCR in response to the MHC-peptide complex, results in an immune rather than a tolerogenic/anergic response.
The changes in receptor expression from DC precursors to iDCs and to mDCs enables DCs as they mature to respond to a new set of ligands with consequent changes in the trafficking pattern of DCs [3]. For example, as precursors of DC become iDCs they become responsive to a wide range of proinflammatory stimuli and migrate towards sites of infections and injuries [2-5]. The iDCs express many receptors for proinflammatory cytokines as well as chemoattractant factors which equip iDCs with the capacity to respond to many exogenous and endogenous danger signals [4]. Chemoattractant receptors enable iDCs to migrate towards inflammatory sites where chemoattractants are being produced. Once they are there, they phagocytize and process self-antigens from cellular debris and foreign antigens from invading microorganisms. The interaction of chemoattractants with their receptors initiates a signal-transduction cascade that, in addition to activating cell migration, activates a number of genes, resulting in the production and secretion of a variety of mediators and effector molecules including cytokines such as IL-1, IL-2, IL-4, IL-12, IL-15, and TGF-β (transforming growth factor-β) chemokines ([6,7] and unpublished observations).
Expression of chemokine receptors by dendritic cells
The presence of 'danger' signals such as inflammatory mediators and components of infectious organisms at the peripheral inflammatory site serve to induce the maturation of iDCs [4]. As DCs mature, they lose many of their receptors, but they gain CCR7 and CXCR5, which enable them to migrate along concentration gradients of constitutively produced chemokines such as SLC/CCL21, ELC/CCL19, and BLC/CXCL13 to the site of origin of these chemokines in lymphoid tissues [5] (Fig. 2). It is in the well-organized lymphoid tissues that mDCs expressing a particular peptide–MHC complex have a better chance of encountering those few T lymphocytes expressing a TCR that can recognize the antigen on the DC, thus initiating an immune response. By directing the trafficking and homing of iDCs and mDCs, respectively, chemokine receptors therefore presumably play a crucial role in the uptake and delivery of antigenic signals and initiation of adaptive immune responses.
Figure 2.
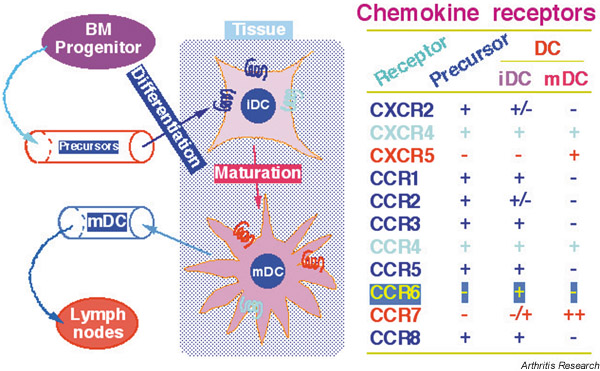
Changes in chemokine receptor expression in the course of maturation of dendritic cells. The in vivo traffic route of dendritic cells as they mature is depicted, along with the changes in the expression of chemokine receptors. BM, bone marrow; CC indicates the presence of adjoining cysteines in sequence; DC, dendritic cell; iDC, immature dendritic cell; mDC, mature dendritic cell.
Role of defensins in adaptive immunity
A number of disparate nonchemokine ligands have been shown to have the ability to activate chemokine receptors on iDCs and as a consequence to have major effects on adaptive immune responses. One example is the defensins, which interact with chemokine receptors. We have shown that β defensins interact with cells expressing CCR6, the receptor for a chemokine known as LARC/MIP-3α/CCL20, present on iDCs, resting memory CD4 cells, and a subset of B lymphocytes [8]. Defensins at micromolar concentrations have direct microbicidal activities, but at 1/10 to 1/100 micromolar concentrations, the β defensins derived from keratinocytes and mucosal epithelial cella activate and induce chemotactic migration of CCR6+ cells. Although the receptor for α defensins has not been identified, it is inhibitable by pertussis toxin and therefore also is a Gαi-protein-coupled receptor, like the chemokine receptors [9]. The possibility that α defensins interact with chemokine-like receptors on iDCs may account for our previous findings showing that α defensins are remarkably potent adjuvants that enhance both T-helper (Th)1 cell-mediated responses and Th2-type humoral antibody responses in mice to concomitantly administered antigens such as keyhole limpet hemocyanin (KLH) and ovalbumin [10,11].
Role of receptors on dendritic cells in breaking tolerance to 'self-tumor' antigens
The observed adjuvant effects of defensins on soluble antigens led us to examine the ability of β defensins to augment a weaker anti-tumor response. The anti-tumor effect of β defensins and selected chemokines was tested by linking them to an idotypic immunoglobulin antigen (Id) or its unique Fv Fragment (sFv) present on malignant murine lymphomas [12]. These linked fusion constructs were administered either as a naked DNA vaccine intra-muscularly or as recombinant fusion proteins. The results revealed that Id or sFv by themselves were not immunogenic [12,13]. Furthermore, mixtures of chemokines or murine β defensins (mBD2 and mBD3) and Id had no significant protective effect. Only when the β defensins or chemokines were covalently linked to the tumor antigen was an immunogenic anti-tumor effect achieved [12,13]. The Id–mBD2 fusion product yielded more cellular than humoral immunity, while Id–mBD3 induced predominantly antibody responses to the tumor antigens. Consequently, mBD2, but not mBD3, had a protective effect and induced prolonged tumor survival in 20–50% of mice challenged with tumors. The humoral immune effects of mBD3 fusion products resembled that of MDC/CCL22, which interacts with CCR4 and is known to favor the induction of TH2 responses. Alternatively, mBD2 mimicked the cell-mediated anti-tumor effects of MIP3α /CCL20, which also interacts with CCR6 and MCP3/CCL7, a chemokine that is known to interact with chemokine receptors CCR1, CCR2, and CCR3, which favors Th1-cell-mediated immune responses [14]. These findings suggest that the identity of the chemokine or defensin–receptor interaction on iDCs plays a role in determining whether humoral or cellular immunity is induced [13]. Because only Id antigens fused to chemokines or defensins were effective in inducing an immune response, and because these fused proteins were still functionally active inducers of in vitro chemotaxis of iDCs [13], presumably interaction of the fusion product with the chemokine receptor favors antigen uptake and processing and subsequent presentation by antigen-presenting cells. This hypothesis is supported by the inability of control fusion constructs (using mutated or an inactive β prodefensin moiety) that do not bind or activate the receptors on iDCs to elicit any immune response [13]. Thus, linking of sFv antigen to a functional chemokine or defensin, like a postage stamp, may help deliver the antigen more effectively to the iDCs in part by directing the migration of iDCs to the site of antigen production when used as a DNA vaccine. Since Id antigen actually represents an overexpressed normal clonal B-cell immunoglobulin product, these experiments actually break tolerance and produce an immune response to a self-antigen: in other words, they induce an 'autoimmune' response to the tumor.
Role of receptors on dendritic cells in autoimmune myositis
There have been several recent reports of a second type of nonchemokine ligand, namely tyrosyl tRNA synthetase (TyrRS) being chemotactic for neutrophils by selectively interacting with one of the receptors for IL-8, CXCR-1 [15,16]. This surprising result motivated us to evaluate the chemoattractant activities of other aminoacyl tRNA synthetases (aaRSs) to which some patients with idiopathic inflammatory myopathic (dermatomyositis and polymyositis or related diseases) develop autoantibodies [17].
Of the five aaRSs tested, two to which patients had autoantibodies were shown to be chemotactic for T lymphocytes, IL-2-activated monocytes, and immature dendritic cells (iDCs). They were not chemotactic for neutrophils, unstimulated monocytes, or mature dendritic cells (mDCs) (Table 1).
Table 1.
Chemotactic activity of aminoacyl tRNA synthetases
| Leukocyte subpopulation | HisRS | MHRS | 1–48 HisRS | AsnRS | SerRS | AspRS | LysRS |
|---|---|---|---|---|---|---|---|
| Neutrophils | - | - | - | - | - | - | - |
| Monocytes | - | - | - | - | - | - | - |
| IL-2-activated monocytes | + | - | + | N.D. | N.D. | N.D. | N.D. |
| Lymphocytes | + | - | + | + | + | ± | ± |
| CD8 T cells | ± | - | ± | N.D. | N.D. | N.D. | N.D. |
| CD4 T cells | + | - | + | + | N.D. | N.D. | N.D. |
| iDCs | + | - | + | + | - | + | - |
| mDCs | - | - | - | - | N.D. | N.D. | N.D. |
-, absent response; +, positive response; ±, variable response; N.D., not done. AsnRS, asparaginyl tRNA synthetase; AspRS, aspartyl tRNA synthetase; HisRS, histidyl tRNA synthetase; iDC, immature dendritic cells; LysRS, lysyl tRNA synthetase; mDCs, mature dendritic cells; MHRS, mutated HisRS; SerRS, seryl tRNA synthetase.
We tested histidyl tRNA synthetase (HisRS), the most frequently targeted aaRS in myositis, since 20–30% of patients have autoantibodies to it. We also tested the N-terminal coiled-coil domain of HisRS 1–48 with which autoantibodies react and found it to be a less potent chemotactic stimulant of IL-2-activated monocytes and T cells but not of iDCs. In contrast, a construct lacking the N-terminal domain (HisRS 61–509) was not chemotactic for any leukocyte subset. Furthermore, a mutated variant of HisRS, MHRS, also lacked any chemotactic activity.
Next, we investigated the possibility that HisRS utilized a G-coupled protein receptor by testing its susceptibility to pertussis toxin. Indeed, the chemotactic response of HisRS was inhibited by this toxin. Furthermore, we examined the desensitizing effects of a panel of chemokines to determine whether HisRS acted on a chemokine receptor. Preincubation with either RANTES/CCL5 or MIP-1β / CCL4, which uses CCR5, inhibited the subsequent chemotactic response of T cells to HisRS. Conversely, preincubation with HisRS selectively blocked the chemotactic response to these chemokines. Furthermore, HisRS was an equipotent chemoattractant for HEK293 cells transfected with CCR5, but not for cells transfected with CCR1 or CCR3. These observations thus identified CCR5 as a receptor for HisRS.
Although the incidence of autoantibody formation to asparaginyl tRNA synthetase (AsnRS) is less than 5%, this synthetase was also chemotactic for T cells and iDCs. We therefore tested AsnRS on a panel of cell lines transfected with chemokine receptors to identify its receptors and established that AsnRS was chemotactic for HEK293 cells transfected with CCR3. In contrast, three additional aaRSs to which no autoantibodies have been detected, namely seryl tRNA synthetase (SerRS), lysyl tRNA synthetase (LysRS), and aspartyl tRNA synthetase (AspRS) were not chemotactic for iDCs. Even though SerRS was chemotactic for lymphocytes expressing CCR3, it had no effect on iDCs. Consequently, the presence of autoantibodies to an aaRS appear to correlate with the capacity of the synthetase to have chemotactic effects on iDCs expressing receptors, suggesting that these antigens may contribute to myositis not only by inducing autoantibodies, but also by attracting inflammatory cells into affected muscle tissue. Indeed, three lines of evidence support this hypothesis. One is that infiltrates of mononuclear cells, including DCs, have been detected in striated muscle of patients with myositis [18]. Another is that CCR5 as well as CCR2 have been detected on cells infiltrating inflamed muscle tissue, and the degenerating and regenerating muscles themselves express low levels of CCR5 in myositis [19-21]. And finally, injection of naked DNA coding for HisRS into mice induces local myositis at the injection site [22].
Role of receptors on dendritic cells in pathogenesis of autoimmune disease
Another group of nonchemokine ligands that are well known to interact with CCR5 and CXCR4 are the envelope proteins gp120 and gp41 of HIV-1 (as reviewed [23]). They initially interact with CD4, and the result is a conformational change that enables the envelope proteins to interact with chemokine receptors as a prerequisite for the entry of HIV-1 into cells. This interaction with chemokine receptors is evident from the inability of monocytotropic HIV-1 to infect subjects homozygous for the Δ32 CCR5 variant [24,25]. This variant is retained in the cytoplasm and is not expressed on the cell surface. Furthermore, heterozygotes develop AIDS more slowly than homozygotes [26]. It is relevant to our findings that subjects heterozygous for Δ32 CCR5 also have a lower incidence of autoimmune diseases, including rheumatoid arthritis, Crohn's disease, and multiple sclerosis [27-30]. The mechanistic basis for the lower incidence of autoimmunity in such heterozygotes is elucidated to some extent by the phenotypic characteristics of CCR5 knockout mice [31]. Such mice have reduced resistance to several microbial pathogens and are protected from lipopolysaccharide-induced endotoxemia. The finding that cellular immune responses of CCR5-deficient mice are diminished suggests that ligation of CCR5 promotes Th1 polarization of immune responses [32]. This hypothesis is supported by observations that ligands, such as MIP-1α and RANTES, that utilize CCR5 favor induction of cytokine production by Th1 cells [32]. These reports suggest that CCR5 ligation may favor the development of Th1-type autoimmune conditions. Consequently, myositis may be a Th1 condition and this may account for the low incidence of autoantibody formation in these patients.
Our studies have shown that diverse chemoattractants for iDCs also have the capacity to break self-tolerance, resulting in the capacity to reject tumors or promulgate autoimmunity [33]. These findings suggest the hypothesis that antigens with chemotactic effects for receptors expressed by iDCs may be more effectively processed and presented to T cells, thus initiating and promulgating autoimmune responses or tumor immunity. Our observations suggest that the proinflammatory chemokine-like properties of some proteins provide a danger signal that not only amplifies innate inflammatory reactions, but also elicits adaptive immune responses, including the formation of autoantibodies.
Furthermore, the observation that the administration of an idiotypic tumor antigen derived from lymphomas, when linked to chemokines or β defensins as a fusion product, can apparently also break the tolerant or anergic state to that antigen further highlights the immuno-enhancing/adjuvant consequences of antigens interacting with receptors on iDCs. This idea is certainly supported by abundantly documented observations that antigens interacting with receptors on antigen-presenting cells are 10,000-fold more effectively processed by the MHC class II pathway than are pinocytosed antigens [34].
Therapeutic implications
Of course, many questions remain unanswered. For example, it is unclear why only a minority of patients with myositis develop autoantibodies selectively to aaRS. Perhaps myositis is predominantly based on Th1 cytokine responses, with a lower incidence of autoantibody production than is seen in Th2 autoimmune-based conditions. Since aaRSs are present in all nucleated cell types, the selective targeting of these particular autoantigens to muscle tissues in myositis is another puzzle awaiting solution. Nevertheless, our observations suggest that the autoantigenic aaRS has a causal role in this disease process and identifies several chemokine receptors, namely CCR5 and CCR3, as potential targets for therapeutic intervention in patients with autoimmune myositis.
Concluding remarks
Overall, our results lead to the prediction that ligands that interact with receptors on DC are likely to have immuno-adjuvant effects. This suggests that the relatively small proportion of self-antigens that are thought to be capable of inducing autoimmune conditions are more likely than most self-antigens to have domains that mimic ligands for receptors on iDCs. The observation that some aaRSs fall into that category is provocative, but additional autoantigens need to be examined to support this hypothesis.
Glossary of terms
aaRS = aminoacyl tRNA synthetase; AsnRS = asparaginyl tRNA synthetase; AspRS = aspartyl tRNA synthetase; CC indicates the presence of adjoining cysteines in sequence; CCL = CC chemokine ligand; CCR = CC chemokine receptor; DC = dendritic cell; HisRS = histidyl tRNA synthetase; Id = idiotypic immunoglobulin; iDC = immature dendritic cell; LARC = liver- and activation-regulated chemokine; LysRS = lysyl tRNA synthetase; mBD = murine β defensin; MCP = monocyte chemotactic protein; MDC = macrophage-derived chemokine; mDC = mature dendritic cell; MHRS = mutated HisRS; MIP = macrophage inflammatory protein; RANTES = regulated-upon-activation normal T cell expressed and secreted; SerRS = seryl tRNA synthetase; sFv = lymphoma-specific single-chain immunoglobulin (consisting of linked Vh and V2 domains of lymphoma immunoglobulin).
London, UK. 24-26 June 2002
Acknowledgements
We are grateful for the infinite patience and fortitude of Ms Cheryl Fogle, who retyped this draft on innumerable occasions, and to Dr Ruth Neta for her critical, constructive comments. The contents of this publication do not necessarily reflect the views or policies of the Department of Health and Human Services, nor does mention of trade names, commercial products, or organizations imply endorsement by the US Government. The publisher or recipient acknowledges the right of the US Government to retain a nonexclusive, royalty-free license in and to any copyright covering the article.
References
- Lanzavecchia A, Sallusto F. The instructive role of dendritic cells on T cell responses: lineages, plasticity and kinetics. Curr Opin Rheumatol. 2001;13:291–298. doi: 10.1097/00002281-200107000-00008. [DOI] [PubMed] [Google Scholar]
- Banchereau J, Steinman RM. Dendritic cells and the control of immunity. Nature. 1998;392:245–251. doi: 10.1038/32588. [DOI] [PubMed] [Google Scholar]
- Sallusto F, Schaerli P, Loetscher P, Schaniel C, Denig D, Mackay CR, Qin S, Lanzavecchia A. Rapid and coordinated switch in chemokine receptor expression during dendritic cell maturation. Eur J Immunol. 1998;28:2760–2769. doi: 10.1002/(SICI)1521-4141(199809)28:09<2760::AID-IMMU2760>3.3.CO;2-E. [DOI] [PubMed] [Google Scholar]
- Matzinger P. An innate sense of danger. Semin Immunol. 1998;10:399–415. doi: 10.1006/smim.1998.0143. [DOI] [PubMed] [Google Scholar]
- Sallusto F, Palermo B, Lenig D, Miettinen M, Matikainen S, Julkunen I, Forster R, Burgstahler R, Lipp M, Lanzavecchia A. Distinct patterns and kinetics of chemokine production regulate dendritic cell function. Eur J Immunol. 1999;29:1617–1625. doi: 10.1002/(SICI)1521-4141(199905)29:05<1617::AID-IMMU1617>3.0.CO;2-3. [DOI] [PubMed] [Google Scholar]
- Granucci F, Vizzardelli C, Pavelka N, Feau S, Persico M, Virzi E, Rescigno M, Moro G, Ricciardi-Castagnoli P. Inducible IL-2 production by dendritic cells revealed by global gene expression analysis. Nat Immunol. 2001;2:882–888. doi: 10.1038/ni0901-882. [DOI] [PubMed] [Google Scholar]
- Vissers JL, Hartgers FC, Lindhout E, Teunissen MB, Fidor CG, Adema G. Quantitative analysis of chemokine expression by dendritic cell subsets in vitro and in vivo. J Leukoc Biol. 2001;69:785–793. [PubMed] [Google Scholar]
- Yang D, Chertov O, Bykovskaia SN, Chen Q, Buffo MJ, Shogan J, Anderson M, Schroder JM, Wang JM, Howard OMZ, Oppenheim JJ. Beta-defensins: linking innate and adaptive immunity through dendritic and T cell CCR6. Science. 1999;286:525–528. doi: 10.1126/science.286.5439.525. [DOI] [PubMed] [Google Scholar]
- Yang D, Chen Q, Chertov O, Oppenheim JJ. Human neutrophil defensins selectively chemoattract naive T and immature dendritic cells. J Leukoc Biol. 2000;68:9–14. [PubMed] [Google Scholar]
- Tani K, Murphy WJ, Chertov O, Salcedo R, Koh CY, Utsunomiya I, Funakoshi S, Asai O, Hermann S, Wang J, Kwak L, Oppenheim JJ. Defensins act as potent adjuvants that promote cellular and humoral immune response in mice to a lymphoma idiotype and carrier antigens. J Int Immunol. 2000;12:691–700. doi: 10.1093/intimm/12.5.691. [DOI] [PubMed] [Google Scholar]
- Lillard JW Jr, Boyaka PN, Chertov O, Oppenheim JJ, McGhee JR. Mechanisms for induction of acquired host immunity by neutrophil peptide defensins. Proc Natl Acad Sci U S A. 1999;96:651–656. doi: 10.1073/pnas.96.2.651. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Biragyn A, Tani K, Grimm MC, Weeks SD, Kwak LW. Genetic fusion of chemokines to a self tumor antigen induces protective, T-cell dependent antitumor immunity. Nat Biotechnol. 1999;17:253–258. doi: 10.1038/6995. [DOI] [PubMed] [Google Scholar]
- Biragyn A, Surehnu M, Yang D, Ruffini PA, Haines BA, Klyush-nenkova E, Oppenheim JJ, Kwak LW. Mediators of innate immunity that target immature, but not mature, dendritic cells induce antitumor immunity when genetically fused with non-immunogenic tumor antigens. J Immunol. 2001;167:6644–6653. doi: 10.4049/jimmunol.167.11.6644. [DOI] [PubMed] [Google Scholar]
- Sato N, Ahuja SK, Quinones M, Kostecki V, Reddick RL, Melby PC, Kuziel WA, Ahuja SS. CC chemokine receptor (CCR)2 is required for langerhans cell migration and localization of T helper cell type 1 (Th1)-inducing dendritic cells. Absence of CCR2 shifts the Leishmania major-resistant phenotype to a susceptible state dominated by Th2 cytokines, b cell outgrowth, and sustained neutrophilic inflammation. J Exp Med. 2000;192:205–218. doi: 10.1084/jem.192.2.205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wakasugi K, Schimmel P. Two distinct cytokines released from a human aminoacyl-tRNA synthetase. Science. 1999;284:147–151. doi: 10.1016/S0304-3940(00)00990-3. [DOI] [PubMed] [Google Scholar]
- Wakasugi K, Schimmel P. Highly differentiated motifs responsible for two cytokine activities of a split human tRNA synthetase. J Biol Chem. 1999;274:23155–23159. doi: 10.1074/jbc.274.33.23155. [DOI] [PubMed] [Google Scholar]
- Targoff IN. Update on myositis-specific and myositis-associated autoantibodies. Curr Opin Rheumatol. 2000;12:475–481. doi: 10.1097/00002281-200011000-00001. [DOI] [PubMed] [Google Scholar]
- Nagaraju K, Raben N, Villalba ML, Danning C, Loeffler LA, Lee E, Tresser N, Abati A, Fetsch P, Plotz PH. Costimulatory markers in muscle of patients with idiopathic inflammatory myopathies and in cultured muscle cells. Clin Immunol. 1999;92:161–169. doi: 10.1006/clim.1999.4743. [DOI] [PubMed] [Google Scholar]
- Adams EM, Kirkley J, Eidelman G, Dohlman J, Plotz PH. The predominance of beta (CC) chemokine transcripts in idiopathic inflammatory muscle diseases. Proc Assoc Am Physicians. 1997;109:275–285. [PubMed] [Google Scholar]
- Confalonieri P, Bernasconi P, Megna P, Galbiati S, Cornelio F, Mantegazza R. Increased expression of beta-chemokines in muscle of patients with inflammatory myopathies. J Neuropathol Exp Neurol. 2000;59:164–169. doi: 10.1093/jnen/59.2.164. [DOI] [PubMed] [Google Scholar]
- De Rossi M, Bernasconi P, Baggi F, de Waal Malefyt R, Mantegazza R. Cytokines and chemokines are both expressed by human myoblasts: possible relevance for the immune pathogenesis of muscle inflammation. Int Immunol. 2000;12:1329–1335. doi: 10.1093/intimm/12.9.1329. [DOI] [PubMed] [Google Scholar]
- Blechynden LM, Lawson MA, Tabarias H, Garlepp MJ, Sherman J, Raben N, Lawson CM. Myositis induced by naked DNA immunization with the gene for histidyl-tRNA synthetase. Hum Gene Ther. 1997;8:1469–1480. doi: 10.1089/hum.1997.8.12-1469. [DOI] [PubMed] [Google Scholar]
- Wang JM, Oppenheim JJ. Interference with the signaling capacity of CC chemokine receptor 5 can compromise its role as an HIV-1 entry coreceptor in primary T lymphocytes. J Exp Med. 1999;190:591–595. doi: 10.1084/jem.190.5.591. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu R, Paxton WA, Choe S, Ceradini D, Martin SR, Horuk R, Mac-Donald ME, Stuhlmann H, Koup RA, Landau NR. Homozygous defect in HIV-1 coreceptor accounts for resistance of some multiply-exposed individuals to HIV-1 infection. Cell. 1996;86:367–377. doi: 10.1016/s0092-8674(00)80110-5. [DOI] [PubMed] [Google Scholar]
- Samson M, Libert F, Doranz BJ, Rucker J, Liesnard C, Farber CM, Saragosti S, Lapoumeroulie C, Cognaux J, Forceille C, Muyldermans G, Collman RG, Doms RW, Vassart G, Parmentier M. Resistance to HIV-1 infection in Caucasian individuals bearing mutant alleles of the CCR-5 chemokine receptor gene. Nature. 1996;382:772–725. doi: 10.1038/382722a0. [DOI] [PubMed] [Google Scholar]
- Dean M, Jacobson LP, McFarlane G, Margolick JB, Jenkins FJ, Howard OMZ, Dong HF, Goedert JJ, Buchbinder S, Gomperts E, Vlahov D, Oppenheim JJ, O'Brien SJ, Carrington M. Reduced risk of AIDS lymphoma in individuals heterozygous for the CCR5-delta32 mutation. Cancer Res. 1999;59:3561–3564. [PubMed] [Google Scholar]
- Zang YC, Samanta AK, Halder JB, Hong J, Tejada-Simon MV, Rivera VM, Zhang JZ. Aberrant T cell migration towards RANTES and MIP-1 alpha in patients with multiple sclerosis. Overexpression of chemokine receptor CCR5. Brain. 2000;123:1874–1882. doi: 10.1093/brain/123.9.1874. [DOI] [PubMed] [Google Scholar]
- Garred P, Madsen HO, Peterson J, Marquart H, Hansen TM, Freisleben Sorensen S, Volck B, Svejgaard A, Anderson V. CC chemokine receptor 5 polymorphism in rheumatoid arthritis. J Rheumatol. 1998;25:1462–1465. [PubMed] [Google Scholar]
- Gomez-Reino JJ, Pablos JL, Carreira PE, Santiago B, Serrano L, Vicario JL, Balsa A, Figueroa M, de Juan MD. Association of rheumatoid arthritis with a functional chemokine receptor, CCR5. Arthritis Rheum. 1999;42:989–992. doi: 10.1002/1529-0131(199905)42:5<989::AID-ANR18>3.0.CO;2-U. [DOI] [PubMed] [Google Scholar]
- Herfarth H, Pollok-Kopp B, Goke M, Press A, Oppermann M. Polymorphism of CC chemokine receptors CCR2 and CCR5 in Crohn's disease. Immunol Lett. 2001;77:113–117. doi: 10.1016/S0165-2478(01)00199-7. [DOI] [PubMed] [Google Scholar]
- Zhou Y, Kurihara T, Ryseck RP, Yang Y, Ryan C, Loy J, Warr G, Bravo R. Impaired macrophage function and enhanced T cell-dependent immune response in mice lacking CCR5, the mouse homologue of the major HIV-1 coreceptor. J Immunol. 1998;160:4018–4025. [PubMed] [Google Scholar]
- Andres PG, Beck PL, Mizoguchi E, Mizoguchi A, bhan AK, Dawson T, Kuziel WA, Maeda N, MacDermott RP, Podolsky DK, Reinecker HC. Mice with a selective deletion of the CC chemokine receptors 5 or 2 are protected from dextran sodium sulfate-mediated colitis: lack of CC chemokine receptor 5 expression results in a NK1.1+lymphocyte-associated Th2-type immune response in the intestine. J Immunol. 2000;164:6303–6312. doi: 10.4049/jimmunol.164.12.6303. [DOI] [PubMed] [Google Scholar]
- Bender A, Ernst N, Iglesias A, Dornmair K, Wekerle, Hohlfeld R. T cell receptor repertoire in polymyositis: clonal expansion of autoaggressive CD8+ T cells. J Exp Med. 1995;181:1863–1868. doi: 10.1084/jem.181.5.1863. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Watts C. Capture and processing of exogeneous antigens for presentation on MHC molecules. Annu Rev Immunol. 1997;15:821–850. doi: 10.1146/annurev.immunol.15.1.821. [DOI] [PubMed] [Google Scholar]